Bakteriyoplankton sayım yöntemleri - Bacterioplankton counting methods
Bakteriyoplankton sayımı ... tahmin bolluğundan bakterioplankton deniz mikrobiyologları için yararlı bilgi olan belirli bir su kütlesinde. Gözlemlenen suda mevcut olan sayıyı belirlemek için yıllar içinde çeşitli sayma metodolojileri geliştirilmiştir. Bakteriyoplankton saymak için kullanılan yöntemler şunları içerir: epifloresan mikroskobu, akış sitometrisi, bölünen hücre sıklığı (FDC) aracılığıyla verimlilik ölçüleri, timin şirketleşme ve lösin kuruluş.
Gibi faktörler tuzluluk, sıcaklık, enlem, çeşitli besin seviyeleri, su hareketi ve diğer organizmaların varlığı bakterioplankton sayımını etkileyebilir.[1][2][3][4][5] Bu faktörlerdeki değişiklikler, bakterioplankton sayısını etkileyerek, su kütlesine, konuma, kıyıdan uzaklığa ve mevsime göre değişmesine neden olur.[6][7][8]
Bakteriyoplankton sayısı genellikle şu şekilde ifade edilir: ml başına hücre (hücre ml−1).
Kullanımlar
Deniz mikrobiyolojisini ve su ekosistemini anlamada bakterioplankton sayımı faydalı olabilir. Bakteriyoplankton sayısının gözlemlenmesi, aşağıdaki konularda daha fazla bilgi sağlayabilir:
- Su sistemlerinde çeşitli besin döngüsünde yer alan süreçler[9][10]
- Suda üretkenlik için [11]
- Çevresel değişiklikleri, özellikle aşırılıkları belirlemek için [12]
- Su ekosistemindeki besin bileşimi [16]
- Diğer suda yaşayan organizmaların (yani karides) bolluğu ve koşulları [17]
Epifloresan mikroskobu
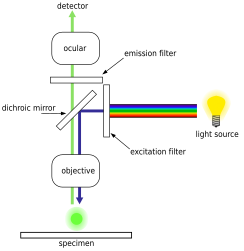
Epifloresan mikroskobu kullanımına dayanan gelişmiş bir optik mikroskop tekniğidir. floresan boyalar belirli biyolojik belirteçlere bağlanan ve daha sonra ayırt edici emisyon spektrumu lens aracılığıyla tanımlanır. Floresan boyalar şunları içerir: DAPI, Akridin Portakal, SYBR Yeşil 1 ve YO-PRO-1, hepsi bakteri ve virüsler gibi biyolojik örneklerde hem DNA hem de RNA yapılarını boyayabilmektedir.[18][19][20][21] Bununla birlikte, DNA boyama öncelikle bakteriyel hücre tanımlaması için kullanılır. Modern epifloresan mikroskobu ile, bakteri hücre miktarlarını tahmin etmek ve saymak için endüstri standardı, DAPI lekesi.[22] Bu teknik, deniz suyu, çeşitli tatlı su kaynakları, toprak ve tortular gibi çok çeşitli ortam ve konumlardan numuneler için uygulanabilir.[22]
Numaralandırma tekniği
Standart bir deneyde, hazırlanan bakteri numuneleri sayım slaytlarına yerleştirilir ve ardından epifloresan mikroskobu altında görüntülenir. Büyütme, sayım slaydı üzerindeki 0,1 X 0,1 mm kare birimlerin açıkça görülebildiği bir seviyeye ayarlanır.[23] Bakterileri ölçmek için, hücreler 5-30 rasgele kare birim görüş alanında sayılır ve alan başına ortalama bir bakteri sayısı tablo haline getirilir.[22] Bu değer daha sonra, lam biriktirme alanındaki toplam görüş alanı sayısını belirleyerek ve bunu sayma birimi başına ortalama bakteri sayısıyla çarparak mL başına toplam bakteri hücre sayısını tahmin etmek için ekstrapole edilir.[23]
Güvenilirlik
İçin numaralandırmak bakteri hücre miktarları, bir numunedeki sadece küçük bakteri bölümleri, lojistik nedenlerden dolayı fiziksel olarak sayılır ve bunun üzerine toplam bolluklar ekstrapolasyon ile tahmin edilir. Ortalama değerler daha sonra numuneler arasında karşılaştırma için kullanılır. Bununla birlikte, toplam bolluk miktarlarını tahmin etmek için yalnızca küçük bir alt kümenin tablolarının kullanıldığı bu tekniğin doğruluğu sorgulanmıştır.[22] Öncelikle, bakteri hücrelerinin sayım slaytlarındaki dağılımının düzensiz ve tutarsız olabileceği gösterilmiştir.[22] Ek olarak, bu tekniği kullanarak bakteri sayımlarının meşru bir tahminini elde etmek için, 20 görüş alanından 350'den fazla bireysel hücrenin ölçülmesi gerektiği öne sürülmüştür.[22] Bu sadece zaman alıcı olmakla kalmaz, aynı zamanda bazı örneklerde elde edilmesi zor olabilir.
Akış Sitometrisi

{kind=link}
Akış sitometrik analizi (veya, akış sitometrisi ) birçok klinik uygulamada yaygın bir prosedürdür. Bununla birlikte, otuz yılı aşkın bir süre önce keşfedilmesine rağmen, bakteriyoplankton sayımında sucul mikrobiyal ekoloji tarafından benimsenmesi nispeten yavaş olmuştur.[24] Kullanımı henüz geçmedi epifloresan mikroskobu.[25] Her iki bolluk tahmin tekniğinin de nispeten doğru olmasına rağmen, akış sitometrisi insan hatasına daha az meyillidir, daha hassastır, daha yüksek bir çözünürlüğe sahiptir ve birkaç dakika içinde on binlerce hücreyi inceleyebilir.[24] Akış sitometrisi, hücre bolluğunun yanı sıra hücrelerin boyutu, aktivitesi ve morfolojisi hakkında da bilgi sağlayabilir.[26]
Akış sitometrisi, hem fotosentetik hem de fotosentetik olmayan bakteriyoplanktonları ayırt etmek ve ölçmek için kullanılabilir.[26] Fotosentetik prokaryotların nicelendirilmesi, örneğin siyanobakteriler ve pikoökaryotik yosun fotosentetik pigmentlerin floresan özelliği sayesinde mümkün olur.[27] Örneğin, iki ana fotosentetik prokaryotta farklı fotosentetik pigment oluşumu, proklorokok ve Synechococcus onların farkına varmalarını sağlar.[28][29][30] Prochlorococcus divinil-klorofiller içerir a ve b synechococcus hem turuncu hem de kırmızı floresan yayarken mavi veya UV ışığı ile uyarılma altında yalnızca kırmızı floresan gösteren; portakaldan fikobilinler ve kırmızıdan klorofil. Floresansın yanı sıra, prochlorococcus ve synechococus önemli ölçüde farklı boyutlardadır ve bu nedenle akış sitometrik analizi üzerine farklı saçılma sinyalleri sunar. Bu, farklılaşmalarına daha da yardımcı olur.[31] Prochlorococcus miktarının belirlenmesi, neredeyse yalnızca akış sitometrisi yoluyla mümkün olduğu için büyük bir buluş olarak kabul edilir. Bunun nedeni, epifloresan mikroskopisinin proklorokokta bulunan düşük klorofil otofloresanı tespit edememesidir.[26]
Fotosentetik bakteriyoplanktonun yanı sıra, fotosentetik olmayan bakteriyoplankton da akış sitometrisi ile numaralandırılabilir. Bu, DNA veya gıda vakuol boyama yoluyla yapılır.[27] Akış sitometrisi, proklorokkusu, benzer boyutlarından dolayı başlangıçta kafa karıştıran heterotrofik bakterilerden ayırmada özellikle başarılı olmuştur.
Epifloresan mikroskobunun birçok mikrobiyal ekoloji laboratuarında akış sitometrisi üzerinde kullanılması bir dizi ekonomik ve pratik faktörden sorumlu tutulabilir. İlk olarak, ticari akış sitometrelerinin kullanımı, titizlikle eğitilmiş bir teknisyenin uzmanlığını gerektirir. İkinci olarak, akış sitometreleri epi-floresan mikroskopi aparatına kıyasla oldukça pahalıdır. Üçüncüsü, birçok akış sitometresi kan hücrelerini incelemek için tasarlanmıştır; okyanus bakterileri nispeten küçüktür ve bu nedenle birçok ticari akış sitometresinde çözünürlük sınırına yaklaşır.[32]
Numaralandırma Süreci
Bakteriyoplanktonun akış sitometrik kantifikasyonu dört adımı içerir: sabitleme, boyama, veri işleme ve veri yorumlama.
Sabitleme
Sabitleme sadece numuneyi korumak için değil, aynı zamanda hücrelerin lekelere geçirgenliğini artırmak için yapılır.[24] Bununla birlikte, çoğu fiksasyon ajanı, boyut, ışığın saçılma şekli gibi belirli yönleri değiştirerek hücreleri değiştirme kapasitesine sahiptir. otofloresans ve nükleik asitler. Hücrelerin akış sitometrik ayrımı bu niteliklere dayandığından bu sorunludur. Bazı fiksatifler ayrıca hücrelerin tamamen kaybolmasına neden olur.[24] Şu anda, fiksasyon işleminde kullanılan ajanlardan bazıları iki varyasyon formaldehit (formalin ve paraformaldehit),% 70 etanol, glutaraldehit ve TCA içerir.[33] Protein ve nükleik asitler için en iyi fiksasyon ajanının, hücrelere hızlı bir şekilde girebilmesi nedeniyle paraformaldehit olduğu varsayılmaktadır.[24]
Boyama
Akış sitometrisinde boyama, bakteriyoplanktonun bakteriyel olmayan partiküllerden ayrılmasını sağlar. Geniş bir yelpazede numunenin inkübasyonunu içerir. florokromlar UV uyarımlı boyalar gibi (DAPI ve Hoechst 33342) ve mavi ışıkla uyarılan nükleik asit boyaları (TO-PRO-1, TOTO-1, SYBR Yeşil I ).[31] Uzun süre, akış sitometreleri incelemek için UV uyarımlı boyalar kullandı. bakterioplankton ya sınırlı duyarlılığa sahip düşük maliyetli akış sitometrelerinde ya da heterotrofik bakterileri ototroflardan ayırmak için gereken yüksek duyarlılığa sahip pahalı akış sitometrelerinde kullanılabilir. SYBR Green I gibi mavi uyarımlı boyaların piyasaya sürülmesi, düşük maliyetli, yüksek hassasiyetli akış sitometrelerinde bakteriyoplanktonun yüksek kaliteli akış sitometrik analizini mümkün kıldı.[31]
Optimum boyama için inkübasyon süreleri bileşikten bileşiğe değişir. UV uyarımlı boyalar bir saat veya daha fazla sürebilirken, mavi ışık uyarımlı boyalar sadece 15 dakika gerektirir.[24]
Boyama gibi tamponlar eşlik edebilir. Triton X-100 Bu, hücreleri lekelere karşı daha geçirgen hale getirir. Özellikle TO-PRO-1 gibi hücre geçirimsiz boyalarda kullanılırlar. Tamponlar ayrıca Picogreen, YO-PRO-1 ve YOYO-1 gibi iyonik güce duyarlı boyaları seyreltmek için kullanılır. Triton-X-100 gibi tamponlar sadece klorofil flüoresansını söndürmekle kalmayıp aynı zamanda istenmeyen arka plan flüoresanı da oluşturduğundan, tamponların kullanımı hücrelere zararlı olabilir. Bu, aralarında ayrım yapmanın zorluğunu artırabilir heterotrofik bakteri ve ototrofik prokaryotlar.[24]
Sayma
{kind=link}
Bir akış sitometrik analizde, her saniye 200'den fazla hücre bir lazer ışınının veya cıva lambasının önünden geçer. Fotoçoğaltıcılar her bir parçacığın saçtığı ışık miktarını ve uyarılma üzerine yayılan floresanı toplar. Bu bilgi daha sonra içselleştirilir ve sistem tarafından bir olay olarak yorumlanır. Bununla birlikte, akış sitometrelerinin çok az çabayla hücreleri sayma kabiliyetine rağmen, çoğunun gerçek hücre konsantrasyonunu belirleme yolu yoktur. Bu, miktarı önceden belirlenmiş (bakterilerin tanelere oranının belirlenmesine yardımcı olan) referans boncukların kullanımı, deneyden önce ve sonra ağırlık ölçümleri ve günlük akış kalibrasyonu dahil olmak üzere çeşitli yöntemlerle belirlenebilir.[24]
Akış sitometrelerinin büyük bir avantajı, farklı bakterioplanton popülasyonlarını tanımlama yetenekleridir. Bu ayrım, dört faktörün analizi yoluyla yapılır; ışık saçılması, yeşil floresan, mavi floresan ve kırmızı floresan. Işık saçılımı analizi tek başına yetersizdir ve genellikle birkaç nedenden dolayı floresanla birlikte incelenir; birincisi, deniz suyu bakteri gibi ışığı yayan birçok parçacığı içerir. İkinci olarak, birçok okyanus bakterisinin boyutları çözünürlük sınırına yaklaşır. Hücreler tarafından saçılan ışık miktarı sadece hücrelerin boyutuna göre değil, aynı zamanda parçacığın iç yapısı, kırılma indisi, şekli ve yönelimi ile de belirlenir. Dağınık ışık ikisinden biri olarak sınıflandırılır ileri dağılım (FSC) veya yan dağılım (SSC). İlki, hücre hacmi ve kütlesi ile ilişkilendirilirken, ikincisi, hücrelerin kırılma indisi, içeriği ve tanecikliği ile ilişkilendirilmiştir. [24]
Hücre konsantrasyonları 2,5 × 10'dan yüksek olduğunda6 ml başına hücre sayısı, hücrenin birden fazla yakınlıktan geçme ve tek bir olay olarak kaydedilme olasılığı büyütülür. Bu tesadüf olarak bilinir ve numuneyi önceden seyrelterek kolayca önlenebilir.[31]
Üretkenlik ölçüleri
Hücreleri bölme sıklığı
Bölünen hücre sıklığı (FDC), suda yaşayan heterotrofik bir bakteri topluluğunun ortalama büyüme oranını tahmin etmek için kullanılan bir yöntemdir.[34] Yöntem, özellikle hücre bölünmesini kullanır septum büyüme oranının bir göstergesi olarak oluşum.[34] Tek tek hücreler arasındaki boşluklar (istila ) altında gözlemlenir epifloresan mikroskobu.[34] FDC, halihazırda bölünen hücrelerin oranı ile bir bakteri topluluğundaki büyüme oranı arasında bir ilişki olduğu varsayımına dayanmaktadır.[35]
Timidin birleşmesi
Timidin dahil etme, bakteri büyümesini tahmin etmek için en yaygın olarak kullanılan yöntemlerden biridir.[36] Timidin bir öncü için DNA ve DNA sentezi, tritiatlı timidin dahil edilerek ölçülebilir. triklorasetik asit (TCA) - yoluyla çözünmeyen malzeme kurtarma yolları.[37] Timidin birleşmesi, DNA sentezini sadece büyüyen hücrelerin radyoaktif timidini DNA sentezlemek için dahil edebileceği varsayımını kullanarak DNA sentezi oranlarına dayalı olarak büyümeyi ölçer.[38]
Bu prosedürün zayıf yönleri, bir numuneye trityumlanmış timidin eklendiğinde DNA'nın yanı sıra diğer moleküllerin etiketlenmesini içerir.[36] Karbon sınırlaması durumlarında timidin, bir DNA öncüsü yerine bir karbon kaynağı olarak da kullanılabilir.[36] Timidin birleştirme deneylerinin sonuçları, diğer moleküllere kıyasla DNA'ya katılan timidin oranı bilinmediğinde yanıltıcı olabilir.[36]
Lösin birleşmesi
Lösin su bakteri topluluklarında protein sentezinin bir ölçüsü olarak kullanılır.[39] Numunelere radyo etiketli lösin ilave edilerek hücrenin sıcak trikloroasetik asit (CA) ile çözünmeyen kısımlarında birikimi belirlenir.[39] Örnekler daha sonra membran filtrede toplanır.[39] Lösin proteini, suda yaşayan bakteri popülasyonlarının% 50'sinden fazlası tarafından alınır ve bakteri topluluğundaki nitrojen kullanımını tahmin etmek için lösin katılımı kullanılabilir.[39]
Deniz mevsimsel veraset dinamikleri
Bakteri popülasyonlarının benzersiz metabolizmaları ve kaynak tercihleri olduğu için, bakteriyel bileşimlerin yüksek çözünürlüklü zaman serileri analizinin kullanılması, mevsimsel bakteri dizisindeki modellerin tanımlanmasına izin verir.[40] Bakteri topluluğu kompozisyonlarındaki farklılıklar, türler arası bakteri etkileşimlerinin fotosentetik plankton ile belirli permütasyonlarına yol açar. protist otlayanlar ve fajlar dolayısıyla mevsimsel dinamikleri etkiler. Nüfus dinamikleri ve kompozisyonundaki kalıpları doğrulamak için kullanılan istatistiksel yöntemlerin birkaç yıl içinde tekrarlanabilir olduğu gösterilmiştir ve çevresel faktörler bu zamansal modellerin öngörücüleri olarak hizmet etmiştir.[41] Bakteriyoplankton için çoğu araştırma aktivitesi şu anda kuzey yarımkürenin ılıman sularında 30 ° N'den Kuzey Kutup Dairesi 66 ° N'de.[40]
Ilıman bölgelerde mevsimsel geçiş
Fitoplankton popülasyonlarının mevsimsel ardışıkları tutarlı bir yinelenen model izledikçe, bakteri dinamikleri ve fitoplankton dizilimi ilişkilendirilebilir.[40] Genel olarak bakteri bileşimindeki mevsimsel değişiklikler, sıcaklıktaki değişiklikleri takip eder ve klorofil a besin bulunabilirliği bakterioplankton büyüme oranlarını sınırlarken.[42][43][44][45][6][46] Sonbahar / kış sonlarında su sütununun karıştırılması sırasında, yüzeye getirilen besinler, belirgin bir diatom bahar çiçeklenmesini ve ardından dinoflagellatların gelmesini sağlar.[40] Sonra bahar çiçeği Fitoplankton bozunmasından Çözünmüş organik maddenin (DOM) salınması nedeniyle bakteri üretimi ve büyümesi artar.[47][48] Bu erken ardıl aşamada, sınıfın üyeleri Flavobakteriler (Bakteroidler) tipik olarak bakteri topluluğunun baskın bileşenleridir.[49][50] Genom analizi ve meta-transkriptomik DOM'un bozunmasını ve asimilasyonunu kolaylaştıran çoklu hidrolitik enzimler içeren bakterilerin varlığını ortaya çıkarmıştır.[51][52][53][54] İlkbahar çiçeklerinde, bazı üyeler Roseobacter clade (Alfaproteobakteriler ) ve bazı Gammaproteobacteria genellikle DOM bozulmasıyla ilişkilendirilir.[48][49] Sıcaklıklar arttıkça ve ilkbahardan gelen besinler tükendikçe, daha küçük fitoplanktonlar ve siyanobakteriler şimdi oligotrofik sularda büyür.[40]
Yaz aylarında sular katmanlaştıkça, Roseobacter, SAR86 (Gammaproteobacteria) ve SAR11 (Alphaproteobacteria) bakteri türleri bollukta artar.[55][56] Sık görülen sonbahar diyatom ve dinoflagellat Baltık Denizi'ndeki çiçeklenmeler, tamamlayıcı besin girdileri ile ilişkilidir ve yüksek frekanslı örnekleme, sonbaharda, Aktinobakteriler genellikle artış ve ardından sonbahara özgü farklı Flavobakteriler SAR11 ve Planctomycetes.[49]
İçinde Akdeniz, yaz tabakalaşması sırasında bir zamanlar baskın olan oligotrofik popülasyonlar yavaş yavaş yok olurken, derin kış karışımı SAR11 sınıfının üyelerinin artan çeşitlilik elde etmesine izin verir.[57] Akdeniz'deki arkeler arasında, Thaumarchaeota Deniz Grubu I (MGI) ve Euryarchaeota Deniz Grubu II (MGII.B) popülasyonları kışın baskın hale geldi.[58] İken Baltık Denizi, kış karışımı getiriyor Epsilon-proteobakteriler ve Archaea popülasyonları derin habitatlarından yüzeye çıkarır.[49]
Referanslar
- ^ Uzun RA, Azam F (2001-12-05). "Deniz suyundaki bakterioplankton topluluğu zenginliğinin mikro ölçekli düzensizliği". Sucul Mikrobiyal Ekoloji. 26 (2): 103–113. doi:10.3354 / ame026103.
- ^ He J, Zhang F, Lin L, Ma Y, Chen J (2012). "Batı Kanada Havzasında 2008 yazında bakterioplankton ve pikofitoplankton bolluğu, biyokütle ve dağılımı". Derin Deniz Araştırmaları Bölüm II: Oşinografide Güncel Çalışmalar. 81-84: 36–45. Bibcode:2012DSRII..81 ... 36H. doi:10.1016 / j.dsr2.2012.08.018.
- ^ Wei C, Bao S, Zhu X, Huang X (2008). "Çin, Chaohu Gölü'ndeki bakteriyoplankton topluluğu kompozisyonunun mekansal-zamansal varyasyonları". Doğa Bilimlerinde İlerleme. 18 (9): 1115–1122. doi:10.1016 / j.pnsc.2008.04.005.
- ^ López-Flores R, Boix D, Badosa A, Brucet S, Quintana XD (2009). "Kapalı Akdeniz tuz bataklıklarında (Kuzeydoğu İspanya) bakteriyoplankton ve fitoplankton dinamiklerini etkileyen çevresel faktörler". Deneysel Deniz Biyolojisi ve Ekoloji Dergisi. 369 (2): 118–126. doi:10.1016 / j.jembe.2008.11.003.
- ^ Medvinsky, Alexander B .; Adamovich, Boris V .; Aliev, Rubin R .; Chakraborty, Amit; Lukyanova, Elena V .; Mikheyeva, Tamara M .; Nikitina, Ljudmila V .; Nurieva, Nailya I .; Rusakov, Alexey V. (2017). "Sıcaklık dalgalanmalarını etkileyen bir faktör olarak ve göl bakteriyoplanktonlarının bolluğunun öngörülebilirliği". Ekolojik Karmaşıklık. 32: 90–98. doi:10.1016 / j.ecocom.2017.10.002.
- ^ a b Andersson AF, Riemann L, Bertilsson S (Şubat 2010). "Pyrosequencing, Baltık Denizi bakterioplankton toplulukları içindeki taksonların zıt mevsimsel dinamiklerini ortaya koyuyor". ISME Dergisi. 4 (2): 171–81. doi:10.1038 / ismej.2009.108. PMID 19829318.
- ^ Ghiglione JF, Murray AE (Mart 2012). "Kıyı Antarktika deniz bakteriyoplanktonunda yazdan kışı belirgin hale getirme ve kışın daha yüksek zenginlik". Çevresel Mikrobiyoloji. 14 (3): 617–29. doi:10.1111 / j.1462-2920.2011.02601.x. PMID 22003839.
- ^ Straza TR, Ducklow HW, Murray AE, Kirchman DL (2010-11-01). "Antarktika kıyı sularında bakteri gruplarının bolluğu ve tek hücre aktivitesi". Limnoloji ve Oşinografi. 55 (6): 2526–2536. Bibcode:2010LimOc..55.2526S. doi:10.4319 / lo.2010.55.6.2526.
- ^ Currie DJ, Kalff J (Mart 1984). "Bakteriyoplankton ve fitoplanktonun tatlı suda fosfor alımındaki göreceli önemi1". Limnoloji ve Oşinografi. 29 (2): 311–321. Bibcode:1984LimOc..29..311C. doi:10.4319 / lo.1984.29.2.0311.
- ^ Lindström ES (Aralık 2001). "Bakteriyoplankton Topluluğu Kompozisyonu Üzerindeki Etkili Faktörlerin Araştırılması: Beş Mezotrofik Gölün Saha Çalışmasının Sonuçları". Mikrobiyal Ekoloji. 42 (4): 598–605. doi:10.1007 / s00248-001-0031-y. PMID 12024242. S2CID 22656746.
- ^ Cotner JB, Biddanda BA (2002-03-01). "Küçük Oyuncular, Büyük Rol: Pelajik Su Ekosistemlerinde Biyojeokimyasal Süreçler Üzerindeki Mikrobiyal Etki". Ekosistemler. 5 (2): 105–121. CiteSeerX 10.1.1.484.7337. doi:10.1007 / s10021-001-0059-3. S2CID 39074312.
- ^ Harnisz M (Mart 2013). "Su ortamındaki değişikliklerin bir göstergesi olarak doğal bakterilerin toplam direnci". Çevre kirliliği. 174: 85–92. doi:10.1016 / j.envpol.2012.11.005. PMID 23246751.
- ^ Chen, Xinxin; Wang, Kai; Guo, Annan; Dong, Zhiying; Zhao, Qunfen; Qian, Jie; Zhang, Demin (2016). "Fazla fosfat yüklemesi, zamanla oligotrofik kıyı suyu mikrokozmoslarındaki bakterioplankton topluluğu kompozisyonunu değiştirir". Deneysel Deniz Biyolojisi ve Ekoloji Dergisi. 483: 139–146. doi:10.1016 / j.jembe.2016.07.009.
- ^ Dai, Wenfang; Zhang, Jinjie; Tu, Qichao; Deng, Ye; Qiu, Qiongfen; Xiong, Jinbo (2017). "Bakteriyoplankton topluluğu ve türler arası etkileşim, artan kıyı ötrofikasyonunu gösterir". Kemosfer. 177: 317–325. Bibcode:2017Chmsp.177..317D. doi:10.1016 / j.chemosphere.2017.03.034. PMID 28319885.
- ^ Urakawa, Hidetoshi; Bernhard, Anne E. (2017). "Mikrobiyal göstergeler kullanarak sulak alan yönetimi". Ekolojik Mühendislik. 108: 456–476. doi:10.1016 / j.ecoleng.2017.07.022.
- ^ Haukka K, Kolmonen E, Hyder R, Hietala J, Vakkilainen K, Kairesalo T, Haario H, Sivonen K (Şubat 2006). "Besin yüklemesinin göl mezokozomlarında bakterioplankton topluluğu kompozisyonuna etkisi". Mikrobiyal Ekoloji. 51 (2): 137–46. doi:10.1007 / s00248-005-0049-7. PMID 16435168. S2CID 35399139.
- ^ Zhang D, Wang X, Xiong J, Zhu J, Wang Y, Zhao Q, Chen H, Guo A, Wu J (2014). "Karideslerin sağlık durumunun biyolojik göstergeleri olarak bakterioplankton toplulukları". Ekolojik Göstergeler. 38: 218–224. doi:10.1016 / j.ecolind.2013.11.002.
- ^ Tanious FA, Dana JM, Buczak H, Ratmeyer LS, Wilson WD (1992-03-31). "DAPI (4 ', 6-diamidino-2-fenilindol), DNA ve RNA'ya farklı şekilde bağlanır: AT bölgelerinde minör-oluk bağlanması ve AU bölgelerinde interkalasyon". Biyokimya. 31 (12): 3103–3112. doi:10.1021 / bi00127a010. PMID 1372825.
- ^ Gonzalez K, Mcvey S, Cunnick J, Udovichenko IP, Takemoto DJ (1995). "Akış sitometrisi kullanılarak kültürde normal ve galaktozemik lens epitel hücrelerinde toplam DNA ve RNA'nın akridin turuncusu diferansiyel boyaması". Güncel Göz Araştırması. 14 (4): 269–273. doi:10.3109/02713689509033525. PMID 7541739.
- ^ Noble RT, Fuhrman JA (1998-02-13). "Deniz virüsleri ve bakterilerinin hızlı epifloresans sayımı için SYBR Green I kullanımı". Sucul Mikrobiyal Ekoloji. 14 (2): 113–118. doi:10.3354 / ame014113.
- ^ Marie D, Vaulot D, Partensky F (Mayıs 1996). "Deniz prokaryotlarının akış sitometrik analizi için yeni nükleik asit boyaları YOYO-1, YO-PRO-1 ve PicoGreen'in uygulanması". Uygulamalı ve Çevresel Mikrobiyoloji. 62 (5): 1649–55. doi:10.1128 / AEM.62.5.1649-1655.1996. PMC 167939. PMID 8633863.
- ^ a b c d e f Muthukrishnan T, Govender A, Dobretsov S, Abed RM (2017/01/08). "Epifloresan Mikroskopi Kullanarak Bakteri Sayımının Güvenilirliğini Değerlendirme". Deniz Bilimleri ve Mühendisliği Dergisi. 5 (1): 4. doi:10.3390 / jmse5010004.
- ^ a b O'Connor JT, O'Connor T, Twait R (2009). Su Arıtma Tesisi Performans Değerlendirmeleri ve İşlemleri. John Wiley & Sons, Inc. s. 193–198. doi:10.1002 / 9780470431474.app1. ISBN 9780470431474.
- ^ a b c d e f g h ben Gasol, Josep M .; Giorgio, Paul A. del (2000-06-30). "Doğal planktonik bakterileri saymak ve planktonik bakteri topluluklarının yapısını anlamak için akış sitometrisini kullanma". Scientia Marina. 64 (2): 197–224. doi:10.3989 / scimar.2000.64n2197. ISSN 1886-8134.
- ^ Giorgio, Paul A. del; Bird, David F .; Prairie, Yves T .; Planas, Dolors (1996-06-01). "Yeşil nükleik asit lekesi SYTO 13 ile göl planktonundaki bakteri bolluğunun akış sitometrik tayini". Limnoloji ve Oşinografi. 41 (4): 783–789. Bibcode:1996LimOc..41..783G. doi:10.4319 / lo.1996.41.4.0783. ISSN 1939-5590.
- ^ a b c Sieracki, Michael E .; Haugen, Elin M .; Cucci, Terry L. (1995-08-01). "Sargasso Denizi'ndeki heterotrofik bakterilerin fazla tahmini: akış ve görüntüleme sitometrisi ile doğrudan kanıt". Derin Deniz Araştırmaları Bölüm I: Oşinografik Araştırma Makaleleri. 42 (8): 1399–1409. Bibcode:1995 DSRI ... 42.1399S. doi:10.1016 / 0967-0637 (95) 00055-B. ISSN 0967-0637.
- ^ a b Zubkov MV, Burkill PH, Topping JN (2007-01-01). "DNA lekeli okyanus planktonik protistlerinin akış sitometrik sayımı". Plankton Araştırma Dergisi. 29 (1): 79–86. doi:10.1093 / plankt / fbl059.
- ^ WATERBURY, JOHN B .; WATSON, STANLEY W .; GUILLARD, ROBERT R. L .; MARKA, LARRY E. (Ocak 1979). "Tek hücreli, denizel, planktonik, siyanobakterinin yaygın oluşumu". Doğa. 277 (5694): 293–294. Bibcode:1979Natur.277..293W. doi:10.1038 / 277293a0. ISSN 1476-4687. S2CID 4270426.
- ^ Chisholm, Sallie W .; Frankel, Sheila L .; Goericke, Ralf; Olson, Robert J .; Palenik, Brian; Waterbury, John B .; West-Johnsrud, Lisa; Zettler, Erik R. (1992-02-01). "Prochlorococcus marinus nov. Gen. Nov. Sp .: divinil klorofil a ve b içeren bir oksifototrofik deniz prokaryotu". Mikrobiyoloji Arşivleri. 157 (3): 297–300. doi:10.1007 / BF00245165. ISSN 0302-8933. S2CID 32682912.
- ^ Chisholm, Sallie W .; Olson, Robert J .; Zettler, Erik R .; Goericke, Ralf; Waterbury, John B .; Welschmeyer, Nicholas A. (Temmuz 1988). "Okyanusal öfotik bölgede bol miktarda bulunan yeni bir serbest yaşayan proklorofit". Doğa. 334 (6180): 340–343. Bibcode:1988Natur.334..340C. doi:10.1038 / 334340a0. ISSN 1476-4687. S2CID 4373102.
- ^ a b c d Marie D, Partensky F, Jacquet S, Vaulot D (Ocak 1997). "Nükleik Asit Boyası SYBR Green I Kullanılarak Akış Sitometrisi ile Deniz Pikoplanktonunun Doğal Popülasyonlarının Numaralandırılması ve Hücre Döngüsü Analizi". Uygulamalı ve Çevresel Mikrobiyoloji. 63 (1): 186–93. doi:10.1128 / AEM.63.1.186-193.1997. PMC 1389098. PMID 16535483.
- ^ Giorgio PA, Bird DF, Prairie YT, Planas D (Haziran 1996). "Yeşil nükleik asit lekesi SYTO 13 ile göl planktonundaki bakteri bolluğunun akış sitometrik tayini". Limnoloji ve Oşinografi. 41 (4): 783–789. Bibcode:1996LimOc..41..783G. doi:10.4319 / lo.1996.41.4.0783.
- ^ Rice J, Kızak MA, Burkill PH, Tarran GA, O'connor CD, Zubkov MV (Mart 1997). "Türe Özgü Floresan Oligonükleotid Problarının Deniz Nanoflagellatlarının rRNA'sına Hibridizasyon Özelliklerinin Akış Sitometrik Analizi". Uygulamalı ve Çevresel Mikrobiyoloji. 63 (3): 938–44. doi:10.1128 / AEM.63.3.938-944.1997. PMC 1389123. PMID 16535558.
- ^ a b c Hagström A, Larsson U, Hörstedt P, Normark S (Mayıs 1979). "Hücreleri bölme sıklığı, su ortamlarında bakteri üreme oranlarının belirlenmesine yeni bir yaklaşım". Uygulamalı ve Çevresel Mikrobiyoloji. 37 (5): 805–12. doi:10.1128 / AEM.37.5.805-812.1979. PMC 243306. PMID 16345378.
- ^ Newell SY, Christian RR (Temmuz 1981). "Bakteri üretkenliğinin tahmin edicisi olarak hücre bölünme sıklığı". Uygulamalı ve Çevresel Mikrobiyoloji. 42 (1): 23–31. doi:10.1128 / AEM.42.1.23-31.1981. PMC 243955. PMID 16345812.
- ^ a b c d Servais P, Martinez J, Billen G, Vives-Rego J (Ağustos 1987). "Bakteriyoplankton DNA'sına [H] Timidin Katılmasının Belirlenmesi: Yöntemin DNaz Tedavisi ile İyileştirilmesi". Uygulamalı ve Çevresel Mikrobiyoloji. 53 (8): 1977–9. doi:10.1128 / AEM.53.8.1977-1979.1987. PMC 204039. PMID 16347424.
- ^ Bell R, Ahlgren G, Ahlgren I (Haziran 1983). "Ötrofik İsveç Gölünde [3H] timidin Birleşimini Ölçerek Bakteriyoplankton Üretiminin Tahmin Edilmesi". Uygulamalı ve Çevresel Mikrobiyoloji. 45 (6): 1709–1721. doi:10.1128 / AEM.45.6.1709-1721.1983. PMC 242528. PMID 16346304.
- ^ Fuhrman J, Azam F (Temmuz 1980). "British Columbia, Kanada, Antarktika ve Kaliforniya, ABD kıyı suları için bakterioplankton ikincil üretim tahminleri". Uygulamalı ve Çevresel Mikrobiyoloji. 39 (6): 1085–1095. doi:10.1128 / AEM.39.6.1085-1095.1980. PMC 291487. PMID 16345577.
- ^ a b c d Kirchman D, K'nees E, Hodson R (Mart 1985). "Lösin birleşimi ve doğal su sistemlerinde bakteriler tarafından protein sentezinin bir ölçüsü olarak potansiyeli". Uygulamalı ve Çevresel Mikrobiyoloji. 49 (3): 599–607. doi:10.1128 / AEM.49.3.599-607.1985. PMC 373556. PMID 3994368.
- ^ a b c d e Bunse C, Pinhassi J (Haziran 2017). "Deniz Bakteriyoplankton Mevsimsel Aktarım Dinamikleri". Mikrobiyolojideki Eğilimler. 25 (6): 494–505. doi:10.1016 / j.tim.2016.12.013. PMID 28108182.
- ^ Fuhrman JA, Hewson I, Schwalbach MS, Steele JA, Brown MV, Naeem S (Ağustos 2006). "Her yıl tekrarlayan bakteri toplulukları okyanus koşullarından tahmin edilebilir". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 103 (35): 13104–9. Bibcode:2006PNAS..10313104F. doi:10.1073 / pnas.0602399103. PMC 1559760. PMID 16938845.
- ^ Pinhassi J, Hagström Å (2000-06-15). "Deniz bakteriyoplanktonunda mevsimsel ardışık". Sucul Mikrobiyal Ekoloji. 21 (3): 245–256. doi:10.3354 / ame021245.
- ^ Pinhassi J, Gómez-Consarnau L, Alonso-Sáez L, Sala MM, Vidal M, Pedrós-Alió C, Gasol JM (2006-10-10). "Bakteriyoplankton besin sınırlamasındaki mevsimsel değişiklikler ve bunların Kuzeybatı Akdeniz'de bakteri topluluğu kompozisyonu üzerindeki etkileri". Sucul Mikrobiyal Ekoloji. 44 (3): 241–252. doi:10.3354 / ame044241.
- ^ Sapp M, Wichels A, Wiltshire KH, Gerdts G (Mart 2007). "Kuzey Denizi'nde kış-ilkbahar geçişi sırasında bakteri topluluğu dinamikleri". FEMS Mikrobiyoloji Ekolojisi. 59 (3): 622–37. doi:10.1111 / j.1574-6941.2006.00238.x. PMID 17381518.
- ^ Gilbert JA, Field D, Swift P, Newbold L, Oliver A, Smyth T, Somerfield PJ, Huse S, Joint I (Aralık 2009). "Batı İngiliz Kanalı'ndaki mikrobiyal toplulukların mevsimsel yapısı" (PDF). Çevresel Mikrobiyoloji. 11 (12): 3132–9. doi:10.1111 / j.1462-2920.2009.02017.x. hdl:1912/3133. PMID 19659500.
- ^ Gilbert JA, Steele JA, Caporaso JG, Steinbrück L, Reeder J, Temperton B, Huse S, McHardy AC, Knight R, Joint I, Somerfield P, Fuhrman JA, Field D (Şubat 2012). "Mevsimsel deniz mikrobiyal topluluk dinamiklerini tanımlama". ISME Dergisi. 6 (2): 298–308. doi:10.1038 / ismej.2011.107. PMC 3260500. PMID 21850055.
- ^ Riemann L, Steward GF, Azam F (Şubat 2000). "Bir mezokozm diatom patlaması sırasında bakteri topluluğu bileşimi ve aktivitesinin dinamikleri". Uygulamalı ve Çevresel Mikrobiyoloji. 66 (2): 578–87. doi:10.1128 / AEM.66.2.578-587.2000. PMC 91866. PMID 10653721.
- ^ a b Buchan A, LeCleir GR, Gulvik CA, González JM (Ekim 2014). "Usta geri dönüştürücüler: fitoplankton çoğalmalarıyla ilişkili bakterilerin özellikleri ve işlevleri". Doğa Yorumları. Mikrobiyoloji. 12 (10): 686–98. doi:10.1038 / nrmicro3326. PMID 25134618. S2CID 26684717.
- ^ a b c d Lindh MV, Sjöstedt J, Andersson AF, Baltar F, Hugerth LW, Lundin D, Muthusamy S, Legrand C, Pinhassi J (Temmuz 2015). "Mevsimsel bakterioplankton popülasyon dinamiklerinin yüksek frekanslı örnekleme ile çözülmesi". Çevresel Mikrobiyoloji. 17 (7): 2459–76. doi:10.1111/1462-2920.12720. PMID 25403576.
- ^ Alderkamp A, Sintes E, Herndl GJ (2006-12-21). "İlkbahar ve yaz aylarında Kuzey Denizi kıyılarındaki başlıca prokaryotik plankton gruplarının bolluğu ve aktivitesi". Sucul Mikrobiyal Ekoloji. 45 (3): 237–246. doi:10.3354 / ame045237.
- ^ Fernández-Gómez B, Richter M, Schüler M, Pinhassi J, Acinas SG, González JM, Pedrós-Alió C (Mayıs 2013). "Deniz Bacteroidetes Ekolojisi: karşılaştırmalı bir genomik yaklaşım". ISME Dergisi. 7 (5): 1026–37. doi:10.1038 / ismej.2012.169. PMC 3635232. PMID 23303374.
- ^ Teeling H, Fuchs BM, Becher D, Klockow C, Gardebrecht A, Bennke CM, Kassabgy M, Huang S, Mann AJ, Waldmann J, Weber M, Klindworth A, Otto A, Lange J, Bernhardt J, Reinsch C, Hecker M , Peplies J, Bockelmann FD, Callies U, Gerdts G, Wichels A, Wiltshire KH, Glöckner FO, Schweder T, Amann R (Mayıs 2012). "Bir fitoplankton patlamasıyla indüklenen deniz bakteriyoplankton popülasyonlarının substrat kontrollü ardışıklığı". Bilim. 336 (6081): 608–11. Bibcode:2012Sci ... 336..608T. doi:10.1126 / science.1218344. PMID 22556258. S2CID 29249533.
- ^ Teeling H, Fuchs BM, Bennke CM, Krüger K, Chafee M, Kappelmann L, Reintjes G, Waldmann J, Quast C, Glöckner FO, Lucas J, Wichels A, Gerdts G, Wiltshire KH, Amann RI (Nisan 2016). "Kıyılardaki bahar yosunlarının çoğalması sırasında bakterioplankton dinamiklerinde tekrarlayan modeller". eLife. 5: e11888. doi:10.7554 / eLife.11888. PMC 4829426. PMID 27054497.
- ^ Taylor JD, Cottingham SD, Billinge J, Cunliffe M (Ocak 2014). "Mevsimsel mikrobiyal topluluk dinamikleri, yüzey kıyı sularında fitoplankton kaynaklı polisakkaritlerle ilişkilidir". ISME Dergisi. 8 (1): 245–8. doi:10.1038 / ismej.2013.178. PMC 3869024. PMID 24132076.
- ^ Agawin NS, Duarte CM, Agustí S (1998-09-03). "Bir Akdeniz Körfezinde Synechococcus sp.'nin büyümesi ve bolluğu: mevsimsellik ve sıcaklıkla ilişki". Deniz Ekolojisi İlerleme Serisi. 170: 45–53. Bibcode:1998MEPS..170 ... 45A. doi:10.3354 / meps170045.
- ^ Alonso-Sáez L, Balagué V, Sà EL, Sánchez O, González JM, Pinhassi J, Massana R, Pernthaler J, Pedrós-Alió C, Gasol JM (Nisan 2007). "Kuzeybatı Akdeniz kıyı sularında bakteri çeşitliliğindeki mevsimsellik: klon kütüphaneleri, parmak izi ve FISH aracılığıyla değerlendirme". FEMS Mikrobiyoloji Ekolojisi. 60 (1): 98–112. doi:10.1111 / j.1574-6941.2006.00276.x. PMID 17250750.
- ^ Salter I, Galand PE, Fagervold SK, Lebaron P, Obernosterer I, Oliver MJ, Suzuki MT, Tricoire C (Şubat 2015). "Oligotrofik Kuzeybatı Akdeniz'deki aktif SAR11 ekotiplerinin mevsimsel dinamikleri". ISME Dergisi. 9 (2): 347–60. doi:10.1038 / ismej.2014.129. PMC 4303628. PMID 25238399.
- ^ Hugoni M, Taib N, Debroas D, Domaizon I, Jouan Dufournel I, Bronner G, Salter I, Agogué H, Mary I, Galand PE (Nisan 2013). "Nadir arkeal biyosferin yapısı ve yüzey kıyı sularındaki aktif ekotiplerin mevsimsel dinamikleri". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 110 (15): 6004–9. Bibcode:2013PNAS..110.6004H. doi:10.1073 / pnas.1216863110. PMC 3625260. PMID 23536290.