Aiphanes - Aiphanes
| Aiphanes | |
|---|---|
 | |
| Aiphanes horrida -de Jena Botanik Bahçesi | |
| bilimsel sınıflandırma | |
| Krallık: | Plantae |
| Clade: | Trakeofitler |
| Clade: | Kapalı tohumlular |
| Clade: | Monokotlar |
| Clade: | Kommelinidler |
| Sipariş: | Arecales |
| Aile: | Arecaceae |
| Alt aile: | Arecoideae |
| Kabile: | Cocoseae |
| Cins: | Aiphanes Willd. |
| Türler | |
| Aiphanes horrida | |
| Çeşitlilik | |
| Yaklaşık 26 tür | |
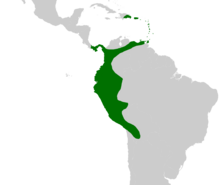 | |
| Yerel dağılımı Aiphanes | |
| Eş anlamlı[1] | |
Martinezia (sensu Kunth, değil Ruiz y Pavón ) | |
Aiphanes bir cins nın-nin dikenli palmiyeler hangisine özgü tropikal bölgeleri Güney ve Orta Amerika ve Karayipler.[2] Yaklaşık 26 tane var cins içindeki türler (aşağıya bakın), boyut olarak alt kat yeraltı ile çalılar kaynaklanıyor alt gölgelik 20 metre (66 ft) uzunluğundaki ağaçlar. Çoğu var sıkıca bileşik yapraklar (merkezi bir eksen boyunca çiftler halinde, tüy benzeri düzenlenmiş yaprakçıklara bölünmüş yapraklar); bir tür var tüm yapraklar. Sapları, yaprakları ve hatta bazen meyveleri dikenlerle kaplıdır. Bitkiler ömürleri boyunca defalarca çiçek açar ve ayrı ayrı erkek ve dişi çiçeklere sahiptir, ancak bunlar aynı şekilde birlikte taşınırlar. çiçeklenme. Tozlayıcıların kayıtları sınırlı olmasına rağmen, çoğu türün böcekler tarafından tozlaştığı görülmektedir. Meyveler, en az iki tür de dahil olmak üzere birçok kuş ve memeli tarafından yenir. amazon papağanları.
Carl Ludwig Willdenow adını icat etti Aiphanes 1801'de. Bundan önce, cinse ait türler yerleştirilmişti. Baktris veya Caryota. İsim Martinezia cinse de uygulanmıştır ve 1847 ile 1932 arasında genellikle Aiphanes. Max Burret adı diriltti Aiphanes 1932'de ve modern cins kavramının temelini attı. Aiphanes en çok diğer birkaç dikenli avuç içi cinsiyle yakından ilgilidir—Akrocomia, Astrocaryum, Baktris ve Desmoncus. İki tür yaygın olarak ekilir süs eşyaları ve meyve, tohumlar veya palmiye kalbi çeşitli türler tarafından yenildi Amerika'nın yerli halkları bin yıldır.
Açıklama
Aiphanes 20 metre (66 ft) yüksekliğindeki alt kanopi ağaçlarından ormanın alt katında büyüyen yer altı gövdeli küçük çalılara kadar değişen dikenli palmiyeler cinsidir.[3] Adı, Antik Yunan ai, "her zaman" anlamına gelir hayalet"açık", "görünür" veya "dikkat çekici" anlamına gelir.[4] 1996'da monografi cins, botanikçiler Finn Borchsenius ve Rodrigo Bernal "ironik bir şekilde, türlerin Aiphanes genellikle yoğun bitki örtüsünde tespit edilmesi ve bulunması çok zordur ve bu nedenle, en kötü toplanan neotropikal palmiyeler arasındadır. "[4]
Kaynaklanıyor

Bazı türler tek gövdeli iken, diğerleri çok gövdeli (Sezgi ) kümeler. Gövde boyutundaki varyasyonla birleştiğinde, bu cins içinde çeşitli büyüme biçimleri üretir - ormanın alt kanopisine dönüşen tek (tek saplı) palmiyeler, ormanın alt kısmında büyüyen soliter veya sezpitoz palmiyeler ve ışıltılı yer üstü sapı olmayan palmiyeler.[3]
İki tür, bir ışıltılı büyüme alışkanlığı—A. acaulis ve A. spicata. Diğer iki tür -A. ulei ve A. weberbaueri - hem aközleşmiş popülasyonlarda hem de yer üstü gövdeleri üreten popülasyonlarda görülür. Bazı türler, alışılmadık bir büyüme formu olan tek gövdeli alt avuç içleridir. Aiphanes grandis ve A. minima 10 metreden (33 ft) daha uzun boylu büyüyen tek gövdeli palmiyelerdir, geri kalanı ise çok gövdeli alt türlerdir. Çok gövdeli palmiyeler, tek bir ana gövdeye sahip bitkilerden ve birkaç bazal emiciler 20 yoğun şekilde paketlenmiş gövdeden oluşan öbekleri keser. Tek bir tür içinde çeşitli büyüme biçimleri bulunabilir ve bu, habitat ve çevresel koşullardan etkileniyor gibi görünmektedir.[3]
Yapraklar
Yaprakları Aiphanes türler genellikle sıkıca bölünmüştür - yaprak ekseninin her iki yanında tüy benzeri veya eğrelti otu benzeri bir modelde yaprakçıklar sıraları ortaya çıkar. Bunun tek istisnası A. macroloba hangisi tüm yapraklar. Genellikle spiral olarak düzenlenirler, ancak bazı türlerin farklı yaprak düzeni, palmiye fidelerinde normal olan ancak yetişkinler arasında nadir görülen bir durumdur. Eski yaprak tabanları, kökler hariç, gövdeden temiz bir şekilde ayrılır A. hirsuta subsp. fosteriorum, genellikle gövdenin yeni kısımlarına tutturulmuş eski yaprak tabanlarına sahiptir.[5]
Yapraklar dikenlidir ancak derecesi tür içinde ve arasında değişir. Yaprak kılıfları her zaman yoğun dikenlidir ancak dikenler genellikle küçülür ve yaprakların uçlarına doğru seyrekleşir.[5]
Dikenler

Dikenler karakteristiktir Aiphanes ve diğer üyeleri subtribe Baktridina. Bitkilerin hemen hemen her yerinde bulunurlar ve özellikle gövdede, yaprak diplerinde ve pedinkül. İçinde Aiphanesdikenler, bitkinin dış dokularından oluşur ve diğer bitki organlarının değiştirilmesiyle elde edilmez. 1 milimetreden (0,04 inç) az ila 25 santimetreden (9,8 inç) daha uzun aralıklar.[6]
Çiçekler
Aiphanes türler pleonantiktir — yaşamları boyunca defalarca çiçek açarlar — ve monoecious Bu, ayrı erkek ve dişi çiçekler olduğu anlamına gelir, ancak bireysel bitkiler her iki tür çiçeği de taşır. İçinde Aiphanes, erkek ve dişi çiçekler aynı anda doğar çiçeklenme. Genellikle her birinde sadece tek bir çiçeklenme taşınır düğüm, olmasına rağmen A. gelatinosa genellikle tek bir düğümde üçlü gruplar halinde bulunur. Çiçeklenme genellikle bir pedinkül ve bir saptan oluşan bir ana eksenden oluşur. Rachis. Rachel, kendileri çiçekleri taşıyan daha küçük dallar olan rachillae'yi taşırken, pedinkül, rachileri bitkinin gövdesine bağlayan ana saptır. Bazı türlerde ikinci dereceden dallanma vardır - raşilaların kendileri dallıdır ve çiçekler bu dallarda taşınır.[7]
Çiçekler genellikle üçlü gruplar halinde doğar - bir dişi çiçek ve iki erkek çiçek. Bazı türlerde dört çiçekten oluşan gruplar (iki erkek ve iki dişi) rapor edilmiştir. Çiçeklenmenin en ucunda, ağacın ekseninden uzakta, erkek ve dişi çiçek üçlülerinin yerini erkek çiçek çiftleri alır. Çiçek rengi yeterince bilinmemektedir. Korunan çiçekler zamanla renklerini yitirdikleri için canlı bitkilerden kaydedilmelidir ve bu türlerin vahşi doğadaki kayıtları eksiktir. Erkek çiçekler iki gruba ayrılır: krem veya sarı çiçekleri olanlar ve çiçeklerinde bir miktar mor olanlar. Dişi çiçekler, erkek çiçeklerden daha az bilinir.[7]
Polen taneleri genellikle küresel ila elipsoid şekildedir, bazen üçgen şeklindedir, uzun eksenleri boyunca yaklaşık 20 ila 30 mikrometre ve çapları 20 ila 30 um'dir. Tipik olarak monosülkat, meridiyonosülkat veya daha nadiren trichotomosulcate.[8] sulkus polen tanesinin yüzeyi boyunca uzanan bir karıktır ve genellikle tozlaşmanın meydana geldiği yerdir. Monosulcate polen, polen tanesinin kutbu boyunca uzanan tek bir oluğa sahiptir. Meridionosulcate poleninde polen tanesinin ekvatoru boyunca uzanan bir karık bulunur.[9] Trichotomosulcate poleninin ise üç oluğu vardır.[10] Polenin dış tabakası büyük veya küçük ölçüde sırtlar, dikenler veya siğillerle kaplıdır. Bu "şekillendirme", sinekle tozlaşan türlerde daha belirgin olma eğilimindedir ve böcekler veya arılar tarafından tozlaşanlarda daha az belirgindir.[8]
Meyve
Meyvesi Aiphanes türler genellikle kırmızı, küresel, tek tohumludur drupe. İnce bir cilt (veya epikarp Düz veya dikenli olabilen) etli olanı örter mezokarp, tipik olarak turuncu ve tatlıdır. Mezokarp A. horrida en yüksek raporlardan birine sahip karoten herhangi bir bitki ürününün içeriği ve ayrıca zengindir protein. endokarp Tohumun etrafını saran, kahverengi veya siyah renkte ve olgunlukta çok serttir.[11] Tohumlar ince ve açık kahverengidir tohum kabuğu (veya testa) ve beyaz endosperm tatlı ve tadı bir şekilde Hindistan cevizi.[11]
Karyotip
Yayınlanan kromozom sayımı iki tür için var, Aiphanes minima ve A. horrida; haploid kromozom sayıları 15 ile 18 arasında değişmektedir. Borchsenius ve Bernal, avuç içlerinde doğru kromozom sayımlarını elde etmenin zor olduğunu ve kromozom sayılarındaki farklılıkların bu zorlukları yansıtabileceğini bildirmektedir.[12]
Taksonomi
| Alt kabile üyeleri arasındaki ilişki Baktridina, dayalı plastid DNA soyoluşu.[13] *Aiphanes aculeata botanik eşanlamlıdır A. horrida. |

Aiphanes yerleştirilir alt aile Arecoideae, kabile Cocoseae ve Bactridinae alt kabilesi, cinslerle birlikte Desmoncus, Baktris, Akrocomia ve Astrocaryum.[14]
Alman botanikçi cinsinin 1932 tarihli revizyonunda Max Burret 32 tür tanıdı. Bunlardan 17'si, çoğunlukla Alman botanikçiler tarafından yapılan koleksiyonlara dayanan yeni türlerdi. Wilhelm Kalbreyer 1877 ile 1881 arasında kuzey Kolombiya'da. tür kavramı ve doğal popülasyonlarda bulunan çeşitliliğe aşina olmayan Burret, hemen hemen her örneği ayrı bir türe yerleştirdi. İkinci Dünya Savaşı sırasında Berlin Herbaryumunun bombalanması, bu 32 türün 13'ünün bilinen tek koleksiyonunu yok etti ve durumu daha da karmaşık hale getirdi.[15]
Uluslararası Botanik İsimlendirme Kodu her türün bir ile temsil edilmesini gerektirir tür koleksiyonu.[16] Burret'in tip koleksiyonlarının yok edilmesi, birçok türü yalnızca orijinal tasvirlerinden bilinen, genellikle resimlerden yoksun bıraktı. Diğer örnekler (neotipler olarak adlandırılır), 1989'da Rodrigo Bernal ve meslektaşları tarafından bunların yerini alacak şekilde belirlendi.[17] veya Borchsenius ve Bernal tarafından cinsin 1996 monografisinde.[18] Bernal ve meslektaşları, Kalbreyer'in kuzey Kolombiya'daki seyahatlerinin izini sürmeye ve orijinal koleksiyonların bulunduğu yere mümkün olduğu kadar yakından örnek toplamaya çalıştılar.[17]
Burret bölündü Aiphanes iki alt türe, Brachyanthera ve Macroanthera. 11 tür yerleştirildi Macroantherageri kalanlar yerleştirilirken Brachyanthera.[15] Borchsenius ve Bernal, 1996 tarihli monografilerinde, bu alt türlerin uygulanabilirliğini sorguladılar. Kabul ettiler eğer Macroanthera üç türe indirildi (A. horrida, A. eggersii ve A. minima) uygulanabilir bir gruplama oluşturabilir, ancak bu, Brachyanthera aşırı heterojen. Sonuç olarak, Burret'in alt türleri kullanımını terk ettiler.[19]
Burret'in cinsi tanımlamasını izleyen otuz yıl içinde, toplam tür sayısını 47'ye çıkaran 15 tür daha tanımlandı.[15] Borchsenius ve Bernal, bu isimlerin çoğu eşanlamlıydı Amerikalı botanikçi olmasına rağmen George Proctor topaklanma kararlarına katılmadılar A. acanthophylla içine A. minima.[20] Borchsenius ve Bernal ayrıca yeni bir tür tanımladılar: Aiphanes spicatatoplam kabul edilen tür sayısı 22'ye çıkarıldı.[3] İki durumda, bilinen yegane koleksiyonların yok edilmesi, bir ismin eşanlamlı olduğundan kesinlikle emin olmayı imkansız hale getirdi.[21] Akım Seçilmiş Bitki Ailelerinin Dünya Kontrol Listesi, Rafaël Govaerts tarafından, Kraliyet Botanik Bahçeleri, Kew, Borchsenius ve Bernal'ın monografisinin yayınlanmasından bu yana açıklanan dört tür dahil olmak üzere 26 türü tanır.[22]
Burret bölündü Aiphanes iki alt türe, Brachyanthera ve Macroanthera. 11 tür yerleştirildi Macroantherageri kalanı ile Brachyanthera.[15] Borchsenius ve Bernal, 1996 tarihli monografilerinde, bu alt türlerin uygulanabilirliğini sorguladılar. Kabul ettiler eğer Macroanthera üç türe indirildi (A. horrida, A. eggersii ve A. minima) uygulanabilir bir gruplama oluşturabilir, ancak bu, Brachyanthera aşırı heterojen. Sonuç olarak, Burret'in alt türleri kullanımını terk ettiler.[19]
Tarih
Bu cinsteki bir türün en erken botanik tanımı Fransız botanikçi tarafından yapılmıştır. Charles Plumier, iki türü ziyaretlerine dayanarak tanımlayan Batı Hint Adaları 1689 ve 1695 arasında. Plumier'in her iki türünün de artık Aiphanes minima. Aynı tür Hollandalı botanikçi tarafından tanımlandı Nikolaus Joseph von Jacquin 1763'te. İspanyol botanikçi José Celestino Mutis ayrıntılı bir açıklama üretti A. lindeniana ve bu türün çizimleri ve ne olduğu düşünülüyor A. horrida 1779'da.[4]
1791'de Joseph Gaertner bir tür dahil Aiphanes onun içinde De Fructibus et Seminibus Plantarum, onu çağırmak Baktris minima. Bu, cinsin herhangi bir üyesi için geçerli olarak yayınlanan en eski isimdir. İsim Aiphanes Alman botanikçi tarafından icat edildi Carl Ludwig Willdenow 1801'de tek bir türü tanımladı, A. aculeata1806'da.[4]
Jacquin adı kullanmıştı Caryota Horrida Willdenow tarafından tanımlanan aynı türe ait (ve aynı birey olabilir) bir bitkiyi tanımlamak. Borchsenius ve Bernal, Jacquin'in açıklaması için Willdenow'un adına öncelik veren 1809 yayın tarihini aktarırlar.[4] Bununla birlikte, daha yeni Dünya Kontrol Listesi (2006), Jacquin'in açıklaması için 1801 yayın tarihi verir. A. horrida türler için doğru isim.[23]
1816'da Alexander von Humboldt, Aimé Bonpland ve Carl Sigismund Kunth tarif Martinezia caryotifolia, listesine başka bir ad eklemek eş anlamlı için A. horrida. Orijinal tanı karakterlerinden beri Martinezia mevcut hiçbir türe uymuyordu, Kunth tarafından yeniden tanımlandı. M. caryotifolia. Sonuç olarak, Martinezia yerine geldi Aiphanes ve ikinci isim 1847 ile 1932 arasında nadiren kullanıldı. 1857'de Hermann Karsten yeni bir cins yarattı, Mararaiki Kolombiyalı türü barındırmak için, M. bicuspidata (daha sonra eşanlamlı olarak gösterilecek A. horrida) ve M. erinacea (şimdi A. erinacea ). Hermann Wendland diriltmeye teşebbüs Aiphanes 1878'de birleşme Martinezia ve Marara içine girdi, ancak teklifi göz ardı edildi. 1901'de Hatip F. Cook iki yeni tür oluşturdu—Curimaiçine koyduğu A. minima, ve Tilmiahangi barındırdı A. horrida.[24] 1932'de bir türü yayınladıktan sonra Martinezia, Burret cins hakkındaki fikrini değiştirdi ve onu Aiphanes. Bu, cinsin mevcut sınırlandırılmasına yol açtı.[15]
Türler
Tarafından kabul edilen türler Seçilmiş Bitki Ailelerinin Dünya Kontrol Listesi:[2]
- Aiphanes acanthophylla (Mart.) Burret - Porto Riko, Dominik Cumhuriyeti
- Aiphanes acaulis Galeano ve R.Bernal - Kolombiya
- Aiphanes bicornis Cerón & R.Bernal - Ekvador
- Aiphanes buenaventurae R.Bernal & Borchs. - Valle del Cauca, Kolombiya
- Aiphanes chiribogensis Borchs. & Balslev - Ekvador
- Aiphanes deltoidea Burret - Kolombiya, Peru, kuzeybatı Brezilya
- Aiphanes duquei Burret - Kolombiya
- Aiphanes eggersii Burret - Ekvador, Peru
- Aiphanes erinacea (H.Karst.) H.Wendl. - Kolombiya, Ekvador
- Aiphanes jelatinosa H.E.Moore - Kolombiya, Ekvador
- Aiphanes graminifolia Galeano ve R.Bernal - Kolombiya
- Aiphanes grandis Borchs. & Balslev - Ekvador
- Aiphanes hirsuta Burret - Kolombiya, Ekvador, Panama, Kosta Rika
- Aiphanes horrida (Jacq.) Burret - Trinidad, Kolombiya, Venezuela, Peru, kuzeybatı Brezilya, Bolivya
- Aiphanes leiostachys Burret - Kolombiya'da Antioquia
- Aiphanes Lindeniana (H.Wendl.) H.Wendl. - Kolombiya
- Aiphanes linearis Burret - Kolombiya'da Antioquia ve Valle del Cauca
- Aiphanes makroloba Burret - Kolombiya, Ekvador
- Aiphanes minima (Gaertn.) Burret - Saint Lucia, Barbados
- Aiphanes multipleks R.Bernal & Borchs. - Valle del Cauca, Kolombiya
- Aiphanes parvifolia Burret - Kolombiya
- Aiphanes pilaris R.Bernal - Kolombiya
- Aiphanes simpleks Burret - Kolombiya
- Aiphanes spicata Borchs. & R.Bernal - Peru
- Aiphanes stergiosii S.M. Niño - Batı Venezuela'daki Portuguesa Eyaleti
- Aiphanes tricuspidata Borchs., M.Ruíz ve Bernal - Kolombiya, Ekvador
- Aiphanes ulei (Dammer) Burret - Kolombiya, Ekvador, Peru, kuzeybatı Brezilya
- Aiphanes verrucosa Borchs. & Balslev - Ekvador
- Aiphanes weberbaueri Burret - Ekvador, Peru
Dağıtım ve durum
Cins Aiphanes aralıkları Dominik Cumhuriyeti ve Panama kuzeyde Trinidad ve Tobago doğuda, karşısında Kolombiya ve aşağı And Dağları -e Bolivya. İçinde Brezilya sadece sınır boyunca meydana gelir Peru. Aiphanes esas olarak Güney Amerika'dır — bir tür (A. hirsuta ) Panama'da ve diğer iki ülkede (A. horrida ve A. minima) Karayipler'de bulunur. Aiphanes minima, hangisi endemik Karayip adalarına kadar, Güney Amerika anakarasında bulunmayan tek türdür. olmasına rağmen A. horrida tarafından bildirildi Guyana ve güney Venezuela bu raporlar ile doğrulanmadı Herbaryum kuponlar.[25]
Aiphanes horrida en yaygın olarak dağılan türdür. Aralıkları Trinidad Bolivya'ya var ama yok Ekvador ve kuzey Peru. Diğer türler, daha dar aralıklara sahiptir. çeşitlilik merkezi batı Kolombiya ve Ekvador'da ve kuzeydoğu Peru'da bir başka küçük.[25] 2006 IUCN Kırmızı Listesi üç tür içerir nesli tükenmekte habitat tahribatı ile—A. grandis,[26] A. leiostachys[27] ve Bir verrükoza[28]—Ve üç kişi daha düşünüldü savunmasız aynı tehdide -A. chiribogensis,[29] A. duquei[30] ve A. lindeniana.[31] Rodrigo Bernal ve Gloria Galeano Bu listeyi, Kolombiya palmiyelerinin durumunun 2005 yılında gözden geçirilmesiyle genişletti. İki türü listelediler kritik tehlike altında —A. graminifolia, ilk olarak 2002'de tanımlanan bir tür ve A. leiostachys (IUCN Kırmızı Listesinde tehlike altında olarak sınıflandırılmıştır). İki türü tehlike altında olarak sınıflandırdılar.A. acaulis ve A. parvifolia - ve savunmasız iki tür -A. gelatinosa ve A. pilaris. Ayrıca altı türü şöyle sınıflandırdılar: yakın tehdit —A. erinacea, A. hirsuta, A. lindeniana (IUCN Kırmızı Listesine göre savunmasız), A. linearis, A. macroloba ve A. simpleks.[32] Bu türlere yönelik tehditler listelenmemiş ancak Jens-Christian Svenning, A. erinacea sınırlı dağılımı ve bozulmuş ormanlarda yenilenme yeteneğinin zayıf olması nedeniyle ağaç kesimi tehdidi altındaydı.[33] Bunlara ek olarak, A. deltoidea batıda yaygın olarak dağıtılan Amazon yağmur ormanları, 1994 yılında Francis Kahn ve Farana Moussa tarafından nadir bir tür olarak sınıflandırıldığı için o kadar düşük yoğunluklarda bulunur.[34]
Habitat ve ekoloji

Aiphanes türler, ormanın alt ve alt kanopisinin palmiyeleri. En çok dağılan türler, A. horrida, ikisinde de oluşur tropikal kuru orman ve daha nemli orman türlerinde, ancak dağılımında yukarıların en yağışlı ormanlarıyla çakışan bir boşluk vardır. Amazon Havzası. Diğer iki tür, A. minima ve A. eggersiidaha kuru ortamlarda da bulunur; A. eggersii yılda 500 mm (20 inç) kadar az yağış alan bölgelerde bulunur. Kalan türler bulunur dağ yüksek rakımlarda veya ıslak - genellikle çok ıslak - ova ormanlarında ormanlar, yıllık 9.000 mm'ye (350 inç) kadar yağış alan alanlar dahil.[25]
Tarafından ziyaret kayıtları tozlayıcılar yalnızca birkaç tür için vardır, ancak bunların çoğu, türlerin böcekler tarafından tozlaştığını göstermektedir. Çiçekler A. chiribogensis küçük miktarlarda üretmek nektar ama kokusuzdur. Meyve sinekleri (Drosophilidae ), mantar sivrisinekleri (Mycetophilidae, Sciaridae ), tatarcıklar (Cecidomyiidae, Ceratopogonidae ) ve mikromotlar (Lepidoptera ) bu çiçekleri ziyaret ederken kaydedildi, ancak arılar ve uçan sinekler değildi. Aiphanes eggersii arılar ve muhtemelen rüzgar tarafından tozlaştığı düşünülüyordu. Meyve sinekleri (Drosophilidae ), uçan sinekler (Syrphidae), tatarcıklar (Ceratopogonidae ) ve yaprak böcekleri (Chrysomelidae) çiçeklerini ziyaret ederken kaydedildi. A. erinaceaama arılar değildi. Aiphanes horrida bildirildiğine göre rüzgarla tozlaştı, arılar (Meliponidae ), yabani otlar (Curculionidae ) ve böcekler (Hemiptera ). Sinek ve bitler A. simpleks.[35]
Meyvesi A. horrida vitamin ve enerji bakımından zengindir ve birçok hayvan tarafından yenme olasılığı yüksektir. Yağlı kuşlar meyvesini yediği ve tohumlarını dağıttığı bildirilmiştir. Ağacın dikenli yapısına rağmen sincapların da meyveyi tükettiği bildirilmektedir.[35] Meyveleri, çiçekleri ve tohumları A. minima tarafından tüketiliyor savunmasız Saint Vincent Amazon (Amazona guildingii)[36] ve aynı zamanda potansiyel olarak önemli bir gıda türü olarak kabul edilir. kritik tehlike altında Porto Rikolu amazon (Amazona vittata).[37]
Bazı türler kümelenmiş dağılımlar gösterir. Yetişkinlerin kümelenmiş dağılımını ve her ikisinde de fidelerin sınırlı sayıda toplanmasını açıklamak için dağılma sınırlamasına başvurulmuştur. A. erinacea Ekvador'da[38] ve A. minima Porto Riko'da.[37] Benzer şekilde, nadir A. lindeniana ve A. simpleks Kolombiya ormanlarında, sınırlı tohum üretimi ve kuş ve memeli meyveciller tarafından tohum dağılımının sınırlı etkinliği ile bağlantılı olabilir.[39]
Kullanımlar
Aiphanes türlerin uzun bir insan kullanım geçmişi vardır. Ait olduğu düşünülen karbonize tohum kalıntıları A. horrida arkeolojik sitelerde bulundu Kolombiya yaklaşık 2800 yılına kadar uzanan BP;[40] Bu türün tohumları halen tüketilmekte ve yerel pazarlarda ticareti yapılmaktadır.[41] Aiphanes horrida olduğu gibi süs olarak da yaygın olarak ekilir. A. minima.[42][43] Meyvesi veya tohumları A. deltoidea,[44] A. eggersii,[45] A. linearis[46] ve A. minima[47] hepsi yerel olarak tüketilmektedir. palmiye kalbi nın-nin A. macroloba tarafından tüketilir Coaiquer Kuzeybatı Güney Amerika halkı.[48] Aiphanol izole edilmiş bir bileşik A. horrida, önemli gösterdi engelleyici karşı aktivite siklooksijenazlar;[49] bu enzimlerin inhibisyonu, aşağıdaki semptomlardan kurtulmayı sağlayabilir: iltihap ve ağrı.[50]
Notlar
- ^ Borchsenius ve Bernal (1996), s. 33–34
- ^ a b Seçilmiş Bitki Ailelerinin Kew Dünya Kontrol Listesi
- ^ a b c d Borchsenius ve Bernal (1996), s. 4
- ^ a b c d e Borchsenius ve Bernal (1996), s. 2
- ^ a b Borchsenius ve Bernal (1996), s. 8-9
- ^ Borchsenius ve Bernal (1996), s. 6-8
- ^ a b Borchsenius ve Bernal (1996), s. 11–14
- ^ a b Borchsenius ve Bernal (1996), s. 16–22
- ^ Hoen, Peter (1999). "Polen ve Spor Terminolojisi Sözlüğü". Paleobotani ve Palinoloji Laboratuvarı, Michigan Teknoloji Üniversitesi. Arşivlenen orijinal 18 Mayıs 2010'da. Alındı 17 Mart 2010.
- ^ Nadot, S .; A. Forchioni; L. Penet; J. Sannier; A. Ressayre (2006). "Monokotlarda erken polen gelişimi ve açıklık düzeni arasındaki bağlantılar". Protoplazma. 228 (1–3): 55–64. doi:10.1007 / s00709-006-0164-4. PMID 16937055. S2CID 28497230.
- ^ a b Borchsenius ve Bernal (1996), s. 14–15
- ^ Borchsenius ve Bernal (1996), s. 26
- ^ Asmussen, Conny B .; John Dransfield; Vinnie Deickmann; Anders S. Barfod; Jean-Christophe Pintaud; William J. Baker (2006). "Palmiye ailesinin (Arecaceae) yeni bir alt aile sınıflandırması: plastid DNA filogenisinden kanıtlar". Linnean Topluluğu Botanik Dergisi. 151 (1): 15–38. doi:10.1111 / j.1095-8339.2006.00521.x.
- ^ Dransfield, John; Natalie W. Uhl; Conny B. Asmussen; William J. Baker; Madeline M. Harley; Carl E. Lewis (2005). "Palmiye Ailesinin Yeni Filogenetik Sınıflandırması, Arecaceae". Kew Bülten. Kraliyet Botanik Bahçeleri, Kew. 60 (4): 559–69. JSTOR 25070242.
- ^ a b c d e Borchsenius ve Bernal (1996), s. 3
- ^ "7. Madde". Uluslararası Botanik İsimlendirme Kodu (Viyana Kodu). Alındı 20 Mart 2010.
- ^ a b Bernal, Rodrigo G .; Gloria Galeano-Garcés; Andrew Henderson (1989). "W. Kalbreyer Tarafından Toplanan Kolombiya Palmiyelerinin Neotipifikasyonu". Takson. Uluslararası Bitki Taksonomisi Derneği (IAPT). 38 (1): 98–107. doi:10.2307/1220905. JSTOR 1220905.
- ^ Borchsenius ve Bernal (1996)
- ^ a b Borchsenius ve Bernal (1996), s. 33
- ^ George R. Proctor, Acevedo-Rodríguez & Strong (2005), s. 138–139
- ^ Borchsenius ve Bernal (1996), s. 87
- ^ Govaerts, R.; J. Henderson; S.F. Zona; D.R. Hodel; A. Henderson (2006). "Aramak Aiphanes". Arecaceae Dünya Kontrol Listesi. Mütevelli Heyeti Kraliyet Botanik Bahçeleri, Kew. Alındı 2010-01-05.
- ^ "Aiphanes horrida". Kraliyet Botanik Bahçeleri, Kew: Seçilmiş Bitki Ailelerinin Dünya Kontrol Listesi. Alındı 2010-01-04.
- ^ Borchsenius ve Bernal (1996), s. 2–3
- ^ a b c Borchsenius ve Bernal (1996), s. 26–30
- ^ Montúfar, R .; N. Pitman (2003). "Aiphanes grandis". IUCN Tehdit Altındaki Türlerin Kırmızı Listesi. 2003: e.T43757A10826566. Alındı 4 Şubat 2020.
- ^ Bernal, R. (1998). "Aiphanes leiostachys". IUCN Tehdit Altındaki Türlerin Kırmızı Listesi. 1998: e.T38942A10158301. Alındı 4 Şubat 2020.
- ^ Montúfar, R .; N. Pitman (2003). "Aiphanes verrucosa". IUCN Tehdit Altındaki Türlerin Kırmızı Listesi. 2003: e.T38069A10095781. Alındı 4 Şubat 2020.
- ^ Montúfar, R .; N. Pitman (2003). "Aiphanes chiribogensis". IUCN Tehdit Altındaki Türlerin Kırmızı Listesi. 2003: e.T38752A10147801. Alındı 4 Şubat 2020.
- ^ Bernal, R. (1998). "Aiphanes duquei". IUCN Tehdit Altındaki Türlerin Kırmızı Listesi. 1998: e.T38941A10158248. Alındı 4 Şubat 2020.
- ^ Bernal, R. (1998). "Aiphanes Lindeniana". IUCN Tehdit Altındaki Türlerin Kırmızı Listesi. 1998: e.T38943A10158352. Alındı 4 Şubat 2020.
- ^ Bernal, Rodrigo; Gloria Galeano (2006). "Kolombiya Palmiyelerinde (Arecaceae) Tehlike: 18 yılda değişim". Linnean Topluluğu Botanik Dergisi. 151 (1): 151––163. doi:10.1111 / j.1095-8339.2006.00530.x.
- ^ Svenning, Jens-Christian (1998). "Kuzeybatı Ekvador'daki bir And yağmur ormanı parçasındaki palmiye türlerinin yerel dağılımı üzerindeki arazi kullanımının etkisi". Biyoçeşitlilik ve Koruma. 7 (12): 1529–1537. doi:10.1023 / A: 1008831600795. S2CID 34175807.
- ^ Kahn, Francis; Farana Moussa (1994). "Peru palmiye ağaçlarının çeşitliliği ve koruma durumu" (PDF). Biyoçeşitlilik ve Koruma. 3 (3): 227–241. doi:10.1007 / BF00055940. S2CID 32827272.
- ^ a b Borchsenius ve Bernal (1996), s. 30–32
- ^ Culzac-Wilson, Lystra (2005). St. Vincent Papağan için Türleri Koruma Planı Amazona guildingii. Puerto de la Cruz, Tenerife: Loro Parque Fundación.
- ^ a b Inman, Faith M .; Thomas R. Wentworth; Martha Groom; Cavell Brownie; Russ Lea (2007). "Porto Rikolu Papağanı restore etmek için yapay gölgelik boşlukları kullanmak (Amazona vittata) tropikal ağaç tarlalarında habitat ". Orman Ekolojisi ve Yönetimi. 243 (2–3): 169–177. doi:10.1016 / j.foreco.2007.02.003.
- ^ Svenning, J.-C. (2001). "Çevresel heterojenlik, işe alım sınırlaması ve tropik dağ yağmur ormanlarında (Maquipucuna, Ekvador) palmiye ağaçlarının orta ölçekli dağılımı". Tropikal Ekoloji Dergisi. 17 (1): 97–113. doi:10.1017 / S0266467401001067.
- ^ Correa-Gómez, Diego F .; Orlando Vargas-Ríos (2009). "Regeneración de palmas in bosques nativos y plantaciones del Sanctuario de Fauna y Flora Otún-Quimbaya (Risaralda, Kolombiya)". Caldasia (ispanyolca'da). 31 (2): 195–212. Arşivlenen orijinal 2011-09-27 tarihinde.
- ^ Morcote-Ríos, Gaspar; Rodrigo Bernal (2001). "Yeni Dünyadaki Arkeolojik Alanlardaki Palmiye Kalıntıları (Palmae): Bir İnceleme". Botanik İnceleme. New York Botanik Bahçesi Basın. 67 (3): 309–350. doi:10.1007 / BF02858098. JSTOR 4354394. S2CID 46582757.
- ^ Borchsenius ve Bernal (1996), s. 47
- ^ Henderson, Andrew; Gloria Galeano; Rodrigo Bernal (1995). Amerika Palms için Saha Rehberi. Princeton, New Jersey: Princeton University Press. s. 171–174. ISBN 0-691-08537-4.
- ^ Riffle Robert Lee (2008). Palmiye Kereste Basın Rehberi. Kereste Basın. s. 30–32. ISBN 978-0-88192-776-4.
- ^ Borchsenius ve Bernal (1996), s. 52
- ^ Borchsenius ve Bernal (1996), s. 54
- ^ Borchsenius ve Bernal (1996), s. 69
- ^ Borchsenius ve Bernal (1996), s. 72
- ^ Borchsenius ve Bernal (1996), s. 71
- ^ Banwell, Martin G .; Anna Bezos; Satish Chand; Gerd Dannhardt; Werner Kiefer; Ulrike Nowe; Christopher R. Parish; G. Paul Savage; Holger Ulbrich (2003). "Stilbenolignan (±) -aiphanol ve çeşitli türdeşlerin yakınsak sentezi ve ön biyolojik değerlendirmeleri". Organik ve Biyomoleküler Kimya. 1 (14): 2427–2429. doi:10.1039 / b305106d. PMID 12956057.
- ^ Goodsell, David S. (2001-05-01). "Siklooksijenaz". RCSB Protein Veri Bankası. Alındı 2010-01-17.
Referanslar
- Acevedo-Rodríguez, Pedro; Mark T. Strong (2005). Porto Riko ve Virgin Adaları Monokotiledonları ve Gymnospermleri. Birleşik Devletler Ulusal Herbaryumundan katkılar. 52. Washington, D.C .: Smithsonian Enstitüsü. s. 1–415.
- Borchsenius, Finn; Rodrigo Bernal (Aralık 1996). "Aiphanes (Palmae) ". Flora Neotropica. Flora Neotropica Organizasyonu adına New York Botanik Bahçesi Basın. 70: 1–94. JSTOR 4393869.