Arkeol - Archaeol
Arkeol ana çekirdek zar lipidlerinden biridir Archaea, hayatın üç alanından biri. Archaea'yı diğerlerinden ayıran temel özelliklerden biri bakteri ve ökarya arkeolün önemli bir rol oynadığı zar lipidleridir. Bu nedenle arkeol, özellikle antik arkealar için bir biyobelirteç olarak geniş bir şekilde kullanılmaktadır. metanojenler, aktivite.[1]
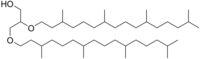
Archaeol, genellikle iki fitanil zincirinin, a'nın sn-2 ve sn-3 pozisyonlarına bağlanmasıyla oluşur. gliserol molekül. Oldukça dallanmış yan zincirlerin, çok düşük olanı hesaba kattığı düşünülmektedir. geçirgenlik Arkeol bazlı zar, arkeanın aşırı ortamlara en önemli uyarlamalarından biri olabilir.
 | |
| İsimler | |
|---|---|
| IUPAC adı 2,3-Bis (3,7,11,15-tetrametilheksadekoksi) propan-1-ol | |
| Diğer isimler Archaeol lipid; 2,3-Di-Ö-fitanil-sn-gliserol; 2,3-Bis [(3,7,11,15-tetrametilheksadesil) oksi] -1-propanol | |
| Tanımlayıcılar | |
3 boyutlu model (JSmol ) | |
| MeSH | arkeol + lipit |
PubChem Müşteri Kimliği | |
CompTox Kontrol Paneli (EPA) | |
| |
| Özellikleri | |
| C43H88Ö3 | |
| Molar kütle | 653.174 g · mol−1 |
Aksi belirtilmedikçe, veriler kendi içlerindeki malzemeler için verilmiştir. standart durum (25 ° C'de [77 ° F], 100 kPa). | |
| Bilgi kutusu referansları | |
Kimya
Archaeol, archaea'da yaygın olarak bulunan bir lipittir. Standart arkeol 2,3-di-O-fitanil-sn-gliserol, ikisiyle fitanil zincirleri gliserolün sn-2 ve sn-3 konumuna bağlanma eter tahviller. 2,3-sn-gliserol yapısı ve eter bağı bağı, arkea lipidlerinin aşağıdakilerden iki temel farkıdır: bakteri ve ökarya 1,2-sn-gliserol ve çoğunlukla ester bağları kullanan.[2] Doğal arkeol, üçü için 3R, 7R, 11R konfigürasyonlarına sahiptir. kiral merkezler içinde izoprenoit zincirler. Karmaşıklığa katkıda bulunan dört yapısal varyasyon vardır. membran lipitleri işlev ve özelliklerde. İki fitanil zinciri, makrosiklik arkeol vermek için 36 üyeli bir halka oluşturabilir. Hidroksile arkeol, ilk başta hidroksile fitanil zincirlerine sahiptir. üçüncül karbon atom, sesterterpanil arkeol ise C2 gliserol veya her iki karbonda ikame eden C25 sesterterpanil zincirleri ile fitanil yan zincirlerine sahiptir. Standart arkeol ile aynı karbon iskeletine sahip, ancak fitanil yan zincirlerinde bir veya birden fazla çift bağa sahip doymamış arkeol de keşfedilmiştir.[3]
İki arkeol molekülü oluşturmak için kafa kafaya bağlantı kurabilir kaldarchaeol (bir tipik gliserol dialkil gliserol tetraeter, GDGT), arkelerde en yaygın tetraeter lipidlerinden biridir.







Biyolojik rol ve sentez
Biyolojik rol
Archaeol, şimdiye kadar analiz edilen tüm arkelerde, en azından eser miktarda bulundu. Çoğu durumda dier çekirdek lipitlerinin% 100'ünü temsil eder. nötrofilikhalofiller[3] ve kükürt bağımlı termofiller (en çekirdek lipitleri tetraeter lipitler olsa da). Metanojenler, standart arkeol dışında hidroksikarkeol ve makrosiklik içerir ve sesterterpanil zinciri içeren arkeol, alkalifilik aşırı halofillerin karakteristiğidir. Tetraeter lipidlerinin arkelerde de yaygın olarak bulunması dikkat çekicidir.[2]
Lipozomlar (en az bir çift lipit tabakasına sahip küresel bir kesecik) arkeadan lipidler tipik olarak son derece düşük geçirgenlik moleküller ve iyonlar için, hatta protonlar dahil. Tarafından indüklenen iyon geçirgenliği iyonoforlar (zarlar boyunca iyon taşıyıcılar) da oldukça düşüktür ve sadece yumurtanınki ile karşılaştırılabilir fosfatidilkolin (çok yaygın bir biyolojik membran bileşeni) 37˚C'de sıcaklık c.a'ya yükseldiğinde. 70˚C.[4][5] Nazaran bakteri ve ökarya, izoprenoit arkeolün yan zincirleri oldukça dallıdır. Bu yapısal farkın, arkelerin aşırı ortamlara uyum sağlamasına olanak tanıyan tüm büyüme sıcaklığı aralığı boyunca arkelerin geçirgenliğini düşürdüğüne inanılmaktadır.[6]
Sentez süreci
Arkeol, genellikle arke hücrelerinde fosfolipid olarak bulunur. Tamamen doymuş arkeol fosfolipidinin sentetik yolu şu şekilde ilerler: izoprenlerin baştan kuyruğa bağlanmasıyla izoprenoid yan zincirlerin sentezi, gliserol-1-fosfat omurgasına eter bağlantısı, CDP arkeol oluşumu, polar baş grubu bağlanması ve çift doygunluk tahviller. Bunu takiben tetraeter lipidler, daha sonra baştan başa bağlantı yoluyla dimerizasyon reaksiyonu ile sentezlenebilir.[7]
Archaea, bakteri ve ökaryaya kıyasla farklı biyosentetik izoprenoid zincir yollarına sahiptir. İzoprenoidin öncülleri C5 birimleridir izopentenil pirofosfat (IPP) ve dimetilalil pirofosfat (DMAPP), yaşamın her üç alanı için evrenseldir. Genel olarak, iki bileşik bakterilerde 2-C-metil-D-eritritol 4-fosfat / 1-deoksi-D-ksilüloz 5-fosfat yolu (MEP / DOXP yolu) ile sentezlenir ve mevalonat (MVA) yolu ile sentezlenir. çoğu ökaryada. Archaea'da IPP ve DMAPP sentezi, son üç adımda klasik MVA yolundan farklı olan ve kalan dört adımı paylaşan alternatif bir MVA yolunu takip eder.[7]
Bakterilerdeki eter lipitleri
İzoprenoid zinciri ile gliserol arasındaki eter bağlantısını içeren arkeol, arkeler için ikna edici bir biyobelirteç olarak kabul edilmekle birlikte, bazılarında eter membran lipidleri de keşfedilmiştir. aerobik ve anaerobik bakteriler, bir ester bağı ve alkil zincirlerine bir eter bağı olan lipitler dahil. Pek çok kesinlikle anoksik bakteri ve birkaç aerobik tür, gliserolün sn-1 konumuna bağlı bir alkil zincirine sahip olan plazmalojenler (Pla) içerir. vinil-eter bağı. Archaea'ya benzer şekilde, bu lipitlerin bakterilerin olumsuz ortamlara direncini artırdığı düşünülmektedir. Daha çarpıcı olan, alkil zincirlerini (ancak izoprenoid zincirleri değil) eter yoluyla gliserol moleküllerine bağlayarak, arkeole benzer şekilde oluşan izoprenoid olmayan dialkil gliserol dieter lipidlerin (DGD) ve dallı dialkil gliserol tetraeter lipidlerin (brGDGT) keşfidir. bağlantı. Bu lipidlerin yalnızca yan zincirlerdeki arkea eter lipidlerinden ve gliserol üzerindeki bağlanma pozisyonlarından farklı olması oldukça dikkat çekicidir. DGD, termofilik bakterilerde rapor edilmiştir. mezofilik bakteri ve toplanma miksobakteriler.[8][9]
Lipid biyobelirteci olarak kullanılır
Sedimanlardaki arkeol tipik olarak diyajenez sırasında arke membran fosfolipitlerinin hidrolizinden kaynaklanır. Yüksek koruma potansiyeli nedeniyle, genellikle organik jeokimyacılar tarafından, özellikle metanojen biyokütlesi ve aktivitesi için arkea aktivitesi için bir biyolojik belirteç olarak tespit edilir ve kullanılır. Bir metanojen vekil olarak, Michinari Sunamura ve diğerleri tarafından kullanılır. sedimentlerdeki metanojenleri doğrudan ölçmek için Tokyo Körfezi,[10] ve ayrıca Katie L. H. Lim ve ark. göstergesi olarak metanojenez suya doymuş topraklarda.[11] C. A. McCartney vd. sığırlarda metan üretimi için bir vekil olarak kullandı.[12]
Bu arada, eski biyojeokimyayı anlamaya yardımcı olmak için de kullanılır. Richard D. Pancost ve arkadaşları tarafından bir biyobelirteç olarak kullanılmıştır. yeniden inşa etmek için Holosen biyojeokimya ombrotrofik turbalıklar.[13] Ian D. Bull ve arkadaşları tarafından yürütülen bir pilot çalışma. ayrıca arkeolü, sindirim sistemlerinin fermente edilmesi arasındaki farkları ortaya çıkarmak için biyobelirteç olarak kullandı. ön bağırsak ve arka bağırsak antik otçul memeliler.[14]
Ek olarak, bozulmamış arkeolün farklı bozunma kinetiği ve kaldarchaeol arkeolün kaldarchaeole oranı, bir tuzluluk yayla göllerinde vekaleten, paleosalite çalışmaları için bir araç sağlar.[15]
Archaeol, yan zincirleri şu şekilde korunarak bazı durumlarda hidrolize de olabilir. fitan veya bozulmamış redoks koşullarına bağlı olarak.[16]
Ölçüm
Arkeolü analiz etmek için, lipidler genellikle geleneksel Bligh-Dyer prosedürü ile ekstrakte edilir,[17] genellikle bunu fraksiyonlama (ince katman veya kolon kromatografisi ile) takip eder ve türetme. Kazuhiro Demizu vd.[18]ve Sadami Ohtsubo vd.[19] çekirdek lipitlerin nihayet tabi tutulduğu asit Bligh ve Dyer ekstraksiyonu, asit muamelesi ve türevlendirmeyi içeren benzer prosesler önerildi. kromatografi.
Bir numunede bulunan arkeol konsantrasyonunu belirlemek için, yaygın olarak kromatografi teknolojileri kullanılır. yüksek performanslı sıvı kromatografisi (HPLC),[18][19][20] gaz kromatografisi (GC),[21] ve süper kritik sıvı kromatografisi (SFC),[22][23] ile kütle spektrometrisi (MS) genellikle tanımlamaya yardımcı olmak için başvurdu.
Ayrıca bakınız
Referanslar
- ^ Ricardo Cavicchioli (2007) tarafından düzenlendi, Archaea, Washington, DC: ASM Press, ISBN 978-1-55581-391-8, OCLC 172964654CS1 bakimi: ek metin: yazarlar listesi (bağlantı)
- ^ a b Koga, Y Nishihara, M Morii, H Akagawa-Matsushita, M (1993). "Metanojenik bakterilerin eter polar lipidleri: yapılar, karşılaştırmalı yönler ve biyosentezler". Mikrobiyolojik İncelemeler. 57 (1): 164–82. doi:10.1128 / MMBR.57.1.164-182.1993. OCLC 680443863. PMC 372904. PMID 8464404.CS1 bakimi: birden çok ad: yazarlar listesi (bağlantı)
- ^ a b Gambacorta, A .; Gliozzi, A .; De Rosa, M. (1995). "Archaeal lipidler ve biyoteknolojik uygulamaları". Dünya Mikrobiyoloji ve Biyoteknoloji Dergisi. 11 (1): 115–131. doi:10.1007 / BF00339140. PMID 24414415.
- ^ Yamauchi, Kiyoshi; Doi, Kuniyuki; Kinoshita, Masayoshi; Kii, Fumiko; Fukuda, Hideki (Ekim 1992). "Archaebacterial lipid modelleri: 1,2-diphytanylglycero-3-phosphocholine'den oldukça tuz toleranslı membranlar". Biochimica et Biophysica Açta (BBA) - Biyomembranlar. 1110 (2): 171–177. doi:10.1016 / 0005-2736 (92) 90355-p. ISSN 0005-2736. PMID 1390846.
- ^ Yamauchi, Kiyoshi; Doi, Kumiyuki; Yoshida, Yoichi; Kinoshita, Masayoshi (Mart 1993). "Archaebacterial lipids: 1,2-diphytanyl-sn-glycero-3-phosphocoline'dan oldukça proton geçirimsiz membranlar". Biochimica et Biophysica Açta (BBA) - Biyomembranlar. 1146 (2): 178–182. doi:10.1016/0005-2736(93)90353-2. ISSN 0005-2736. PMID 8383997.
- ^ Koga Yosuke (2012). "Arkaeal ve Bakteriyel Lipid Membranlarının Termal Adaptasyonu". Archaea. 2012: 789652. doi:10.1155/2012/789652. ISSN 1472-3646. PMC 3426160. PMID 22927779.
- ^ a b Jain Samta (2014). "Arkeal membran eter lipitlerinin biyosentezi". Mikrobiyolojide Sınırlar. 5: 641. doi:10.3389 / fmicb.2014.00641. PMC 4244643. PMID 25505460.
- ^ Grossi, Vincent; Mollex, Damien; Vinçon-Laugier, Arnauld; Hakil, Floransa; Pacton, Muriel; Cravo-Laureau, Cristiana (2015). "Anaerobik Bakterilerde Mono- ve Dialkyl Gliserol Eter Lipidler: Mezofilik Sülfat İndirgeyici Desulfatibacillum alkenivorans PF2803T'den Biyosentetik İçgörüler". Uygulamalı ve Çevresel Mikrobiyoloji. 81 (9): 3157–3168. doi:10.1128 / AEM.03794-14. PMC 4393425. PMID 25724965.
- ^ Lorenzen, Wolfram; Ahrendt, Tilman; Bozhüyük, Kenan A J; Bode, Helge B (2014-05-11). "Bakteriyel eter lipit biyosentezinde çok işlevli bir enzim rol oynar". Doğa Kimyasal Biyoloji. 10 (6): 425–427. doi:10.1038 / nchembio.1526. ISSN 1552-4450. PMID 24814673.
- ^ Sunamura, Michinari; Koga, Yosuke; Ohwada, Kouichi (1999-11-01). "Tokyo Körfezi Sedimanlarında Arkeol Lipitleri Kullanılarak Metanojenlerin Biyokütle Ölçümü". Deniz Biyoteknolojisi. 1 (6): 562–568. doi:10.1007 / PL00011811. ISSN 1436-2228. PMID 10612681.
- ^ Lim, Katie L. H .; Pancost, Richard D .; Hornibrook, Edward R. C .; Maxfield, Peter J .; Evershed Richard P. (2012). "Arkeol: Suya Doymuş Topraklarda Metanogenezin Bir Göstergesi". Archaea. 2012: 896727. doi:10.1155/2012/896727. ISSN 1472-3646. PMC 3512251. PMID 23226972.
- ^ Dewhurst, R. J .; Yan, T .; Bull, I.D .; McCartney, C.A. (2013-02-01). "Sığırlarda metan üretimi için moleküler bir vekil olarak arkeolün değerlendirilmesi". Journal of Dairy Science. 96 (2): 1211–1217. doi:10.3168 / jds.2012-6042. ISSN 0022-0302. PMID 23261373.
- ^ Pancost, Richard D .; McClymont, Erin L .; Bingham, Elizabeth M .; Roberts, Zoë; Charman, Dan J .; Hornibrook, Edward R.C .; Blundell, Anthony; Chambers, Frank M .; Lim, Katie L.H. (Kasım 2011). "Ombrotrofik bataklıklarda bir metanojen biyolojik belirteç olarak arkeol". Organik Jeokimya. 42 (10): 1279–1287. doi:10.1016 / j.orggeochem.2011.07.003.
- ^ Gill, Fiona L .; Dewhurst, Richard J .; Dungait, Jennifer A.J .; Evershed, Richard P .; Ives, Luke; Li, Cheng-Sen; Pancost, Richard D .; Sullivan, Martin; Bera, Subir (Mayıs 2010). "Archaeol - modern ve eski otçul memelilerde ön bağırsak fermantasyonu için bir biyobelirteç mi?". Organik Jeokimya. 41 (5): 467–472. doi:10.1016 / j.orggeochem.2010.02.001.
- ^ Wang, Huanye; Liu, Weiguo; Zhang, Chuanlun L .; Jiang, Hongchen; Dong, Hailiang; Lu, Hongxuan; Wang, Jinxiang (Ocak 2013). "Kuzeydoğu Qinghai-Tibet Platosu'ndaki dağlık göllerde bir tuzluluk göstergesi olarak arkeolün kaldarchaeol'e oranının değerlendirilmesi". Organik Jeokimya. 54: 69–77. doi:10.1016 / j.orggeochem.2012.09.011.
- ^ Rowland, S.J. (Ocak 1990). "Metanojenik bakterilerin laboratuvarda olgunlaşmasıyla asiklik izoprenoid hidrokarbon üretimi". Organik Jeokimya. 15 (1): 9–16. doi:10.1016 / 0146-6380 (90) 90181-x. ISSN 0146-6380.
- ^ Bligh, E. G .; Dyer, W.J. (Ağustos 1959). "Toplam Lipit Ekstraksiyonu ve Saflaştırmanın Hızlı Bir Yöntemi". Kanada Biyokimya ve Fizyoloji Dergisi. 37 (8): 911–917. doi:10.1139 / o59-099. ISSN 0576-5544. PMID 13671378. S2CID 7311923.
- ^ a b Demizu, Kazuhiro; Ohtsubo, Sadami; Kohno, Shuhei; Miura, Isao; Nishihara, Masateru; Koga Yosuke (1992). "Yüksek performanslı sıvı kromatografisi ile etere bağlı gliserolipidlerin analizine dayalı metanojenik hücrelerin kantitatif tespiti". Fermentasyon ve Biyomühendislik Dergisi. 73 (2): 135–139. doi:10.1016 / 0922-338x (92) 90553-7. ISSN 0922-338X.
- ^ a b Ohtsubo, S (Mayıs 1993). "Asetiklastik metanojenlerin miktarının belirlenmesi ve etere bağlı gliserolipidlerin analizine dayalı olarak doğal ortamlarda toplam metanojenik hücrelerin tahmini için hassas bir yöntem". FEMS Mikrobiyoloji Ekolojisi. 12 (1): 39–50. doi:10.1016 / 0168-6496 (93) 90023-z. ISSN 0168-6496.
- ^ Martz, Robert F .; Sebacher, Daniel I .; White, David C. (Şubat 1983). "Çevresel örneklerde metan oluşturan bakterilerin biyokütle ölçümü". Mikrobiyolojik Yöntemler Dergisi. 1 (1): 53–61. doi:10.1016/0167-7012(83)90007-6. ISSN 0167-7012. PMID 11540801.
- ^ Smith, G.C .; Floodgate, G.D. (Ekim 1992). "Metanojenik biyokütleyi tahmin etmek için kimyasal bir yöntem". Kıta Sahanlığı Araştırması. 12 (10): 1187–1196. Bibcode:1992CSR .... 12.1187S. doi:10.1016 / 0278-4343 (92) 90078-x. ISSN 0278-4343.
- ^ Holzer, Gunther U .; Kelly, Patrick J .; Jones, William J. (Temmuz 1988). "Bir hidrotermal delikli metanojenden lipitlerin ve süper kritik akışkan kromatografisiyle ilişkili havalandırma tortusunun analizi". Mikrobiyolojik Yöntemler Dergisi. 8 (3): 161–173. doi:10.1016/0167-7012(88)90017-6. ISSN 0167-7012.
- ^ King, Jerry (2002-01-22), "Lipid Ekstraksiyonu, Fraksiyonasyonu ve Reaksiyonları için Süper Kritik Sıvı Teknolojisi", Lipid Biyoteknolojisi, CRC Press, doi:10.1201 / 9780203908198.ch34, ISBN 9780824706197