Heterodontosauridae - Heterodontosauridae - Wikipedia
| Heterodontosauridler | |
|---|---|
 | |
| SAM-PK-K1332 örnek döküm Heterodontosaurus tucki, Kaliforniya Üniversitesi Paleontoloji Müzesi | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Sipariş: | †Ornithischia |
| Aile: | †Heterodontosauridae Romer, 1966 |
| Alt gruplar | |
| Eş anlamlı | |
| |
Heterodontosauridae bir aile erken ornitorik dinozorlar bu muhtemelen en çok baz alınan grubun (ilkel) üyeleri. Onların fosiller nispeten nadirdir ve grupları sayıca azdır, hariç tüm kıtalarda yaşadılar Avustralya yaklaşık 100 milyon yıldır Geç Triyas için Erken Kretase.
Heterodontosauridler tilki Uzun kuyruk dahil 2 metreden (6,6 fit) daha kısa boyutlu dinozorlar. Çoğunlukla genişlemiş dişler dahil karakteristik dişleriyle bilinirler. köpek dişler ve yanak dişleri çiğnemeye uygun hale getirilmiş, benzer Kretase'dekilere hadrosauridler. Diyetleri otçul veya muhtemelen her yerde yaşayan.
Açıklama


Heterodontozoridler arasında sadece Heterodontozorlar kendisi tam bir iskeletten bilinmektedir. Parçalı iskelet kalıntıları Abrictosaurus diğer birçok heterodontosaurid sadece çene parçalarından ve dişlerden bilinirken biliniyor ancak tam olarak tanımlanmadı. Sonuç olarak, çoğu heterodontosaurid sinapomorfiler (tanımlayıcı özellikler) dişlerden ve çene kemiklerinden tarif edilmiştir.[2][3] Heterodontozorlar 1 metreden (3,3 ft) biraz daha uzun ölçülmüş,[4] parça parça kalırken Lycorhinus daha büyük bir bireyi gösterebilir.[5]
Tianyulong Bazı theropodlarda bulunan proto-tüylerin bir varyantı olarak yorumlanan ipliksi bütünlüğü koruyor gibi görünüyor. Bu iplikler, kuyruğu boyunca bir tepe içerir. Bu ipliksi bütünlüğün varlığı, hem ornitoşluların hem de sauriskianların endotermik.[6]
Kafatası ve dişler
Her ikisi de Abrictosaurus ve Heterodontozorlar çok büyük gözleri vardı. Gözlerin altında jugal kemik yanlara doğru yansıtılan bir özellik, aynı zamanda Ceratopsia'lılar. Çoğu ornitosçunun çenesinde olduğu gibi, ön Kenarı premaksilla (üst çenenin ucundaki bir kemik) dişsizdi ve muhtemelen keratinli gaga (Rhamphotheca ), heterodontosauridlerin dişleri olmasına rağmen arka premaksilla bölümü. A denen büyük bir boşluk diastema, bu premaksiller dişleri dişlerin dişlerinden ayırdı. üst çene (ana üst çene kemiği) pek çok ornitoskiyende, ancak bu diastema karakteristik olarak heterodontosauridlerde kavisliydi. çene (alt çene) tarafından eğildi öncül ornitoriklere özgü bir kemik. Bu kemik ayrıca premaksillada bulunana benzer bir gagayı da destekledi. Alt çenede bulunan tüm dişler diş hekimi kemik.[2]
Heterodontosauridler, güçlü oldukları için adlandırılır. heterodont dişlenme. Üç adet premaksiller diş vardı. İçinde Erken Jura Abrictosaurus, Heterodontozorlar, ve Lycorhinusİlk iki premaksiller diş küçük ve koni şeklindeyken, çok daha büyük olan üçüncü dişin köpek dişlerine benziyordu. etobur memeliler ve genellikle caniniform veya "savunma dişi" olarak adlandırılır. Yukarıdan daha büyük olan bir alt kaniniform, diş hekimliğinde ilk pozisyonu aldı ve ağız kapalıyken üst çenenin kemerli diasteması tarafından yerleştirildi.[2] Bu kaniniformlar, hem ön hem de arka kenarlarda tırtıklıydı. Heterodontozorlar ve Lycorhinus, bunlar Abrictosaurus sadece ön kenarda çentikler deldi.[7][8] İçinde Erken Kretase EchinodonPremaksilla yerine maksillada bulunan iki üst kaniniform olabilir,[9] ve Fruitadens -den Geç Jura her dişte iki alt kaniniformu olabilir.[10][11]

Karakteristik dişler gibi, yanak dişleri türetilmiş heterodontosauridler de erken ornitozanlar arasında benzersizdi. Küçük sırtlar veya dişler, bitki örtüsünü kırpmak için ornitorik yanak dişlerinin kenarlarını sıraladı. Bu dentiküller, tüm heterodontosauridlerde diş taçının ucundan aşağıya doğru yolun yalnızca üçte birini uzatır; diğer ornitoriklerde, dişler köke doğru daha da aşağıya uzanır. Bazal formlar Abrictosaurus hem maksilla hem de diş dişlerinde genellikle diğer ornitoriklere benzer yanak dişleri vardı: geniş aralıklı, her biri alçak bir krona ve kronu kökünden ayıran güçlü bir çıkıntıya (cingulum) sahipti. Daha fazlası türetilmiş gibi formlar Lycorhinus ve Heterodontozorlardişler keski şeklindeydi daha yüksek kronlar ve cingula yok, böylece kronlar ve kökler arasında genişlik farkı yoktu.[2]
Bu türetilmiş yanak dişleri üst üste biniyordu, böylece kronları yiyeceklerin çiğnenebileceği sürekli bir yüzey oluşturuyordu. Diş sıraları ağzın kenarından hafifçe içeri girerek dişlerin dışında kas tarafından sınırlanmış olabilecek bir boşluk bırakıldı. yanak çiğnemek için gerekli olacaktı. Kretase Dönemi hadrosaurları ve ceratopsianları ile birçok otçul memelinin yakınsak gelişmek biraz benzer diş pilleri. Yüzlerce dişin sürekli değiştirildiği hadrosaurların aksine, heterodontosauridlerde diş değişimi çok daha yavaş gerçekleşti ve beklemede tek bir yedek diş olmadan birkaç örnek bulundu. Karakteristik olarak, heterodontosauridler, diğer ornitoriklerin çoğunda diş gelişimine yardımcı olduğu düşünülen çene kemiklerinin iç tarafındaki küçük açıklıklardan (foramina) yoksundu. Heterodontosaurids ayrıca benzersiz bir küremsi dişler ve ön diş arasındaki eklem, ağız kapalıyken alt çenelerin dışa doğru dönmesine izin vererek yanak dişlerini birbirine karşı gıcırdatır. Yavaş değiştirme oranı nedeniyle, bu taşlama, kronların artan yüksekliği her bir dişe uzun bir ömür vermesine rağmen, genellikle daha yaşlı dişlerdeki diş etlerinin çoğunu yok eden aşırı diş aşınmasına neden oldu.[12]
İskelet

Postkraniyal anatomisi Heterodontosaurus tucki iyi tanımlanmış olmasına rağmen H. tucki Genellikle Erken Jura heterodontosauridlerinin en çok türevi olarak kabul edilir, bu nedenle özelliklerinin kaçının diğer türlerle paylaşıldığını bilmek imkansızdır.[2] Ön ayaklar bir dinozor için uzundu, arka bacakların uzunluğunun% 70'inden fazlası. İyi gelişmiş deltopektoral kret (eklenmesi için bir sırt göğüs ve omuz kaslar) humerus ve öne çıkan olekranon süreci (nerede ön kolu uzatan kaslar eklendi) ulna ön ayağın da güçlü olduğunu gösterir. Beş basamak vardı Manus ('el'). İlki büyüktü, keskin bir şekilde kavisli bir pençe ile uçluydu ve esnetildiğinde içe doğru dönüyordu; Robert Bakker buna "bükülme-başparmak" adını verdi.[13] İkinci rakam en uzundu, üçüncü rakamdan biraz daha uzundu. Bu rakamların her ikisi de pençelere sahipken, pençesiz dördüncü ve beşinci rakamlar karşılaştırıldığında çok küçük ve basitti. Arka bacakta tibia % 30 daha uzundu uyluk, genellikle hız için bir adaptasyon olarak kabul edilir. Tibia ve fibula bacağın alt kısmının astragalus ve kalkaneum ayak bileğinintibiofibiotarsus 'modern ile yakınsak kuşlar. Ayrıca kuşlara benzer şekilde, daha düşük tarsal (ayak bileği) kemikler ve metatarsallar bir 'oluşturmak için kaynaştıtarsometatarsus. ' İçinde dört hane vardır pes (arka ayak), sadece ikinci, üçüncü ve dördüncü yerle temas eder. Kuyruk, diğer pek çok ornitosçunun aksine, kemikleşmiş tendonlar sert bir duruş sürdürmek için ve muhtemelen esnekti.[4] Parçalı iskelet Abrictosaurus ön ayaklar ve manus, daha küçük olmasına rağmen, hiçbir zaman tam olarak tarif edilmemiştir. Heterodontozorlar. Ayrıca, ön ayağın dördüncü ve beşinci rakamlarının her birinde bir tane daha az bulunur falanks kemiği.[14]
Sınıflandırma

Güney Afrikalı paleontolog Robert Süpürge adı yarattı Geranosaurus 1911'de dinozor çene kemikleri için tüm dişleri ve bazı kısmi ilişkili uzuv kemiklerini kaybetti.[15] 1924'te, Lycorhinus olarak adlandırıldı ve bir olarak sınıflandırıldı Cynodont, tarafından Sidney Haughton.[16] Heterodontozorlar 1962'de seçildi ve Lycorhinus ve Geranosaurus yakından ilişkili ornitorik dinozorlar olarak kabul edildi.[17] Alfred Romer 1966'da Heterodontosauridae olarak adlandırılan ornithischian dinozorları ailesi olarak adlandırıldı. Heterodontozorlar ve Lycorhinus.[18] Olarak tanımlandı clade tarafından 1998'de Paul Sereno[19] 2005 yılında kendisi tarafından şunlardan oluşan gövde grubu olarak yeniden tanımlanmıştır. Heterodontosaurus tucki ve tüm türler ile daha yakından ilişkili Heterodontozorlar daha Parasaurolophus walkeri, Pachycephalosaurus wyomingensis, Triceratops horridus veya Ankylosaurus magniventris.[20]
Heterodontosauridae cinsleri içerir Abrictosaurus, Lycorhinus, ve Heterodontozorlarhepsi Güney Afrika'dan. Süre Richard Thulborn bir kez üçü de yeniden atandı Lycorhinus,[14] diğer tüm yazarlar bu üç cinsi farklı kabul eder.[8] Aile içinde Heterodontozorlar ve Lycorhinus dikkate alındı kardeş takson, ile Abrictosaurus bazal üye olarak.[3] Geranosaurus aynı zamanda bir heterodontosauriddir, ancak genellikle bir nomen dubium çünkü tip numunenin tüm dişleri eksiktir, bu da onu ailedeki diğer türlerden ayırt edilemez hale getirir.[2] Daha yakın zamanlarda cins Echinodon çeşitli çalışmalarda bir heterodontosaurid olarak kabul edilmiştir.[9][10] Lanasaurus 1975'te bir üst çene için seçildi,[21] ancak daha yeni keşifler, Lycorhinus bunun yerine yapmak Lanasaurus a küçük eşanlamlı bu cinsin.[5] Dianchungosaurus bir zamanlar bir heterodontosaurid olarak kabul edildi Asya,[22] ama o zamandan beri kalıntıların bir kimera nın-nin prosauropod ve mezoeucrocodylian kalır.[23] José Bonaparte Güney Amerikalıları da sınıflandırdı Pisanosaurus bir seferde heterodontosaurid olarak,[24] ancak bu hayvanın artık daha bazal ornitorik olduğu biliniyor.[25]

Heterodontosauridae üyeliği, belirsizliğine kıyasla sağlam bir şekilde kurulmuştur. filogenetik durum. Birkaç erken çalışma, heterodontosauridlerin çok ilkel ornitorikler olduğunu öne sürdü.[4][17] Ön ayakların morfolojisindeki varsayılan benzerlikler nedeniyle, Robert Bakker, heterodontosauridler ve erken dönemler arasında bir ilişki önerdi. Sauropodomorflar sevmek Anchisaurus, emirleri birleştirmek Saurischia ve Ornithischia.[13] Son birkaç on yıldaki baskın hipotez, heterodontosauridleri bazal ornitopodlar olarak yerleştirdi.[2][3][7][26] Bununla birlikte, diğerleri heterodontosauridlerin bunun yerine Marginocephalia (ceratopsians ve pachycephalosaurs ),[27][28] 21. yüzyılın başlarında yapılan bazı çalışmalarda destek bulan bir hipotez.[29][30] Heterodontosauridler ve marjinosefaliler içeren sınıf seçildi Heterodontosauriformes.[31] Heterodontosauridler, hem ornitopodlar hem de marjinosefaliler için bazal olarak görülmüştür.[32][33] 2007 yılında kladistik analiz, heterodontosauridlerin bilinen tüm ornitosauridler için bazal olduğunu ileri sürdü. Pisanosaurus, aile üzerindeki en eski çalışmaların bazılarını yansıtan bir sonuç.[34][35] Bununla birlikte, Bonaparte tarafından yapılan bir araştırma, Pisanosauridae'nin Heterodontosauridae ile eşanlamlı olduğunu ve kendi başına ayrı bir aile olmadığını buldu. Pisanosaurus bir heterodontosaur olarak.[36] Butler vd. (2010), Heterodontosauridae'yi bilinen en önemli ornitorik radyasyon olarak buldu.[37]
kladogram Aşağıda, Heterodontosauridae içindeki karşılıklı ilişkiler gösterilmektedir ve Sereno, 2012 tarafından yapılan analizi takip etmektedir:[38]
| Heterodontosauridae |
| |||||||||||||||||||||||||||||||||||||||
Bir 2020 yeniden çalışması Cerapoda Dieudonné tarafından et al. geleneksel, kubbe başlı pachycephalosaurlara göre Pachycephalosauria içinde bir bazal grup olarak 'heterodontosauridler' olarak kabul edilen hayvanları buldu.[39]

Dağıtım

Başlangıçta yalnızca Güney Afrika'nın Erken Jura döneminden bilinirken, heterodontosaurid kalıntıları artık dört kıtalar. Heterodontosaurid tarihinin başlarında, süper kıta Pangea hala büyük ölçüde bozulmamıştı ve ailenin neredeyse tüm dünyaya dağıtım.[9] Bilinen en eski olası heterodontosaurid kalıntıları, bir çene parçası ve izole dişlerdir. Laguna Colorada Formasyonu nın-nin Arjantin hangi tarihe kadar uzanır Geç Triyas. Bu kalıntıların benzer bir türetilmiş morfolojisi vardır. Heterodontozorlar, hem ön hem de arka kenarlarda tırtıklı bir kaniniform ve ayrıca singulumu olmayan yüksek taçlı maksiller dişler dahil.[40] Irmis et al. (2007), bu fosil materyalin bir heterodontosauridi temsil ettiğini geçici olarak kabul etti, ancak bu atamayı doğrulamak için ek materyale ihtiyaç duyulduğunu, çünkü örneğin kötü bir şekilde korunduğunu belirtti.[41] Sereno (2012) ise sadece bu materyalin bir ornitosiyeni veya hatta özellikle bir heterodontosauridi temsil edebileceğini belirtmiştir.[42] Olsen Kent & Whiteside (2010), Laguna Colorada Formasyonunun yaşının zayıf bir şekilde sınırlandırıldığını ve bu nedenle bu formasyondaki varsayılan heterodontosauridin Triyas veya Jura çağında olup olmadığının kesin olarak belirlenmediğini belirtti.[43] En çeşitli heterodontosaurid fauna fosillerin bulunduğu güney Afrika'nın Erken Jura'dan geliyor Heterodontozorlar, Abrictosaurus, Lycorhinusve şüpheli Geranosaurus bulunan.[2]
Tanımlanmamış Erken Jura heterodontosauridleri de Amerika Birleşik Devletleri[44] ve Meksika,[45] sırasıyla. Ek olarak, 1970'lerden başlayarak, Geç Jura dönemine ait çok sayıda fosil materyali keşfedildi. Morrison Formasyonu yakın Fruita, Colorado Birleşik Devletlerde.[10] 2009 yılında baskıda açıklanan bu materyal, cinsin içine yerleştirildi. Fruitadens.[11] Bir singulumu olmayan heterodontozorid dişler, aynı zamanda Geç Jura ve Erken Kretase oluşumlarından da tanımlanmıştır. ispanya ve Portekiz.[46] Kalıntıları Echinodon 2002'de yeniden tanımlandı ve bu, geç hayatta kalan bir heterodontosauridi temsil edebileceğini gösteriyor. Berriasiyen Güney İngiltere'deki Erken Kretase aşaması.[9] Dianchungosaurus Çin'in Erken Jura'sından artık bir heterodontosaurid olarak kabul edilmiyor; Geç kalan bir Asya formu bilinmesine rağmen (Tianyulong ).[6]
Paleobiyoloji

Heterodontosaurid fosillerinin çoğu, jeolojik oluşumlar kurak ve yarı kurak ortamları temsil eden Üst Elliot Formasyonu nın-nin Güney Afrika ve Purbeck Yatakları Güney İngiltere.[3] Heterodontosauridlerin mevsimsel geçirdiği öne sürülmüştür. doğum veya kış uykusu yılın en kurak zamanlarında. Çoğu heterodontosauridde yedek diş eksikliği nedeniyle, dişlerin sürekli ve ara sıra değiştirilmesinin diş sırasının işlevini tek bir çiğneme yüzeyi olarak kesintiye uğratacağı görüldüğünden, bu hareketsiz dönemde tüm diş setinin değiştirilmesi önerildi. .[14] Ancak bu, heterodontosaurid çene mekaniğinin yanlış anlaşılmasına dayanıyordu.[47] Heterodontosauridlerin aslında dişlerini diğerlerinden daha yavaş olsa da sürekli olarak değiştirdiği düşünülüyordu. sürüngenler, fakat CT taraması genç ve olgun kafataslarının Heterodontozorlar yedek diş göstermez.[48] Şu anda bunu destekleyen hiçbir kanıt yok hipotez heterodontosauridlerde aestivasyon,[2] ancak kafatası taramalarına göre reddedilemez.[48]
Heterodontozoridlerin yanak dişleri, sert bitki materyalini öğütmek için açıkça uyarlanmış olsa da, diyetleri omnivor olabilir. Sivri ön-maksiller dişler ve ön ayaklardaki keskin, kavisli pençeler bir dereceye kadar yırtıcı davranışa işaret ediyor. Uzun, güçlü ön ayaklarının Heterodontozorlar modern modellere benzer şekilde böcek yuvalarını yırtmak için yararlı olabilirdi. karıncayiyenler. Bu ön ayaklar aynı zamanda kazma araçları olarak da işlev görmüş olabilir. kökler ve yumrular.[2]
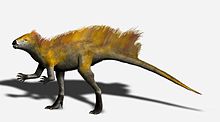
Arka ekstremiteye kıyasla ön ayağın uzunluğu şunu göstermektedir: Heterodontozorlar kısmen olabilirdi dört ayaklı ve öne çıkan olekranon süreci ve ön ayağın aşırı uzatılabilir rakamları birçok dört ayaklıda bulunur. Bununla birlikte, manüs açıkça ağırlık desteği için değil, kavrama için tasarlanmıştır. Uzun tibia ve ayağın yanı sıra tibiofibiotarsus ve tarsometatarsus füzyonu da dahil olmak üzere arka bacağın birçok özelliği, heterodontosauridlerin arka ayaklar üzerinde hızlı bir şekilde çalışacak şekilde adapte edildiğini gösterir, bu nedenle olası değildir Heterodontozorlar belki beslenme zamanı dışında dört uzuvda da hareket etti.[4]
Bilinen tüm heterodontozoridlerde bulunan kısa dişler, günümüzde bulunan dişlere büyük ölçüde benzer. misk geyiği, pekari ve domuzlar. Bu hayvanların çoğunda (hem de uzun dişli mors ve Asya filleri ), bu bir cinsel olarak dimorfik trait, sadece erkeklerde bulunan dişlerle. tip numune nın-nin Abrictosaurus dişleri yoktur ve başlangıçta dişi olarak tanımlanmıştır.[14] Bu mümkün olmaya devam ederken, kullanılmayan sakral omur ve kısa yüz, bu örneğin genç bir hayvanı temsil ettiğini göstermektedir. Başlangıçta ait olduğu önerilen ikinci, daha büyük bir örnek Abrictosaurus Dişlerin sadece yetişkinlerde bulunduğu fikrini desteklemek için kullanılan dişlere sahip olduğu açıkça görülüyor. ikincil cinsel özellik erkeklerin. Bu dişler, savaş için kullanılmış olabilir veya Görüntüle aynı türün üyeleriyle veya diğer türlerle.[2] Gençlikte dişlerin olmaması Abrictosaurus gençlerde dişler bilindiği için onu diğer heterodontozoridlerden ayıran başka bir özellik de olabilir. Heterodontozorlar. Dişler için önerilen diğer işlevler arasında savunma ve ara sıra omnivor diyette kullanım yer alır.[48] Ancak, bu örnek alternatif olarak yeniden atandı Lycorhinus Halihazırda dişlere sahip olduğu ve bu nedenle bunların yokluğu bilinen 2012'de Sereno tarafından Abrictosaurus yaşın bir sonucu olmayabilir.[42]
2005 yılında, 200 milyon yıldan daha eski, fosilleşmiş küçük bir heterodontosaurid iskeleti keşfedildi. Güney Afrika. Temmuz 2016'da Güney Afrikalı araştırmacılardan oluşan bir ekip tarafından Avrupa Sinkrotron Radyasyon Tesisi; dişlenme taraması, bir milimetreden daha ince damak kemiklerini ortaya çıkardı.[49]
Referanslar
- ^ R.A. Thulborn, 1971, "Ornithischian dinozorlarının kökenleri ve evrimi", Doğa 234(5324): 75-78
- ^ a b c d e f g h ben j k Weishampel, David B.; Witmer, Lawrence M. (1990). "Heterodontosauridae". Weishampel, David B .; Dodson, Peter; Osmólska, Halszka (editörler). Dinosauria. Berkeley: California Üniversitesi Yayınları. sayfa 486–497. ISBN 978-0-520-06727-1.
- ^ a b c d Norman, David B.; Sues, Hans-Dieter; Witmer, Lawrence M .; Coria, Rodolfo A. (2004). "Bazal Ornitopoda". Weishampel, David B .; Dodson, Peter; Osmólska, Halszka (editörler). Dinosauria (İkinci baskı). Berkeley: California Üniversitesi Yayınları. s. 393–412. ISBN 978-0-520-24209-8.
- ^ a b c d Santa Luca, Albert P. (1980). "Postkraniyal iskelet Heterodontosaurus tucki (Reptilia, Ornithischia) Güney Afrika'nın Stormberg'inden ". Güney Afrika Müzesi Yıllıkları. 79 (7): 159–211.
- ^ a b Gow, Christopher E. (1990). "Diş taşıyan bir çene Lycorhinus angustidens Haughton, 1924 (Dinosauria, Ornithischia) ". Güney Afrika Müzesi Yıllıkları. 99 (10): 367–380.
- ^ a b Zheng, Xiao-Ting; Sen, Hai-Lu; Xu, Xing; Dong, Zhi-Ming (19 Mart 2009). "İpliksi bütünleşik yapılara sahip bir Erken Kretase heterodontosaurid dinozor". Doğa. 458 (7236): 333–336. doi:10.1038 / nature07856. PMID 19295609. S2CID 4423110.
- ^ a b Thulborn Richard A. (1970). "Triyas dönemine ait ornitorik dinozorun sistematik konumu Lycorhinus angustidens". Linnean Society'nin Zooloji Dergisi. 49 (3): 235–245. doi:10.1111 / j.1096-3642.1970.tb00739.x.
- ^ a b Hopson, James A. (1975). "Ornitorik dinozorların genel ayrımı üzerine Lycorhinus ve Heterodontozorlar Güney Afrika Stormberg Serisinden (Yukarı Triyas) ". Güney Afrika Bilim Dergisi. 71: 302–305.
- ^ a b c d Norman, David B.; Barrett, Paul M. (2002). "İngiltere'nin Alt Kretase (Berriasiyen) bölgesinden Ornitoş dinozorları". Milner, Andrew'da; Batten, David J. (editörler). Purbeck Times'da Yaşam ve Ortamlar. Paleontolojide Özel Makaleler 68. Londra: Paleontoloji Derneği. s. 161–189. ISBN 978-0-901702-73-9.
- ^ a b c Galton, Peter M. (2007). "Batı Amerika Birleşik Devletleri Morrison Formasyonundan (Üst Jura) ornithischian dinozorlarının (çoğunlukla Ornithopoda) dişleri." Carpenter, Kenneth (ed.). Boynuzlar ve Gagalar: Ceratopsian ve Ornitopod Dinozorları. Bloomington: Indiana University Press. sayfa 17–47. ISBN 978-0-253-34817-3.
- ^ a b Butler, Richard J .; Galton, Peter M .; Porro, Laura B .; Chiappe, Luis M .; Henderson, D. M .; Erickson, Gregory M. (2010). "Kuzey Amerika'dan yeni bir Üst Jurassic heterodontosaurid'den çıkarılan ornitosiyan dinozor vücut boyutunun alt sınırları" (PDF). Royal Society B Tutanakları. 277 (1680): 375–381. doi:10.1098 / rspb.2009.1494. PMC 2842649. PMID 19846460.
- ^ Weishampel, David B. (1984). Ornitopod dinozorlarda çene mekaniğindeki evrim. Anatomi, Embriyoloji ve Hücre Biyolojisindeki Gelişmeler. Anatomi, Embriyoloji ve Hücre Biyolojisindeki Gelişmeler 87. 87. Berlin; New York: Springer-Verlag. s. 1–109. doi:10.1007/978-3-642-69533-9_1. ISBN 978-0-387-13114-6. PMID 6464809.
- ^ a b Bakker, Robert T. (1986). Dinozor Heresies: Dinozorların Gizemini ve Yok Olmalarını Ortaya Çıkaran Yeni Teoriler. New York: William Morrow. s. 453 pp. ISBN 978-0-14-010055-6.
- ^ a b c d Thulborn Richard A. (1974). "Lesotho'nun Üst Triyas Kırmızı Yataklarından yeni bir heterodontosaurid dinozor (Reptilia: Ornithischia)". Linnean Society'nin Zooloji Dergisi. 55 (2): 151–175. doi:10.1111 / j.1096-3642.1974.tb01591.x.
- ^ Süpürge, Robert. (1911). "Güney Afrika'daki Stormberg'in dinozorları üzerine". Güney Afrika Müzesi Yıllıkları. 7: 291–308.
- ^ Haughton, Sidney H. (1924). "Stormberg Serisinin faunası ve stratigrafisi". Güney Afrika Müzesi Yıllıkları. 12: 323–497.
- ^ a b Crompton, A.W .; Charig, Alan (1962). "Güney Afrika'nın Yukarı Triyasından yeni bir ornitoşçu". Doğa. 196 (4859): 1074–1077. doi:10.1038 / 1961074a0. S2CID 4198113.
- ^ Romer, Alfred S. (1966). Omurgalı Paleontoloji (Üçüncü baskı). Chicago: Chicago Press Üniversitesi. s.468 s. ISBN 978-0-7167-1822-2.
- ^ Sereno, Paul C. (1998). "Dinosauria'nın üst düzey taksonomisine uygulanarak filogenetik tanımlar için bir gerekçe". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 210 (1): 41–83. doi:10.1127 / njgpa / 210/1998/41.
- ^ Sereno, Paul C. (2005-11-07). "Kök Archosauria - TaxonSearch". Arşivlenen orijinal 2007-02-19 tarihinde. Alındı 2007-02-24.
- ^ Gow, Christopher E. (1975). "Güney Afrika'nın Kızıl Yataklarından yeni bir heterodontosaurid diş replasmanına dair açık kanıtlar gösteriyor". Linnean Society'nin Zooloji Dergisi. 57 (4): 335–339. doi:10.1111 / j.1096-3642.1975.tb01895.x.
- ^ Genç, C.C. (1982). "[Yunnan Eyaleti, Lufeng İlçesinden yeni bir dinozor cinsi]". Zhou M.'de (ed.). [Yang Zhongjian'ın toplanan eserleri] (Çin'de). Pekin: Academica Sinica. sayfa 38–42.
- ^ Barrett, Paul M .; Xu Xing (2005). "Yeniden değerlendirme Dianchungosaurus lufengensis Yang, 1982a, Çin Halk Cumhuriyeti, Yunnan Eyaletindeki Aşağı Lufeng Formasyonundan (Alt Jura) esrarengiz bir sürüngen ". Paleontoloji. 79 (5): 981–986. doi:10.1666 / 0022-3360 (2005) 079 [0981: ARODLY] 2.0.CO; 2. ISSN 0022-3360.
- ^ Bonaparte, Jose F. (1976). "Pisanosaurus mertii Casamiquela ve Ornithischia'nın kökeni ". Paleontoloji Dergisi. 50: 808–820.
- ^ Weishampel, David B.; Witmer, Lawrence M. (1990). "Lesothosaurus, Pisanosaurus, ve Technosaurus". Weishampel, David B .; Dodson, Peter; Osmólska, Halszka (editörler). Dinosauria (İlk baskı). Berkeley: California Üniversitesi Yayınları. s. 416–425. ISBN 978-0-520-06727-1.
- ^ Sereno, Paul C. (1986). "Kuş kalçalı dinozorların filogenisi". National Geographic Araştırma. 2 (2): 234–256.
- ^ Zhao Xijin (1983). "Filogeni ve Dinosauria'nın evrim aşamaları". Acta Palaeontologica Polonica. 28 (1–2): 295–306.
- ^ Cooper, Michael A. (1985). "Ornitosiyan dinozorunun bir revizyonu Kangnasaurus coetzeei Haughton, Ornithischia sınıflandırmasıyla ". Güney Afrika Müzesi Yıllıkları. 95: 281–317.
- ^ Zhao Xijin; Cheng Zhengwu; Xu Xing (1999). "Çin, Liaoning'deki Tuchengzi Formasyonundaki en eski ceratopsian". Omurgalı Paleontoloji Dergisi. 19 (4): 681–691. doi:10.1080/02724634.1999.10011181.
- ^ Sen Hailu; Xu Xing; Wang Xiaolin (2003). "Yeni bir Psittacosauridae (Dinosauria: Ornithopoda) cinsi ve marginosefalili dinozorların kökeni ve erken evrimi". Acta Geologica Sinica (İngilizce Sürümü). 77 (1): 15–20. doi:10.1111 / j.1755-6724.2003.tb00105.x.
- ^ Xu Xing, X; Forster, Catherine A .; Clark, James M .; Mo Jinyou. (2006). "Kuzeybatı Çin'in Geç Jura dönemine ait geçiş özelliklerine sahip bir bazal ceratopsia". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 273 (1598): 2135–2140. doi:10.1098 / rspb.2006.3566. PMC 1635516. PMID 16901832.
- ^ Maryanska, Teresa; Osmólska, Halszka. (1985). "Ornitorik soyoluş üzerine". Acta Palaeontologica Polonica. 30: 137–150.
- ^ Butler, Richard J. (2005). "Güney Afrika ve Lesoto'nun Üst Elliot Formasyonunun (Alt Jura) 'fabrosaurid' ornitorik dinozorları". Linnean Society'nin Zooloji Dergisi. 145 (2): 175–218. doi:10.1111 / j.1096-3642.2005.00182.x.
- ^ Butler, Richard J.; Smith, Roger M.H .; Norman, David B. (2007). "Güney Afrika'nın Geç Triyas döneminden ilkel bir ornitorik dinozor ve Ornithischia'nın erken evrimi ve çeşitliliği". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 274 (çevrimiçi yayınlanmıştır): 2041–6. doi:10.1098 / rspb.2007.0367. PMC 2275175. PMID 17567562.
- ^ Butler, Richard J .; Upchurch, Paul; Norman, David B. (2008). "Ornitorik dinozorların soyoluşu". Sistematik Paleontoloji Dergisi. 6 (1): 1–40. doi:10.1017 / S1477201907002271. S2CID 86728076.
- ^ Bonaparte, J.F. (1976). "Pisanosaurus mertii Casamiquela ve Ornithischia'nın kökeni". Journal of Paleontology. 50 (5): 808–820. JSTOR 1303575.
- ^ R. J. Butler. 2010. Güney Afrika'nın aşağı Elliot Formasyonundan (Geç Triyas) bazal ornitosiyen dinozor Eocursor parvus'un anatomisi. Linnean Society'nin Zooloji Dergisi 160: 648-684
- ^ Sereno, P.C. (2012). s. 193-206.
- ^ Dieudonné, P.E .; Cruzado-Caballero, P .; Godefroit, P .; Tortosa, T. (2020). "Cerapodan dinozorlarının yeni bir filogeni". Tarihsel Biyoloji: 1–21. doi:10.1080/08912963.2020.1793979.
- ^ Báez, Ana Maria; Marsicano Claudia A. (2001). "Patagonya'nın Üst Triyasından bir heterodontosaurid ornithischian dinozor". Ameghiniana. 38 (3): 271–279.
- ^ Randall B. Irmis; William G. Parker; Sterling J. Nesbitt; Haziran Liu (2007). "Erken ornitorik dinozorlar: Triyas rekoru". Tarihsel Biyoloji. 19 (1): 3–22. CiteSeerX 10.1.1.539.8311. doi:10.1080/08912960600719988. S2CID 11006994.
- ^ a b Sereno, Paul C. (2012). "Heterodontosaurid dinozorların taksonomisi, morfolojisi, çiğneme işlevi ve filogenisi". ZooKeys (226): 1–225. doi:10.3897 / zookeys.226.2840. PMC 3491919. PMID 23166462.
- ^ Paul E. Olsen; Dennis V. Kent; Jessica H. Whiteside (2010). "Newark Supergroup tabanlı astrokronoloji ve jeomanyetik polarite zaman ölçeğinin (Newark-APTS) Dinosauria'nın erken çeşitliliğinin temposu ve modu için etkileri". Edinburgh Kraliyet Topluluğu'nun Dünya ve Çevre Bilimleri İşlemleri. 101 (3–4): 201–229. doi:10.1017 / S1755691011020032.
- ^ Sues, Hans-Dieter; Clark, James M .; Jenkins, Farish A. (1994). "Güneybatı Amerika’daki Glen Kanyon Grubu’ndan Erken Jurassic dörtayaklıların bir incelemesi". Fraser, Nicholas C .; Sues, Hans-Dieter (editörler). Dinozorların Gölgesinde: Erken Mezozoik Tetrapodlar. Cambridge: Cambridge University Press. s. 285–294. ISBN 978-0-521-45899-3.
- ^ Clark, James; Montellano, Marisol; Hopson, James A .; Hernandez, Rene; Fastovsky, David A. (1994). "Meksika'nın kuzeydoğusundaki La Boca Formasyonundan bir Erken veya Orta Jura tetrapod topluluğu". Fraser, N.C .; Sues, H.-D. (eds.). Dinozorların Gölgesinde: Erken Mezozoik Tetrapodlar. Cambridge: Cambridge University Press. s. 295–302. ISBN 978-0-521-45899-3.
- ^ Sánchez-Hernández, Barbara; Benton, Michael J.; Naish Darren (2007). "Kuzeydoğu İspanya Galve bölgesinin Geç Jura ve Erken Kretase dönemine ait dinozorlar ve diğer omurgalı fosilleri". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 249 (1–2): 180–215. doi:10.1016 / j.palaeo.2007.01.009.
- ^ Hopson, James A. (1980). "Erken Mezozoik ornitorik dinozorlarda diş fonksiyonu ve yerine koyma: aestivasyon için çıkarımlar". Lethaia. 13 (1): 93–105. doi:10.1111 / j.1502-3931.1980.tb01035.x.
- ^ a b c Butler, Richard J .; Porro, Laura B .; Norman, David B. (2008). "Ornitorik dinozorun ilkel kafatası Heterodontosaurus tucki Güney Afrika'daki "Stormberg" den. Omurgalı Paleontoloji Dergisi. 28 (3): 702–711. doi:10.1671 / 0272-4634 (2008) 28 [702: AJSOTP] 2.0.CO; 2. ISSN 0272-4634.
- ^ "SA'da bulunan dinozor fosili nihayet sırlarından vazgeçiyor". 2016-07-27. Arşivlendi 2016-07-28 tarihinde orjinalinden. Alındı 2016-07-27.
| ||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||
