Haplogrup D-M174 - Haplogroup D-M174
| Haplogrup D-M174 | |
|---|---|
| Muhtemel menşe zamanı | 50,000[1] - 60,000[2] yıl BP 65.200 [% 95 CI 62.100 <-> 68.300] ybp[3] |
| Birleşme yaşı | 46.300 [% 95 CI 43.500 <-> 49.100] ybp[3] |
| Olası menşe yeri | Asya[2][4][5] (muhtemelen Orta Asya[6] veya Güneydoğu Asya[2]) |
| Ata | D (D-CTS3946) |
| Torunları | D-Z27276 (D1a1) D-M55 (D1a2a) D-Y34637 (D1a2b) |
| Mutasyonları tanımlama | M174, IMS-JST021355, SAYFALAR00003 |
Haplogrup D1 veya D-M174 alt sınıfı Haplogrup D-CTS3946. Bu haplogrup öncelikle şurada bulunur: Doğu Asya ve Andaman Adaları aynı zamanda düşük sıklıkta düzenli olarak bulunmasına rağmen Orta Asya ve Güneydoğu Asya.
Kökenler
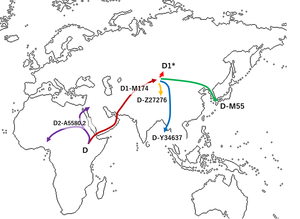
Haplogrup D-M174'ün ortaya çıktığına inanılıyor. Asya Günümüzden yaklaşık 60.000 yıl önce, Shi ve arkadaşları tarafından önerilmiştir. (2008), Kuzey ve Orta Asya'da nadir olması nedeniyle Güneydoğu Asya (Kuzey veya Orta Asya yerine) kökenlidir.[2][4] Haplogroup D-M174 ile birlikte haplogrup E ayırt edici YAP polimorfizm (ortak atalarını gösterir), Asya dışında hiçbir haplogrup D-M174 kromozomu bulunmamıştır.[4]
Birkaç çalışma (Hammer ve diğerleri 2006, Shinoda 2008, Matsumoto 2009, Cabrera ve diğerleri 2018), baba haplogrup D-M174'ün Orta Asya. Hammer ve diğerlerine göre, haplogroup D-M174, Tibet ve Altay dağları arasında ortaya çıktı. Doğu Avrasya'da çok sayıda dalga olduğunu öne sürüyor.[7][neden? ]
Mondal ve diğerleri tarafından 2017 yılında yapılan bir çalışma. bulur Riang insanları (bir Tibeto-Birmanya nüfusu) ve Andamanlıların en yakın akraba soyları Doğu Asya'da bulunmaktadır. Jarawa ve Onge, bu D1a2b soylarını son ~ 7000 yıl içinde birbirleriyle paylaştılar, ancak ~ 53000 yıl önce Japon D1a2a soyundan ayrıldılar. "Ayrıca şunu da öne sürüyorlar:" Bu, haplogrup D'nin ayrı bir ataya işaret etmediğini gösteriyor. Daha ziyade, haplogrup D, OOA genişlemesinin taşıdığı ayakta varyasyonun bir parçasıydı ve daha sonra Andaman dışında ve kısmen Japonya ve Tibet'te popülasyonların çoğunda kayboldu ”.[8]
Bir çalışma (Haber ve diğerleri 2019), üç Nijeryalı numunede "D0" adlı önerilen bir haplogrup buldu. Yazarlar, kısmen haplogroup "D0" ın olası derin köklenmesinin yanı sıra kendisi ve ana haplogroup için yakın zamanda hesaplanan erken sapma süreleri nedeniyle, D0 ve ana haplogroup DE için bir Afrika kökenini öneriyorlar. "D0" haplogrubu M174'ün dışında, ancak E'nin sahip olmadığı 7 SNP paylaşıyor.[9] "D0" aynı zamanda "D2" olarak adlandırıldı ve eski D (D-M174) artık bu keşif nedeniyle "D1" olarak da adlandırıldı.
Diğer üç D2 örneği Batı Asya'da da bulundu (yine 2019'da): ikisi Suudi Arabistan'da ve diğeri Suriye'de. Suriye'de bulunan örnek, bugüne kadar D2'nin en bazal örneğidir. Son kanıtlar (Haber ve diğerleri tarafından da önerildiği gibi), D2'nin DE bölünmesine yakın ancak D dalında oldukça farklı bir haplogrup olduğunu ve diğer D soylarının sahip olduğu M174 mutasyonundan yoksun olduğunu göstermektedir.[10]
Genel Bakış
Günümüzde popülasyonlar arasında yüksek sıklıkta bulunur. Tibet, kuzey Myanmar, Qinghai, Japon takımadaları, ve Andaman Adaları merakla geri kalanında olduğu kadar olmasa da Hindistan. Ainu Japonya ve çeşitli Tibeto-Birmanya insanlar (örneğin Tripuri insanlar ) neredeyse sadece Haplogroup D-M174 kromozomlarına sahip olduğu için dikkat çekicidir. Haplogrup D-M174 kromozomları ayrıca düşük ila orta frekanslarda bulunur. Bai, Dai, Han, Hui, Mançu, Miao, Tujia, Xibe, Yao, ve Zhuang nın-nin Çin ve birkaç azınlık nüfusu arasında Siçuan ve Yunnan bu konuşma Tibeto-Burman dilleri ve Tibetlilere yakın bir yerde ikamet ediyorlar. Jingpo, Jino, Mosuo, Naxi, Pumi, Qiang, ve Yi.[11]
Haplogrup D aynı zamanda Çin ve Kore popülasyonlarında da bulunur, ancak Tibet ve Japonya popülasyonlarına göre çok daha düşük sıklıkta. 2011'de yayınlanan bir çalışmada, Han Çinli erkeklerinin% 2.49'unda (43/1729) D-M174 bulundu ve bu haplogrubun sıklıkları, ülkenin kuzeyine ve batısına doğru ortalamanın üzerinde olma eğiliminde (5/56 =% 8.9 D-M174 Shaanxi Han, 13/221 =% 5,9 D-M174 Gansu Han, 6/136 =% 4,4 D-M174 Yunnan Han, 1/27 =% 3,7 D-M174 Guangxi Han, 2/61 =% 3,3 D- M174 Hunan Han, 2/62 =% 3.2 D-M174 Sichuan Han).[12] Han Çinli Y-DNA'nın 2011'de yayınlanan başka bir çalışmasında, Haplogroup D-M174, Şangay'daki Fudan Üniversitesi'ndeki ilgisiz Han Çinli erkek gönüllülerin bir örneğinin% 1.94'ünde (7/361), gönüllülerin çoğunun kökeniyle gözlemlendi. Doğu Çin'e (Jiangsu, Zhejiang, Şangay ve Anhui) kadar izleniyor.[13] Kore'de, Haplogroup D-M174, Daejeon'dan alınan bir numunenin% 3,8'inde (5/133) gözlemlenmiştir.[14] 3/85 = Seul'den bir numunenin% 3,5'i,[15] Jeolla'dan bir numunenin% 3,3'ü (3/90),[16] Gyeongsang'dan bir örneğin% 2.4'ü (2/84),[16] Seul'den başka bir numunenin% 2.3'ü (13/573),[14] Chungcheong'dan bir örneğin% 1,4'ü (1/72),[16] Jeju'dan bir örneğin% 1,1'i (1/87),[16] ve Seoul-Gyeonggi'den üçüncü bir numunenin% 0.9'u (1/110).[16] Diğer çalışmalarda, haplogrup D-M174% 6,7 (3/45) olarak gözlenmiştir.[17] ve% 4.0 (3/75)[18] Kore'den alınan numunelerin, numune alanı daha fazla belirtilmeksizin. Y-DNA Haplogrup D'nin Han Çinli ve Koreli üyelerinin filogenetik pozisyonuna ilişkin çok az yüksek çözünürlüklü veri yayınlandı, ancak mevcut veriler, Haplogrup D'nin çoğu Han Çinli üyesinin Tibetliler arasında (ve özellikle de D-M15 sınıfı, aynı zamanda bazı Lolo-Burma ve Hmong-Mien dillerinin konuşmacıları arasında da bulunur), oysa Haplogroup D'nin çoğu Koreli üyesi, aralarında sıklıkla bulunan D-M55 sınıfına ait olmalıdır. Ainu, Ryukyuan, ve Japonca insanlar.[18][16][3]
Haplogrup D Y-DNA, modern popülasyonlar arasında (düşük sıklıkta da olsa) bulunmuştur. Avrasya bozkır, gibi Güney Altaylar (6/96 =% 6,3 D-M174 (xM15),[19] 6/120 =% 5,0 D-P47[20]), Kazaklar (1/54 =% 1,9 D-M174,[17] 6/1294 =% 0,5 D[21]), Nogais (4/76 =% 5,3 D-M174 Kara Nogai,[22] 1/87 =% 1,1 D-M174 Kuban Nogai[22]), Khalkhas (1/24 =% 4,2 D-M174,[17] 3/85 =% 3,5 D-M174,[15] 2/149 D-M15 + 2/149 D-P47 = 4/149 =% 2.7 D-M174 toplam[18]), Zakhchin (2/60 =% 3,3 D-M174[15]), Uriankhai (1/60 =% 1,7 D-M174[15]), ve Kalmyks (5/426 =% 1,2 D-M174[23]). Ayrıca dilbilimsel olarak benzer (Türk - veya Moğol - konuşan) çölün modern popülasyonları ve bozkır güneyindeki vaha kuşağı, örneğin Yugurlar, Bao'an, Monguors, Uygurlar, ve Özbekler. Ticari testlerde, üyeler batıya kadar bulundu. Romanya Avrupa'da ve Irak Batı Asya'da.[24]
Haplogroup C-M217'den farklı olarak, Haplogroup D-M174 Yeni Dünya'da bulunmaz; herhangi bir modern Kızılderili (Kuzey, Orta veya Güney) popülasyonunda mevcut değildir. Haplogroup C-M217 gibi Yeni Dünya'ya seyahat etmiş olması mümkün olsa da, görünüşe göre bu soyların nesli tükendi.
Haplogrup D-M174, Y kromozomları Haplogroup D-M174'e ait olan bireylerin büyük bir yüzdesini içeren popülasyonların her birinde özel olarak bulunan Haplogroup D-M174 kromozomlarının farklı bir alt kümesiyle, oldukça aşırı coğrafi farklılaşması ile de dikkat çekicidir: Haplogrup D-M15 arasında Tibetliler (ve diğerleri arasında Doğu Asya ve Güneydoğu Asyalı Haplogroup D-M174 Y-kromozomlarının düşük frekanslarını gösteren popülasyonlar), Haplogrup D-M55 Japon Takımadalarının çeşitli popülasyonları arasında ve düşük sıklıkta Koreliler, Haplogroup D-P99 Tibet sakinleri ve orta Avrasya'nın diğer bazı bölgeleri arasında (Örneğin. Moğolistan[25] ve Altay[18][19][20]). D-M15 veya D-M55 alt kanatları pozitif test edilmemiş D-M174 *, yüksek frekanslarda bulunur. Andaman Adalılar ve son zamanlarda Andamanese alt sınıfının D-Y34637 (D1a2b).[26] Test edilmiş pozitif D-M15, D-P47 veya D-M55 alt kanatları olmayan başka bir D-M174 * paragraf grubu türü (veya türü), aşağıdakiler arasında çok düşük bir frekansta bulunur. Türk ve Moğol popülasyonları Orta Asya toplamda en fazla% 1 tutarında. Haplogrup D-M174'ün bu görünüşte eski çeşitliliği, bunun bir "süper haplogrup" veya "makro haplogrup" olarak daha iyi karakterize edilebileceğini göstermektedir.
Bir çalışmada, Thais arasında bulunan test edilmiş pozitif alt bıçaklar olmadan Haplogroup D-M174 sıklığı% 10'du.[2] Su et al. (2000), Tayland'dan alınan üç numunenin% 11.1'inde (5/45) DE-YAP / DYS287 (xM15) buldu (% 20 (4/20) Kuzey Tayland,% 20 (1/5) dahil) Yani ve% 0 (0/20) Kuzeydoğu Tayland) ve% 16,7 (1/6) Guam.[27] Bu arada, yazarlar D-M15'i bir çift örneğin% 15'inde buldular. Yao (% 30 (3/10) Yao dahil Jinxiu ve% 0 (0/10) Yao Nandan ), Bir örneklemin% 14,3'ü (2/14) Yi Örneklemin% 3,8'i (1/26) Kamboçyalılar ve bir numunenin% 3.6'sı (1/28) Zhuang.[27] Dong et al. (2002), DE-YAP Y kromozomlarını bir numunenin% 12.5'inde (2/16) buldu. Jingpo itibaren Luxi Şehri, Yunnan,% 10.0 (2/20) Dai itibaren Luxi Şehri, Yunnan ve% 1,82 (1/55) örnek Nu itibaren Gongshan ve Fugong Yunnan ilçeleri.[28]
Dağıtım
Haplogrup D-M174 Y kromozomları arasında bulunan Tibeto-Burman popülasyonlar ve Japon Takımadaları (haplogroup D1a2b, D1a2a ve D1a1) halkı. D-M55 (D1a2a) özellikle belirgindir, Haplogroup D-M174'ün bir iç dalı boyunca en az beş ayrı mutasyondan oluşan bir kompleks taşır. soyoluş, böylece onları Tibetliler ve Andaman Adalıları arasında bulunan diğer Haplogroup D-M174 kromozomlarından açıkça ayırır ve Y kromozomu Haplogroup D-M55'in tarihöncesini geliştiren ata popülasyonundaki modal haplogrup olduğuna dair kanıt sağlar. Jōmon Japon adalarında kültür.
D-M174 Y kromozom taşıyıcılarının çoğunun Orta Asya -e Doğu Asya. Bir grup, Andaman Adaları, böylece oluşturan veya oluşturmaya yardımcı olan Andamanese insanlar. Başka bir grup, modern Tibet ve güney Çin'de kaldı (bugün Tibeto-Burman halklar) ve başka bir grup muhtemelen Japonya'ya göç etti. Kore Yarımadası (öncesiJōmon insanlar ).[2][18]
Alt kanat
D-Z27276 (D1a1)
Haplogrup D-Z27276 D-M15 ve D-P99'un ortak atasıdır.
D-M15 (D1a1a)
D-M15'in ilk olarak Kamboçya ve Laos'tan alınan bir örnekte (1/18 =% 5,6) ve Japonya'dan bir örnekte (1/23 =% 4,3), Y-DNA varyasyonunun dünya çapında ön araştırmasında bulunduğu bildirilmiştir. mevcut insan popülasyonları.[29]
Daha sonra, Haplogroup D-M15'e ait olan Y-DNA, Tibeto-Burman konuşan nüfus Güneybatı Çin (yaklaşık% 23'ü dahil Qiang,[2][30][31] yaklaşık% 12,5'i Tibetliler,[2] ve yaklaşık% 9'u Yi[2][32]) ve arasında Yao insanlar kuzeydoğuda yaşayan Guangxi (6/31 =% 19,4 Lowland Yao, 5/41 =% 12,2 Yerli Mien, 3/41 =% 7,3 Lowland Kimmun)[33] geneline ılımlı bir dağılımla Orta Asya, Doğu Asya ve kıtasal Güneydoğu Asya (Çinhindi ).[2]
2011'de yayınlanan bir araştırma, D-M15'i bir örneklemin% 7,8'inde (4/51) buldu. Hmong Daw ve bir numunenin% 3,4'ünde (1/29) Xinhmul kuzeyden Laos.[33]
D-P47 (D1a1b1)
Aralarında yüksek sıklıkta bulundu Pumi,[2] Naxi,[2] ve Tibetliler,[34][2] ılımlı bir dağılımla Orta Asya.[2] Bir araştırmaya göre, Tibetliler haplogrup D-P47'nin yaklaşık% 41.31'lik bir frekansına sahipler.[2] Haplogrup D-P47, Xi'an Han'ın% 9'unda bulundu.[35]
D-M55 (D1a2a)
Daha önce olarak biliniyordu D-M55, D-M64.1 / Page44.1 (D1a2a), Ainu[36] ve orta sıklıkta Japonca[37] ve Ryukyuans.[37]
Kim et al. (2011), Haplogroup D-M55'i bir numunenin% 2.0'ında (1/51) buldu. Pekin Han ve Jeolla bölgesinden% 3,3 (3/90), Gyeongsang bölgesinden% 2,4 (2/84),% 1,4 (1/72) dahil olmak üzere Güney Kore'den bir numune havuzunun% 1,6'sında (8/506) Chungcheong bölgesinden, Jeju bölgesinden% 1.1 (1/87), Seul-Gyeonggi bölgesinden% 0.9 (1/110) ve Gangwon bölgesinden% 0 (0/63).[16] Çekiç et al. (2006) Haplogroup D-P37.1'i Güney Kore'den bir numunenin% 4.0'ında (3/75) buldu.[18]
Günümüzde erkekler arasında düşük D-M116.1 seviyeleri (D-M55'in bir alt sınıfı) Timor (Erkeklerin% 0,2'si),[38] Erkeklerin% 9.5'inde bulunur Mikronezya (Hammer ve ark. 2006,[18] Japonya'dan gelen son karışımı yansıttığına inanılıyor. Diğer bir deyişle, D-M116.1 (D1a2a1) 'in genellikle D-M64.1'in (D1a2a) birincil alt sınıfı olduğuna inanılmaktadır, muhtemelen Güneydoğu Asya'nın Japon askeri işgali sırasında Dünya Savaşı II.
Göre Mitsuru Sakitani Haplogroup D1 Orta Asya'dan kuzeye geldi Kyushu aracılığıyla Altay Dağları ve Kore Yarımadası 40.000 yıldan fazla bir süre önce ve Haplogroup D-M55 (D1a2a) Japon takımadalarında doğdu.[39]
Son zamanlarda, haplogroup D-M55'in Japon şubesinin 53.000 yıldan beri diğer D şubelerinden farklı ve izole olduğu doğrulandı. D1a arasındaki bölünme muhtemelen Orta Asya'da meydana geldi, bazıları ise haplogroup D'nin başlangıcında ani bir bölünme olduğunu öne sürüyor, çünkü Japon dalı, başka hiçbir D dalında bulunmayan beş benzersiz mutasyona sahip.[8]
(D *) Paragroup D1a2b (D34637)
D1a2b, yüksek frekanslarda bulunur: Andaman Adalılar[2] (özellikle Onge (23/23 = 100%), Jarawa (4/4 = 100%).[40][26]
D1a *
D1a * (xM15, P99, M55) bazı Tibet azınlık kabilelerinde bulunur. Kuzeydoğu Hindistan (oranlar sıfır ile% 65 arasında değişmektedir).[5][41][42][43]
D-M175 *
Bazal D-M174 * (xM15, P47, M55) yaklaşık% 5'inde bulunmuştur. Altaylar.[18] Kharkov et al. Haplogroup D * (xD-M15), özellikle Kulada (5/46 =% 10.9) ve Kosh-Agach (1/7) olmak üzere üç farklı bölgeden Güney Altaylıların örnek havuzunun% 6,3'ünde (6/96) bulundu. =% 14), ancak alt sınıf D-M55 veya D-P99'un herhangi bir markörü için test edilmemişlerdir. Kharkov et al. ayrıca Beshpeltir'den bir Güney Altaylı bireyde haplogroup DE-M1 (xD-M174) Y-DNA bulduklarını bildirdi (1/43 =% 2.3).[19]
Filogenetik
Filogenetik tarih
2002'den önce, akademik literatürde Y-Kromozom Filogenetik ağacı için en az yedi adlandırma sistemi vardı. Bu, ciddi bir kafa karışıklığına yol açtı. 2002 yılında, büyük araştırma grupları bir araya gelerek Y-Kromozom Konsorsiyumu'nu (YCC) oluşturdu. Herkesin kullanmayı kabul ettiği tek bir yeni ağaç oluşturan ortak bir makale yayınladılar. Daha sonra popülasyon genetiği ve genetik şecere ile ilgilenen bir grup yurttaş bilim adamı, her şeyden önce zamanında olmayı amaçlayan amatör bir ağaç oluşturmak için bir çalışma grubu oluşturdu. Aşağıdaki tablo, tüm bu çalışmaları, dönüm noktası olan 2002 YCC Ağacı noktasında bir araya getiriyor. Bu, eski yayınlanmış literatürü gözden geçiren bir araştırmacının adlandırmalar arasında hızla hareket etmesini sağlar.
| YCC 2002/2008 (Kısaltma) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | YCC 2002 (Uzun El) | YCC 2005 (Uzun El) | YCC 2008 (Uzun El) | YCC 2010r (Uzun El) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D-M174 | * | * | * | * | * | * | * | * | D | D | D | D | D | D | D | D | D | D |
| D-M15 | 4 | IV | 3G | 12 | Eu5 | H3 | B | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 | D1 |
| D-M55 | * | * | * | * | * | * | * | * | D2 | D2 | D2 | D2 | D2 | D2 | D2 | D2 | D2 | D2 |
| D-P12 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2a | D2a | D2a1a1 | D2a1a1 | D2 | D2 | D2a1a1 | D2a1a1 | D2a1a1 | kaldırıldı | kaldırıldı |
| D-M116.1 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2b * | D2a | D2a | D2a | D2a | D2a | D2a | D2a | D2a | kaldırıldı | kaldırıldı |
| D-M125 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2b1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 | D2a1 |
| D-M151 | 4 | IV | 3G | 11 | Eu5 | H2 | B | D2b2 | D2a1 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 | D2a2 |
Araştırma yayınları
Yayınlarına göre aşağıdaki araştırma ekipleri YCC ağacının oluşturulmasında temsil edildi.
Filogenetik ağaçlar
ISOGG ağacına göre (Sürüm: 14.151).[44]
- DE (YAP)
- D (CTS3946)
- D1 (M174 / Sayfa30, IMS-JST021355)
- D1a (CTS11577)
- D1a1 (F6251 / Z27276)
- D1a1a (M15)Çin toprakları, Tibet
- D1a1b (S99)Çin toprakları, Tibet, Moğol, Orta Asya
- D1a2(Z3660)
- D1a2a (M64.1 / Sayfa44.1, M55) Japonya (Yamato insanlar 、Ryukyuan insanlar 、Ainu insanlar )
- D1a2b (Y34637)Andaman Adaları (Onge insanlar 、Jarawa insanlar )
- D1a1 (F6251 / Z27276)
- D1b (L1378) Filipinler[45]
- D1a (CTS11577)
- D2 (A5580.2)Nijerya, Suudi Arabistan, Suriye
- D1 (M174 / Sayfa30, IMS-JST021355)
- D (CTS3946)
Ayrıca bakınız
Genetik
Y-DNA D-M174 alt kanatları
Y-DNA omurga ağacı
| İnsanın filogenetik ağacı Y kromozom DNA haplogrupları [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y kromozomal Adam " | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ben | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | T | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | P [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HAYIR | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | Ö | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Referanslar
- ^ "Y-DNA Haplogrup D-M174 ve Alt Kladları - 2017".
- ^ a b c d e f g h ben j k l m n Ö p Shi H, Zhong H, Peng Y, Dong YL, Qi XB, Zhang F, Liu LF, Tan SJ, Ma RZ, Xiao CJ, Wells RS, Jin L, Su B (Ekim 2008). "Doğu Asya'daki en eski modern insan yerleşiminin ve Tibet ve Japon popülasyonlarının çeşitli kökenlerinin Y kromozomu kanıtı". BMC Biyoloji. 6: 45. doi:10.1186/1741-7007-6-45. PMC 2605740. PMID 18959782.
- ^ a b c YFull Haplogroup YTree v6.02, 02 Nisan 2018. Erişim tarihi 7 Temmuz 2018.
- ^ a b c Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (Mayıs 2008). "Yeni ikili polimorfizmler, insan Y kromozom haplogrup ağacını yeniden şekillendirir ve çözünürlüğünü artırır". Genom Araştırması. 18 (5): 830–8. doi:10.1101 / gr.7172008. PMC 2336805. PMID 18385274.
- ^ a b Su B, Xiao C, Deka R, Seielstad MT, Kangwanpong D, Xiao J, Lu D, Underhill P, Cavalli-Sforza L, Chakraborty R, Jin L (Aralık 2000). "Y kromozom haplotipleri Himalayalara tarih öncesi göçleri ortaya koyuyor". İnsan Genetiği. 107 (6): 582–90. doi:10.1007 / s004390000406. PMID 11153912. S2CID 36788262.
- ^ Gayden, Tenzin; Cadenas, Alicia M .; Regueiro, Maria; Singh, Nanda B .; Zhivotovsky, Lev A .; Underhill, Peter A .; Cavalli-Sforza, Luigi L .; Herrera, Rene J. (Mayıs 2007). "Gen Akışına Yönel Bir Bariyer Olarak Himalayalar". Amerikan İnsan Genetiği Dergisi. 80 (5): 884–894. doi:10.1086/516757. PMC 1852741. PMID 17436243.
- ^ Matsumoto H (Şubat 2009). "Japon ırkının kökeni, immünoglobulin G'nin genetik belirteçlerine dayanmaktadır". Japonya Akademisi Tutanakları. Seri B, Fiziksel ve Biyolojik Bilimler. 85 (2): 69–82. Bibcode:2009PJAB ... 85 ... 69 milyon. doi:10.2183 / pjab.85.69. PMC 3524296. PMID 19212099.
- ^ a b Mondal, Mayukh; Bergström, Anders; Xue, Yalı; Calafell, Francesc; Laayouni, Hafid; Casals, Ferran; Majumder, Partha P .; Tyler-Smith, Chris; Bertranpetit, Jaume (2017/05/01). "Çeşitli Hint popülasyonlarının Y-kromozom dizileri ve Andamanese soyları". İnsan Genetiği. 136 (5): 499–510. doi:10.1007 / s00439-017-1800-0. hdl:10230/34399. ISSN 1432-1203. PMID 28444560. S2CID 3725426.
Buna karşılık, Riang (Tibeto-Burman konuşan) ve Andamanlıların en yakın komşuları Doğu Asya'da var. Jarawa ve Onge, haplogroup D soylarını son ~ 7000 yıl içinde birbirleriyle paylaştılar, ancak ~ 53000 yıl önce Japon haplogroup D Y kromozomlarından, büyük olasılıkla paylaşılan bir ata popülasyonundan ayrılma nedeniyle ayrıldılar.
- ^ Tyler-Smith, Chris; Xue, Yalı; Thomas, Mark G .; Yang, Huanming; Arciero, Elena; Asan; Connell, Bruce A .; Jones, Abigail L .; Haber, Marc (2019-06-13). "Nadir Bir Derin Köklere Sahip D0 Afrika Y-Kromozomal Haplogrubu ve Modern İnsanların Afrika'dan Genişlemesi İçin Etkileri". Genetik. 212 (4): 1421–1428. doi:10.1534 / genetik.119.302368. ISSN 0016-6731. PMC 6707464. PMID 31196864.
- ^ Estes, Roberta (2019-06-21). "Heyecan verici Yeni Y DNA Haplogrup D Keşifleri!". DNAeXplained - Genetik Şecere. Alındı 2019-07-08.
- ^ Y 染色体 单 倍 群 D 在 東亞 的 分布 及其 意義
- ^ Zhong H, Shi H, Qi XB, Duan ZY, Tan PP, Jin L, Su B, Ma RZ (2011). "Genişletilmiş Y Kromozomu Araştırması Modern İnsanların Kuzey Rotası Üzerinden Doğu Asya'ya Buzul Sonrası Göçlerini Öneriyor". Mol. Biol. Evol. 28 (1): 717–727. doi:10.1093 / molbev / msq247. PMID 20837606.
- ^ Yan S, Wang CC, Li H, Li SL, Jin L, ve diğerleri. (The Genographic Consortium) (Eylül 2011). "Y kromozom Haplogrup O'nun güncellenmiş bir ağacı ve P164 ve PK4 mutasyonlarının filogenetik pozisyonları revize edildi". Avrupa İnsan Genetiği Dergisi. 19 (9): 1013–5. doi:10.1038 / ejhg.2011.64. PMC 3179364. PMID 21505448.
- ^ a b Park MJ, Lee HY, Yang WI, Shin KJ (Temmuz 2012). "Kore'deki Y kromozom varyasyonunu anlama - birleşik haplogrup ve haplotip analizlerinin alaka düzeyi". Uluslararası Adli Tıp Dergisi. 126 (4): 589–99. doi:10.1007 / s00414-012-0703-9. PMID 22569803. S2CID 27644576.
- ^ a b c d Katoh T, Munkhbat B, Tounai K, Mano S, Ando H, Oyungerel G, Chae GT, Han H, Jia GJ, Tokunaga K, Munkhtuvshin N, Tamiya G, Inoko H (Şubat 2005). "Y kromozom analizi ile ortaya çıkan Moğol etnik gruplarının genetik özellikleri". Gen. 346: 63–70. doi:10.1016 / j.gene.2004.10.023. PMID 15716011.
- ^ a b c d e f g Kim, Soon-Hee; Kim, Ki-Cheol; Shin, Dong-Jik; Jin, Han-Jun; Kwak, Kyoung-Don; Han, Myun-Soo; Song, Joon-Myong; Kim, Won; Kim, Wook (2011). "Kore'de Y kromozom haplogrup O2b-SRY465 soylarının yüksek frekansları: Kore halkına genetik bir bakış açısı". Araştırmacı Genetik. 2 (1): 10. doi:10.1186/2041-2223-2-10. PMC 3087676. PMID 21463511.
- ^ a b c Wells RS, Yuldasheva N, Ruzibakiev R, Underhill PA, Evseeva I, Blue-Smith J, Jin L, Su B, Pitchappan R, Shanmugalakshmi S, Balakrishnan K, Read M, Pearson NM, Zerjal T, Webster MT, Zholoshvili I, Jamarjashvili E, Gambarov S, Nikbin B, Dostiev A, Aknazarov O, Zalloua P, Tsoy I, Kitaev M, Mirrakhimov M, Chariev A, Bodmer WF (Ağustos 2001). "Avrasya'nın merkezi: Y kromozom çeşitliliğine kıtasal bir bakış açısı". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 98 (18): 10244–9. Bibcode:2001PNAS ... 9810244W. doi:10.1073 / pnas.171305098. PMC 56946. PMID 11526236.
- ^ a b c d e f g h Hammer MF, Karafet TM, Park H, Omoto K, Harihara S, Stoneking M, Horai S (2006). "Japonların ikili kökeni: avcı-toplayıcı ve çiftçi Y kromozomları için ortak zemin". İnsan Genetiği Dergisi. 51 (1): 47–58. doi:10.1007 / s10038-005-0322-0. PMID 16328082.
- ^ a b c Khar'kov VN, Stepanov VA, Medvedeva OF, Spiridonova MG, Voevoda MI, Tadinova VN, Puzyrev VP (Mayıs 2007). "[Kuzey ve güney Altaylılar arasındaki gen havuzu farklılıkları, Y kromozom haplogrupları hakkındaki verilerden çıkarılmıştır]". Genetika (Rusça). 43 (5): 675–87. doi:10.1134 / S1022795407050110. PMID 17633562. S2CID 566825.
- ^ a b Dulik MC, Zhadanov SI, Osipova LP, Askapuli A, Gau L, Gokcumen O, Rubinstein S, Schurr TG (Şubat 2012). "Mitokondriyal DNA ve Y kromozomu varyasyonu, Yerli Amerikalılar ve Yerli Altaylılar arasındaki son zamanlardaki ortak atalara kanıt sağlıyor". Amerikan İnsan Genetiği Dergisi. 90 (2): 229–46. doi:10.1016 / j.ajhg.2011.12.014. PMC 3276666. PMID 22281367.
- ^ EE Ashirbekov, DM Botbaev, AM Belkozhaev, AO Abayldaev, AS Neupokoeva, JE Mukhataev, B. Alzhanuly, DA Sharafutdinova, DD Mukushkina, MB Rakhymgozhin, AK Khanseitova, SA Limborska ve NA Aytkhozhina "Y-Chromosome Dağılımı Güney Kazakistan, Jambıl ve Almatı Bölgelerinden Kazak. " Kazakistan Cumhuriyeti Ulusal Bilimler Akademisi Raporları, ISSN 2224-5227, Cilt 6, Sayı 316 (2017), 85-95.
- ^ a b Yunusbayev B, Metspalu M, Järve M, Kutuev I, Rootsi S, Metspalu E, Behar DM, Varendi K, Sahakyan H, Khusainova R, Yepiskoposyan L, Khusnutdinova EK, Underhill PA, Kivisild T, Villems R (Ocak 2012). "Eski insan göçlerine karşı asimetrik yarı geçirgen bir bariyer olarak Kafkasya". Moleküler Biyoloji ve Evrim. 29 (1): 359–65. doi:10.1093 / molbev / msr221. PMID 21917723.
- ^ Malyarchuk B, Derenko M, Denisova G, Khoyt S, Woźniak M, Grzybowski T, Zakharov I (Aralık 2013). "Kalmyks'te etnik ve kabile düzeyinde Y kromozom çeşitliliği". İnsan Genetiği Dergisi. 58 (12): 804–11. doi:10.1038 / jhg.2013.108. PMID 24132124.
- ^ Y-DNA D Haplogrup Projesi Family Tree DNA şirketinde
- ^ Di Cristofaro J, Pennarun E, Mazières S, Myres NM, Lin AA, Temori SA, Metspalu M, Metspalu E, Witzel M, King RJ, Underhill PA, Villems R, Chiaroni J (2013). "Afgan Hindu Kush: Avrasya alt kıta gen akışlarının birleştiği yer". PLOS ONE. 8 (10): e76748. Bibcode:2013PLoSO ... 876748D. doi:10.1371 / journal.pone.0076748. PMC 3799995. PMID 24204668.
- ^ a b Y-Dolu
- ^ a b Su, Bing; Jin, Li; Underhill, Peter; Martinson, Jeremy; Saha, Nilmani; McGarvey, Stephen T .; Shriver, Mark D .; Chu, Jiayou; Oefner, Peter; Chakraborty, Ranajit; Deka, Ranjan (18 Temmuz 2000). "Polinezya kökenleri: Y kromozomundan içgörüler". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 97 (15): 8225–8228. Bibcode:2000PNAS ... 97.8225S. doi:10.1073 / pnas.97.15.8225. PMC 26928. PMID 10899994.
- ^ DONG Yongli, SHI Hong, LI Weixiang, YANG Jie, ZENG Weimin, LI Kaiyuan ve XIAO Chunjie, "YAP lokusunda, Salween Nehri'nin büyük geçidinde ve aşağı havzada bulunan yedi Yunnan etnik azınlık popülasyonunda polimorfizm çalışması" (orijinal Çince başlık: "怒江 大 峡谷 及 下游 地区 7 个 云南 少数民族 YAP 位 点 的 多态性 研究"), Acta Anthropologica Sinica, Cilt. 21, No. 3 (Ağustos 2002). http://www.ivpp.cas.cn/cbw/rlxxb/xbwzxz/201203/t20120320_3512811.html
- ^ Underhill PA, Shen P, Lin AA, Jin L, Passarino G, Yang WH, Kauffman E, Bonné-Tamir B, Bertranpetit J, Francalacci P, Ibrahim M, Jenkins T, Kidd JR, Mehdi SQ, Seielstad MT, Wells RS, Piazza A, Davis RW, Feldman MW, Cavalli-Sforza LL, Oefner PJ (Kasım 2000). "Y kromozom dizisi varyasyonu ve insan popülasyonlarının tarihi". Doğa Genetiği. 26 (3): 358–61. doi:10.1038/81685. PMID 11062480. S2CID 12893406.
- ^ Xue Y, Zerjal T, Bao W, Zhu S, Shu Q, Xu J, Du R, Fu S, Li P, Hurles ME, Yang H, Tyler-Smith C (Nisan 2006). "Doğu Asya'da erkek demografisi: insan nüfusu artış zamanlarında bir kuzey-güney zıtlığı". Genetik. 172 (4): 2431–9. doi:10.1534 / genetik.105.054270. PMC 1456369. PMID 16489223.
- ^ Wang CC, Wang LX, Shrestha R, Zhang M, Huang XY, Hu K, Jin L, Li H (2014). "Batı Siçuan koridorunda ikamet eden Qiangic popülasyonlarının genetik yapısı". PLOS ONE. 9 (8): e103772. Bibcode:2014PLoSO ... 9j3772W. doi:10.1371 / journal.pone.0103772. PMC 4121179. PMID 25090432.
- ^ Wen B, Xie X, Gao S, Li H, Shi H, Song X, Qian T, Xiao C, Jin J, Su B, Lu D, Chakraborty R, Jin L (Mayıs 2004). "Tibeto-Burman popülasyonlarının genetik yapısının analizleri, güney Tibeto-Burmanlarda cinsiyete dayalı bir karışım ortaya koyuyor". Amerikan İnsan Genetiği Dergisi. 74 (5): 856–65. doi:10.1086/386292. PMC 1181980. PMID 15042512.
- ^ a b Cai X, Qin Z, Wen B, Xu S, Wang Y, Lu Y, Wei L, Wang C, Li S, Huang X, Jin L, Li H (2011). "Son Buzul Maksimumda Güneydoğu Asya'dan Doğu Asya'ya darboğazlar yoluyla insan göçü Y kromozomları tarafından ortaya çıkarıldı". PLOS ONE. 6 (8): e24282. Bibcode:2011PLoSO ... 624282C. doi:10.1371 / journal.pone.0024282. PMC 3164178. PMID 21904623.
- ^ Dongsheng Lu et al. Tibet Dağlılarının Atalarının Kökenleri ve Genetik Tarihi, 25 Ağustos 2016
- ^ Kim 2011
- ^ Tajima A, Hayami M, Tokunaga K, Juji T, Matsuo M, Marzuki S, Omoto K, Horai S (2004). "Ainu'nun genetik kökenleri, anne ve baba soylarının birleşik DNA analizlerinden çıkarılmıştır". İnsan Genetiği Dergisi. 49 (4): 187–93. doi:10.1007 / s10038-004-0131-x. PMID 14997363.
- ^ a b YOUICHI SATO, TOSHIKATSU SHINKA, ASHRAF A. EWIS, AIKO YAMAUCHI, TERUAKI IWAMOTO, YUTAKA NAKAHORI Modern Japon erkeklerinin Y kromozomundaki genetik varyasyona genel bakış.
- ^ Tumonggor, Meryanne K; Karafet, Tatiana M; Downey, Sean; Lansing, J Stephen; Norquest, Peter; Sudoyo, Herawati; Hammer, Michael F; Cox, Murray P (Eylül 2014). "Batı Timor'da izolasyon, temas ve sosyal davranış biçimindeki genetik çeşitlilik". İnsan Genetiği Dergisi. 59 (9): 494–503. doi:10.1038 / jhg.2014.62. PMC 4521296. PMID 25078354.
- ^ 崎 谷 満 『DNA ・ 考古 ・ 言語 の 学 際 研究 が 示 す 新 ・ 日本 列島 史』 (勉 誠 出版 2009 年) (Japonca)
- ^ Thangaraj K, Singh L, Reddy AG, Rao VR, Sehgal SC, Underhill PA, Pierson M, Frame IG, Hagelberg E (Ocak 2003). "Andaman Adalılarının genetik yakınlıkları, yok olan bir insan popülasyonu". Güncel Biyoloji. 13 (2): 86–93. doi:10.1016 / S0960-9822 (02) 01336-2. PMID 12546781. S2CID 12155496.
- ^ Cordaux R, Weiss G, Saha N, Stoneking M (Ağustos 2004). "Kuzeydoğu Hindistan geçidi: insan göçleri için bir engel mi yoksa koridor mu?". Moleküler Biyoloji ve Evrim. 21 (8): 1525–33. doi:10.1093 / molbev / msh151. PMID 15128876.
- ^ Chandrasekar A, Saheb SY, Gangopadyaya P, Gangopadyaya S, Mukherjee A, Basu D, Lakshmi GR, Sahani AK, Das B, Battacharya S, Kumar S, Xaviour D, Sun D, Rao VR (2007). "Güney Asya'da YAP ekleme imzası". İnsan Biyolojisi Yıllıkları. 34 (5): 582–6. doi:10.1080/03014460701556262. PMID 17786594. S2CID 11860142.
- ^ Reddy BM, Langstieh BT, Kumar V, Nagaraja T, Reddy AN, Meka A, Reddy AG, Thangaraj K, Singh L (Kasım 2007). "Kuzeydoğu Hindistan'ın Avusturya-Asya kabileleri, Güney ve Güneydoğu Asya arasında şimdiye kadar eksik olan genetik bağı sağlıyor". PLOS ONE. 2 (11): e1141. Bibcode:2007PLoSO ... 2.1141R. doi:10.1371 / journal.pone.0001141. PMC 2065843. PMID 17989774.
- ^ Y-DNA Haplogrup D ve Alt Kladları - 2019
- ^ Y-DNA Haplogrup D ve Alt Kladları - 2014
- Underhill PA, Kivisild T (2007). "Y kromozomunun ve mitokondriyal DNA popülasyon yapısının insan göçlerinin izlenmesinde kullanılması". Genetik Yıllık İnceleme. 41: 539–64. doi:10.1146 / annurev.genet.41.110306.130407. PMID 18076332. S2CID 24904955.