Paraceratherium - Paraceratherium
| Paraceratherium | |
|---|---|
 | |
| İskelet döküm P. transouralicum, Ulusal Doğa ve Bilim Müzesi, Tokyo | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Sınıf: | Memeli |
| Sipariş: | Perissodactyla |
| Aile: | †Hyracodontidae |
| Alt aile: | †Indricotheriinae |
| Cins: | †Paraceratherium Forster-Cooper, 1911 |
| Türler | |
| †Aceratherium bugtiense Hacı, 1908 | |
| Türler | |
| |
| Eş anlamlı | |
Cins eşanlamlı
Tür eşanlamlı
| |
Paraceratherium soyu tükenmiş cins boynuzsuz gergedan. Biridir en büyük karasal memeliler var olan ve erken dönemden geçe kadar yaşayan Oligosen dönem (34–23 milyon yıl önce). Kalıntıları boyunca bulundu Avrasya Çin ile Balkanlar. Üye olarak sınıflandırılır Hyracodont alt aile Indricotheriinae. Paraceratherium "boynuzsuz canavarın yanında" anlamına gelir. Aceratherium, içinde bulunduğu cins türler A. bugtiense başlangıçta yerleştirildi.
Tam boyutu Paraceratherium eksikliğinden dolayı bilinmiyor fosiller. Omuz yüksekliği yaklaşık 4.8 metre (15.7 fit) ve uzunluk yaklaşık 7.4 metredir (24.3 fit). Ağırlığının yaklaşık 15 ila 20 ton (33.000 ila 44.000 lb) olduğu tahmin edilmektedir. Uzun boyun, yaklaşık 1.3 metre (4.3 ft) uzunluğundaki bir kafatasını destekledi. Geniş, diş gibiydi kesici dişler ve bir burun kesiği olduğunu düşündüren bir burun kesiği kavrayıcı üst dudak veya hortum (gövde). Bacaklar uzun ve sütun gibiydi. Yaşam tarzı Paraceratherium modern büyük memelilerinkine benzer olabilir. filler ve mevcut gergedanlar. Boyutu nedeniyle çok az avcılar ve yavaş bir üreme hızı. O bir tarayıcı, çoğunlukla yaprakları, yumuşak bitkileri ve çalıları yer. Birkaç dağınık ağaçlı kurak çöllerden subtropikal ormanlara kadar değişen habitatlarda yaşadı. Hayvanın neslinin tükenmesinin nedenleri bilinmemektedir, ancak çeşitli faktörler öne sürülmüştür.
taksonomi cinsinin ve içindeki türlerin uzun ve karmaşık bir geçmişi vardır. Diğer Oligosen indricotheres cinsleri, örneğin Baluchitherium, Indrikoterium, ve Pristinotherium, adlandırılmıştır, ancak tam örnek yoktur, bu da karşılaştırma ve sınıflandırmayı zorlaştırır. Çoğu modern bilim adamı, bu cinsin küçük eş anlamlılar nın-nin Paraceratheriumve en az üç ayırt edilebilir tür içerdiği düşünülmektedir; P. bugtiense, P. transouralicum, ve P. huangheense. En çok bilinen tür P. transouralicum, bu yüzden cinsin çoğu rekonstrüksiyonu buna dayanmaktadır. Arasındaki farklar P. bugtiense ve P. transouralicum nedeniyle olabilir cinsel dimorfizm, bu da onları aynı tür yapar.
Taksonomi
taksonomik tarihi Paraceratherium bilinen fosillerin parçalı doğası nedeniyle karmaşıktır ve Batılı, Sovyet ve Çinli bilim adamları 20. yüzyılın büyük bir bölümünde birbirlerinden izole olarak çalıştılar ve araştırmaları esas olarak kendi dillerinde yayınladılar.[1] Dünyanın farklı yerlerinden bilim adamları, bu hayvanların daha eksiksiz bir resmini elde etmek için bulduklarını karşılaştırmaya çalıştılar, ancak siyaset ve savaşlar tarafından engellendi.[2] Karşıt taksonomik eğilimler "topaklanma ve bölme "ayrıca soruna katkıda bulundu.[3] Yanlış jeolojik tarihleme önceden bilim adamlarının çeşitli jeolojik oluşumlar şimdi çağdaş olduğu bilinen farklı yaşlardaydı. Birçok cins, içindeki ince farklılıklar temelinde adlandırılmıştır. azı dişi diş özellikleri - diğer gergedanların popülasyonları içinde değişen özellikler takson - ve bu nedenle çoğu bilim adamı tarafından türleri ayırt etmeleri nedeniyle kabul edilmemektedir.[4]
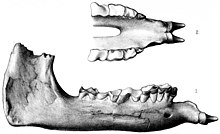
Erken keşifler Indricotheres Asya'ya çeşitli kolonyal bağlantılar aracılığıyla yapıldı.[2] Bilinen ilk indriko fosilleri Belucistan (günümüz Pakistan'ında) 1846'da Vickary adlı bir asker tarafından yapıldı, ancak bu parçalar o zamanlar tanımlanamıyordu.[5] İlk fosiller artık şu şekilde kabul edilmektedir: Paraceratherium İngiliz jeolog tarafından keşfedildi Guy Ellcock Pilgrim Belucistan'da 1907-1908'de. Materyalleri bir üst çene, alt dişler ve bir çenenin arkasından oluşuyordu. Fosiller, Chitarwata Formasyonu nın-nin Dera Bugti Pilgrim'in daha önce araştırdığı yer. 1908'de, soyu tükenmiş gergedan cinsinin yeni bir türü için fosilleri temel aldı. Aceratherium; A. bugtiense. Aceratherium o zamana kadar çöp sepeti taksonu; birçok alakasız boynuzsuz gergedan türünü içeriyordu ve bunların çoğu o zamandan beri diğer cinslere aktarıldı.[1][6] Fosil kesici dişler Pilgrim'in daha önce ilgisiz cinse tahsis ettiği Bugtitherium daha sonra yeni türe ait olduğu gösterildi.[7]
1910'da, İngiliz paleontolog tarafından yapılan bir keşif gezisi sırasında Dera Bugti'de daha fazla kısmi fosil bulundu. Clive Forster-Cooper. Bu kalıntılara dayanarak, Foster-Cooper taşındı A. bugtiense yeni cinse Paraceratherium, "boynuzsuz canavarın yanında" anlamına gelir. Aceratherium.[1][8] Bu yeniden sınıflandırmanın mantığı, türün belirgin bir şekilde aşağıya çevrilmiş dişleriydi.[7] 1913'te Forster-Cooper yeni bir cins ve tür seçti. Thaumastotherium ("harika canavar") Osborni, aynı kazılardan elde edilen daha büyük fosillere dayanmaktadır (daha önce bazılarının erkeklere ait olduğunu öne sürmüştü. P. bugtiense), ancak cinsi yeniden adlandırdı Baluchitherium o yıl daha sonra çünkü eski adı meşgul, zaten bir hemipteran böcek.[9][10][8] Fosilleri Baluchitherium o kadar parçalandı ki, Foster-Cooper bunu yalnızca bir tür garip toynaklı, ama karışıklık olasılığından bahsetti Paraceratherium.[11] Amerikalı paleontolog Henry Fairfield Osborn, daha sonra B. osborni adlandırıldı, önerildi Titanothere.[2]
Bir Rusya Bilimler Akademisi keşif daha sonra fosiller buldu Aral Oluşumu yakınında Aral denizi Kazakistan'da; o bilinen en eksiksiz indriko iskeletiydi, ancak kafatasından yoksundu. 1916'da bu kalıntılara dayanarak, Aleksei Alekseeivich Borissiak cinsi dikti. Indrikoterium mitolojik bir canavarın adı, "Indrik canavar ". Bir tür adı vermedi, I. asiaticum, 1923'e kadar, ancak Rus paleontolog Maria Pavlova zaten adlandırmıştı I. transouralicum 1922'de.[1][12] Ayrıca 1923'te Borissiak, o zamana kadar bilinen çeşitli ilgili formları içerecek şekilde Indricotheriinae alt ailesini yarattı.[13]
1922'de Amerikalı kaşif Roy Chapman Andrews Çin'e iyi belgelenmiş bir seferi yönetti ve Moğolistan sponsorluğunda Amerikan Doğa Tarihi Müzesi. Moğol oluşumlarında çeşitli indriko kalıntıları bulundu. Gobi Çölü dik pozisyonda duran bir numunenin bacakları dahil, kapana kısılmışken öldüğünü gösterir. bataklık hem de çok eksiksiz bir kafatası. Bu kalıntıların temeli oldu Baluchitherium grangeri, 1923'te Osborn tarafından seçildi.[14][15]
2017'de yeni bir tür, P. huangheense, Çinli paleontolog Yong-Xiang Li ve meslektaşları tarafından Hanjiajing Formasyonundaki çene elemanlarına dayanarak seçildi. Gansu Eyaleti Çin'in; isim yakındaki anlamına gelir Huanghe Nehri.[16] Çoğunlukla büyüklük, burun şekli ve ön diş dizilişindeki farklılıklara dayanan çok sayıda başka tür ve cins isimleri, çeşitli indrikotere kalıntıları için icat edilmiştir. Atfedilebilir fosiller Paraceratherium Avrasya genelinde keşfedilmeye devam ediyor, ancak Pakistan'daki siyasi durum, orada daha fazla kazı yapılamayacak kadar istikrarsız hale geldi.[5]
Türler ve eşanlamlılar
1922'de Forster-Cooper yeni türe isim verdi Metamynodon bugtiensis Dera Bugti'den bir damak ve diğer parçalara dayanarak, bu cinsin dev bir üyesine ait olduğu sanılıyor. Bu fosillerin artık bir sapkınlığa ait olduğu düşünülüyor. Paraceratherium bugtiense M3 azı dişinden yoksun örnek.[17][18] 1936'da Amerikalı paleontologlar Walter Granger ve William K. Gregory Forster-Cooper'ın Baluchitherium osborni muhtemelen bir küçük eşanlamlı (aynı takson için geçersiz bir ad) Paraceratherium bugtienseçünkü bu örnekler aynı yerde toplanmış ve muhtemelen aynı morfolojik olarak değişken türler.[19] Amerikalı paleontolog William Diller Matthew ve Forster-Cooper'ın kendisi de benzer şüpheleri birkaç yıl önce ifade etmişti. Zaten küçük eşanlamlı olarak ilan edilmiş olmasına rağmen, cins adı Baluchitherium Osborn'u çevreleyen tanıtım nedeniyle çeşitli medyada popüler kaldı. B. grangeri.[3][20]

1989'da Amerikalı paleontologlar Spencer G. Lucas ve Jay C. Sobus indricothere taksonlarının bir revizyonunu yayınladı ve bunu bugün çoğu batılı bilim insanı izledi. Sonucuna vardılar Paraceratheriumen eski isim olarak, Oligosen'den gelen tek geçerli indriko cinsiydi ve dört geçerli tür içeriyordu, P. bugtiense, P. transouralicum (başlangıçta Indrikoterium), P. prohorovi (başlangıçta Aralotherium ), ve P. orgosensis (başlangıçta Dzungariotherium ). Diğer isimlerin çoğunun bu taksonların küçük eşanlamlıları olduğunu veya şüpheli isimler, doğru bir şekilde tanımlanamayacak kadar parçalı kalıntılara dayanıyor. Lucas ve Sobus, adlandırılmış cinsler ve türler arasındaki iddia edilen farklılıkları analiz ederek, bunların büyük olasılıkla popülasyonlar içindeki varyasyonu temsil ettiğini ve 1930'larda işaret edildiği gibi çoğu özelliğin örnekler arasında ayırt edilemez olduğunu buldular. Tek kafatasının atanmış olması P. transouralicum veya Indrikoterium kubbelidir, diğerleri ise tepede düzdür. cinsel dimorfizm; bu mümkündür P. bugtiense fosiller dişiyi temsil ederken P. transouralicum aynı türün erkeğini temsil eder.[3][21][22]
Lucas ve Sobus'a göre, türler P. bugtiense Pakistan'ın geç Oligoseninden itibaren aşağıdaki gibi küçük eş anlamlılar içerir B. osborni ve P. zhajremensis. P. transouralicum Geç Oligosen, Kazakistan, Moğolistan ve Kuzey Çin'den B. grangeri ve I. eksi.[21] 2013 yılında Amerikalı paleontolog Donald Prothero bunu önerdi P. orgosensis orijinal cins adını garanti edecek kadar farklı olabilir Dzungariotherium, tam konumu değerlendirme gerektirse de. P. prohorovi Kazakistan Oligosen sonundan itibaren konumu diğer türlere göre çözümlenemeyecek kadar eksik olabilir; aynı şey önerilen türler için de geçerlidir. I. ara ve P. tienshanensisyanı sıra cins Benaratherium.[3][21] Cins adı olmasına rağmen Indrikoterium artık küçük eşanlamlı Paraceratherium, Indricotheriinae alt aile adı hala kullanımdadır çünkü cins adı eşanlamlılığı, bunlardan türetilen daha yüksek seviyeli taksonların adlarını etkilemez. Alt ailenin üyelerine bu nedenle hala yaygın olarak indrikoterler olarak atıfta bulunulmaktadır.[23]
Lucas ve Sobus'un revizyonunun aksine, Çinli paleontolog Jie Ye ve meslektaşlarının 2003 tarihli bir makalesi Indrikoterium ve Dzungariotherium geçerli cinslerdi ve bu P. prohorovi ait değildi Paraceratherium. Ayrıca türlerin geçerliliğini de kabul ettiler. P. lipidus, P. tienshanensis, ve P. sui.[24] Çinli paleontolog tarafından hazırlanan 2004 tarihli bir makale Tao Deng ve meslektaşları ayrıca üç farklı cinsi tanıdı.[25] Bazı Batılı yazarlar benzer şekilde 1989 revizyonundan bu yana geçersiz kabul edilen isimleri kullandılar, ancak ayrıntılı analiz ve gerekçe sunmadan.[3]
Evrim

üst aile Rhinocerotoidea modern gergedanları içeren, geriye doğru erken Eosen —Yaklaşık 50 milyon yıl önce — gibi erken öncüllerle Hyrachyus. Rhinocerotoidea üç aile içerir; Amynodontidae, Rhinocerotidae ("gerçek gergedanlar") ve Hyracodontidae. Gergedan grubu içindeki çeşitlilik tarih öncesi çağlarda çok daha fazlaydı; köpek büyüklüğünden köpek boyutuna kadar değişiyorlardı. Paraceratherium. Uzun bacaklı vardı cursorial koşma ve çömelme için uyarlanmış formlar, yarı suda yaşayan formlar. Çoğu türün boynuzları yoktu. Gergedan fosilleri, esas olarak, hayvanların en çok korunma ihtimali olan parçası olan dişlerinin özelliklerine göre tanımlanır. Çoğu gergedanın üst azı dişlerinde pi Taçta (π) şekilli (π) desen ve her bir alt azı dişi eşleştirilmiş L şeklindedir. Fosil gergedanların tanımlanmasında çeşitli kafatası özellikleri de kullanılmaktadır.[26]
Indricotheriinae alt ailesi, Paraceratherium aittir, ilk olarak Amerikalı paleontolog tarafından Hyracodontidae ailesinin bir parçası olarak sınıflandırılmıştır. Leonard B. Radinsky Daha önce, Rhinocerotidea'da bir alt aile, hatta tam bir aile olan Indricotheriidae olarak görülüyorlardı.[27] 1999'da kladistik çalışma tapiromorflar Amerikalı paleontolog Luke Holbrook, indricother'leri hyracodontid'in dışında buldu. clade ve onların olmayabileceğini yazdı monofiletik (doğal) gruplama.[28] Radinsky'nin planı, bugün yaygın olan hipotezdir. Hyracodont ailesi, koşmaya uyarlanmış uzun bacaklı üyeler içerir. Hyracodon, ve kesici özellikleriyle ayırt edildi. Indricotheres, diğer hyracodonlardan daha büyük boyutları ve türetilmiş burunlarının, kesici dişlerinin ve köpek dişlerinin yapısı. Bilinen en eski indrico, köpek büyüklüğündedir. Forstercooperia ortasından ve geç Eosen Batı Kuzey Amerika ve Asya. İnek boyunda Juxia Orta Eosenden bilinmektedir; Eosen sonlarında cins Urtinoterium Asya'nın büyüklüğüne neredeyse ulaştı Paraceratherium.[21][26] Paraceratherium kendisi Avrasya'da yaşadı Oligosen 23 ila 34 milyon yıl önce.[29] Cins, büyük boyutu, kaslı bir burnu destekleyebilecek burun kesiği ve aşağı dönük olmasıyla diğer indrikoterlerden ayrılır. premaksilla.[21] Ayrıca ikinci ve üçüncü alt kesici dişleri, alt köpek dişlerini ve birinci küçük azı dişleri.[26]

kladogram Aşağıda Lucas ve Sobus tarafından Indricotheriinae'nin 1989 analizini izler ve en yakın akrabalarını gösterir. Paraceratherium:[21]
| Hyracodontidae |
| ||||||||||||||||||||||||
Lucas ve meslektaşları, 1981'de yapılan önceki bir analizde benzer sonuçlara ulaşmıştı. Forstercooperia, burada hala tuttular Paraceratherium ve Indrikoterium ayrı cins olarak.[30] 2016 yılında Çinli araştırmacılar Haibing Wang ve meslektaşları, aile için Paraceratheriidae ve alt aile için paraceratheriine adını kullandılar ve onları Hyracodontidae'nin dışına yerleştirdiler.[31]
Açıklama

Paraceratherium biridir bilinen en büyük kara memelileri şimdiye kadar var olmuş, ancak tam örneklerin olmaması nedeniyle kesin boyutu belirsizdir.[4] Toplam vücut uzunluğunun 1936'da Granger ve Gregory tarafından önden arkaya 8,7 m (28,5 ft) ve Sovyet paleontoloğu tarafından 7,4 m (24,3 ft) olduğu tahmin edildi. Vera Gromova 1959'da, ancak önceki tahmin artık abartılı kabul ediliyor. Ağırlığı Paraceratherium bazı soyu tükenmiş olanlara benziyordu hortumlular bilinen en büyük tam iskelete sahip olan bozkır mamut (Mammuthus trogontherii).[32][33] Kabaca eşdeğer kütlesine rağmen, Paraceratherium herhangi bir hortumdan daha uzun olabilirdi.[4] Omuz yüksekliği, Granger ve Gregory tarafından omuzlarda 5,25 m (17,2 ft), ancak Amerikalı paleontolog tarafından 4,8 m (15,7 ft) olarak tahmin edilmiştir. Gregory S. Paul 1997'de.[34] Boyun paleontologlar tarafından 2 ila 2,5 m (6,6 ila 8,2 ft) uzunluğunda olduğu tahmin edildi. Michael P. Taylor ve 2013'te Mathew J. Wedel.[35]

30 tonluk (66.000 lb) ilk tahminler artık abartılı kabul ediliyor; maksimum 15 ila 20 ton (33.000 ila 44.000 lb) aralığında ve ortalama olarak 11 ton (24.000 lb) kadar düşük olabilir. Hesaplamalar esas olarak şu fosillere dayanmaktadır: P. transouralicum çünkü bu tür en eksiksiz kalıntılardan bilinmektedir.[4] Tahminler kafatası, dişler ve uzuv kemiği ölçümlerine dayanmaktadır, ancak bilinen kemik elemanları farklı boyutlardaki bireyler tarafından temsil edilmektedir, bu nedenle tüm iskelet rekonstrüksiyonları birkaç ağırlık aralığıyla sonuçlanan kompozit ekstrapolasyonlardır.[32][36]
Hayvanın rengine ve deri dokusuna dair hiçbir belirti yoktur çünkü deri izleri veya mumyalar bilinmemektedir. Çoğu yaşam restorasyonu, yaratığın derisini modern gergedanlara göre kalın, katlanmış, gri ve tüysüz olarak gösterir. Saç vücut ısısını koruduğu için, filler ve gergedanlar gibi modern büyük memeliler büyük ölçüde tüysüzdür. Prothero, çoğu tasvirin aksine, Paraceratherium büyük fil benzeri kulakları vardı. termoregülasyon. Fillerin kulakları vücudun yüzey alanını genişletir ve kan damarlarıyla doludur, bu da aşırı ısının daha kolay atılmasını sağlar. Prothero'ya göre, bu doğru olurdu Paraceratheriumkulak açıklıklarının etrafındaki sağlam kemiklerin gösterdiği gibi.[4] Paleontologlar Pierre-Olivier Antoine ve Darren Naish bu fikre karşı şüphelerinizi dile getirdi.[37][38]
Kafatası
En büyük kafatasları Paraceratherium kafatasının arkasında yaklaşık 1,3 metre (4,3 ft) uzunluğunda, 33 ila 38 santimetre (13 ila 15 inç) ve boyunda 61 santimetre (24 inç) genişliğindedir. zigomatik kemerler. Paraceratherium düz ve diğer gergedanların boynuzları için bağlantı noktası görevi gören pürüzlü bölgeden yoksun uzun bir alnı vardı. Burun bölgesinin üstündeki kemikler uzundur ve burun kesiği kafatasına kadar uzanır. Bu şunu gösterir Paraceratherium vardı kavrayıcı üst dudak siyah gergedan ve Hintli gergedan veya kısa hortum (gövde) olduğu gibi tapirler. Kafatasının arkası alçak ve dardı, büyük olmadan Lambdoid tepede ve boyunca sırtlar sagital kret, aksi takdirde itmek ve savaşmak için güçlü kaslara ihtiyaç duyan boynuzlu ve dişli hayvanlarda bulunur. Ayrıca kafatasını otomatik olarak tutan ense bağlarının tutturulması için derin bir çukur vardı. oksipital kondil çok genişti ve Paraceratherium Dallardan yiyecek ararken başını güçlü bir şekilde aşağı doğru kaydırmasına izin veren büyük, güçlü boyun kaslarına sahipmiş gibi görünüyor.[4] Bir kafatası P. transouralicum kubbeli bir alnı vardır, diğerleri ise muhtemelen cinsel dimorfizm nedeniyle düz alınlara sahiptir.[21] Beyin endocast nın-nin P. transouralicum Kızılderili gergedanının beyninin kafatası uzunluğunun yüzde 17,7'sini oluştururken kafatası uzunluğunun sadece yüzde 8'i olduğunu gösteriyor.[19]
Türleri Paraceratherium kafatası özellikleriyle ayırt edilebilir. P. bugtiense nispeten ince gibi özelliklere sahipti üst çene ve premaksilla, sığ kafatası çatıları, nispeten ince olan ve kafatasına geri yerleştirilen mastoid-paroksipital süreçler, daha az geriye uzanan lambdoid kret ve yatay yönelimli oksipital kondil ile paylaştığı Dzungariotherium. P. transouralicum sağlam üst çene ve premaksillalara sahipti, kalkık zigomata, kubbeli ön kemikler, kalın mastoid-paroksipital süreçler, geriye doğru uzanan lambdoid kret ve dikey yönelimle oksipital kondiller.[3] P. huangheense farklı P. bugtiense sadece çenenin arka kısmının anatomisinde ve daha büyük boyutunda.[16]
Çoğu ilkel gergedanların aksine, ön dişleri Paraceratherium her iki çenede de büyük ve konik olan ve dişler olarak tanımlanan tek bir kesici dişe indirgenmiştir. Üst kesici dişler aşağıya dönüktü; alt olanlar daha kısaydı ve ileriyi gösteriyordu. Bilinen gergedanlar arasında bu düzenleme benzersizdir. Paraceratherium ve ilgili Urtinoterium. Erkeklerde kesici dişler daha büyük olabilir. köpek dişi aksi takdirde kesici dişlerin arkasında bulundu. Kesici dişler yanak dişlerinden büyük bir diastema (boşluk).[4] Bu özellik, kesici dişlerin ve yanak dişlerinin farklı uzmanlıklara sahip olduğu memelilerde bulunur.[26] Üst azı dişleri, V şeklindeki üçüncü üst azı dişleri hariç, pi şeklinde (π) bir desene ve küçültülmüş bir metastil. Küçük azı dişleri pi modelini sadece kısmen oluşturdu. Her bir azı dişi bir insan yumruğu büyüklüğündeydi; memeliler arasında, kafatasının boyutuna göre küçük olmalarına rağmen, sadece hortumlu hayvanlar tarafından aşılmışlardı. Alt yanak dişleri, gergedanların tipik özelliği olan L şeklindedir.[4]
Postkraniyal iskelet
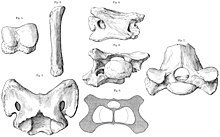
Tam set yok omur ve kaburga Paraceratherium henüz bulundu ve kuyruk tamamen bilinmiyor. Atlas ve eksen Boynun omurları, modern gergedanların çoğundan daha genişti ve büyük kafayı tutmak için gerekli olan güçlü bağlar ve kaslar için alan vardı. Omurların geri kalanı da çok genişti ve büyüktü zigapofizler Baş, boyun ve omurgayı desteklemek için kaslar, tendonlar, bağlar ve sinirler için çok yer vardır. sinir dikenleri uzundu ve sırt boyunca boyun kaslarının ve kafatasını tutmak için ense bağlarının bağlandığı uzun bir "tümsek" oluşturdu. Kaburgalar modern gergedanlara benziyordu, ancak göğüs kafesi uzun bacaklar ve büyük bedenlere oranla daha küçük görünebilirdi çünkü modern gergedanlar nispeten kısa uzuvludur. Alt sırtın son omuru, sakrum, ileri gergedanlarda bulunan bir özellik.[4] Sevmek Sauropod dinozorları, Paraceratherium vardı pleurocoel sakral öncesi omurlarında muhtemelen iskeletin hafiflemesine yardımcı olan açıklıklar (kemiğin içi boş kısımları).[39]

Uzuvlar, hayvanın büyük ağırlığını desteklemek için büyük ve sağlamdı ve bazı açılardan ve yakınsak aynı şekilde graviportal (ağır ve yavaş hareket eden) yapılarına sahip filler ve sauropod dinozorlarınkilerle. Alt ekstremite, el ve ayak kemiklerini kısaltırken, kaynaştırırken ve sıkıştırırken üst ekstremite kemiklerini uzatma eğiliminde olan bu tür hayvanların aksine, Paraceratherium kısa üst ekstremite kemiklerine ve uzun el ve ayak kemiklerine sahipti - disk şeklindeki olanlar hariç falankslar - aşağı indikleri akan gergedanlara benzer. Bazı ayak kemikleri neredeyse 50 santimetre (20 inç) uzunluğundaydı. Uyluk kemikleri tipik olarak 1,5 m'dir (4,9 ft), bu boyut yalnızca bazı filler ve dinozorlar için aşılmıştır. Uyluk kemikleri sütun şeklindeydi ve diğer gergedanlardan çok daha kalın ve daha sağlamdı. tüccarlar bu sağlamlık önemlerini azalttığı için taraflar çok azaldı. Uzuvlar, daha küçük hayvanlarda olduğu gibi bükülmek yerine sütun benzeri bir duruşta tutuldu, bu da büyük uzuv kaslarına olan ihtiyacı azalttı.[4] Ön uzuvların üç parmağı vardı.[40]
Bilinen parçalı yapısı nedeniyle Paraceratherium Hayvanın iskeleti olan fosiller, keşfedildiği günden bu yana birkaç farklı şekilde yeniden yapılandırıldı.[41] 1923'te Matthew, daha az eksiksiz olanı temel alarak iskeletin yeniden inşasını çizmesi için bir sanatçıyı denetledi. P. transouralicum O zamana kadar bilinen örnekler, modern bir gergedanın oranlarını bir rehber olarak kullanıyor.[42] Sonuç çok çömelmiş ve kompakttı ve Osborn'un aynı yıl daha ince bir versiyonu çekilmişti. Daha sonraki bazı yaşam restorasyonları, altta yatan iskelete pek bakılmaksızın hayvanı çok ince yaptı.[4] Gromova, 1959'da daha eksiksiz bir iskelet rekonstrüksiyonu yayınladı. P. transouralicum Aral Formasyonundan iskelet, ancak bunda birkaç boyun omuru da yoktu.[43]
Paleobiyoloji

İngiliz zoolog Robert M. Alexander 1988'de aşırı ısınmanın şu ülkelerde ciddi bir sorun olabileceğini öne sürdü Paraceratherium boyutu nedeniyle.[44] Prothero'ya göre, yaşayanlar için en iyi analoglar Paraceratherium filler, gergedanlar ve hipopotamlar gibi büyük memeliler olabilir. Termoregülasyona yardımcı olmak için, bu hayvanlar gün boyunca gölgede dinlenerek veya su ve çamur içinde yuvarlanarak soğurlar. Ayrıca çoğunlukla geceleri yiyecek arar ve hareket ederler. Büyük boyutu nedeniyle, Paraceratherium hızlı koşamaz ve hareket edemezlerdi, ancak yiyecek kıtlığı olan bir ortamda gerekli olan büyük mesafeleri geçebilirlerdi. Bu nedenle geniş yaşam alanlarına sahip olabilirler ve göçmen olmuş olabilirler.[4] Prothero, indricotheres kadar büyük hayvanların çok büyük ev aralıklarına veya en az 1.000 kilometre karelik (250.000 dönüm) bölgelere ihtiyaç duyacağını ve kaynakların kıtlığı nedeniyle Asya'da birçok popülasyon veya çok sayıda insan için çok az yer olacağını öne sürüyor. neredeyse aynı tür ve cinslerin. Bu ilkeye rekabetçi dışlama; siyah gergedanın (bir tarayıcı) nasıl olduğunu açıklamak için kullanılır ve beyaz gergedan (bir otlakçı) Afrika'nın aynı bölgelerinde farklı nişlerden yararlanır.[3]
Yaşam alanlarındaki çoğu karasal yırtıcı, modern bir avcıdan büyük değildi. Kurt ve tehdit değildi Paraceratherium.[29] Yetişkin bireyler, herhangi bir kara avcısının saldıramayacağı kadar büyük olurdu, ancak gençler savunmasız olurdu. Bugti yataklarındaki kemikler üzerindeki ısırık izleri, yetişkinlerin bile 10-11 metre (33-36 ft) uzunluğundaki timsahlar tarafından avlanmış olabileceğini gösteriyor. Crocodylus bugtiensis. Fillerde olduğu gibi, hamilelik dönemi Paraceratherium uzun olabilir ve bireyler uzun ömürlü olmuş olabilir.[4] Paraceratherium belki de dişilerden ve yavrularından oluşan ve yırtıcı hayvanlardan korudukları küçük sürülerde yaşamış olabilir.[29] Kara memelileri için mümkün olan maksimum ağırlığın 20 ton (44.000 lb) olabileceği öne sürülmüştür ve Paraceratherium bu sınıra yakındı.[45] Memelilerin çok daha büyük boyuttaki sauropod dinozorlarına ulaşamamasının nedenleri bilinmemektedir. Nedeni, bunun yerine ekolojik olabilir biyomekanik ve belki de üreme stratejileriyle ilgilidir.[32] Görülen hareket, ses ve diğer davranışlar CGI gibi belgeseller Hayvanlarla Yürüyüş tamamen varsayımsaldır.[4]
Diyet ve beslenme

Basit, düşük taçlı dişler şunu gösterir: Paraceratherium nispeten yumuşak yapraklar ve çalılardan oluşan bir diyete sahip bir tarayıcıydı. Daha sonra rinoserozlar, yüksek taçlı dişlere sahip otlayıcıydı çünkü diyetleri dişlerini çabucak aşındıran grit içeriyordu. Çalışmaları Mezowear açık Paraceratherium dişler, yaratıkların yumuşak yapraklardan beslendiğini doğrular; mikro yıl çalışmalar henüz yapılmadı.[4] İzotop analizi gösterir ki Paraceratherium esas olarak beslendi C3 tesisleri esas olarak yapraklardır.[46][47] Perissodactyl akrabaları, atlar, tapirler ve diğer gergedanlar gibi, Paraceratherium olabilirdi arka bağırsak fermentörü; yiyeceklerinden nispeten az besin çıkaracak ve hayatta kalmak için büyük miktarlarda yemesi gerekecekti. Diğer büyük otçullar gibi, Paraceratherium büyük bir sindirim sistemine sahip olacaktı.[4]
Granger ve Gregory, büyük kesici dişlerin savunma için veya boynu aşağı doğru hareket ettirerek çalıları gevşetmek için kullanıldığını ve böylece kazma ve kaldıraç görevi gördüğünü savundu.[19] Tapirler, ön dişleriyle kabuğu soyarken, dalların etrafını sarmak için hortumlarını kullanırlar; bu yetenek yardımcı olabilirdi Paraceratherium. Bazı Rus yazarlar, dişlerin muhtemelen dalları kırmak, kabuğu soymak ve yüksek dalları bükmek için kullanıldığını ve erken Oligosen türlerinin daha sonraki türlere göre daha büyük dişlere sahip olduklarını, muhtemelen yaprak temelli diyetten daha fazla kabuğa sahip olduklarını öne sürdüler. İlgili türlerin artık çağdaş olduğu bilindiğinden ve dişlerdeki farklılıkların artık cinsel açıdan dimorfik olduğu düşünüldüğünden, ikinci fikir bugün kabul edilmiyor.[4] Sürüleri Paraceratherium küçük memelilerin ulaşamadığı uzun ağaçlardan sürekli yiyecek ararken göç etmiş olabilir.[29] Osborn, yiyecek arama yönteminin yüksek göz atma tarzına benzer olacağını öne sürdü. zürafa ve okapi kafaları yere yakın taşınan modern gergedanlar yerine.[42]
dağılım ve yaşam alanı
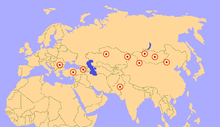
Atanabilir kalır Paraceratherium Avrasya genelinde erken ve geç Oligosen oluşumlarında (34–23 milyon yıl önce), günümüz Çin, Moğolistan, Hindistan, Pakistan, Kazakistan, Gürcistan, Türkiye, Romanya, Bulgaristan ve Balkanlar.[5] Dağılımları ile ilişkili olabilir paleocoğrafik Alp-Himalaya dağ kuşağının gelişimi. Aralığı Paraceratherium buluntular, üzerinde benzer bir çevreye sahip sürekli bir kara kütlesinde yaşadıklarını ima eder, ancak bu, bu bölgenin çeşitli deniz engellerine sahip olduğunu gösteren paleocoğrafik haritalarla çelişmektedir, bu nedenle cins, buna rağmen yaygın olarak dağılmakta başarılı olmuştur.[48] Bir arada bulunan fauna Paraceratherium diğer rinoserozlar dahil, artiodaktiller kemirgenler sakallı gelincikler hyaenodonts, nimravidler ve kediler.[29]
Yaşam alanı Paraceratherium Bulunduğu jeolojik oluşumların türlerine bağlı olarak, aralığı boyunca değiştiği görülmektedir.[29] Hsanda Gol Oluşumu Moğolistan, kurak bir çöl havzasını temsil ediyor ve faunanın çoğunlukla ağaç tepelerinden veya yere yakın beslenen hayvanlardan oluşması nedeniyle, çevrenin birkaç uzun ağaç ve sınırlı çalı örtüsüne sahip olduğu düşünülüyor.[49] Fosil polenleri üzerine yapılan bir araştırma, Çin'in büyük bir kısmının odunsu çalılık olduğunu gösterdi. tuzlu çalı, mormon çayı (Efedra), ve nitre çalı (Nitraria), hepsi kurak ortamlara uyarlanmıştır. Ağaçlar nadirdi ve yeraltı sularının yakınında yoğunlaşmıştı.[50] Çin'in bölgeleri Paraceratherium yaşadı kuru göller ve bol kum tepecikleri ve en yaygın bitki fosilleri, çöle adapte olmuş yapraklardır. Palibinia. Moğolistan ve Çin'deki ağaçlar arasında huş, karaağaç, meşe ve diğer yaprak döken ağaçlar, Sibirya ve Kazakistan'da da ceviz ağaçları vardı.[29] Pakistan'daki Dera Bugti kuru, ılıman ve subtropikal ormanlara sahipti.[46]
Yok olma
Nedenler Paraceratherium Yaklaşık 11 milyon yıl hayatta kaldıktan sonra soyu tükendi bilinmemektedir, ancak tek bir neden olması muhtemel değildir.[29] Teoriler şunları içerir iklim değişikliği, düşük üreme oranı ve istila gomphothere Oligosen'in sonlarında (28 ila 23 milyon yıl önce) Afrika'dan proboscideans. Gomphotheres, girdikleri habitatları önemli ölçüde değiştirmiş olabilir, aynı şekilde Afrika filleri ağaçları yok ederek ve ormanları otlaklara çevirerek bugün yapın. Yiyecek kaynakları azaldığında ve sayıları azaldığında, Paraceratherium halklar diğer tehditlere karşı daha savunmasız hale gelirdi.[51] Gibi büyük yırtıcılar Hyainailurus ve Amphicyon ayrıca Afrika'dan Asya'ya girdi. erken Miyosen (23 ila 16 milyon yıl önce); bunlar avlanmış olabilir Paraceratherium buzağılar. Diğer otoburlar da bu süre zarfında Asya'yı işgal etti.[29]
Referanslar
- ^ a b c d Prothero, 2013. s. 17–34
- ^ a b c Manias, C. (2014). "Bina Baluchitherium ve Indrikoterium: Erken Yirminci Yüzyıl Paleontolojisinde İmparatorluk ve Uluslararası Ağlar " (PDF). Biyoloji Tarihi Dergisi (Gönderilen makale). 48 (2): 237–78. doi:10.1007 / s10739-014-9395-y. PMID 25537636. S2CID 207150574.
- ^ a b c d e f g Prothero, 2013. s. 67–86
- ^ a b c d e f g h ben j k l m n Ö p q Prothero, 2013. s. 87–106
- ^ a b c Prothero, 2013. s. 35–52
- ^ Pilgrim, G.E. (1910). "Hindistan'ın Tersiyerinden yeni memeli cins ve türlerine ilişkin bildirimler". Hindistan Jeolojik Araştırmalarının Kayıtları. 40 (1): 63–71.
- ^ a b Cooper, C.F (1924). "Kafatasında ve dişlerinde Paraceratherium bugtiense: Alt Miyosen yaşlı Dera Bugti yataklarından anormal gergedan cinsi ". Kraliyet Topluluğu'nun Felsefi İşlemleri B: Biyolojik Bilimler. 212 (391–401): 369–394. Bibcode:1924RSPTB.212..369F. doi:10.1098 / rstb.1924.0009.
- ^ a b Forster-Cooper, C. (1911). "LXXVIII.Paraceratherium bugtiense, Belucistan'ın Bugti Tepelerinden yeni bir Rhinocerotidae cinsi. - Ön bildirim ". Annals ve Doğa Tarihi Dergisi. Seri 8. 8 (48): 711–716. doi:10.1080/00222931108693085. Arşivlendi 20 Kasım 2015 tarihinde orjinalinden. Alındı 28 Ağustos 2015.
- ^ Forster-Cooper, C. (1913). "XLIV. Thaumastotherium osborni, Belucistan'ın Bugti tepelerinin Üst Oligosen yataklarından yeni bir perissodactyles cinsi. - Ön bildirim ". Annals ve Doğa Tarihi Dergisi. Seri 8. 12 (70): 376–381. doi:10.1080/00222931308693412. Arşivlendi 7 Ekim 2015 tarihinde orjinalinden. Alındı 28 Ağustos 2015.
- ^ Forster-Cooper, C. (1913). "Genel adın düzeltilmesi". Annals ve Doğa Tarihi Dergisi. Seri 8. 12 (71): 504. doi:10.1080/00222931308693431. Arşivlendi 12 Ağustos 2018'deki orjinalinden. Alındı 13 Ocak 2018.
- ^ Forster-Cooper, C. (1923). "Baluchitherium osborni (? syn. Indricotherium turgaicum, Borrissyak) ". Londra Kraliyet Cemiyeti'nin Felsefi İşlemleri. 212 (391–401): 35–66. doi:10.1098 / rstb.1924.0002. JSTOR 92060.
- ^ Pavlova, M. (1922). "Indricotherium transouralicum n. sp. provenant du district de Tourgay ". Bulletin de la Société des Naturalistes de Moscou, Section Géologique (Fransızcada). 31: 95–116.
- ^ Borissiak, A.A. (1924). "Über die Unterfamilie Indricotheriinae Boriss. = Baluchitheriinae Osb". Zentralblatt für Mineralogie, Geologie und Paläontologie (Almanca'da). 18: 571–575.
- ^ Prothero, 2013. s. 1–16
- ^ Osborn, H.F. (1923). "Soyu tükenmiş dev gergedan Baluchitherium Batı ve Orta Asya ". Doğal Tarih. 3. 23: 208–228. Arşivlendi 9 Mart 2016 tarihinde orjinalinden. Alındı 8 Eylül 2014.
- ^ a b Yong-Xiang, L .; Yun-Xiang, Z .; Ji, L .; Zhi-Chao, L .; Kun, X. (2017). "Çin'in Gansu Eyaleti, Lanzhou Havzası'nın Erken Oligoseninden yeni paraceratheres (Perissodactyla, Mammalia) fosilleri". Vertebrata PalAsiatica. 55 (4): 367–381. doi:10.19615 / j.cnki.1000-3118.170922. Arşivlendi 12 Ekim 2017'deki orjinalinden. Alındı 11 Ekim 2017.
- ^ Forster-Cooper, C. (1922). "LXXIV.Metamynodon bugtiensis, sp. n., Belucistan'daki Dera Bugti yataklarından. - ön bildirim ". Annals ve Doğa Tarihi Dergisi. 9 (53): 617–620. doi:10.1080/00222932208632717.
- ^ Tissier, J .; Becker, D .; Codrea, V .; Costeur, L .; Fărcaș, C .; Solomon, A .; Venczel, M .; Maridet, O .; Smith, T. (2018). "Doğu Avrupa'dan Amynodontidae (Mammalia, Perissodactyla) hakkında yeni veriler: Eosen-Oligosen geçişi etrafındaki filogenetik ve paleobiyocoğrafik çıkarımlar". PLOS ONE. 13 (4): e0193774. Bibcode:2018PLoSO..1393774T. doi:10.1371 / journal.pone.0193774. PMC 5905962. PMID 29668673.
- ^ a b c Granger, W .; Gregory, W. K. (1936). "Soyu tükenmiş devasa gergedan hakkında ek notlar, Baluchitherium, Moğolistan Oligoseninden ". Amerikan Doğa Tarihi Müzesi Bülteni. 72: 1–73. hdl:2246/363.
- ^ Forster-Cooper, C. (1934). "Belucistan'ın soyu tükenmiş gergedanları". Kraliyet Topluluğu'nun Felsefi İşlemleri B: Biyolojik Bilimler. 223 (494–508): 569–616. Bibcode:1934RSPTB.223..569F. doi:10.1098 / rstb.1934.0013.
- ^ a b c d e f g Lucas, S. G .; Sobus, J.C. (1989), "Indricotheres'in sistematiği" Prothero, D. R .; Schoch, R.M. (editörler), Perissodactyl'lerin Evrimi, New York, New York ve Oxford, İngiltere: Oxford University Press, s. 358–378, ISBN 978-0-19-506039-3, OCLC 19268080
- ^ Zhan-Xiang, S. (1973). "Sinkang, Dzungaria oligoseninden yeni bir dev gergedan cinsi" (PDF). Vertebrata PalAsiatica. 11 (Çince ve İngilizce). 2: 182–191. Arşivlendi (PDF) 22 Ekim 2014 tarihinde orjinalinden. Alındı 22 Ekim 2014.
- ^ Antoine, P. O .; Karadenizli, L .; Saraç, G. E .; Sen, S. (2008). "Kuzey-Orta Anadolu'nun (Türkiye) Geç Oligoseninden dev bir rhinocerotoid (Mammalia, Perissodactyla)". Linnean Society'nin Zooloji Dergisi. 152 (3): 581–592. doi:10.1111 / j.1096-3642.2007.00366.x.
- ^ Ye, Y .; J., Meng; Yu, W.W. (2003). "Keşfi Paraceratherium Sincan'ın kuzey Junggar Havzasında " (PDF). Vertebrata PalAsiatica (Çince ve İngilizce). 41: 220–229. Arşivlendi (PDF) 3 Mart 2016'daki orjinalinden. Alındı 22 Ekim 2014.
- ^ Qui, Z.-X .; Wang, B.-Y .; Deng, T. (2004). "Linxia Basin'deki Oligocene'den Indricotheres (Perissodactyla, Mammalia), Gansu, Çin" (PDF). Vertebrata PalAsiatica (Çince ve İngilizce). 42 (3): 177–192. Arşivlenen orijinal (PDF) 22 Ekim 2014. Alındı 22 Ekim 2014.
- ^ a b c d Prothero, 2013. pp. 53–66
- ^ Radinsky, L. B. (1966). "The families of the Rhinocerotoidea (Mammalia, Perissodactyla)". Journal of Mammalogy. 47 (4): 631–639. doi:10.2307/1377893. JSTOR 1377893.
- ^ Holbrook, L. (1999). "The phylogeny and classification of tapiromorph perissodactyls (Mammalia)". Cladistics. 15 (3): 331–350. doi:10.1006/clad.1999.0107.
- ^ a b c d e f g h ben Prothero, 2013. pp. 107–121
- ^ Lucas, S. G .; Schoch, R. M.; Manning, E. (1981). "Sistematiği Forstercooperia, a Middle to Late Eocene hyracodontid (Perissodactyla: Rhinocerotoidea) from Asia and Western North America". Paleontoloji Dergisi. 55 (4): 826–841. JSTOR 1304430.
- ^ Wang, H .; Bai, B.; Meng, J.; Wang, Y. (2016). "Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids". Bilimsel Raporlar. 6 (1): 39607. Bibcode:2016NatSR...639607W. doi:10.1038/srep39607. PMC 5175171. PMID 28000789.
- ^ a b c Fortelius, M.; Kappelman, J. (1993). "The largest land mammal ever imagined". Linnean Society'nin Zooloji Dergisi. 108: 85–101. doi:10.1111/j.1096-3642.1993.tb02560.x.
- ^ Larramendi, A. (2016). "Omuz yüksekliği, vücut kütlesi ve hortumların şekli" (PDF). Acta Palaeontologica Polonica. 61. doi:10.4202 / app.00136.2014. S2CID 2092950. Arşivlendi (PDF) 24 Ağustos 2016 tarihinde orjinalinden.
- ^ Paul, G. S. (1997). "Dinosaur models: The good, the bad, and using them to estimate the mass of dinosaurs" (PDF). Dinofest International Proceedings: 129–142. Arşivlendi (PDF) 4 Mart 2016'daki orjinalinden. Alındı 3 Ocak 2015.
- ^ Taylor, M. P .; Wedel, M. J. (2013). "Why sauropods had long necks; and why giraffes have short necks". PeerJ. 1: e36. doi:10.7717/peerj.36. PMC 3628838. PMID 23638372.
- ^ Tsubamoto, T. (2012). "Estimating body mass from the astragalus in mammals". Acta Palaeontologica Polonica: 259–265. doi:10.4202/app.2011.0067. S2CID 54686160.
- ^ Antoine, P. O. (2014). "There were giants upon the earth in those days (Book Review)". Palaeovertebrata. 38: 1–3. doi:10.18563/pv.38.1.e4. Arşivlendi 12 Kasım 2014 tarihinde orjinalinden. Alındı 20 Ekim 2014.
- ^ Naish, D. (29 June 2013). "Tet Zoo Bookshelf: van Grouw's Tüysüz Kuş, Bodio's Eternity of Eagles, Witton's Pterozorlar, Van Duzer's Sea Monsters on Medieval and Renaissance Maps!". blogs.scientificamerican.com/tetrapod-zoology. Arşivlendi 6 Ekim 2014 tarihinde orjinalinden. Alındı 5 Ekim 2014.
- ^ Sander, P. M.; Christian, A .; Clauss, M .; Fechner, R .; Gee, C. T.; Griebeler, E. M.; Gunga, H. C.; Hummel, J. R.; Mallison, H .; Perry, S. F.; Preuschoft, H.; Rauhut, O. W. M.; Remes, K.; Tütken, T.; Wings, O.; Witzel, U. (2011). "Biology of the sauropod dinosaurs: The evolution of gigantism". Biyolojik İncelemeler. 86 (1): 117–155. doi:10.1111 / j.1469-185X.2010.00137.x. PMC 3045712. PMID 21251189.
- ^ Antoine, P. O.; Ibrahim Shah, S. M.; Cheema, I. U.; Crochet, J. Y.; Franceschi, D. D.; Marivaux, L .; Métais, G. G.; Welcomme, J. L. (2004). "New remains of the baluchithere Paraceratherium bugtiense from the Late/latest Oligocene of the Bugti hills, Balochistan, Pakistan". Asya Yer Bilimleri Dergisi. 24 (1): 71–77. Bibcode:2004JAESc..24...71A. doi:10.1016/j.jaes.2003.09.005.
- ^ Granger, W.; Gregory, W. K. (1935). "A revised restoration of the skeleton of Baluchitherium, gigantic fossil rhinoceros of Central Asia". Amerikan Müzesi Novitates. 787: 1–3. hdl:2246/2123.
- ^ a b Osborn, H. F. (1923). "Baluchitherium grangeri, a giant hornless rhinoceros from Mongolia". Amerikan Müzesi Novitates. 78: 1–15. hdl:2246/3262.
- ^ Gromova, V. L. (1959). "Gigantskie nosorogi". Trudy Paleontology Institut Akademii Nauk SSSR (Rusça). 71: 154–156.
- ^ Alexander, R.M. (1998). "All-time giants: the largest animals and their problems". Paleontoloji. 41 (6): 1231–1245. Arşivlendi 27 Eylül 2016 tarihinde orjinalinden. Alındı 1 Mart 2016.
- ^ Clauss, M .; Frey, R.; Kiefer, B.; Lechner-Doll, M.; Loehlein, W.; Polster, C.; Rössner, G. E.; Streich, W. J. (2003). "The maximum attainable body size of herbivorous mammals: Morphophysiological constraints on foregut, and adaptations of hindgut fermenters" (PDF). Oekoloji (Gönderilen makale). 136 (1): 14–27. Bibcode:2003Oecol.136...14C. doi:10.1007/s00442-003-1254-z. PMID 12712314. S2CID 206989975. Arşivlendi (PDF) 8 Haziran 2019 tarihinde orjinalinden. Alındı 6 Ağustos 2018.
- ^ a b Martin, C.; Bentaleb, I.; Antoine, P. -O. (2011). "Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 311 (1–2): 19–29. Bibcode:2011PPP...311...19M. doi:10.1016/j.palaeo.2011.07.010.
- ^ Wang, Y .; Deng, T. (2005). "A 25 m.y. Isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau". Dünya ve Gezegen Bilimi Mektupları. 236 (1–2): 322–338. Bibcode:2005E&PSL.236..322W. doi:10.1016/j.epsl.2005.05.006.
- ^ Sen, S .; Antoine, P. O.; Varol, B.; Ayyildiz, T.; Sözeri, K. (2011). "Giant rhinoceros Paraceratherium and other vertebrates from Oligocene and middle Miocene deposits of the Kağızman-Tuzluca Basin, Eastern Turkey". Naturwissenschaften. 98 (5): 407–423. Bibcode:2011NW.....98..407S. doi:10.1007/s00114-011-0786-z. PMID 21465174. S2CID 19968371.
- ^ Meng, J.; McKenna, M. C. (1998). "Faunal turnovers of Palaeogene mammals from the Mongolian Plateau". Doğa. 394 (6691): 364–367. Bibcode:1998Natur.394..364M. doi:10.1038/28603. S2CID 204998953.
- ^ Leopold, E. B.; Liu, G .; Clay-Poole, S. (1992), "Low-biomass vegetation in the Oligocene?", in Prothero, D. R.; Berggren, W.A. (eds.), Eocene-Oligocene Climatic and Biotic Evolution, Princeton: Princeton University Press, pp. 399–420, ISBN 978-0-691-02542-1
- ^ Putshkov, P. V. (2001). ""Proboscidean agent" of some Tertiary megafaunal extinctions". Terra Degli Elefanti Congresso Internazionale: The World of Elephants: 133–136.
Kaynakça
- Prothero, D. (2013). Rhinoceros Giants: The Palaeobiology of Indricotheres. Indiana: Indiana University Press. ISBN 978-0-253-00819-0.