Macroolithus - Macroolithus - Wikipedia
| Macroolithus | |
|---|---|
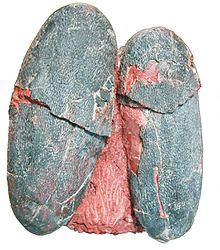 | |
| Bir çift Macroolithus yaotunensis yumurtalar | |
| Yumurta fosili sınıflandırması | |
| Temel kabuk tipi: | Ornitoid |
| Morfotip: | Ornitoid-ratit |
| Oofamily: | †Elongatoolithidae |
| Oogenus: | †Macroolithus Zhao, 1975 |
| Oospecies yazın | |
| †Oolithes rugustus Genç, 1965 | |
| Oospecies | |
| |
Macroolithus bir Oogenus (fosil yumurta cinsi) dinozor yumurtası e ait oofamily Elongatoolithidae. oospecies yazın, M. rugustus, başlangıçta artık geçersiz olan oogenus adı altında tanımlandı Oolithes. Diğer üç oospecies bilinmektedir: M. yaotunensis, M. mutabilis, ve M. lashuyuanensis. Nispeten büyük, iki katmanlı yumurta kabuğuna sahip uzun yumurtalardır. Yuvaları büyük, eşmerkezli çift yumurta halkalarından oluşur. Kabuğunda yuvaların kamufle edilmesine yardımcı olabilecek mavi-yeşil pigmentasyon kanıtı var.
Macroolithus içeren yumurtalar bulundu oviraptorid Dinozor embriyolar benzeyen Heyuannia. Oviraptorid ve elongatoolithids arasında birden fazla başka ilişki (embriyo içeren diğer yumurtalar, yuvalarda düşünen ebeveynler ve bir çift kabuklu Macroolithus-bir oviraptoridin pelvisinde korunan yumurtalar gibi), ebeveynin Macroolithus bir oviraptoriddi.
İçinde bulunur Üst Kretase orta ve doğu Asya oluşumları; fosiller Moğolistan, Kazakistan ve Çin'de bulundu. Güney Çin'deki Nanxiong oluşumunda, Macroolithus fosiller, muhtemelen Kretase-Tersiyer sınırı, geleneksel olarak olmayanın yok oluşunu işaret ettiği varsayılır.kuş dinozorlar. Bazı paleontologlar, bu oluşumdaki dinozor yumurtalarının kaydını ani bir felaketten ziyade kademeli bir yok olma olayını destekleyici olarak yorumladılar. Bununla birlikte, diğer paleontologlar bu yorumların yalnızca erken dönemlerde erozyon ve yeniden birikme eserlerine dayandığına inanmaktadır. Paleojen.
Açıklama

Macroolithus yumurtalar, 16 ila 21 cm (6,3 ila 8,3 inç) uzunluğunda büyük boyları ve özellikle kaba süslemeleriyle karakterize edilir.[1][2] Mikroyapıları literatürde iyi tanımlanmamıştır,[1] ancak genellikle tipik uzama taşı desenini takip eder:[2] Yumurta kabuğu, iki yapısal katman halinde düzenlenmiştir (memeli katman ve sürekli katman). Sürekli katman, yumurta kabuğunun dış kısmını oluşturur; yumurta kabuğu birimleri, katman bir süreklilik gibi görünecek şekilde birbirine kaynaşmıştır. Memeli veya koni olarak bilinen iç katman, yumurta kabuğu birimlerinin tabanını oluşturan koni şeklindeki yapılardan oluşur.[1] İçinde Macroolithussürekli katman, memeliler katmandan iki ila üç kat daha kalındır.[1][2] Yumurtaların çok çeşitli gözenek yapısı vardır. Gaz iletkenliği gözenek boyutu ve yoğunluğu ile ilişkili olduğundan, bu, yumurtaların değişken ortamlarda bırakıldığı anlamına gelebilir.[1]
Oospecies
Dört oospecies Macroolithus şu anda tanınıyor:
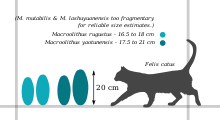
- M. rugustus - oospecies yazın Bu yumurtalar şurada bulunur: Nanxiong ve Jiangxi güney Çin'de; aynı zamanda Nemegt Oluşumu nın-nin Moğolistan ve Manrakskaya Svita içinde Kazakistan.[3][4] Boyut olarak 16,5 ila 18 cm (6,5 ila 7,1 inç) uzunluğunda ve 7,5 ila 8,5 cm (3,0 ila 3,3 inç) arasında değişir. Yumurta kabuğu 0,8 ila 1,7 mm (0,031 ila 0,067 inç) kalınlığındadır ve bunun yaklaşık 1 / 4'ü memeliler katmanıdır.[5] Dış yüzey, yumurtanın dış merkezi etrafında düzensiz düğüm zincirleriyle (ramotüberküloz süslemesi) kaplanır ve uçlarda daha çukurlu, ağ benzeri bir desene (sagenotüberküloz) geçer.[6][7]
- M. yaotunensis yerli Henan ve Nanxiong.[8][5] Benzer mikro yapıya ve süslemeye sahiptir. M. rugustus, ancak biraz daha büyük (17,5 - 21 cm (6,9 - 8,3 inç) uzunluğunda) ve daha kalın bir kabuğa (1,4 - 1,9 mm (0,055 - 0,075 inç)) sahip olmasıyla ayırt edilebilir.[1][2] Ayrıca aksine M. rugustus, M. yaotunensis memeli tabakası ile sürekli tabakası arasında dalgalı bir sınır gösterir.[5]
- M. mutabilis - Bu Moğol oospecies birkaç yumurta kabuğu parçalarından bilinmektedir. Parçalar 1.3 ila 2.0 mm (0.051 ila 0.079 inç) kalınlığında olup, M. rugustus. Tamamlandığında, yumurta muhtemelen 17 cm (6,7 inç) üzerinde ölçülmüştür. Mikhailov'un "anormal" özellikler olarak adlandırdığı özellik için benzersizdir: birkaç parça yarık şeklinde gözenek kanallarına sahiptir ve süslemesizdir.[2]
- M. lashuyuanensis - bu oospecies, içinde bulunan bir yumurta kabuğu parçasına dayanmaktadır. Nanxiong Havzası nın-nin Guangdong. 2,3 ila 2,7 mm (0,091 ila 0,106 inç) kalınlığıyla yumurta kabuğu en kalın olanıdır. Macroolithus oospecies. Ayrıca, sürekli katmanının üst kısmına yakın koyu, yoğun bir bant ve sürekli ve memeli katmanları arasında kademeli bir geçiş ile ayırt edilir.[9]
Renklendirme
Bazı örnekler M. yaotunensis kırmızımsı kahverengi pigmentin izlerini korumak protoporfirin ve mavi-yeşil pigment Biliverdin, birçok modern kuşun yumurtalarında kullanılan aynı pigmentler. Kabukta biliverdinin baskınlığı ile gösterildiği gibi yumurtalar büyük olasılıkla yoğun bir mavi-yeşil renkteydi. Renklendirmenin amacı belirsizdir, ancak modern kuşlarda renklendirme veya çiftleşme sonrası cinsel işaret olarak hizmet edebilir. Renkli yumurtalar aynı zamanda modern kuşlarda aktif baba bakımı ile ilişkilendirilir ve oviraptorid erkeklerin yumurtalarına baktıkları hipotezine daha fazla destek verir.[10]
İsimler ve etimoloji
İsim Macroolithus Yunan köklerinden türetilmiştir makro ("büyük" anlamına gelir)[11] ve oolithus ("taş yumurta" anlamına gelir), oogenus isimleri için geleneksel son ek.[12] Çince adı (巨 形 蛋 Jùxíng dàn) benzer şekilde "dev yumurta" anlamına gelir.[5] Oospesifik epifetler yaotunensis (Çince : 窑 屯 yáotún) ve Lashuyuanensis (Çince : 腊 树 园 Làshùyuán) oospecies'in orijinal olarak keşfedildiği yerleri onurlandırın (Yaotun[5] ve Lashuyuan[9] sırasıyla Guangdong bölgeleri), Latince son eki ekleyerek -ensis menşe yerini belirtmek için;[13] isim Mutabilis Latince değişkendir.[2] Adını verdiğinde "Oolithes" engebeli, Yang (1965) 'ın etimolojisini vermedi engebeli, ancak Çince formu (粗 皮 cūpí) "pürüzlü cilt" anlamına gelir.[3]
Paleobiyoloji ve ebeveynlik

Yetişkin oviraptorosaur iskeletleri veya elongatoolithid yumurtalı embriyolar arasındaki birçok ilişki ( Macroolithus) bunu gösterin Macroolithus ve diğer elongatoolithids, oviraptorosaurlar tarafından atıldı.[1][14] Bir oviraptorosaur iskeleti Üst Kretase nın-nin Çin 2005 yılında tanımlanmıştır; pelvisinin içinde iki kabuklu uzatma taşı yumurtası korunmuştur. Bu, oviraptorozorların iki işlevsel yumurta kanalları her ikisinin de aynı anda yumurta üreteceği yer. Sato iken et al. yumurtaları özellikle Macroolithus, yumurtaların birbirine çok benzediğini belirttiler M. yaotunensis, daha ince bir yumurta kabuğu ile. Bununla birlikte, ince yumurta kabuğu, basitçe, anne öldüğünde kabuğun oluşumunu tamamlamamış olmasından veya fosilleşmeden önce kabuğun biyokimyasal çözünmesinden kaynaklanıyor olabilir.[15]
Elongatoolithid yuvaların üzerinde veya yakınında çok sayıda farklı oviraptorid türü bulunmuştur, bu da oviraptorid ebeveynlerin kara kara düşünmek yumurtalarında, büyük olasılıkla uzun süreler boyunca.[16] Oviraptorid ve Troodontid yumurtalara baba tarafından bakılıyordu, belki de çok eşli sistemi.[17][10] Ebeveyne göre büyük yumurta büyüklüğü göz önüne alındığında, bir anne bir seferde yalnızca iki yumurta bırakırdı, bu nedenle tek bir yuvanın yumurtalarına birden fazla dişi katkıda bulunmuş olabilir.[15][17]
Yuvalar

Çok iyi korunmuş Macroolithus yuvalar biliniyor, temsil ediyor M. rugustus ve M. yaotunensis.[3][5] Çok iyi korunmuş bir debriyaj M. yaotunensis iki, muhtemelen üç dairesel katman halinde dizilmiş 20 yumurta içerir. Bu yuva tamamlandığında 40 veya daha fazla yumurta içerebilir.[3]
Tanaka et al. (2015), modern arkozorlar (kuşlar, timsahlar ve dinozorları içeren grup) arasında, yumurta kabuklarının gözenekliliğinin, yumurtaların açık yuvalara gömülüp gömülmediğini doğru bir şekilde tahmin etmek için kullanılabileceğini buldu. Sonucuna vardılar Macroolithus yuvalar ya açıktı ya da kısmen bitki örtüsüyle kaplıydı.[18] Yumurtaların mavi-yeşil rengi, yuvayı avcılardan gizlemek için bir kamuflaj görevi görebilirdi.[10]
M. rugustus yumurtalar Tsagan Khushu Moğolistan'da bir kıyı koloni yuvalama alanını temsil eder.[2]
Embriyolar
Birkaç Macroolithus yumurtalar, içindeki oviraptoridlerin embriyonik kalıntılarını korur.[19] İçinde embriyo bulunan iki yumurta Üst Kretase Nanxiong Oluşumu yakın Ganzhou, Jiangxi sevk edildi M. yaotunensis Bu embriyolardan biri çok daha yüksek derecede kemik gelişimi gösterir (kemikleşme ) diğerinden; kemikleşmiş arka bacakları ve birkaç omuru korur. Servikal omurların sadece kemikleşmemiş olması centra ve sinir kemerleri ama aynı zamanda kemikleşmiş zigapofizler, Cheng'i yönetti et al. bu embriyonun aslında bir yumurtayı temsil edebileceği sonucuna varmak, çünkü modern tavuklar ve skuas bu zigafizler yumurtadan çıkana kadar kıkırdaklıdır. Ayrıca, embriyonun ayak anatomisi (özellikle, embriyonun ayak anatomisi) metatarslar ) oviraptoridinkine benzer Heyuannia huangi, bu yumurtaların ait olduğunu belirten H. huangi veya benzer bir tür. Diğer yumurtanın sadece arka bacakları korunmuş, çok daha az gelişmiş bir embriyo vardır. Bu, tür düzeyinde karşılaştırmaya izin vermezken, tibia (incik kemiği) onun bir oviraptorosaur olduğunu doğrular.[19]

Aynı oluşumdan embriyo içeren üç yumurta daha 2016 yılında Wang tarafından tanımlanmıştır. et al. Bu yumurtaların birbirine çok benzediğini belirtmiş olsalar da M. yaotunensis, Elongatoolithidae'den daha düşük herhangi bir ootaksona yönlendirmeyi reddettiler çünkü Macroolithus açıkça tanımlanmamış ve revizyona ihtiyacı var. Bu yumurtalardaki embriyolar, oviraptoridler arasında en iyi korunmuş olanlardır ve oviraptorid ontogenisi hakkında yeni bilgiler sağlar. Örnekler, oviraptoridler olgunlaştıkça kafataslarının anteroposteriora (önden arkaya) göre dorsoventral olarak (yukarıdan aşağıya) daha hızlı büyüdüğünü gösteren nispeten sığ bir kafa gösterir. Bu büyüme modeli, theropodlar arasında alışılmadık bir durumdur, ancak aynı zamanda türetilmiş ("ileri") tyrannosaurids. Ayrıca olağandışı, gelişimin bu erken aşamasında bile, burun kemikleri kaynaşmış. Tesadüfen, tyrannosaurids de gelişimin erken dönemlerinde nazallerin kaynaşmasını gösterir.[14]
Patoloji ve yok olma
Birçok Macroolithus Güney Çin'deki örnekler yumurta kabuğunun iç yüzeyinde çift veya çok katmanlı konilere sahiptir. patolojik durum ovo'da yumurta olarak bilinir.[20] Özellikle en yakın yumurtalar arasında yaygındır. Kretase-Tersiyer (K-T) sınırı, sonunu temsil eden Mesozoik Dönem. Bu patoloji, daha yüksek konsantrasyonla ilişkilidir. eser elementler sevmek Co, Cr, Cu, Mn, Ni, Pb, Sr, V, ve Zn.[20] Modern kuşlar üzerinde yapılan deneyler, bu elementlerin yüksek seviyelerine maruz kalmanın onları yumurta kabuğuna dahil etmelerine neden olacağını göstermiştir, ancak patolojik çok tabakalamanın arkasındaki kesin mekanizma bilinmemektedir. Bu anormallikler muhtemelen yumurtaların çıkma kabiliyetini etkiledi ve dinozorların yok oluşunda rol oynamış olabilir.[14][20] Bununla birlikte, embriyonik, üç çok katmanlı MacroolithusGanzhou'dan gelen yumurta benzeri yumurtalar etkilenmemiş görünüyor.[14]
K-T sınırı ile ilişkilidir anormal derecede yüksek miktarda iridyum, Dünya'nın kabuğunda nadir bulunan, ancak asteroitlerde ve Dünya'nın çekirdeğinde nispeten yaygın olan bir element. Bu, bir göktaşı etkisinin dinozorların yok olmasına neden olduğunun kanıtı olarak kullanılmıştır. Alvarez hipotezi ). Bununla birlikte, bazı paleontologlar yok olma olayını ve iridyum anomalisini, volkanik faaliyetin neden olduğu daha kademeli iklim değişikliğine bağlamaktadır. Deccan Tuzakları. Nanxiong Havzası, K-T sınırını içerdiği için bu tartışmayla özel bir ilişkiye sahiptir. Bununla birlikte, sınırın konumu ve yok olma süresi tartışmaya konu olmuştur. Zhao et al. (2002 ve 2009), Deccan Tuzakları volkanizması ile tutarlı bir zaman dilimi içinde en az iki iridyum salgılayan olay olduğunu, çeşitliliğin kademeli olarak azalması ve Nanxiong Havzasından fosil yumurtalarının nihayet kaybolmasıyla bağlantılı olduğunu varsaydılar. Yorumlarına göre, Macroolithus 250.000 yıl boyunca hayatta kaldı Paleosen.[21][22][23] Ancak Buck et al. (2004) bu iddialara itiraz etti. enkaz akar karıştırıldı ve yeniden işlendi sedimanlar sınırın yakınında, iridyum anomalisinin bulanıklaşmasına ve her iki tarafta yumurta kabuğu parçalarının ortaya çıkmasına neden olur.[24]
Sınıflandırma
Fosilleşmiş yumurtalar, kendi parataksonomik sistemlerine paralel olarak sınıflandırılır. Linnaean taksonomisi. Macroolithus, Elongatoolithus, ve Nanhsiungoolithus bu sistemde adı geçen ilk oogenera idi; ailede sınıflandırıldılar Elongatoolithidae.[5] Kladistik analiz aynı zamanda Macroolithus yakın Elongatoolithusdiğer elongatoolithids ile birlikte bir clade.[25][26] Dört oospecies bilinmektedir: M. rugustus, M. yaotunensis, M. mutabilis, ve M. lashuyuanensis.[1][27] Oogenus'un mikro yapıları yetersiz tanımlanmıştır[1] ve bu nedenle revizyona ihtiyaç duyabilir.[14] Mihaylov tarif ettiğinde M. mutabilis, Zhao'nun orijinal çizimlerinde hiçbir mikro yapısal farklılık bulamadı. M. rugustus ve M. yaotunensis, ama yapmadı eş anlamlı hale getirmek oospecies çünkü Çinli paleontologlar onları farklı görüyordu.[2]
Tarih
Keşifler
Macroolithus Yumurtalar ilk olarak Güney Çin'de, öncü Çinli paleontolog tarafından keşfedildi. Yang Zhongjian. O bölgeden birkaç fosil yumurta kalıntılarını 1965'te tanımladı. Modern fosil yumurta parataksonomisinin ortaya çıkmasından önce çalışarak, onlara türler olarak isimler verdi. Oolithes,[3] Eskiden çeşitli fosil yumurtaları için kullanılan ve artık kullanılmayan bir isim.[28][29] 1975'te Çinli paleontolog Zhao Zikui, modern parataksonomik sistemi prototipleyerek oofamiller, oogenera ve oospecies hiyerarşik bir sistem yarattı. Zhao, Yang'ın O. rugustus yeni oogenusa, Macroolithus, onu iki oospecise bölmek, M. yaotunensis ve M. rugustus. Ayrıca Amerikan vahşetlerinin "Oolithes" carlylensis (şimdi farklı bir elongatoolithid oogenus olarak sınıflandırılmıştır, Macroelongatoolithus[1]) olarak yeniden birleştirilebilir Macroolithus carlylensis.[5]
1991'de Rus paleontolog Konstantin Mihaylov, Zhao'nun parataksonomik adlandırma sistemine dayalı olarak fosil yumurtaların modern sınıflandırmasını tanıttı. Sınıflandırdı "Ö." carlylensis ailede Spheroolithidae, ancak aksi takdirde Zhao'nun 1975 Macroolithus.[30] 1994 yılında M. mutabilis, yeni bir oospecies Macroolithus, Moğolistan'da bulunan kalıntılara dayanıyor.[2]
2000 yılında, dördüncü bir oospecies, "M." turolensis, İspanyol paleontologlar Olga Amo-Sanjuán, José Ignacio Canudo ve Gloria Cuenca-Bescós tarafından İspanya'dan gelen materyallere dayanarak tanımlandı. Ancak, bu oospecies'in yeni materyalleri 2014 yılında ortaya çıkarıldığında, kendi özüne taşınmıştır. Guegoolithus, ayrıca Spheroolithidae'ye taşındı.[27]
2005 yılında, yumurtalar muhtemelen M. yaotunensis bir fosil oviraptorid iskeletinin pelvisinin içinde eşleştirilmiş olarak keşfedildi.[15] 2008'de Çinli paleontologlar Cheng Yen-nien, Ji Qiang, Wu Xiao-chun ve Shan Hsi-yin, Çin'de oviraptorozorların embriyonik kalıntılarını içeren ilk yumurtayı temsil eden bir çift yumurta keşfettiler. Her iki yumurtaya da sevk edildi M. yaotunensis.[19] Dördüncü oospecies, M. lashuyuanensis, 2009 yılında Çinli paleontologlar Fang Xiao-si, Li Pei-xian, Zhang Zhi-jun, Zhang Xian-qiu, Lin You-li, Guo Sheng-bin, Cheng Ye-ming, Li Zhen-yu, Zhang tarafından tanımlanmıştır. Xiao-jun ve Cheng Zheng-wu.[9]
Paleobiyolojik yorumlar

1994 yılında Mihaylov bunu önerdi Macroolithus büyük bir theropodun yumurtalarını temsil etti, özellikle Tarbosaurus, yumurtaların büyüklüğüne ve büyük ölçüde örtüşen dağılıma dayanmaktadır.[2] Bununla birlikte, bir elongatoolithid yumurtanın içinde bir oviraptorid embriyonun keşfi, bu hipoteze şüphe düşürdü.[31] 1990'ların sonlarında birden fazla oviraptorosaur-elongatoolithid birlikteliğinin keşfedilmesiyle,[32] elongatoolithids yumurtalarının oviraptorosaurian dinozorlarına ait olduğu kabul edildi.[1] Oviraptorid ebeveyni, Macroolithus ne zaman, 2008'de oviraptorid embriyoları andırıyor Heyuannia bir çiftin içinde keşfedildi M. yaotunensis Kiangsi'de yumurta.[19]
Erken dönemlerde paleontologlar, Macroolithus gömülü olan yuvalar. Mou 1992, yüksek gaz iletkenlik değerlerini kaydetti Macroolithus yumurtalar ve bu nedenle çok nemli bir ortama bırakıldıkları, yer altına veya bir höyüğün içine gömüldükleri sonucuna varmıştır.[33] Deeming (2006) benzer bir sonuç buldu. Bu, oviraptorosauridlerin yumurtalarında kuş gibi kuluçkaya yattığına dair kanıtlarla çelişiyor gibi görünüyor, ancak Deeming şunu önerdi: Oviraptor yumurtalarını bir tepeye gömdü ve sonra Oviraptor ve akrabaları yumurtalarla doğrudan temas kurmak yerine kuluçka yapmak için bir yuva tepesinin üstüne oturdu.[34] Ancak Tanaka et al. (2015) bu sonuçları istatistiksel kesinlik eksikliği nedeniyle eleştirdi. Modern yumurtalarla yapılan karşılaştırmalara dayanarak, Macroolithus açık veya kısmen kapalı yuvalara döşendiği tahmin edilmektedir.[18] Wiemann et al. (2017) ayrıca Mou ve Deeming'i sadece yumurtaların orta bölümünde yumurta kabuğu gözenekliliğini ölçtükleri ve gözenek yoğunluğunun kutupların yakınında çok daha düşük olduğu gerçeğini hesaba katmadıkları için eleştirdiler. Bu, toplam yumurta kabuğu gözenekliliğinin fazla tahmin edilmesine ve dolayısıyla gaz iletkenlik değerinin fazla tahmin edilmesine yol açacaktır.[10]
Güney Çin'den Macroolithus ve diğer yumurtaların neslinin tükenmesi de farklı yorumlara sahip bir tarihe sahipti. 1990'larda, aralarında ünlü yumurta uzmanı Zhao Zikui'nin de bulunduğu Çinli paleontologlar, Kretase'nin son 200.000 ila 300.000 yılı boyunca dinozor yumurtası çeşitliliğinde kademeli bir azalma gözlemledi. Macroolithus sınıra kadar uzanan.[20][35] Etki hipotezinin aksine, yok oluşun, iz ağır metallerin konsantrasyonunu artıran, dinozorların yumurta kabuğu ve embriyo gelişimini olumsuz yönde etkileyen, popülasyonun kademeli olarak azalmasına ve çökmesine neden olan uzun süreli bir kuraklığın sonucu olduğunu varsaydılar.[35] Zhao et al. 2002'de bu hipotezi revize ederek, Macroolithus Deccan Tuzaklarının volkanizmasından kaynaklanır.[21] 2004 yılında, Buck et al. Bu yoruma itiraz etti ve görünürdeki kademeli yok oluşun çökeltilerin yeniden işlenmesinin neden olduğu bir yanılsama olduğunu iddia etti.[24] Zhao et al. (2009) yok olma olayının kademeli olduğunu ileri sürdü.[22]
Dağıtım ve paleoekoloji
Macroolithus Çin, Moğolistan ve Kazakistan'daki sayısız Geç Kretase bölgesinden bilinmektedir.[1]
Henan
İçinde Henan, Macroolithus yaotunensis diğer elongatoolithids ile bir arada bulunur Elongatoolithus andrewsi ve E. elongatus, Hem de Ovaloolithus ve Paraspheroolithus of Hugang, Luyemiao, ve Sigou Oluşumları. Bu oluşumlar Geç Kretase'de göl veya palustrine çevre. Dinozor vücut fosilleri nadiren aynı birimlerde bulunur, ancak Troodontidler, tiranozorlar ve hadrosaurlar aynı bölgeden bilinmektedir.[8]
Shandong
Bazı yumurtalar geçici olarak atanmış Macroolithus -den bilinmektedir Wangshi Grubu içinde Laiyang, Shandong.[36] Oluşumları Wangshi Grubu yatırıldı Alüvyonlu fanlar, örgülü kanallar ve sığ göller. Yaygın fosiller arasında hadrosauridler ve dinozor yumurtaları bulunur.[37]
Güney Çin
M. rugustus, M. yaotunensis, ve M. lashuyuanensis hepsi bilinmektedir Nanxiong Havzası içinde Guangdong.[5][9] Bu oluşum birincil olarak o bölgedeki akarsular, nehirler ve göllerden biriktirildi.[21][35] Boyunca yayılır K / T sınırı, bu noktada dinozor yumurtalarının çoğu kaybolur, ancak Macroolithus görünüşe göre sınırın her iki tarafında da var (bu, bazı dinozor türlerinin Tersiyerin başlarına kadar hayatta kaldığı anlamına gelir).[21][22] Bununla birlikte, sedimantolojik kanıtlar, bu fosillerin aslında Tersiyer kayalarına döküntü akışlarıyla yeniden işlendiğini göstermektedir.[24]
Nanxiong Havzası, ağırlıklı olarak oviraptorisaurian olmak üzere, bol miktarda fosil yumurtası ile bilinir.[1] yumurtalar Elongatoolithus ve Macroolithus.[21] Diğer yumurta türleri arasında diğer uzama taşı türleri ve ayrıca prizmaolitler, megaloolithids, ve ovaloolithids.[22] Ayak izleri, Nanxiong Havzası'nın ornitopodlar, theropodlar ve muhtemelen Sauropodlar.[38]
Güney Çin, özellikle Nanxiong Oluşumu içinde Jiangxi Eyaleti, muhtemelen dünyadaki en büyük oviraptorosaur çeşitliliğine sahiptir,[39] ve ayrıca oviraptorosaurların çeşitli birlikteliklerini içerir. Macroolithus veya benzeri yumurtalar.[19][15][14] Titanosaurlar ve tyrannosaurids bu bölgeden de bilinmektedir.[14] Dinozor dışı fauna kertenkeleleri ve karasal hayvanları içerir. Nanhsiungchelyid kaplumbağalar.[40]
Moğolistan
Macroolithus rugustus bol miktarda bulunur Nemegt Oluşumu geç tarihlenen Kampaniyen çok erken Maastrihtiyen.[41][42] Bu oluşum, kıvrımlı bir nehrin birikimlerini temsil eder.[42] Nemegt Formasyonunda iyi korunmuş dinozor kalıntıları yaygındır.[43] oviraptorosaurlar dahil,[42] titanosaurlar,[44] troodontids, tyrannosaurs, ankylosaurs,[45] pachycephalosaurs,[46] hadrosaurlar[47] ornitomimozorlar, alvarezsaurs,[48] ve Therizinosaurlar.[49] Küçük hayvan kalıntıları nispeten nadirdir, ancak Nemegt'ten birkaç tür kuş bilinmektedir,[48] yanı sıra birkaç tür çok tüberküloz memeliler.[50] Nemegt Formasyonundaki diğer fosil yumurtalar şunları içerir: Ovaloolithus, Spheroolithus, Elongatoolithus, ve Laevisoolithus.[41]
M. mutabilis sadece Ikh-Shunkht mevkiinde bilinmektedir. Barun Goyot Formasyonu, dan kalma Santoniyen -e Kampaniyen.[41][2] M. rugustus Barun Goyot Formasyonundan da bilinir, bu da kumul dolu bir kumulu temsil eder. eolian çevre. Nemegt Formasyonu ile karşılaştırıldığında, faunaya protoceratopsidler, oviraptoridler ve ankilosauridlerin hakim olduğu Barun Goyot'ta büyük dinozorlar nadirdir.[51] Barun Goyot Formasyonundaki diğer fosil yumurta türleri şunlardır: Protoceratopsidovum, Gobioolithus, Faveoloolithus, Dendroolithus, Spheroolithus, ve Subtiliolithus.[41]
Kazakistan
M. rugustus da bulundu Manrak Oluşumu (Manrakskaya Svita olarak da bilinir) Zaisan Havzası içinde Doğu Kazakistan Bölgesi. Bu oluşum yakın Tayzhuzgen Nehri ve muhtemelen Maastrictian olan geç Kretase'de bir zamana tarihlenir.[2][4][31][52]
Ayrıca bakınız
Referanslar
- ^ a b c d e f g h ben j k l m Simon, D.J. (2014). "Güneydoğu Idaho'dan Dev Dinozor (theropod) Oogenus Macroelongatoolithus (Elongatoolithidae) Yumurtaları: Taksonomik, Paleobiyocoğrafik ve Üreme Etkileri. "(Doktora tezi, Montana Eyalet Üniversitesi, Bozeman).
- ^ a b c d e f g h ben j k l Mihailov, Konstantin K.E. (1994). "Moğolistan ve Kazakistan'ın Kretase döneminden Theropod ve protoceratopsian dinozor yumurtaları". Paleontological Journal. 28 (2): 101–120.
- ^ a b c d e Genç, Chung-chien (1965). "Nanhsiung, Kwangtun ve Kanchou, Kiangsi'den yumurta fosili" (PDF). Vertebrata PalAsiatica. 9 (2): 141–170.
- ^ a b Mihailov, Konstantin (2000). "Moğolistan'ın Kretase'sinden dinozorların ve kuşların yumurtaları ve yumurta kabukları". Benton, M.J .; Shishkin, M.A .; Unwin, D.M .; Kurochkin, E.N. (eds.). Rusya ve Moğolistan'da Dinozorlar Çağı. Cambridge University Press. s. 560–572.
- ^ a b c d e f g h ben Zhao, Zikui (1975). "Guangdong Eyaleti, Nanxiong'daki dinozor yumurtası kabuklarının mikro yapısı - dinozor yumurtalarının sınıflandırılması hakkında" (PDF). Vertebrata PalAsiatica. 13 (2): 105–117.
- ^ Carpenter, K. (1999). Yumurtalar, Yuvalar ve Yavru Dinozorlar: Dinozorların Üremesine Bir Bakış. Indiana University Press. pp.294–295.
- ^ Carpenter, Kenneth (1999). "Fosil Yumurta Nasıl Çalışılır". Yumurtalar, Yuvalar ve Yavru Dinozorlar: Dinozorların Üremesine Bir Bakış (Geçmişin Hayatı). Bloomington, Indiana: Indiana University Press. pp.122–144. ISBN 978-0-253-33497-8.
- ^ a b Liang, Xinquan; Wan, Shunü; Yang, Dongsheng; Zhou, Shiquan; Wu, Shichong (2009). "Çin'in Henan Eyaletindeki dinozor yumurtaları ve dinozor yumurtası yatakları (Üst Kretase): Oluşumlar, paleoortamlar, tafonomi ve koruma". Doğa Bilimlerinde İlerleme. 19 (11): 1587–1601. doi:10.1016 / j.pnsc.2009.06.012.
- ^ a b c d Fang, Xiaosi; Li, Peixian; Zhang, Zhijun; Lin, Youli; Guo, Shengbin; Cheng, Yeming; Li, Zhenyu; Zhang, Xiaojun; Cheng, Zhengwu (2009). "广东 南雄 白垩 系 及 恐龙 蛋 到 鸟蛋 演化 研究" [Guangdong Nanxiong Havzasındaki Kretase Tabakaları ve Dinozor Yumurtasından Kuş Yumurtasına Evrim]. Acta Geoscientia Sinica (Çin'de). 30 (2): 167–186.
- ^ a b c d Wiemann, Jasmina; Yang, Tzu-Ruei; Sander, Philipp N .; Schneider, Marion; Engeser, Marianne; Kath-Schorr, Stephanie; Müller, Christa E .; Sander, Martin P. (2017). "Yumurta renginin dinozor kökeni: oviraptorlar mavi-yeşil yumurta bıraktı". PeerJ. 5 (e3706): e3706. doi:10.7717 / peerj.3706. PMC 5580385. PMID 28875070.
- ^ Banay, G.L. (1948). "Tıbbi terminolojiye giriş i. Yunanca ve Latin türevleri". Tıp Kütüphanesi Derneği Bülteni. 36 (1): 763–769. PMC 194697. PMID 16016791.
- ^ Mihaylov, Konstantin E .; Bray, Emily S .; Hirsch, Karl F. (1996). "Fosil yumurta kalıntılarının parataksonomisi (Veterovata): İlkeler ve uygulamalar". Omurgalı Paleontoloji Dergisi. 16 (4): 763–769. doi:10.1080/02724634.1996.10011364.
- ^ Beyaz, John Tahourdin (1858). Latince son ekler. Londra: Spottiswoode and Co. s.80.
ensis latin kökü.
- ^ a b c d e f g Wang, Shuo; Zhang, Shukang; Sullivan, Corwin; Xu, Xing (2016). "Güney Çin'in Üst Kretase'sinden oviraptorid (Theropoda, Oviraptorosauria) embriyoları içeren elongatoolithid yumurtaları". BMC Evrimsel Biyoloji. 16 (67): 67. doi:10.1186 / s12862-016-0633-0. PMC 4807547. PMID 27012653.
- ^ a b c d Sato, Tamaki; Cheng, Yen-nien; Wu, Xiao-chun; Zelenitsky, Darla; Hsiao, Yu-fu (2005). "Bir Dişi Dinozorun İçinde Bir Çift Kabuklu Yumurta". Bilim. 308 (5720): 375. doi:10.1126 / science.1110578. PMID 15831749.
- ^ Fanti, F; Currie, PJ; Badamgarav, D (2012). "Yeni Örnekler Nemegtomaia Moğolistan'ın Baruungoyot ve Nemegt Formasyonlarından (Geç Kretase) ". PLoS ONE. 7 (2): e31330. Bibcode:2012PLoSO ... 731330F. doi:10.1371 / journal.pone.0031330. PMC 3275628. PMID 22347465.
- ^ a b Varricchio, David J .; Moore, Jason R .; Erickson, Gregory M .; Norell, Mark A .; Jackson, Frankie D .; Borkowski, John J. (2008). "Avian Babalık Bakımı Dinozor Kökenlidir". Bilim. 322 (5909): 1826–1827. Bibcode:2008Sci ... 322.1826V. doi:10.1126 / science.1163245. PMID 19095938.
- ^ a b Tanaka, Kohei; Zelenitsky, Darla K .; Therrien François (2015). "Yumurta Kabuğu Gözenekliliği Dinozorlarda Yuvalanmanın Evrimi Hakkında Bilgi Sağlıyor". PLoS ONE. 10 (11): e0142829. Bibcode:2015PLoSO..1042829T. doi:10.1371 / journal.pone.0142829. PMC 4659668. PMID 26605799.
- ^ a b c d e Cheng, Yen-nien; Ji, Qiang; Wu, Xiao-chun; Shan, Hsi-y (2008). "Çin'de İlk Kez Keşfedilen Embriyonik İskeletli Oviraptorosaurian Yumurtaları (Dinosauria)". Acta Geologica Sinica. 82 (6): 1089–1094. doi:10.1111 / j.1755-6724.2008.tb00708.x.
- ^ a b c d Zhao, Zi-kui (1994). "Çin'de dinozor yumurtaları: Yumurta kabuklarının yapısı ve evrimi üzerine". Carpenter, Kenneth; Hirsch, Karl F .; Horner, John R. (editörler). Dinozor Yumurtaları ve Bebekler. Cambridge University Press. s. 184–203. ISBN 978-0-521-44342-5.
- ^ a b c d e Zhao, Zikui; Mao, Xueming; Chai, Zhifang; Yang, Gaochuang; Kong, Ping; Ebihara, Mitsuru; Zhao, Zhenhua (2002). "Güney Çin, Nanxiong Havzasında dinozorların neslinin tükenmesi ile K / T iridyum zenginleşmesi arasında olası bir nedensel ilişki: dinozor yumurta kabuklarından kanıtlar". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 178 (1–2): 1–17. Bibcode:2002PPP ... 178 .... 1Z. doi:10.1016 / S0031-0182 (01) 00361-3.
- ^ a b c d Zhao, Zikui; Mao, Xueying; Chai, Zhifang; Yang, Gaochuang; Zhang, Fucheng; Yan, Zheng (2009). "Güney Çin, Nanxiong Havzasında Kretase-Paleojen (K / T) geçişi sırasında jeokimyasal çevresel değişiklikler ve dinozor neslinin tükenmesi: Dinozor yumurta kabuklarından kanıtlar". Çin Bilim Bülteni. 54 (5): 806. doi:10.1007 / s11434-008-0565-1.
- ^ Wang, Qiang; Wang, Xiaolin; Zhao, Zikui (2010). "Çin'deki Dinozor Yumurtaları Çalışmasında Son Gelişmeler" (PDF). Çin Bilimler Akademisi Bülteni. 24 (2): 95–97. Arşivlenen orijinal (PDF) 2017-01-09 tarihinde. Alındı 2017-01-08.
- ^ a b c Buck, Brenda J .; Hanson, Andrew D .; Hengst, Richard A .; Hu, Shu-sheng (2004). """Güney Çin'deki Nanxiong Havzasındaki Üçüncül Dinozorlar Kretase'den Yeniden Çalışıyor". Jeoloji Dergisi. 112 (1): 111–118. Bibcode:2004JG .... 112..111B. doi:10.1086/379695.
- ^ Varricctio, David J .; Barta, Daniel E. (2015). "Gobi Kretase'den Şabat'ın" Daha Büyük Kuş Yumurtaları "nı Yeniden Canlandırmak. Acta Palaeontologica Polonica. 60 (1): 11–25. doi:10.4202 / app.00085.2014.
- ^ Garcia, Geraldine; Marivaux, Laurent; Pélissié, Thierry; Vianey-Liaud, Monique (2006). "En erken Laurisian sauropod yumurta kabuğu" (PDF). Acta Palaeontologica Polonica. 51 (1): 99–104.
- ^ a b Moreno-Azanza, M .; Canudo, J.I .; Gasca, J.M. (2014). "Avrupa'nın Alt Kretase bölgesindeki Sferolit yumurta kabukları. Ornitosiyan dinozorlarında yumurta kabuğu evrimi için çıkarımlar" (PDF). Kretase Araştırmaları. 51: 75–87. doi:10.1016 / j.cretres.2014.05.017.
- ^ Lawver, Daniel R .; Jackson, Frankie D. (2015). "Kaplumbağa Üremesinin Fosil Kayıtlarının İncelenmesi: Yumurtalar, Embriyolar, Yuvalar ve Çiftleşen Çiftler" (PDF). Peabody Doğa Tarihi Müzesi Bülteni. 55 (2): 215–236. doi:10.3374/014.055.0210.
- ^ Vianey-Liaud, Monique; Zelenitsky, Darla (2003). "Fosil yumurtaların parataksonomi üzerine tarihsel ve yeni perspektifler". Paleovertebrata. 32 (2–4): 189–195.
- ^ Mihaylov, Konstantin E. (1991). "Amniyotik Omurgalıların Fosil Yumurta Kabuklarının Sınıflandırılması" (PDF). Acta Palaeontologica Polonica. 36 (2): 193–238.
- ^ a b Mikhailov, K.E. (1997). "Amniyotik Omurgalılarda Fosil ve Yeni Yumurta Kabuğu: İnce Yapı, Karşılaştırmalı Morfoloji ve Sınıflandırma". Paleontolojide Özel Makaleler. 56: 64.
- ^ Zelenitsky, Darla K .; Carpenter, Kenneth; Currie, Philip J. (2000). "Elongatoolithus Theropod Yumurta Kabuğunun Kuzey Amerika'dan İlk Kaydı: Asya Oogenusu Macroelongatoolithus Utah'ın aşağı Kretase'sinden ". Omurgalı Paleontoloji Dergisi. 20 (1): 130–138. doi:10.1671 / 0272-4634 (2000) 020 [0130: FROETE] 2.0.CO; 2.
- ^ Mou, Yun (1992). "Guangdong Eyaleti, Nanxiong Havzasından Geç Kretase Dinozor yumurtalarının Yuva Ortamları" (PDF). Vertebrata PalAsiatica. 30 (2): 120–134.
- ^ Deeming, D.C. (2006). "Yumurta Kabuklarının Ultrastrüktürel ve Fonksiyonel Morfolojisi, Dinozor Yumurtalarının Bir Substratta İnkübe Edildiği Fikrini Destekler". Paleontoloji. 49 (1): 171–185. doi:10.1111 / j.1475-4983.2005.00536.x.
- ^ a b c Zhao, Zikui; Evet, Jie; Li, Huamei; Zhao, Zhenhua; Yan, Zheng (1991). "Guangdong Eyaleti, Nanxiong Havzasında Kretase-Tersiyer Sınırındaki dinozorların neslinin tükenmesi" (PDF). Açta Paleontologica Sinica. 29 (1): 1–20.
- ^ Zhao, Zikui (1979). "Dinozor Yumurtalarının Araştırılmasında İlerleme" (PDF). Güney Çin'in Mesozoik ve Senozoik Kırmızı Yatakları. Nanxiong, Guangdong Eyaleti, "Kretase-Tersiyer Çalıştayı" ndan Seçilmiş Makaleler: 330–340.
- ^ Wang, Qiang; Wang, Xiaolin; Zhao, Zikui; Zhang, Jialiang; Jiang, Shunxing (2013). "Çin'in Shandong Eyaleti, Laiyang Havzasının Üst Kretase'sinden yeni kaplumbağa yumurtası fosili". Anais da Academia Brasileira de Ciências. 85 (1): 103–111. doi:10.1590 / S0001-37652013000100008. PMID 23538955.
- ^ Xing, Lida; Harris, Jerald D; Dong, Zhiming; Lin, Youli; Chen, Wei; Guo, Shengbin; Ji, Qiang (2009). "Ornithopod (Dinosauria: Ornithischia), Nanxiong havzası, Guangdong, Çin'deki Üst Kretase Zhutian Formasyonundan izler ve büyük Çin ornitopu ayak izleri üzerine genel gözlemler" (PDF). Çin Jeoloji Bülteni. 28 (7): 829–843.
- ^ Lü, Junchang; Pu, Hanyong; Kobayashi, Yoshitsugu; Xu, Li; Chang, Huali; Shang, Yuhua; Liu, Di; Lee, Yuong-Nam; Kundrát, Martin; Shen, Caizhi (2015). "Güney Çin'in Geç Kretase Dönemi'nden Yeni Bir Oviraptorid Dinozoru (Dinosauria: Oviraptorosauria) ve Paleobiyocoğrafik Etkileri". Bilimsel Raporlar. 5: 11490. Bibcode:2015NatSR ... 511490L. doi:10.1038 / srep11490. PMC 4489096. PMID 26133245.
- ^ Tong, Haiyan; Mo, Jinyou (2010). "Jiangxichelys, Çin, Jiangxi Eyaleti, Ganzhou'nun Geç Kretase'sinden yeni bir nanhsiungchelyid kaplumbağa". Jeoloji Dergisi. 147 (6): 981–986. Bibcode:2010GeoM..147..981T. doi:10.1017 / S0016756810000671.
- ^ a b c d Mihailov, Konstantin (1995). "Moğolistan'daki Kretase Yumurtalarının Sistematik, Faunistik ve Stratigrafik Çeşitliliği: Çin ile Karşılaştırma". Altıncı Mezozoik Karasal Ekosistemler ve Biyota Sempozyumu, Kısa Makaleler: 165–168.
- ^ a b c Weishampel, David B .; Fastovsky, David E .; Watabe, Mahito; Varricchio, David; Jackson, Frankie; Tsogtbaatar, Khishigjav; Barsbold Rinchen (2008). "Bugin Tsav'dan Yeni Oviraptorid Embriyoları, Nemegt Formasyonu (Üst Kretase), Moğolistan, Yaşam Alanları ve Kalabalıkları Hakkında İçgörüler". Omurgalı Paleontoloji Dergisi. 28 (4): 1110–1119. doi:10.1671/0272-4634-28.4.1110.
- ^ Clarke, Julia A .; Norell, Mark A. (2004). "Yeni Avialan Kalıntıları ve Moğolistan'ın Geç Kretase Nemegt Formasyonundan Bilinen Avifauna'nın İncelenmesi". Amerikan Müzesi Novitates. 3447: 1–12. doi:10.1206 / 0003-0082 (2004) 447 <0001: NARAAR> 2.0.CO; 2. hdl:2246/2812.
- ^ Wilson, Jeffrey A. (2005). "Moğol Sauropod'un Yeniden Tanımı Nemegrosaurus mongoliensis Nowinski (Dinosauria: Saurischia) ve Geç Kretase Sauropod çeşitliliği hakkında yorumlar ". Sistematik Paleontoloji Dergisi. 3 (3): 283–318. doi:10.1017 / S1477201905001628.
- ^ Arbor, Victoria M .; Currie, Philip J .; Badamgarav, Demchig (2014). "Moğolistan'ın Üst Kretase Baruungoyot ve Nemegt oluşumlarının ankylosaurid dinozorları". Linnean Society'nin Zooloji Dergisi. 172 (3): 631–652. doi:10.1111 / zoj.12185.
- ^ Evans, David C .; Brown, Caled Marshall; Ryan, Michael J .; Tsogtbaatar, Khishigjav (2011). "Nemegt Formasyonundan Homalocephale calathocercos'un (Ornithischia: Pachycephalosauria) Kraniyal Süsleme ve Ontogenetik Durumu, Moğolistan". Omurgalı Paleontoloji Dergisi. 31 (1): 84–92. doi:10.1080/02724634.2011.546287.
- ^ Maryanska, Teresa; Osmolska, Halszka (1982). "Nemegt Formasyonundan Firth Lambeosaurine Dinozor, Üst Kretase, Moğolistan". Acta Palaeontologica Polonica. 26 (3/4): 243–255.
- ^ a b Clarke, Julia A .; Norrell, Mark A. (2004). "Yeni Avialan Kalıntıları ve Moğolistan'ın Geç Kretase Nemegt Formasyonundan Bilinen Avifauna'nın Bir İncelemesi" (PDF). Amerikan Müzesi Novitates. 3447: 1–12. doi:10.1206 / 0003-0082 (2004) 447 <0001: NARAAR> 2.0.CO; 2. hdl:2246/2812.
- ^ Bell, Phil R .; Currie, Philip J .; Lee, Yuong-Nam (2012). "Tyrannosaur izleri besliyor Deinocheirus (Theropod:? Ornithomimosauria) Nemegt Formasyonundan (Geç Kretase) kalıntıları, Moğolistan ". Kretase Araştırmaları. 37: 186–190. doi:10.1016 / j.cretres.2012.03.018.
- ^ Wible, John R .; Rougier Guillermo W. (2000). "Kafatası Anatomisi Kryptobaatar dashzevegi (Mammalia, Multituberculara) ve memeli karakterlerinin evrimi üzerindeki etkisi " (PDF). Amerikan Doğa Tarihi Müzesi Bülteni. 247: 6.
- ^ Longrich, Nicholas R .; Currie, Philip J .; Dong, Zhi-Ming (2010). "İç Moğolistan Yazarlar Bayan Mandahu'nun Üst Kretase'sinden yeni bir oviraptorid (Dinosauria: Theropoda)". Paleontoloji. 53 (5): 945–960. doi:10.1111 / j.1475-4983.2010.00968.x.
- ^ Averianov, Alexander O. (2007). "Kazakistan'ın kuzeydoğu Aral Denizi bölgesinde Geç Kretacoues yataklarından gelen Theropod dinozorları". Kretase Araştırmaları. 28 (3): 532–544. doi:10.1016 / j.cretres.2006.08.008.