Moleküler paleontoloji - Molecular paleontology
| Bir dizinin parçası |
| Paleontoloji |
|---|
 |
Paleontoloji tarihi |
| Paleontoloji Portalı Kategori |
Moleküler paleontoloji geri kazanımı ve analizini ifade eder DNA, proteinler, karbonhidratlar veya lipidler, ve onların diyajenetik eski insan, hayvan ve bitki kalıntılarından ürünler.[1][2] Moleküler paleontoloji alanı, evrimsel olaylara, türlere ilişkin önemli bilgiler sağlamıştır. diasporalar soyu tükenmiş olanların keşfi ve karakterizasyonu Türler. DNA'ya moleküler analitik teknikler uygulayarak fosiller DNA'nın geri kazanıldığı herhangi iki organizma arasındaki akrabalık seviyesi ölçülebilir.[3]
Moleküler paleontoloji alanındaki gelişmeler, bilim adamlarının evrimsel güvenmek yerine genetik düzeyde sorular fenotipik tek başına varyasyon. Gibi çeşitli biyoteknolojik teknikleri kullanma DNA izolasyonu, amplifikasyon, ve sıralama[4] bilim adamları, sayısız organizmanın farklılaşması ve evrimsel geçmişine ilişkin genişletilmiş yeni anlayışlar elde etmeyi başardılar.
Tarih
Moleküler paleontoloji çalışmasının, Abelson'un fosil kabuklarda korunmuş 360 milyon yıllık amino asitleri keşfetmesiyle başladığı söyleniyor.[5] Ancak, Svante Pääbo genellikle moleküler paleontoloji alanının kurucusu olarak kabul edilen kişidir.[6]
Moleküler paleontoloji alanı, 1950'lerden bu yana birçok büyük ilerleme kaydetmiştir ve sürekli büyüyen bir alandır. Aşağıda, yapılan dikkate değer katkıları gösteren bir zaman çizelgesi bulunmaktadır.
Zaman çizelgesi

1950'lerin ortası: Abelson korunmuş olarak bulundu amino asitler yaklaşık 360 milyon yıllık fosil kabuklarında. Moleküler evrimin incelenebilmesi için fosil amino asit dizilerinin mevcut organizma ile karşılaştırılması fikri üretildi.[5]
1970'ler: Fosil peptidler, amino asit analizi.[7] Tüm peptitleri kullanmaya başlayın ve immünolojik yöntemler.[8]
1970'lerin sonu: Paleobotanistler (Paleobotanistler olarak da yazılabilir) iyi korunmuş fosil bitkilerinden molekülleri inceledi.[9]
1984: Soyu tükenmiş bir türün ilk başarılı DNA dizilimi olan Quagga, zebra benzeri bir tür.[1]
1991: Bir dinozorun fosil kemiğinden, özellikle de sismosaurus'tan başarılı bir şekilde protein çıkarılması üzerine yayınlanan makale.[10]
2005: Bilim adamları nesli tükenmiş olanı diriltiyor 1918 grip virüsü.[11]
2006: Neandertaller nükleer DNA dizisi segmentleri analiz edilmeye ve yayınlanmaya başlanır.[16]
2007: Bilim adamları soyu tükenmiş olanın tamamını sentezliyor insan endojen retrovirüsü (HERV-K) sıfırdan.[12]
2010: Yeni bir erken hominid türü olan Denisovalılar, Sibirya'daki bir mağarada bulunan kemikten elde edilen mitokondriyal ve nükleer genomlardan keşfedildi. Analiz, Denisovan örneğinin yaklaşık 41.000 yıl önce yaşadığını ve yaklaşık 1 milyon yıl önce Afrika'da hem modern insanlarla hem de Neandertallerle ortak bir atayı paylaştığını gösterdi.[13]
2013: İlk bütün Neandertal genomu başarıyla sıralandı. Daha fazla bilgi şu adreste bulunabilir: Neandertal genom projesi.[14]
2013: Kalıntı içeren 400.000 yıllık bir örnek mitokondriyal DNA dizilenmiş ve Neandertallerin ve Denisovalıların ortak bir atası olduğu bulunmuştur. Homo heidelbergensis.[15]
2015: DNA içeren 110.000 yıllık fosil diş Denisovalılar rapor edildi.[17][18]
The quagga
İlk başarılı DNA dizilimi nesli tükenmiş türler 1984 yılında, zebra benzeri bir tür olan quagga'nın 150 yıllık bir müze örneğinden alınmıştır.[1] Mitokondriyal DNA (mtDNA olarak da bilinir), kurutulmuş quagga kası ve bir dağ zebrasının mitokondriyal DNA'sından 12 baz ikamesi ile farklı olduğu bulundu. Bu iki türün 3-4 milyon yıl önce ortak bir ataya sahip olduğu sonucuna varıldı ki bu da bilinenle tutarlıdır. fosil türlerin kanıtı.[19]
Denisovalılar
Denisovalılar nın-nin Avrasya Neandertallerle ve insanlarla ilişkili bir hominid türü olan, 2008'de elde edilen 41.000 yıllık bir örneğin DNA dizilişinin doğrudan bir sonucu olarak keşfedildi. Alınan bir parmak kemiğinden mitokondriyal DNA'nın analizi, örneğin her ikisinden de genetik olarak farklı olduğunu gösterdi. insanlar ve Neandertaller. Daha sonra iki diş ve bir ayak kemiğinin aynı popülasyona sahip farklı bireylere ait olduğu bulundu. Analizler, modern insanlar geldiğinde hem Neandertallerin hem de Denisovanların Avrasya'da zaten mevcut olduğunu gösteriyor.[14] Kasım 2015'te bilim adamları, DNA içeren bir fosil diş bulduklarını bildirdi. Denisovalılar ve yaşının 110.000 yaşında olduğu tahmin ediliyor.[17][18]
Mitokondriyal DNA analizi

Denisovalı parmak kemiğindeki mtDNA, modern insanlarınkinden 385 baz ile farklıdır (nükleotidler ) yaklaşık 16.500 mtDNA dizisinde, modern insan ve Neandertaller yaklaşık 202 bazdır. Aksine, arasındaki fark şempanzeler ve modern insanlar yaklaşık olarak 1,462 mtDNA baz çiftidir.[20] Bu, yaklaşık bir milyon yıl önce bir sapma zamanı önerdi. Bir dişin mtDNA'sı, parmak kemiğininkine yüksek bir benzerlik taşıyordu, bu da aynı popülasyona ait olduklarını gösteriyordu.[21] İkinci bir dişten, diğer dişte ve parmakta bulunandan beklenmedik şekilde çok sayıda genetik farklılık gösteren bir mtDNA dizisi elde edildi ve bu da yüksek derecede mtDNA çeşitliliği olduğunu düşündürdü. Aynı mağaradan bu iki birey, tüm Avrasya'dan örneklenmiş Neandertallerde görülenden daha fazla çeşitlilik gösterdi ve farklı kıtalardan günümüz insanları kadar farklıydı.[22]
Nükleer genom analizi
Denisova parmak kemiğinden nükleer DNA'nın izolasyonu ve dizilimi de gerçekleştirildi. Bu örnek, alışılmadık derecede DNA koruması ve düşük seviyede kontaminasyon gösterdi. Neandertal ve modern insanlarla ayrıntılı bir karşılaştırmaya olanak tanıyan neredeyse eksiksiz genomik dizileme elde etmeyi başardılar. Bu analizden, mitokondriyal dizilimlerinin belirgin farklılığına rağmen, Denisova popülasyonunun Neandertal ile birlikte modern Afrika insanlarına giden soydan ortak bir dalı paylaştığı sonucuna vardılar. Denisovan ve Neandertal dizileri arasındaki tahmini ortalama uzaklaşma süresi 640.000 yıl önceydi ve bunların ikisi ile modern Afrikalıların dizileri arasındaki süre 804.000 yıl önceydi. Denisova mtDNA'sının ayrışmasının, insanlığın diğer dallarından arındırılmış bir soyun kalıcılığından kaynaklandığını öne sürüyorlar. genetik sürüklenme veya başka bir introgression eski bir hominin soyundan.[21]
Homo heidelbergensis
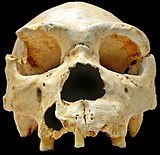
Homo heidelbergensis ilk olarak 1907'de Almanya'nın Heidelberg yakınlarında keşfedildi ve daha sonra Avrupa, Afrika ve Asya'da başka yerlerde de bulundu.[23][24]Bununla birlikte, 2013 yılına kadar, elde edilebilir DNA'ya sahip bir örnek, yaklaşık 400.000 yıllık bir femurda bulundu. Sima de los Huesos Mağarası ispanyada. Femurun hem mtDNA hem de nükleer DNA içerdiği bulundu. DNA ekstraksiyonu ve kütüphane hazırlama tekniklerindeki gelişmeler, mtDNA'nın başarılı bir şekilde izole edilmesine ve dizilenmesine izin verdi, ancak nükleer DNA'nın gözlemlenen örnekte çok bozulmuş olduğu ve ayrıca eski bir mağara ayısından alınan DNA ile kontamine olduğu bulundu (Ursus deningeri ) mağarada bulunur.[25] MtDNA analizi, örnek ile Denisovalılar arasında şaşırtıcı bir bağlantı buldu ve bu bulgu birçok soruyu gündeme getirdi. Ocak 2014 tarihli "Sima de los Huesos'tan bir homininin mitokondriyal genom dizisi" başlıklı bir makalede, bilim camiasında nasıl yakınsama eksikliğini açıklığa kavuşturan birkaç senaryo önerildi. Homo heidelbergensis bilinen diğer hominin grupları ile ilgilidir. Yazarların önerdiği makul bir senaryo şuydu: H. heidelbergensis hem Denisovanların hem de Neandertallerin atasıydı.[25] Hem Denisovalılardan hem de Neandertallerden tamamen sıralanan nükleer genomlar, yaklaşık 700.000 yıl önce ortak bir ata olduğunu gösteriyor ve bu alanda önde gelen bir araştırmacı olan Svante Paabo, belki de bu yeni hominin grubunun bu erken ata olduğunu öne sürüyor.[15]
Başvurular
Yeni türlerin keşfi ve karakterizasyonu
Fosillere uygulanan moleküler paleontoloji teknikleri, Denisovalılar ve diğerleri de dahil olmak üzere birkaç yeni türün keşfedilmesine ve karakterizasyonuna katkıda bulunmuştur. Homo heidelbergensis. İnsanların yeryüzünü doldururken izledikleri yolu ve bu sırada hangi türlerin mevcut olduğunu daha iyi anlayabildik. diaspora.
Yok olma

Canlandırmak artık mümkün nesli tükenmiş moleküler paleontoloji tekniklerini kullanarak türler. Bu ilk olarak şu yolla gerçekleştirildi: klonlama 2003 yılında Pirene dağ keçisi, 2000 yılında nesli tükenen bir tür yaban keçisi. Pirene dağ keçisinin hücrelerinden alınan çekirdekler, kendi DNA'ları boşaltılan keçi yumurtalarına enjekte edildi ve taşıyıcı keçi annelerine implante edildi.[26] Yavru, ciğerlerindeki kusurlar nedeniyle doğumdan yalnızca yedi dakika sonra yaşadı. Diğer klonlanmış hayvanların da benzer akciğer kusurlarına sahip olduğu gözlenmiştir.[27]
İnsan faaliyetlerinin doğrudan bir sonucu olarak nesli tükenen birçok tür var. Bazı örnekler şunları içerir: dodo, harika auk, Tazmanya kaplanı, Çin nehir yunusu, ve yolcu güvercini. Soyu tükenmiş bir tür kullanılarak yeniden canlandırılabilir. alelik değiştirme[28] Hala yaşayan yakından ilişkili bir türün. Sadece birkaçını değiştirerek genler Bir organizmanın içinde, soyu tükenmiş türlerin genomunu sıfırdan inşa etmek zorunda kalmak yerine, birkaç türü, hatta Neandertalleri bile bu şekilde geri getirmek mümkün olabilir.[kaynak belirtilmeli ]
Soyu tükenmiş türlerin yeniden ortaya çıkmasını çevreleyen etik çok tartışmalı. Soyu tükenmiş türleri hayata döndürme eleştirmenleri, sınırlı para ve kaynakları dünyanın mevcut durumundan korumaktan başka yöne çevireceğini iddia ediyor. biyolojik çeşitlilik sorunlar.[29] Arka plan yok olma oranının yaklaşık 100 ila 1.000 katı olan mevcut yok olma oranları ile,[30] Bu türlerin basitçe hayata döndürülebileceğine inanılırsa, bir yok oluştan arındırma programının mevcut kitlesel yok olma kriziyle ilgili halkın endişelerini azaltabileceğinden korkuluyor. Soyu tükenme pozuyla ilgili bir Scientific American makalesinin editörleri olarak: Yünlü mamutu sadece bu arada fillerin neslinin tükenmesine izin vermek için geri mi getirmeliyiz?[29] Bu çağda (MÖ 10.000 sonrası) çoğu türün yok olmasının ana itici faktörü, habitat kaybıdır ve nesli tükenmiş bir türün geçici olarak geri getirilmesi, bir zamanlar yaşadıkları ortamı yeniden yaratmayacaktır.[31]
Yok oluşun savunucuları, örneğin George Kilisesi, birçok potansiyel faydadan bahsedin. Soyu tükenmiş bir kilit taşı türünün yeniden tanıtılması. tüylü mamut, bir zamanlar onlara bağlı olan ekosistemlerin yeniden dengelenmesine yardımcı olabilir. Soyu tükenmiş bazı türler, iade edilirse, bir zamanlar yaşadıkları ortamlar için geniş faydalar sağlayabilir. Örneğin, yünlü mamutlar, Rus ve Kuzey Kutbu tundralarının erimesini, yeni çimenlerin büyüyüp kök salması için ölü ot yemek ve aşağıdaki zemini arktik bölgeye maruz bırakarak periyodik olarak karı kırmak gibi çeşitli yollarla yavaşlatabilir. hava. Bu teknikler, tehdit altındaki bir türdeki genetik çeşitliliği yeniden tanıtmak, hatta hayvanların değişen bir ortamda daha iyi rekabet etmelerine izin vermek için yeni genler ve özellikler getirmek için de kullanılabilir.[32]
Araştırma ve teknoloji
Yeni bir potansiyel örnek bulunduğunda, bilim adamları normalde ilk olarak hücre ve doku korumasını analiz eder. histolojik teknikler ve DNA'nın hayatta kalma koşullarını test edin. Daha sonra aşağıda açıklanan tekniği kullanarak bir DNA örneğini izole etmeye çalışacaklar ve PCR amplifikasyonu DNA'nın test için mevcut DNA miktarını artırmak için. Bu büyütülmüş DNA daha sonra dizilenir. Dizinin organizmanın filogenetik özellikleriyle eşleştiğini doğrulamak için özen gösterilir.[1] Bir organizma öldüğünde, adı verilen bir teknik amino asit yaş tayini organizmayı yaşlandırmak için kullanılabilir. Derecesini denetler rasemizasyon nın-nin aspartik asit, lösin, ve alanin doku içinde. Zaman geçtikçe, D / L oranı ("D" ve "L" birbirlerinin ayna görüntüleridir) 0'dan 1'e yükselir.[33] Aspartik asidin D / L oranının 0.08'den yüksek olduğu örneklerde, eski DNA dizileri alınamamaktadır (1996 itibariyle).[34]
Mitokondriyal DNA ve nükleer DNA
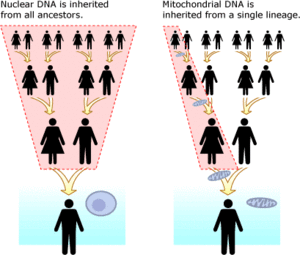
Mitokondriyal DNA (mtDNA) kişinin nükleer DNA'sından ayrıdır. İçinde mevcut organeller aranan mitokondri her birinde hücre. Aksine nükleer DNA Her iki ebeveynden miras kalan ve her nesilde yeniden düzenlenen, mitokondriyal DNA'nın tam bir kopyası anneden oğullarına ve kızlarına aktarılır. Mitokondriyal DNA ile DNA analizi yapmanın faydaları, nükleer DNA'dan çok daha küçük bir mutasyon oranına sahip olması ve on binlerce yıl ölçeğinde soyları takip etmeyi çok daha kolay hale getirmesidir. MtDNA için baz mutasyon oranını bilmek,[35] (insanlarda bu oran aynı zamanda İnsan mitokondriyal moleküler saat ) herhangi iki soyun ne kadar zaman ayrıldığı belirlenebilir. MtDNA'nın bir başka avantajı da, her hücrede binlerce kopyası bulunurken, her hücrede yalnızca iki nükleer DNA kopyası bulunmasıdır.[36] Herşey ökaryotlar tüm bitkileri, hayvanları ve mantarları içeren bir grup, mtDNA'ya sahiptir.[37] MtDNA'nın bir dezavantajı, yalnızca maternal çizginin temsil edilmesidir. Örneğin, bir çocuk DNA'sının 1 / 8'ini sekiz büyük büyükbabasının her birinden miras alacak, ancak anneannesinin mtDNA'sının tam bir klonunu miras alacaktır. Bu, yalnızca babasının büyük büyükbabasının soyadını miras alan bir çocuğa benziyor ve sekiz soyadının tümünün bir karışımı değil.
İzolasyon
Ne zaman düşünülmesi gereken birçok şey var izolasyon bir madde. İlk olarak, ne olduğuna ve nerede bulunduğuna bağlı olarak, kontaminasyonu ve numunenin daha fazla bozulmasını önlemek için gerçekleştirilmesi gereken protokoller vardır.[4] Daha sonra, malzemelerin işlenmesi genellikle fiziksel olarak izole edilmiş bir çalışma alanında ve belirli koşullar altında (yani, belirli Sıcaklık, nem, vb.) Ayrıca kontaminasyonu ve daha fazla numune kaybını önlemek için yapılır.[4]
Materyal elde edildikten sonra, ne olduğuna bağlı olarak, onu izole etmenin ve saflaştırmanın farklı yolları vardır. Fosillerden DNA ekstraksiyonu daha popüler uygulamalardan biridir ve istenen numuneyi elde etmek için atılabilecek farklı adımlar vardır.[4] Amber gömülüden ekstrakte edilen DNA fosiller küçük numunelerden alınarak farklı maddelerle karıştırılabilir, santrifüjlenmiş, inkübe edildi ve tekrar santrifüjlendi.[38] Öte yandan, böceklerden DNA ekstraksiyonu numunenin öğütülmesi, tamponla karıştırılması ve arınma cam elyaf kolonlar aracılığıyla.[39] Sonunda, bu fosiller için numunenin nasıl izole edildiğine bakılmaksızın, izole edilen DNA'nın geçebilmesi gerekir. amplifikasyon.[4][38][39]
Amplifikasyon

Moleküler paleontoloji alanı, polimeraz zincir reaksiyonu (PCR), DNA'nın korunmuş tek bir kopyasından bir DNA parçasının milyarlarca kopyasının yapılmasına izin verir. Bu noktaya kadar en büyük zorluklardan biri, DNA'nın zamanla bozulması nedeniyle geri kazanılan DNA'daki aşırı kıtlıktı.[1]
Sıralama
DNA dizilimi nükleotidlerin ve genlerin sırasını belirlemek için yapılır.[40] DNA'nın çıkarılabileceği birçok farklı malzeme vardır. Hayvanlarda mitokondriyal kromozom, moleküler çalışma için kullanılabilir. Kloroplastlar bitkilerde sekans verilerinin birincil kaynağı olarak incelenebilir.[40]
Sonunda, oluşturulan diziler oluşturmak için kullanılır evrimsel ağaçlar.[40] Veri kümelerini eşleştirme yöntemleri şunları içerir: maksimum olasılık, minimum evrim (Ayrıca şöyle bilinir komşu birleştirme ) en kısa toplam uzunluğa sahip ağacı arayan ve azami cimrilik en az karakter durumu değişikliği gerektiren ağacı bulan yöntem.[40] Bir ağaç içinde tanımlanan tür grupları daha sonra aşağıdaki gibi istatistiksel testlerle de değerlendirilebilir: önyükleme yöntemi, gerçekten önemli olup olmadıklarını görmek için.[40]
Sınırlamalar ve zorluklar
Organizmanın kurutulduğu ve açığa çıkarıldığı DNA'yı korumak için ideal çevresel koşullara ulaşmak ve analize kadar durumlarını korumak zordur. Nükleer DNA normalde ölümden sonra hızla bozulur. endojen hidrolitik süreçler,[34] UV radyasyonu ile,[1] ve diğer çevresel stres faktörleri.
Ayrıca, çevredeki toprağın organik parçalanma ürünleri ile etkileşimlerin biyomoleküler materyallerin korunmasına yardımcı olduğu bulunmuştur.[41] Bununla birlikte, bunlar üzerinde uygun analizi yapabilmek için çeşitli bileşenleri ayırabilmek gibi ek bir zorluk da yarattılar.[42] Bu bozulmalardan bazılarının, PCR sırasında kullanılan bazı enzimlerin faaliyetine müdahale ettiği de bulunmuştur.[41]
Son olarak, antik DNA'nın çıkarılmasındaki en büyük zorluklardan biri, özellikle eski insan DNA'sında, PCR sırasında kontaminasyondur. Küçük miktarlarda insan DNA'sı, eski DNA'nın ekstraksiyonu ve PCR'si için kullanılan reaktifleri kontamine edebilir. Bu sorunlar, tüm çözümlerin yanı sıra işlemde kullanılan cam eşyalar ve diğer araçların kullanımında titiz bir özen gösterilerek aşılabilir. Ayrıca, mevcut farklı DNA türlerini en aza indirmek için ekstraksiyonları yalnızca bir kişi yaparsa yardımcı olabilir.[34]
Ayrıca bakınız
Referanslar
- ^ a b c d e f g Marota, Isolina; Franco Rollo (2002). "Moleküler paleontoloji". Hücresel ve Moleküler Yaşam Bilimleri. 59 (1): 97–111. doi:10.1007 / s00018-002-8408-8. PMID 11846037. S2CID 6377604.
- ^ Higby Schweitzer, Mary (Nisan 2004). "Moleküler paleontoloji: bazı güncel gelişmeler ve sorunlar". Annales de Paléontologie. 90 (2): 81–102. doi:10.1016 / j.annpal.2004.02.001. Alındı 22 Nisan 2014.
- ^ Shapiro, B .; Hofreiter, M. (23 Ocak 2014). "Evrim ve Gen Fonksiyonu Üzerine Paleogenomik Bir Bakış Açısı: Antik DNA'dan Yeni Görüşler". Bilim. 343 (6169): 1236573. doi:10.1126 / science.1236573. PMID 24458647. S2CID 1015148.
- ^ a b c d e Arabacı, Ben. "Moleküler Paleontoloji" (PDF). Nature Publishing Group.
- ^ a b c Abelson, PH (1954). "Fosillerin organik bileşenleri". Carnegie Institute of Washington Yearbook. 53: 97–101.
- ^ Hreha, Sarah. "2013 Gruber Genetics Prize Basın Bülteni". Gruber Vakfı.
- ^ a b de Jong, EW; Westbroek P; Westbroek JF; Bruining JW (1974). "70 Myr üzerinde makromoleküllerin antijenik özelliklerinin korunması". Doğa. 252 (5478): 63–64. Bibcode:1974Natur.252 ... 63D. doi:10.1038 / 252063a0. PMID 4139661. S2CID 4286786.
- ^ a b Westbrock P, van der Meide PH; van der Wey-Kloppers JS; et al. (1979). "Kafadanbacaklı kabuklardan fosil makromoleküller: Karakterizasyon, immünolojik yanıt ve diyajenez". Paleobiyoloji. 5 (2): 151–167. doi:10.1017 / S0094837300006448.
- ^ a b Nitecki MH (1982). Evrimsel Biyolojinin Biyokimyasal Yönleri. Chicago: Chicago Press Üniversitesi. s. 29–91.
- ^ a b Gurley, L.R; J. G. Valdez; W. D. Spall; B. F Smith; D. D. Gillette (Şubat 1991). "Dinozorun fosil kemiğindeki proteinler, seismosaurus". Protein Kimyası Dergisi. 10 (1): 75–90. doi:10.1007 / BF01024658. PMID 2054066. S2CID 9046093.
- ^ a b Kaiser, J. (7 Ekim 2005). "VİROLOJİ: Dirilen Grip Virüsü, Ölümcül 1918 Salgınının Sırlarını Veriyor" (PDF). Bilim. 310 (5745): 28–29. doi:10.1126 / science.310.5745.28. PMID 16210501. S2CID 26252589. Alındı 9 Nisan 2014.
- ^ a b Rockefeller Üniversitesi (1 Mart 2007). "Antik Retrovirüs Dirildi". Günlük Bilim. Alındı 9 Nisan 2014.
- ^ a b Krause, Johannes; Fu, Qiaomei; Güzel, Jeffrey M .; Viola, Bence; Shunkov, Michael V .; Derevianko, Anatoli P .; Pääbo, Svante (24 Mart 2010). "Güney Sibirya'dan bilinmeyen bir homininin tam mitokondriyal DNA genomu". Doğa. 464 (7290): 894–897. Bibcode:2010Natur.464..894K. doi:10.1038 / nature08976. PMID 20336068.
- ^ a b c Prüfer K, Racimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, Heinze A, Renaud G, Sudmant PH, de Filippo C, Li H, Mallick S, Dannemann M, Fu Q, Kircher M, Kuhlwilm M, Lachmann M, Meyer M, Ongyerth M, Siebauer M, Theunert C, Tandon A, Moorjani P, Pickrell J, Mullikin JC, Vohr SH, Green RE, Hellmann I, Johnson PL, Blanche H, Cann H, Kitzman JO, Shendure J, Eichler EE, Lein ES, Bakken TE, Golovanova LV, Doronichev VB, Shunkov MV, Derevianko AP, Viola B, Slatkin M, Reich D, Kelso J, Pääbo S (18 Aralık 2013). "Altay Dağları'ndan bir Neandertal'in tam genom dizisi". Doğa. 505 (7481): 43–49. Bibcode:2014Natur.505 ... 43P. doi:10.1038 / nature12886. PMC 4031459. PMID 24352235.
- ^ a b c Callaway, Ewen (4 Aralık 2013). "Hominin DNA'sı uzmanları şaşırtıyor". Doğa. 504 (7478): 16–17. Bibcode:2013Natur.504 ... 16C. doi:10.1038 / 504016a. PMID 24305130.
- ^ a b Dalton, Rex (15 Mayıs 2006). "Neandertal DNA'sı genom dalgalanmasına yol açar". Doğa. 441 (7091): 260–1. Bibcode:2006Natur.441..260D. doi:10.1038 / 441260b. PMID 16710377.
- ^ a b Zimmer, Carl (16 Kasım 2015). "Bir Dişte, Bazı Çok Eski Kuzenlerin DNA'sı, Denisovalılar". New York Times. Alındı 16 Kasım 2015.
- ^ a b Sawyer, Susanna; Renaud, Gabriel; Viola, Bence; Hublin, Jean-Jacques; Gansauge, Marie-Theres; Shunkov, Michael V .; Derevianko, Anatoly P .; Prüfer, Kay; Kelso, Janet; Pääbo, Svante (11 Kasım 2015). "İki Denisovalı bireyden nükleer ve mitokondriyal DNA dizileri". PNAS. 112 (51): 15696–700. Bibcode:2015PNAS..11215696S. doi:10.1073 / pnas.1519905112. PMC 4697428. PMID 26630009.
- ^ Higuchi, Russell; Bowman, Barbara; Freiberger, Mary; Ryder, Oliver A .; Wilson, Allan C. (15 Kasım 1984). "At ailesinin soyu tükenmiş bir üyesi olan quagga'dan DNA dizileri". Doğa. 312 (5991): 282–284. Bibcode:1984Natur.312..282H. doi:10.1038 / 312282a0. PMID 6504142. S2CID 4313241.
- ^ Krause, Johannes; Fu, Qiaomei; Güzel, Jeffrey M .; Viola, Bence; Shunkov, Michael V .; Derevianko, Anatoli P .; Pääbo, Svante (24 Mart 2010). "Güney Sibirya'dan bilinmeyen bir homininin tam mitokondriyal DNA genomu". Doğa. 464 (7290): 894–897. Bibcode:2010Natur.464..894K. doi:10.1038 / nature08976. PMID 20336068.
- ^ a b Reich D, Green RE, Kircher M, Krause J, Patterson N, Durand EY, Viola B, Briggs AW, Stenzel U, Johnson PL, Maricic T, Good JM, Marques-Bonet T, Alkan C, Fu Q, Mallick S, Li H, Meyer M, Eichler EE, Stoneking M, Richards M, Talamo S, Shunkov MV, Derevianko AP, Hublin JJ, Kelso J, Slatkin M, Pääbo S (23 Aralık 2010). "Sibirya'daki Denisova Mağarasından bir arkaik hominin grubunun genetik geçmişi". Doğa. 468 (7327): 1053–1060. Bibcode:2010Natur.468.1053R. doi:10.1038 / nature09710. PMC 4306417. PMID 21179161.
- ^ Pennisi, E. (16 Mayıs 2013). "Denisova Mağarasından Daha Fazla Genom, İlk İnsan Gruplarının Karışımını Gösteriyor". Bilim. 340 (6134): 799. Bibcode:2013Sci ... 340..799P. doi:10.1126 / science.340.6134.799. PMID 23687020.
- ^ Mounier, Aurélien; Marchal, François; Condemi, Silvana (Mart 2009). "Homo heidelbergensis ayrı bir tür mü? Mauer mandibulası hakkında yeni bilgiler". İnsan Evrimi Dergisi. 56 (3): 219–246. doi:10.1016 / j.jhevol.2008.12.006. PMID 19249816.
- ^ Cartmill, Matt ve Smith, Fred H. (2009). İnsan Soyu. John Wiley & Sons. ISBN 978-0471214915. Alındı 2013-04-21.
- ^ a b Meyer, Matthias; Fu, Qiaomei; Aximu-Petri, Ayinuer; Glocke, Isabelle; Nikel, Birgit; Arsuaga, Juan-Luis; Martínez, Ignacio; Gracia, Ana; de Castro, José María Bermúdez; Carbonell, Eudald; Pääbo, Svante (4 Aralık 2013). "Sima de los Huesos'tan bir homininin mitokondriyal genom dizisi" (PDF). Doğa. 505 (7483): 403–406. Bibcode:2014Natur.505..403M. doi:10.1038 / nature12788. PMID 24305051. S2CID 4456221. Arşivlenen orijinal (PDF) 12 Şubat 2014. Alındı 23 Nisan 2014.
- ^ Zimmer, Carl. "Soyu Tükenmiş Türleri Hayata Döndürmek". National Geographic. Alındı 26 Mart 2014.
- ^ Gray, Richard (31 Ocak 2009). "Soyu tükenmiş dağ keçisi klonlanarak yeniden dirildi". Telgraf. Alındı 9 Nisan 2014.
- ^ Kilise, George. "Multiplex Automated Genomic Engineering (MAGE): Evrimi hızlandıran bir makine, genom tasarımında devrim yaratıyor". Wyss Enstitüsü. Alındı 26 Mart 2014.
- ^ a b Editörler (14 Mayıs 2013). "Soyu Tükenmiş Türleri Ölülerden Geri Getirme Çabaları Neden Noktayı Kaçırıyor?". Bilimsel amerikalı. Alındı 9 Nisan 2014.
- ^ Pimm, S. L .; Russell, G. J .; Gittleman, J. L .; Brooks, T.M. (21 Temmuz 1995). "Biyoçeşitliliğin Geleceği" (PDF). Bilim. 269 (5222): 347–350. Bibcode:1995Sci ... 269..347P. doi:10.1126 / science.269.5222.347. PMID 17841251. S2CID 35154695. Alındı 9 Nisan 2014.
- ^ Hogan, Michael. "Yok Olmanın Nedenleri". Dünya Ansiklopedisi. Alındı 9 Nisan 2014.
- ^ Kilise, George (20 Ağustos 2013). "Yok Olmak İyi Bir Fikirdir". Bilimsel amerikalı. Alındı 9 Nisan 2014.
- ^ "Yöntem: İlkeler". Amino Asit Jeokronoloji Laboratuvarı. Kuzey Arizona Üniversitesi. Arşivlenen orijinal 14 Mart 2012 tarihinde. Alındı 26 Mart 2014.
- ^ a b c Poinar, H. N .; Hoss, M .; Bada, J. L .; Paabo, S. (10 Mayıs 1996). "Amino Asit Rasemizasyonu ve Antik DNA'nın Korunması". Bilim. 272 (5263): 864–866. Bibcode:1996Sci ... 272..864P. doi:10.1126 / science.272.5263.864. hdl:2060/19980202342. PMID 8629020. S2CID 20626782.
- ^ Henn, B. M .; Gignoux, C. R .; Feldman, M. W .; Mountain, J.L. (6 Ekim 2008). "İnsan Mitokondriyal DNA Mutasyon Hızı Tahminlerinin Zaman Bağımlılığını Karakterize Etme". Moleküler Biyoloji ve Evrim. 26 (1): 217–230. doi:10.1093 / molbev / msn244. PMID 18984905.
- ^ "Mitokondriyal DNA". DNA Analisti Eğitimi. Adalete Hizmet Eden NFSTC Bilim. Arşivlenen orijinal 1 Şubat 2014. Alındı 23 Nisan 2014.
- ^ Lang, BF; Gri, MW; Burger, G (1999). "Mitokondriyal genom evrimi ve ökaryotların kökeni". Genetik Yıllık İnceleme. 33: 351–97. doi:10.1146 / annurev.genet.33.1.351. PMID 10690412.
- ^ a b Cano, RJ; Poinar, HN (Eylül 1993). "PCR için uygun fosil ve müze örneklerinden DNA'nın hızlı izolasyonu". BioTeknikler. 15 (3): 432–4, 436. PMID 8217155.
- ^ a b Tagliavia, Marcello; Massa, Bruno; Arnavutça, Ida; La Farina, Mario (29 Mart 2011). "Orthoptera Müzesi Örneklerinden DNA Ekstraksiyonu" (PDF). Analitik Mektuplar. 44 (6): 1058–1062. doi:10.1080/00032719.2010.506939. hdl:10447/57711. S2CID 95380144.
- ^ a b c d e S. Blair Hodges; Mary H. Schweitzer. Moleküler Paleontoloji (PDF). Paleontoloji Ansiklopedisi. s. 752–754.
- ^ a b Tuross, N (1994). "Kemikteki eski DNA'nın biyokimyası". Experientia. 50 (6): 50:530–535. doi:10.1007 / bf01921721. PMID 7517371. S2CID 27896535.
- ^ Tuross, N; Stathoplos, L (1993). Fosil kemiklerdeki antik proteinler. Enzimolojide Yöntemler. 224. s. 121–129. doi:10.1016 / 0076-6879 (93) 24010-r. ISBN 9780121821258. PMID 8264383.
| Tarih | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Metodoloji | |||||||||
| Alt disiplinler |
| ||||||||
| |||||||||
| Anahtar bileşenler | |
|---|---|
| Alanlar | |
| Arkeogenetik nın-nin | |
| İlgili konular | |
| Listeler | |
| |