Bitki morfolojisi - Plant morphology
Fitomorfoloji çalışmasıdır fiziksel form ve dış yapı nın-nin bitkiler.[1] Bu genellikle şundan farklı kabul edilir: bitki anatomisi,[1] hangi içsel çalışma yapı bitkilerin, özellikle mikroskobik düzeyde.[2] Bitki morfolojisi, bitkilerin görsel olarak tanımlanmasında yararlıdır. İçinde son çalışmalar moleküler Biyoloji bitki morfolojilerinin korunması ve çeşitlendirilmesinde yer alan moleküler süreçleri araştırmaya başladı. Bu çalışmalarda transkriptom koruma kalıplarının çok önemli olduğu bulundu ontogenetik sırasındaki geçişler bitki çeşitliliği sınırlayan evrimsel kısıtlamalara neden olabilecek yaşam döngüsü[3].

Dürbün


Bitki morfolojisi "bitkilerin gelişimi, biçimi ve yapısı üzerine bir çalışmayı ve dolaylı olarak bunları plan ve köken benzerliği temelinde yorumlama girişimini" temsil eder.[4] Bitki morfolojisinde dört ana araştırma alanı vardır ve her biri başka bir alanla çakışır. Biyolojik Bilimler.
Her şeyden önce morfoloji karşılaştırmalıBu, morfologun aynı veya farklı türden birçok farklı bitkideki yapıları incelediği, ardından benzerlikler hakkında karşılaştırmalar yaptığı ve fikirler geliştirdiği anlamına gelir. Farklı türlerdeki yapıların ortak, kalıtsal bir sonucu olarak var olduğuna ve geliştiğine inanılırsa genetik yollar, bu yapılar olarak adlandırılır homolog. Örneğin, yapraklar nın-nin çam, meşe, ve lahana hepsi çok farklı görünür, ancak belirli temel yapıları ve parçaların düzenini paylaşırlar. Yaprakların homolojisi, çıkarılması kolay bir sonuçtur. Bitki morfoloğu daha da ileri gider ve dikenler nın-nin kaktüs ayrıca diğer bitkilerdeki yapraklarla aynı temel yapı ve gelişmeyi paylaşır ve bu nedenle kaktüs dikenleri de yapraklara homologdur. Bitki morfolojisinin bu yönü bitki çalışmasıyla örtüşmektedir. evrim ve paleobotanik.
İkinci olarak, bitki morfolojisi, hem bitkisel (somatik) bitkilerin yapılarının yanı sıra üreme yapılar. Bitkisel yapılar damarlı Bitkiler şunlardan oluşan atış sistemi çalışmasını içerir kaynaklanıyor ve yapraklar yanı sıra kök sistemi. Üreme yapıları daha çeşitlidir ve genellikle belirli bir bitki grubuna özgüdür. Çiçekler ve tohumlar, eğreltiotu Sori, ve yosun kapsüller. Bitkilerdeki üreme yapılarının ayrıntılı incelenmesi, nesillerin değişimi tüm bitkilerde ve çoğu yosun. Bu bitki morfolojisi alanı, biyolojik çeşitlilik ve bitki sistematiği.
Üçüncüsü, bitki morfolojisi, bitki yapısını çeşitli ölçeklerde inceler. En küçük ölçeklerde üst yapı, hücrelerin genel yapısal özellikleri ancak bir elektron mikroskobu, ve sitoloji, kullanan hücrelerin incelenmesi Optik mikroskopi. Bu ölçekte, bitki morfolojisi ile örtüşür bitki anatomisi bir çalışma alanı olarak. En büyük ölçekte bitki çalışması büyüme alışkanlığı, bir bitkinin genel mimarisi. Bir dallanma modeli ağaç türden türe değişeceği gibi, bir bitkinin görünüşü de ağaç, ot veya çimen.
Dördüncü olarak, bitki morfolojisi şunların modelini inceler. gelişme, bitki büyüdükçe yapıların ortaya çıktığı ve olgunlaştığı süreç. Hayvanlar, hayatlarının erken dönemlerinden itibaren sahip olacakları tüm vücut parçalarını üretirken, bitkiler yaşamları boyunca sürekli olarak yeni dokular ve yapılar üretirler. Canlı bir bitkinin her zaman embriyonik dokuları vardır. Yeni yapıların üretildikçe olgunlaşma biçimleri, bitkinin yaşamında gelişmeye başladıkları noktadan ve yapıların maruz kaldığı ortamdan etkilenebilir. Bir morfolog bu süreci, nedenleri ve sonucunu inceler. Bu bitki morfolojisi alanı, bitki Fizyolojisi ve ekoloji.
Karşılaştırmalı bir bilim
Bir bitki morfoloğu, aynı veya farklı türden birçok farklı bitkideki yapılar arasında karşılaştırmalar yapar. Farklı bitkilerdeki benzer yapılar arasında bu tür karşılaştırmalar yapmak şu soruyu ele alır: neden yapılar benzer. Genetik, fizyoloji veya çevreye tepkinin altında yatan benzer nedenlerin görünüşte bu benzerliğe yol açması oldukça muhtemeldir. Bu nedenlere yönelik bilimsel araştırmanın sonucu, temel biyolojiye ilişkin iki kavrayıştan birine yol açabilir:
- Homoloji - yapı, ortak soy ve ortak genetik nedeniyle iki tür arasında benzerdir.
- Yakınsama - ortak çevresel baskılara bağımsız adaptasyon nedeniyle iki tür arasındaki yapı benzerdir.
Her türe hangi özelliklerin ve yapıların ait olduğunu anlamak, bitkiyi anlamanın önemli bir parçasıdır. evrim. Evrimsel biyolog, yapıları yorumlamak için bitki morfoloğuna güvenir ve sırayla filojenler yeni morfolojik anlayışlara yol açabilecek bitki ilişkileri.
Homoloji
Farklı türlerdeki yapıların, ortak, kalıtsal genetik yolların bir sonucu olarak var olduğuna ve geliştiğine inanılırsa, bu yapılar homolog. Örneğin, çam, meşe ve lahana yaprakları çok farklı görünür, ancak belirli temel yapıları ve parçaların düzenini paylaşır. Yaprakların homolojisi, çıkarılması kolay bir sonuçtur. Bitki morfoloğu daha da ileri giderek kaktüsün dikenlerinin diğer bitkilerdeki yapraklarla aynı temel yapı ve gelişmeyi paylaştığını ve bu nedenle kaktüs dikenlerinin de yapraklara homolog olduğunu keşfeder.
Yakınsama
Farklı türlerdeki yapıların, çevresel baskıya ortak adaptif tepkilerin bir sonucu olarak var olduğuna ve geliştiğine inanılırsa, bu yapılar yakınsak. Örneğin, fronds of Bryopsis plumosa ve kaynaklanıyor Kuşkonmaz setaceus biri alg ve diğeri çiçekli bitki olmasına rağmen ikisi de aynı tüylü dallanma görünümüne sahiptir. Genel yapıdaki benzerlik, yakınsamanın bir sonucu olarak bağımsız olarak ortaya çıkar. Çoğunun büyüme formu kaktüsler ve türleri Sütleğen çok uzak ailelere ait olmalarına rağmen çok benzer. Benzerlik, sıcak ve kuru bir ortamda hayatta kalma sorununa ortak çözümlerden kaynaklanmaktadır.
Bitkisel ve üreme özellikleri
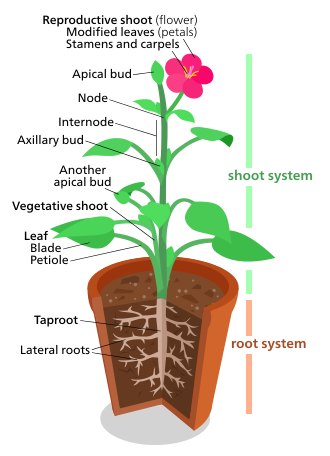
Bitki morfolojisi, hem bitkilerin bitkisel yapılarını hem de üreme yapılarını tedavi eder.
bitkisel (somatik) yapıları damarlı Bitkiler iki ana organ sistemini içerir: (1) a atış sistemi, oluşan kaynaklanıyor ve yapraklar ve (2) a kök sistem. Bu iki sistem neredeyse tüm vasküler bitkiler için ortaktır ve bitki morfolojisi çalışması için birleştirici bir tema sağlar.
Aksine, üreme yapılar çeşitlidir ve genellikle belirli bir bitki grubuna özgüdür. Gibi yapılar Çiçekler ve meyveler sadece içinde bulunur anjiyospermler; Sori sadece şurada bulunur eğrelti otları; ve tohum kozalakları sadece şurada bulunur iğne yapraklılar ve diğer açık tohumlular. Bu nedenle üreme karakterleri, bitkilerin sınıflandırılmasında bitkisel karakterlerden daha yararlı olarak kabul edilir.
Tanımlamada kullanın
Bitki biyologları, bitki taksonlarındaki farklılıkları veya benzerlikleri değerlendirmek için karşılaştırılabilen, ölçülebilen, sayılabilen ve tanımlanabilen bitkilerin morfolojik karakterlerini kullanırlar ve bu karakterleri bitki tanımlama, sınıflandırma ve açıklamalar için kullanırlar.
Açıklamalarda veya tanımlama için karakterler kullanıldığında bunlar denir tanı veya anahtar karakterler bu niteliksel ve niceliksel olabilir.
- Nicel karakterler, sayılabilen veya ölçülebilen morfolojik özelliklerdir, örneğin bir bitki türünün 10-12 mm genişliğinde çiçek yaprakları vardır.
- Niteliksel karakterler, yaprak şekli, çiçek rengi veya tüylenme gibi morfolojik özelliklerdir.
Her iki tür karakter de bitkilerin tanımlanmasında çok faydalı olabilir.
Nesillerin değişimi
Bitkilerdeki üreme yapılarının ayrıntılı incelenmesi, nesillerin değişimi, tüm bitkilerde ve çoğu yosun Alman botanikçi tarafından Wilhelm Hofmeister. Bu keşif, tüm bitkilerin yaşam döngüsünü anlamak için ortak bir temel sağladığından, tüm bitki morfolojisinde yapılan en önemli keşiflerden biridir.
Bitkilerde pigmentasyon
Bitkilerdeki pigmentlerin temel işlevi fotosentez yeşil pigmenti kullanan klorofil Mümkün olduğunca fazla ışık enerjisi yakalamaya yardımcı olan birkaç kırmızı ve sarı pigment ile birlikte. Pigmentler aynı zamanda dikkat çekmede önemli bir faktördür. haşarat cesaretlendirmek için çiçeklere tozlaşma.
Bitki pigmentleri, çeşitli farklı molekül türlerini içerir. porfirinler, karotenoidler, antosiyaninler ve Betalains. Tüm biyolojik pigmentler, belirli dalga boylarını seçici olarak emer. ışık süre yansıtan diğerleri. Absorbe edilen ışık bitki tarafından kimyasal reaksiyonlara güç sağlamak için kullanılabilirken, ışığın yansıyan dalga boyları pigmentin göze görüneceği rengi belirler.
Gelişimde morfoloji
Bitki gelişme bitki büyüdükçe yapıların ortaya çıktığı ve olgunlaştığı süreçtir. Bir konu çalışmadır bitki anatomisi ve bitki Fizyolojisi bitki morfolojisinin yanı sıra.
Bitkilerdeki gelişme süreci, temelde aşağıda görülenden farklıdır. omurgalı hayvanlar. Bir hayvan embriyo gelişmeye başlarsa, ömrü boyunca sahip olacağı tüm vücut parçalarını çok erken üretecektir. Hayvan doğduğunda (veya yumurtadan çıktığında), tüm vücut kısımlarına sahip olur ve bu noktadan itibaren yalnızca daha büyük ve daha olgunlaşır. Aksine, bitkiler yaşamları boyunca sürekli olarak yeni dokular ve yapılar üretirler. meristemler[5] organların uçlarında veya olgun dokular arasında bulunur. Böylece canlı bir bitki her zaman embriyonik dokulara sahiptir.
Bir fabrikada görülen organizasyonun özellikleri ortaya çıkan özellikler tek tek parçaların toplamından daha fazlasıdır. "Bu dokuların ve işlevlerin entegre bir çok hücreli organizma içinde bir araya getirilmesi, yalnızca ayrı parçaların ve işlemlerin özelliklerini değil, aynı zamanda ayrı parçaların incelenmesi temelinde tahmin edilemeyecek oldukça yeni bir özellikler dizisi de sağlar."[6] Yani bir bitkideki moleküller hakkında her şeyi bilmek, hücrelerin özelliklerini tahmin etmek için yeterli değildir; ve hücrelerin tüm özelliklerini bilmek, bir bitkinin yapısının tüm özelliklerini tahmin etmeyecektir.
Büyüme
Bir vasküler bitki tek hücreden başlar zigot, tarafından oluşturuldu döllenme bir yumurta hücresinin sperm hücresi tarafından O noktadan itibaren bir bitki oluşturmak için bölünmeye başlar embriyo süreci boyunca embriyojenez. Bu olurken, ortaya çıkan hücreler, bir ucu ilk kök, diğer ucu ise sürgünün ucunu oluşturacak şekilde organize olur. İçinde tohum bitkiler, embriyo bir veya daha fazla "tohum yaprağı" geliştirecektir (tohumdan çıkan ilk yaprak ). Embriyojenezin sonunda genç bitki, yaşamına başlamak için gerekli tüm parçalara sahip olacaktır.
Bir kez embriyo filizlenir tohumundan veya ana bitkisinden, işlem yoluyla ek organlar (yapraklar, saplar ve kökler) üretmeye başlar. organogenez. Yeni kökler kökten büyür meristemler kökün ucunda bulunur ve sürgünden yeni saplar ve yapraklar büyür meristemler çekimin ucunda.[7] Dallanma, meristem tarafından geride bırakılan ve henüz geçirilmemiş küçük hücre kümeleri olduğunda meydana gelir. hücresel farklılaşma özel bir doku oluşturmak için yeni bir kök veya sürgünün ucu olarak büyümeye başlar. Bir kökün veya sürgünün ucundaki bu tür herhangi bir meristemden büyümeye denir. birincil büyüme ve bu kökün veya sürgünün uzamasına neden olur. İkincil büyüme bir kök veya sürgünün hücre bölünmelerinden genişlemesine neden olur. kambiyum.[8]
Tarafından büyümeye ek olarak hücre bölünme, bir bitki büyüyebilir hücre uzaması. Bu, tek tek hücreler veya hücre grupları daha uzun büyüdüğünde ortaya çıkar. Tüm bitki hücreleri aynı uzunlukta büyümeyecektir. Bir kökün bir tarafındaki hücreler diğer taraftaki hücrelerden daha uzun ve daha hızlı büyüdüğünde, kök, sonuç olarak daha yavaş büyüyen hücrelerin tarafına doğru bükülür. Bu yönlü büyüme, bir bitkinin ışık gibi belirli bir uyarıcıya tepkisi yoluyla gerçekleşebilir (fototropizm ), Yerçekimi (yerçekimi ), Su, (hidrotropizm ) ve fiziksel temas (tigmotropizm ).
Bitki büyümesi ve gelişimi, belirli bitki hormonları ve bitki büyüme düzenleyicileri (PGR'ler) (Ross ve diğerleri, 1983).[9] Endojen hormon seviyeleri, bitki yaşı, soğuğa dayanıklılık, uyku hali ve diğer metabolik koşullardan etkilenir; fotoperiyot, kuraklık, sıcaklık ve diğer dış çevre koşulları; ve örneğin harici olarak uygulanan ve rizosfer kaynaklı PGR'lerin eksojen kaynakları.
Morfolojik varyasyon
Bitkiler, şekil ve yapılarında doğal çeşitlilik gösterirler. Tüm organizmalar kişiden kişiye değişirken, bitkiler ek bir çeşitlilik sergiler. Tek bir birey içinde, diğer benzer parçalardan şekil ve yapı bakımından farklılık gösterebilen parçalar tekrarlanır. Bu çeşitlilik en kolay şekilde bir bitkinin yapraklarında görülür, ancak gövdeler ve çiçekler gibi diğer organlar da benzer çeşitlilik gösterebilir. Bu varyasyonun üç temel nedeni vardır: konumsal etkiler, çevresel etkiler ve gençlik.
Bitki morfolojisinin evrimi
Transkripsiyon faktörleri ve transkripsiyonel düzenleyici ağlar, bitki morfogenezinde ve bunların evriminde anahtar rol oynar. Bitki inişi sırasında, birçok yeni transkripsiyon faktör ailesi ortaya çıktı ve tercihen çok hücreli gelişim, üreme ve organ gelişimi ağlarına bağlanarak kara bitkilerinin daha karmaşık morfogenezine katkıda bulundu.[10]
Konumsal etkiler

Bitkiler yaşamları boyunca aynı organın çok sayıda kopyasını üretmelerine rağmen, belirli bir organın tüm kopyaları aynı olmayacaktır. Olgun bir bitkinin parçaları arasında, organın üretildiği göreceli konumdan kaynaklanan farklılıklar vardır. Örneğin, yeni bir dal boyunca yapraklar, dal boyunca tutarlı bir modelde değişebilir. Dalın tabanının yakınında üretilen yaprakların şekli, bitkinin ucunda üretilen yapraklardan farklı olacaktır ve bu farklılık, belirli bir bitkide ve belirli bir türde daldan dala tutarlıdır. Bu farklılık dalın her iki ucundaki yapraklar olgunlaştıktan sonra da devam eder ve bazı yaprakların diğerlerinden daha genç olmasının bir sonucu değildir.
Çevresel etkiler
Yeni yapıların üretildikçe olgunlaşma biçimleri, bitkilerin yaşamında gelişmeye başladıkları noktadan ve yapıların maruz kaldığı ortamdan etkilenebilir. Bu görülebilir su bitkileri ve ortaya çıkan bitkiler.
Sıcaklık
Sıcaklığın bitkiler üzerinde, bitkinin boyutu ve durumu ve maruziyetin sıcaklığı ve süresi gibi çeşitli faktörlere bağlı olarak çok sayıda etkisi vardır. Daha küçük ve daha fazlası bitki etli, çok yüksek veya çok düşük sıcaklıklardan kaynaklanan hasar veya ölüme yatkınlık o kadar yüksek olur. Sıcaklık, biyokimyasal ve fizyolojik süreçlerin oranını etkiler, hızlar genellikle (sınırlar dahilinde) sıcaklıkla artar. Bununla birlikte, monomoleküler reaksiyonlar için Van't Hoff ilişkisi (bir reaksiyonun hızının 10 ° C'lik bir sıcaklık artışıyla iki veya üç katına çıktığını belirtir), özellikle düşük ve yüksek sıcaklıklarda biyolojik süreçler için kesin olarak geçerli değildir.
Bitkilerde su donduğunda, bitki için sonuçlar, donmanın hücre içinde mi (hücrelerin içinde) yoksa hücreler arası (hücre dışı) boşluklarda hücrelerin dışında mı oluştuğuna bağlıdır.[11] Hücre içi donma, genellikle bitkinin ve dokularının dayanıklılığına bakılmaksızın hücreyi öldürür.[12] Hücre içi donma doğada nadiren meydana gelir, ancak sıcaklıkta orta dereceli düşüşler, örneğin 1 ° C ila 6 ° C / saat, hücreler arası buzun oluşmasına neden olur ve bu "ekstraorganik buz"[13] dokunun sertliğine bağlı olarak ölümcül olabilir veya olmayabilir.
Dondurucu sıcaklıklarda, bitki dokularının hücreler arası boşluklarındaki su önce donar, ancak su, sıcaklıklar 7 ° C'nin altına düşene kadar donmadan kalabilir.[11] Hücreler arası olarak ilk buz oluşumundan sonra, su ayrılmış buzda kaybolurken hücreler küçülür. Hücreler dondurularak kurutulur, dehidrasyon donma hasarının temel nedenidir.
Soğutma hızının dokuların dona karşı direncini etkilediği gösterilmiştir,[14] ancak gerçek donma hızı, yalnızca soğutma hızına değil, aynı zamanda aşırı soğutma derecesine ve dokunun özelliklerine de bağlı olacaktır.[15] Sakai (1979a)[14] Yavaş yavaş 30 ° C ila -40 ° C'ye soğutulduğunda Alaska beyaz ve siyah ladinlerin sürgün primordia'larında buz ayrışması gösterdi. Bu dondurularak kurutulmuş tomurcuklar, sıvı nitrojen yavaşça yeniden ısıtıldığında. Floral primordia da benzer şekilde yanıt verdi. Primordia'daki ekstraorgan donması, kuzey kozalaklı ağaçların en sertinin, hava sıcaklıklarının genellikle -50 ° C veya altına düştüğü bölgelerde kış aylarında hayatta kalma yeteneğini açıklar.[13] Böyle kış tomurcuklarının sertliği iğne yapraklılar tomurcukların küçüklüğü, suyun daha hızlı yer değiştirmesinin evrimi ve yoğun donma dehidrasyonunu tolere etme yeteneği ile güçlendirilmiştir. Kuzey türlerinde Picea ve Pinus1 yaşındaki fidelerin dona dayanıklılığı olgun bitkilerle aynı seviyededir,[16] benzer uyku hali durumları göz önüne alındığında.
Çocukluk

Genç bir bitki tarafından üretilen organ ve dokular fide, genellikle aynı bitkinin büyüdüğünde ürettiğinden farklıdır. Bu fenomen olarak bilinir çocukluk veya heteroblasti. Örneğin genç ağaçlar, tamamen büyümüş bir ağaç olarak üretecekleri dallardan daha fazla yukarı doğru büyüyen daha uzun, daha zayıf dallar üreteceklerdir. Ek olarak, erken büyüme sırasında üretilen yapraklar, yetişkin bitkideki yapraklardan daha büyük, daha ince ve daha düzensiz olma eğilimindedir. Yavru bitki örnekleri, aynı türün yetişkin bitkilerinden o kadar farklı görünebilir ki, yumurtlayan böcekler bitkiyi yavruları için besin olarak tanımaz. Köklenebilirlik ve çiçeklenmede farklılıklar görülür ve aynı olgun ağaçta görülebilir. Bir ağacın tabanından alınan genç kesimler, orta tepeden üst tepeye kadar olan kesimlere göre çok daha kolay kökler oluşturacaktır. Bir ağacın tabanına yakın çiçeklenme, özellikle genç bir ağaç çiçeklenme yaşına ilk ulaştığında, yüksek dallarda çiçeklenmeden daha az ya da yoktur.[17]
Erken büyüme formlarından geç büyüme formlarına geçiş, 'bitkisel faz değişimi ', ancak terminoloji konusunda bazı anlaşmazlıklar var.[18]
Modern bitki morfolojisi
Rolf Sattler kavramı gibi karşılaştırmalı morfolojinin temel kavramlarını revize etmiştir. homoloji. Homolojinin kısmi homoloji ve niceliksel homolojiyi de içermesi gerektiğini vurguladı.[19][20] Bu, kök, sürgün, gövde (kakulom), yaprak (filom) ve saç (trikom) morfolojik kategorileri arasında bir süreklilik gösteren sürekli bir morfolojiye yol açar. Kategoriler arasındaki ara maddelerin en iyi nasıl açıklandığı Bruce K. Kirchoff ve diğerleri tarafından tartışılmıştır.[21] Stalk Enstitüsü tarafından yapılan yeni bir çalışma, her bitkinin tabanına karşılık gelen koordinatları çıkardı ve 3B alanda bıraktı. Grafikteki bitkiler, gerçek besin hareket mesafelerine ve toplam dal uzunluklarına göre yerleştirildiğinde, bitkiler Pareto eğrisinde neredeyse mükemmel bir şekilde düştü. "Bu, bitkilerin mimarilerini büyütme biçiminin aynı zamanda çok yaygın bir ağ tasarımı ödünleşimini de optimize ettiği anlamına geliyor. Çevre ve türlere bağlı olarak, bitki bu belirli çevresel koşullar için ödün vermek için farklı yollar seçiyor." [22]
Yazarı Agnes Arber, yaprağın kısmi atış teorisi, Rutishauser ve Isler süreklilik yaklaşımı Bulanık Arberya Morfolojisi (FAM) olarak adlandırdı. "Bulanık", Bulanık mantık, "Arberya" dan Agnes Arber. Rutishauser ve Isler, bu yaklaşımın yalnızca birçok morfolojik veri ile değil, aynı zamanda moleküler genetik kanıtlarla da desteklendiğini vurguladı.[23] Moleküler genetikten daha yeni kanıtlar, süreklilik morfolojisi için daha fazla destek sağlar. James (2009) şu sonuca varmıştır: "Radyalite [çoğu sapın özelliği] ve dorsiventralite [yaprakların karakteristiği] sürekli bir spektrumun uçlarıdır. Aslında, bu sadece KNOX geninin zamanlamasıdır. ifade !. "[24] Eckardt ve Baum (2010), "artık bileşik yaprakların hem yaprak hem de sürgün özelliklerini ifade ettiği genel olarak kabul edilmektedir" sonucuna varmıştır.[25]
Süreç morfolojisi, bitki formunun dinamik sürekliliğini tanımlar ve analiz eder. Bu yaklaşıma göre yapılar, Sahip olmak süreç (ler), onlar vardır süreç (ler).[26][27][28] Böylelikle yapı / süreç ikilemi, canlı organizmada bunun yalnızca bir uzamsal yapı meselesi olmadığını ve bunun üzerinde veya aleyhinde bir şey olarak bir 'etkinlik' ile bir uzamsal yapı sorunu olmadığını kabul etmek için 'yapı' kavramımızın genişlemesi ile aşılır. ama somut organizma bir mekansalgeçici yapı ve bu uzamsal-zamansal yapının faaliyetin kendisidir ".[29]
Jeune, Barabé ve Lacroix için, klasik morfoloji (yani, birbirini dışlayan kategorileri ima eden nitel homoloji kavramına dayanan ana akım morfoloji) ve sürekli morfoloji, daha kapsamlı süreç morfolojisinin (dinamik morfoloji) alt sınıflarıdır.[30]
Klasik morfoloji, süreklilik morfolojisi ve süreç morfolojisi, özellikle bitki morfolojisi ile bitki moleküler genetiğini bütünleştirmeye çalışan bitki evrimsel biyolojisi (bitki evrimi) alanı olmak üzere bitki evrimi ile oldukça ilgilidir.[31] Rutishauser (2016), alışılmadık morfolojiler üzerine ayrıntılı bir vaka çalışmasında, morfolojik kavramların belirsizliği (sürekliliği), yapısal kategoriler ve gen ifadesi arasında bire bir yazışmanın olmaması gibi bitki evriminin çeşitli konularını açıkladı ve tartıştı. , morfospace kavramı, bauplan özelliklerinin veranda ludens'e karşı uyarlanabilir değeri, fizyolojik adaptasyonlar, umut dolu canavarlar ve tuzlu evrim, gelişimsel sağlamlığın önemi ve sınırları, vb.[32]
Hoşumuza gitsin ya da gitmesin, morfolojik araştırma ya / veya mantık, bulanık mantık, yapı / süreç düalizmi ya da onun aşkınlığı gibi felsefi varsayımlardan etkilenir. Ve ampirik bulgular felsefi varsayımları etkileyebilir. Dolayısıyla felsefe ve deneysel bulgular arasında etkileşimler vardır. Bu etkileşimler, bitki morfolojisi felsefesi olarak adlandırılan şeyin konusudur.[33]
Ayrıca bakınız
- Bitki morfolojisi sözlüğü
- Bitki anatomisi
- Bitki tanımlama
- Bitki Fizyolojisi
- Bitki evrimsel gelişim biyolojisi
- Taksonomi
Referanslar
- ^ a b Raven, P. H., R. F. Evert ve S. E. Eichhorn. Bitki Biyolojisi, 7. baskı, sayfa 9. (New York: W.H. Freeman, 2005). ISBN 0-7167-1007-2.
- ^ Evert, Ray Franklin ve Esau, Katherine (2006) Esau'nun Bitki Anatomisi: Meristemler, hücreler ve bitki gövdesinin dokuları - yapıları, işlevleri ve gelişimi Wiley, Hoboken, New Jersey, sayfa xv, ISBN 0-471-73843-3
- ^ Drost, Hajk-Georg; Bellstaedt, Julia; Ó'Maoiléidigh, Diarmuid S .; Silva, Anderson T .; Gabel, İskender; Weinholdt, Claus; Ryan, Patrick T .; Dekkers, Bas J.W .; Bentsink, Leónie; Hilhorst, Henk W.M .; Ligterink, Wilco; Wellmer, Frank; Grosse, Ivo; Quint, Marcel (2016-02-23). "Post-embriyonik Kum Saati Modelleri Bitki Gelişiminde Ontogenetik Geçişlere İşaret Ediyor". Moleküler Biyoloji ve Evrim. 33 (5): 1158–1163. doi:10.1093 / molbev / msw039. PMID 26912813.
- ^ Harold C. Bold, C. J. Alexopoulos ve T. Delevoryas. Bitki ve Mantar Morfolojisi, 5. baskı, sayfa 3. (New York: Harper-Collins, 1987). ISBN 0-06-040839-1.
- ^ Bäurle, I; Laux, T (2003). "Apikal meristemler: Bitkinin gençlik pınarı". BioEssays. 25 (10): 961–70. doi:10.1002 / bies.10341. PMID 14505363. Gözden geçirmek.
- ^ Leopold, A. C. Bitki Büyümesi ve Gelişimi, sayfa 183. (New York: McGraw-Hill, 1964).
- ^ Marka, U; Hobe, M; Simon, R (2001). "Bitki sürgün meristemlerindeki işlevsel alanlar". BioEssays. 23 (2): 134–41. doi:10.1002 / 1521-1878 (200102) 23: 2 <134 :: AID-BIES1020> 3.0.CO; 2-3. PMID 11169586. Gözden geçirmek.
- ^ Barlow, P (2005). "Bir bitki dokusunda desenli hücre tayini: Ağaçların ikincil floem". BioEssays. 27 (5): 533–41. doi:10.1002 / bies.20214. PMID 15832381.
- ^ Ross, S.D .; Pharis, R.P .; Binder, W.D. 1983. Büyüme düzenleyicileri ve iğne yapraklılar: fizyolojileri ve ormancılıkta potansiyel kullanımları. s. 35–78 içinde Nickell, L.G. (Ed.), Bitki büyümesini düzenleyen kimyasallar. Cilt 2, CRC Press, Boca Raton FL.
- ^ Jin JP; et al. (Temmuz 2015). "Bir Arabidopsis transkripsiyonel düzenleyici harita, yeni transkripsiyon faktörlerinin farklı fonksiyonel ve evrimsel özelliklerini ortaya koymaktadır". Moleküler Biyoloji ve Evrim. 32 (7): 1767–1773. doi:10.1093 / molbev / msv058. PMC 4476157. PMID 25750178.
- ^ a b Glerum, C. 1985. İğne yapraklı fidelerin dona dayanıklılığı: ilkeler ve uygulamalar. s. 107–123 içinde Duryea, M.L. (Ed.). Bildiriler: Fide kalitesinin değerlendirilmesi: büyük testlerin ilkeleri, prosedürleri ve tahmin yetenekleri. Çalıştay, Ekim 1984, Oregon Eyalet Üniv., For. Res. Lab., Corvallis OR.
- ^ Lyons, J.M .; Raison, J.K .; Steponkus, P.L. 1979. Düşük sıcaklığa tepki olarak bitki zarı: genel bir bakış. s. 1–24 içinde Lyons, J.M .; Graham, D .; Raison, J.K. (Eds.). Bitkisel Bitkilerde Düşük Sıcaklık Gerilimi. Academic Press, New York NY.
- ^ a b Sakai, A .; Larcher, W. (Eds.) 1987. Frost Survival of Plants. Springer-Verlag.
- ^ a b Sakai, A. 1979a. Kozalaklı tomurcukların ilkel sürgünlerinin donmadan kaçınma mekanizması. Plant Cell Physiol. 20: 1381–1390.
- ^ Levitt, J. 1980. Bitkilerin Çevresel Streslere Tepkileri. Cilt 1. Soğutma, Dondurma ve Yüksek Sıcaklık Gerilmeleri, 2. baskı. Academic Press, New York NY. 497 s.
- ^ Sakai, A .; Okada, S. 1971. Kozalaklı ağaçların donma direnci. Silvae Genet. 20 (3): 91–97.
- ^ Michael A Dirr; Charles W Heuser, jr. (2006). "2". Odunsu Bitki Yayılımının Referans El Kitabı (İkinci baskı). Varsity Press Inc. s. 26, 28, 29. ISBN 0942375092.
- ^ Jones, Cynthia S. (1999-11-01). "Tohum Bitkilerde Gençlik, Faz Değişimi ve Heteroblasti Üzerine Bir Deneme". Uluslararası Bitki Bilimleri Dergisi. 160 (S6): –105 – S111. doi:10.1086/314215. ISSN 1058-5893. PMID 10572025.
- ^ Sattler, R. (1984). "Homoloji - devam eden bir meydan okuma". Sistematik Botanik. 9 (4): 382–394. doi:10.2307/2418787. JSTOR 2418787.
- ^ Sattler, R., 1994, Bitkilerde homoloji, homeosis ve süreç morfolojisi. İçinde: B.K. Hall (ed.) Kökendeşlik: Karşılaştırmalı morfolojinin hiyerarşik temeli. New York: Academic Press, s. 423-475.
- ^ Kirchoff, BK; Pfeifer, E; Rutishauser, R (2008). "Bitki yapısı ontolojisi: Bitki yapılarını şüpheli veya karışık kimliklerle nasıl etiketlemeliyiz?". Zootaxa. 1950: 103–122. doi:10.11646 / zootaxa.1950.1.10.
- ^ Conn, Adam; Pedmale, Ullas; Chory, Joanne (2017). "Yüksek Çözünürlüklü Lazer Tarama, Evrensel Ağ Tasarım İlkelerini Yansıtan Tesis Mimarilerini Ortaya Çıkarıyor". Hücre Sistemleri. 5 (1): 103–122. doi:10.1016 / j.cels.2017.06.017. PMID 28750198.
- ^ Rutishauser, R .; İşler, B. (2001). "Çiçekli Bitkilerin, Özellikle Mesane Sularının (Utricularia) Gelişimsel Genetiği ve Morfolojik Evrimi: Bulanık Arberya Morfolojisi Klasik Morfolojiyi Tamamlar" (PDF). Botanik Yıllıkları. 88 (6): 1173–1202. doi:10.1006 / anbo.2001.1498.
- ^ James, P.J. (2009). "'Tree and Leaf ': Farklı bir açı ". Linnean. 25: 13–19.
- ^ Eckardt, NA; Baum, D (2010). "Podostemad Bulmacası: Podostemaceae'deki Olağandışı Morfolojinin Evrimi". Bitki Hücresi. 22 (7): 2131–2140. doi:10.1105 / tpc.110.220711. PMC 2929115. PMID 20647343.
- ^ Sattler, R. (1992). "Süreç morfolojisi: Gelişim ve evrimde yapısal dinamikler". Kanada Botanik Dergisi. 70 (4): 708–714. doi:10.1139 / b92-091.
- ^ Vergara-Silva, F. (2003). "Bitkiler ve Evrimsel Gelişim Biyolojisinin Kavramsal Eklemlenmesi". Biyoloji ve Felsefe. 18 (2): 261–264. doi:10.1023 / A: 1023936102602. S2CID 81013686.
- ^ Sattler, R. 2019. Bitki formunun gelişimi ve evrimine yapısal ve dinamik yaklaşımlar. İçinde: Fusco, G. (ed) Evrimsel ve Gelişimsel Biyoloji Üzerine Perspektifler. Alessandro Minelli için Denemeler. Bölüm 6, sayfa 57-70 [1]
- ^ Woodger, J.H. 1967. Biyolojik Prensipler. Londra: Routledge & Kegoan Paul (yeni bir Giriş ile yeniden yayınlandı).
- ^ Jeune, B; Barabé, D; Lacroix, C (2006). "Klasik ve dinamik morfoloji: Formlar uzayı aracılığıyla bir senteze doğru". Acta Biotheoretica. 54 (4): 277–293. doi:10.1007 / s10441-007-9007-8. PMID 17486414. S2CID 25928998.
- ^ Minelli, A. 2018. Bitki Evrimsel Gelişim Biyolojisi. Fenotipin Evrilebilirliği. New York: Cambridge University Press.
- ^ Rutishauser, R. (2016). "Lentibulariaceae (mesane suları ve müttefikleri) ve Podostemonaceae'de (nehir otları) alışılmadık morfolojilerin evrimi: gelişimsel biyoloji ve morfolojik çeşitlilik arayüzünde resimli bir rapor". Botanik Yıllıkları. 117 (5): 811–832. doi:10.1093 / aob / mcv172. PMC 4845801. PMID 26589968.
- ^ Sattler, R. (2018). "Bitki morfolojisi felsefesi". Elemente der Naturwissenschaft. 108: 55–79.(bu makalenin genişletilmiş versiyonu için bkz. [2] )
Dış bağlantılar
| Alanlar |  | |
|---|---|---|
| Bakteri | ||
| Protistler |
| |
| Bitkiler | ||
| Omurgasızlar | ||
| Memeliler | ||
| Diğer omurgalılar | ||
| Diğer başlıklar | ||
| ||
| Alt disiplinler | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bitki grupları | |||||||||||
| |||||||||||
| |||||||||||
| Bitki büyümesi ve alışkanlığı | |||||||||||
| Üreme | |||||||||||
| Bitki taksonomisi | |||||||||||
| Uygulama | |||||||||||
| |||||||||||
| |||||||||||