Pachycephalosauria - Pachycephalosauria
| Pachycephalosaurs | |
|---|---|
 | |
| Oyuncu kadrosu Pachycephalosaurus wyomingensis kafatası, Oxford Üniversitesi Doğa Tarihi Müzesi | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Sipariş: | †Ornithischia |
| Clade: | †Cerapoda |
| Clade: | †Marginocephalia |
| Alttakım: | †Pachycephalosauria Maryańska & Osmólska, 1974 |
| Aile: | †Pachycephalosauridae Sternberg, 1945 (korunan isim ) |
| Türler | |
| †Pachycephalosaurus wyomingensis Gilmore, 1931 (korunan ad) | |
| Genera | |
| |
Pachycephalosauria (/ˌpækɪsɛfələˈsɔːrbenə,-ˌkɛf-/;[1] itibaren Yunan παχυκεφαλόσαυρος 'kalın başlı kertenkeleler' anlamına gelir) bir clade nın-nin ornitorik dinozorlar. İle birlikte Ceratopsia, sınıfı oluşturur Marginocephalia. Genera içerir Pachycephalosaurus, Stegoceras, ve Prenocephale. İki tür haricinde çoğu pachycephalosaur, Geç Kretase Yaklaşık 85,8 ila 65,5 milyon yıl öncesine dayanan dönem.[2] Onlar için özeldir Kuzey yarımküre hepsi burada bulunuyor Kuzey Amerika ve Asya. Hepsi iki ayaklı, otçul /her yerde yaşayan kalın kafatasları olan hayvanlar. Kafatasları türlere bağlı olarak kubbeli, düz veya kama şeklinde olabilir ve hepsi ağır bir şekilde kemiklenmiştir. Kubbeler genellikle düğümler ve / veya sivri uçlarla çevriliydi. Birkaç pachycephalosaur türüne ait kısmi iskeletler bulundu, ancak bugüne kadar tam bir iskelet keşfedilmedi. Genellikle izole edilmiş kafatası parçaları, bulunan tek kemiktir.[3]
Bilinen en eski pachycephalosaur için adaylar arasında Ferganocephale adenticulatum itibaren Orta Jura Periyot tabakaları Kırgızistan ve Stenopelix valdensis itibaren Erken Kretase tabakaları Almanya R.M. Sullivan, bu türlerden herhangi birinin pachycephalosaur olduğundan şüphe etti.[2] 2017'de Han ve meslektaşları tarafından yürütülen bir filogenetik analiz, Stenopelix üyesi olarak Ceratopsia.[4]
Paleobiyoloji
Anatomi
Pachycephalosaurs iki ayaklı ornithischians kalınlaşmış kafatasları ile karakterizedir. Geniş bir bağırsak boşluğu ve geniş kalçaları, kısa ön ayakları, uzun bacakları, kısa, kalın bir boynu ve ağır bir kuyruğu olan hacimli bir gövdeleri vardı. Büyük yörüngeler ve büyük optik sinir iyi görüşe sahip ve alışılmadık derecede büyük pachycephalosaurlara işaret edin koku alma lobları diğer dinozorlara göre iyi bir koku alma duyusuna sahip olduklarını gösterir.[3] Oldukça küçük dinozorlardı, çoğu 2–3 metre (6,6-9,8 fit) uzunluğunda ve en büyüğüdür. Pachycephalosaurus wyomingensis, 4,5 metre (14,8 fit) uzunluğunda ve 450 kilogram (990 pound) ağırlığında olduğu tahmin edilmektedir.[2][5] Pachycephalosaurların karakteristik kafatası, füzyon ve kalınlaşmanın bir sonucudur. cephe ve parietaller kapanış eşliğinde supratemporal fenestra. Bazı türlerde bu, yükseltilmiş bir kubbe şeklini alır; diğerlerinde kafatası düz veya kama şeklindedir. Düz başlı pachycephalosaurlar geleneksel olarak ayrı türler ve hatta aileler olarak kabul edilirken, kubbe başlı yetişkinlerin yavrularını temsil edebilirler.[2][3] Tüm ekranlar oldukça süslü testiler, squamosals, ve postorbitaller künt boynuzlar ve düğümler şeklinde. Pek çok tür sadece kafatası parçalarından biliniyor ve tam bir pachycephalosaur iskeleti henüz bulunmadı.[3]
Kafa atma davranışı

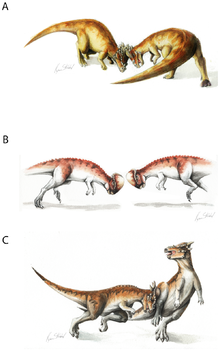
Kafatası kubbesinin uyarlanabilir önemi yoğun bir şekilde tartışıldı. Halk arasında, kafatasının bir tür dinosauri koçu olarak kafa atmada kullanıldığına dair popüler hipotezi ilk olarak Colbert (1955), s.[sayfa gerekli ]. Bu görüş, 1956 bilim kurgu öyküsünde popüler hale geldi "Dinozor Silahı " tarafından L. Sprague de Camp. O zamandan beri birçok paleontolog, kafa atma hipotezi için tartıştılar. Galton (1970) ve Sues (1978). Bu hipotezde, pachycephalosaurlar, günümüzde olduğu gibi birbirleriyle kafa kafaya çarpıştı. dağ keçileri ve misk öküz.
Savaşçı davranış için anatomik kanıtlar arasında omurga sertliği sağlayan vertebral eklemler ve güçlü boyun kas sistemini gösteren sırtın şekli yer alır.[6] Bir pachycephalosaur'un çarpma sırasında stresi iletmek için başını, boynunu ve vücudunu yatay olarak düzleştirebileceği öne sürüldü. Ancak bilinen hiçbir dinozorda baş, boyun ve vücut böyle bir konuma yönlendirilemezdi. Bunun yerine, pachycephalosaurların servikal ve ön dorsal omurları, boynun "S" veya "U" şeklinde bir eğri içinde taşındığını gösterir.[7]
Ayrıca kafatasının yuvarlak şekli, kafa atma sırasında temas eden yüzey alanını azaltarak göz atma darbelerine neden olur. Diğer olasılıklar arasında, yan tarafa vurma, avcılara karşı savunma veya her ikisi bulunur. Göreceli olarak geniş pachycephalosaurs yapıları (bu, hayati iç organları, kanat dikme sırasında zarar görmekten koruyacaktı) ve yassı boynuzları Stygimoloch (kanat dikme sırasında büyük bir etki için kullanılmış olan) kanat dikme hipotezine güven katıyor.
Tarafından yürütülen histolojik bir çalışma Goodwin ve Horner (2004) çarpma ram hipotezine karşı çıktı. Kubbenin "geçici bir ontogenetik aşama" olduğunu, süngerimsi kemik yapısının savaş darbelerini kaldıramadığını ve radyal modelin basitçe hızlı büyümenin bir etkisi olduğunu savundular.[8] Daha sonra biyomekanik analizler Snively ve Cox (2008) ve Snively ve Theodor (2011) Ancak, kubbelerin savaş streslerine dayanabileceği sonucuna vardı.[6]Lehman (2010) Goodwin ve Horner tarafından tartışılan büyüme modellerinin kafa atma davranışıyla tutarsız olmadığını savundu.[9]
Goodwin ve Horner (2004) bunun yerine kubbenin türlerin tanınması için işlev gördüğünü savundu. Kubbenin bir tür dış kaplamaya sahip olduğuna dair kanıtlar vardır ve kubbenin parlak bir şekilde kaplanmış olabileceğini veya mevsimsel olarak renginin değişebileceğini düşünmek mantıklıdır.[8] Ancak fosil kayıtlarının doğası gereği kubbe işlevinde rengin rol oynayıp oynamadığı gözlemlenememektedir.
Longrich, Sankey ve Tanke (2010) türlerin tanınmasının kubbe için olası bir evrim nedeni olduğunu savundu, çünkü kubbe formları türler arasında belirgin bir şekilde farklı değildir. Bu genel benzerlik nedeniyle, bazı Pachycephalosauridae cinsleri bazen yanlış bir şekilde bir araya toplanmıştır. Bu, ceratopsia'lılardaki durumdan farklıdır ve hadrosauridler çok daha belirgin kafatası süslemesine sahip olan. Longrich ve diğerleri Bunun yerine kubbenin, kaynak yatırımını haklı çıkaracak kadar önemli olan, savaş gibi mekanik bir işlevi olduğunu savundu.[10]
Paleopatoloji ile kafa tutma desteği

Peterson, Dischler ve Longrich (2013) Pachycephalosauridae arasında kraniyal patolojileri inceledi ve incelenen tüm kubbelerin% 22'sinin, osteomiyelit Penetran travmadan kaynaklanan kemik enfeksiyonu veya kafatasını örten dokuda kemik dokusunda enfeksiyona yol açan travma. Bu yüksek patoloji oranı, pachycephalosaurid kubbelerin intra-spesifik savaşta kullanıldığı hipotezine daha fazla destek vermektedir.[11] Travma sıklığı, bu konuda farklı cinsler arasında karşılaştırılabilirdi. aile bu cinslerin kubbelerinin boyutuna ve mimarisine göre farklılık göstermelerine ve değişen jeolojik dönemlerde var oldukları gerçeğine rağmen.[11] Bu bulgular, patolojinin olmadığı nispeten düz başlı pachycephalosauridlerin analizinden elde edilen sonuçlarla tam bir tezat içindeydi. Bu, bu bireylerin kadınları veya gençleri temsil ettiği hipotezini destekleyecektir.[10] belirli bir savaş davranışının beklenmediği yerlerde.
Histolojik inceleme, pachycephalosaurid kubbelerin benzersiz bir fibrolamellar kemik formundan oluştuğunu ortaya koymaktadır.[12] içeren fibroblastlar yara iyileşmesinde kritik bir rol oynayan ve yeniden modelleme sırasında kemiği hızla biriktirebilen.[13] Peterson ve diğerleri (2013), birlikte ele alındığında, lezyon dağılımının sıklığı ve frontoparietal kubbelerin kemik yapısının, pachycephalosauridlerin benzersiz kraniyal yapılarını bunun için kullandıkları hipotezine güçlü bir destek sağladığı sonucuna varmıştır. agonistik davranış.[11]
Diyet
Çoğu pachycephalosaur türünün küçük boyutu ve iskelet uyumunun olmaması, onların dağcı olmadıklarını ve öncelikle yere yakın yiyecek yediklerini gösterir. Mallon vd. (2013), ada kıtasında otçulların bir arada varlığını inceledi. Laramidia Geç Kretase döneminde ve pachycephalosauridlerin genellikle 1 metre yükseklikte veya daha düşük bitki örtüsüyle beslenmekle sınırlı olduğu sonucuna varmıştır.[14] Sergiliyorlar heterodonluk arasında farklı diş morfolojisine sahip premaksiller dişler ve maksiller diş. Ön dişler küçüktür ve yumurtalık bir kesite sahiptir ve büyük olasılıkla yiyecek kapmak için kullanılmıştır. Bazı türlerde son premaksiller diş büyümüş ve köpeklere benzerdi. Arka dişler küçük ve üçgendir. dişler ağzı işleme için kullanılan tepenin önünde ve arkasında. Olduğu türlerde diş hekimi bulunmuştur, alt çene dişleri boyut ve şekil olarak üst çenedeki dişlere benzerdir. Dişlerdeki yıpranma kalıpları türe göre değişir, bu da tohumlar, gövdeler, yapraklar, meyveler ve muhtemelen böcekleri içerebilecek bir dizi gıda tercihini gösterir. Kuyruğun tabanına kadar uzanan çok geniş bir göğüs kafesi ve geniş bağırsak boşluğu, yiyecekleri sindirmek için fermantasyonun kullanılmasını önerir.[3]
Biyocoğrafya
Dağıtım
Pachycephalosaurs yalnızca Laurasia batıda bulunan Kuzey Amerika ve merkezi Asya. Pachycephalosaurlar Asya'da ortaya çıktı ve iki ana dağılma olayına sahipti, bu da Asya'da gözlemlenen iki ayrı pachycephalosaur evrim dalgasına neden oldu. Birincisi, geç saatlerden önce meydana geliyor Santoniyen veya erken Kampaniyen, Asya'dan Kuzey Amerika'ya, büyük olasılıkla Bering Kara Köprüsü. Bu göç, ortak bir atadan Stygimoloch, Stegoceras, Tylocephale, Prenocephale, ve Pachycephalosaurus. İkinci olay ortadan önce meydana geldi Kampaniyen ve ortak bir atadan Kuzey Amerika'dan Asya'ya göçü içeriyordu. Prenocephale ve Tylocephale. Başlangıçta bu aralığın dışında keşfedilen pachycephalosaurlar olduğu bildirilen iki tür, Yaverlandia bitholus nın-nin İngiltere ve Majungatholus atopus nın-nin Madagaskar, son zamanlarda aslında theropodlar.[3][15]
Çevre
Asya ve Kuzey Amerika pachycephalosaur türleri oldukça farklı ortamlarda yaşadılar. Asya örnekleri normalde daha sağlamdır, bu da fosilleşmeden önce ölüm yerlerinden çok uzağa taşınmadıklarını gösterir. Büyük bir olasılıkla Orta Asya'da sıcak ve soğuk bir çöl bölgesinde yaşıyorlardı. kurak iklim. Kuzey Amerika örnekleri tipik olarak, denizden gelen erozyonla oluşan kayalarda bulunur. kayalık Dağlar. Örnekler çok daha az sağlamdır; genellikle sadece kafatası başlıkları çıkarılır ve düzenli olarak bulunanlar yüzeyde pul pul dökülme ve fosilleşmeden önce su ile uzun mesafeler taşındığına dair diğer işaretler sergiler. Dağlarda bir dağda yaşadıkları varsayılmaktadır. ılıman iklim ve ölümden sonra erozyonla son istirahat yerlerine taşındı.[16]
Sınıflandırma

Çoğu pachycephalosaurid kalıntısı tam değildir, genellikle belirgin kubbeyi oluşturan frontoparietal kemiğin bölümlerinden oluşur. Bu yapabilir taksonomik içindeki cins ve türlerin sınıflandırılması olarak zor bir görevi tanımlama Pachycephalosauria neredeyse tamamen kafatası özelliklerine dayanır. Sonuç olarak, uygun olmayan türler tarihsel olarak küme atandı. Örneğin, Majungatholus, bir zamanlar bir pachycephalosaur olduğu düşünülen, şimdi bir örnek olarak kabul edilmektedir. Abelisaurid Theropod Majungasaurus, ve Yaverlandia, başlangıçta bir pachycephalosaurid olarak tanımlanan başka bir dinozor da yakın zamanda bir Coelurosaur (Naish in Sullivan (2006) ). Daha karmaşık konular, ontogenetik ve cinsel özellikler pachycephalosaurlarda.
2009 tarihli bir makale, Dracorex ve Stygimoloch sadece erken büyüme aşamalarıydı Pachycephalosaurus, farklı cinsler yerine.[13]
Bir 2020 yeniden çalışması Cerapoda Dieudonné tarafından et al. geleneksel olarak kabul edilen hayvanları kurtardı 'heterodontosauridler 'Pachycephalosauria içinde bir bazal grup olarak, geleneksel, kubbe başlı pachycephalosaurlara göre parafiletik.[17]
Taksonomi


Pachycephalosauria ilk olarak bir alttakım düzenin Ornithischia tarafından Maryańska ve Osmólska (1974). İçine sadece bir tane dahil ettiler aile Pachycephalosauridae.[18] Michael Benton gibi daha sonraki araştırmacılar, bunu bir infraorder bir alt siparişin Cerapoda birleştiren Ceratopsia'cılar ve ornitopodlar.[19] 2006'da Robert Sullivan, pachycephalosaur taksonomisinin yeniden değerlendirmesini yayınladı. Sullivan, Maryańska ve Osmólska'nın Pachycephalosauria tanımını, Pachycephalosauridae ile gereksiz olan tanımını kısıtlama girişimlerini değerlendirdi, çünkü bunlar aynı anatomik karakterlerle teşhis edildi. Sullivan tarafından yapılan girişimleri de reddetti Sereno (1986) onun içinde filogenetik çalışmalar,[20] Pachycephalosauridae'yi yalnızca "kubbe kafataslı" türleri içerecek şekilde yeniden tanımlamak ( Stegoceras ve Pachycephalosaurus), Pachycephalosauria'da bu ailenin dışında daha fazla "bazal" tür bırakırken. Bu nedenle Sullivan'ın Pachycephalosauridae'yi kullanımı, Sereno ve Benton'ın Pachycephalosauria kullanımıyla eşdeğerdir.
Sullivan, Pachycephalosauridae'yi sadece kafatasının karakterlerine dayanarak teşhis etti, tanımlayıcı karakter kubbe şeklinde bir frontoparietal kafatası kemiğiydi. Sullivan'a göre, ilkel olduğu varsayılan bazı türlerde bu özelliğin yokluğu, kubbeli ve kubbeli olmayan pachycephalosaurlar arasında sınıflandırmanın ayrılmasına yol açtı; ancak, düz kafatasları olan daha gelişmiş ve muhtemelen daha genç pachycephalosaurların keşfi (örneğin Dracorex hogwartsia) bu ayrımın yanlış olduğunu gösterir. Sullivan ayrıca, Pachycephalosauridae'nin orijinal teşhisinin "düz ila kubbe benzeri" kafatasları etrafında merkezlendiğini, bu nedenle düz başlı formların aileye dahil edilmesi gerektiğini belirtti.[21]

Filogeni
kladogram burada sunulan bir analizi takip eder Williamson ve Carr (2002).[22]
| Pachycephalosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cladogram sonra Longrich, Sankey & Tanke (2010).[10]
| Pachycephalosauridae | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Aşağıda bir kladogram -den değiştirildi Evans vd. (2013).[23]
| Pachycephalosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ayrıca bakınız
Notlar
- ^ "Pachycephalosauria". Oxford Sözlükleri İngiltere Sözlüğü. Oxford University Press. Alındı 2016-01-21.
- ^ a b Holtz, Thomas R. Jr. (2007) [28 Ocak 2011'de güncellenmiştir]. "Holtz (2007) Dinozorları İçin Cins Listesi" (PDF).
- ^ a b c d e Maryanska, T .; Chapman, R.E .; Weishampel, D.B. (2004). "Pachycephalosauria". Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. sayfa 464–477. ISBN 978-0520242098. OCLC 493366196.
- ^ Han, F.-L .; Forster, C.A .; Clark, J.M .; Xu, X. (2016). "Kafatası anatomisi Yinlong downsi (Ornithischia: Ceratopsia) Sincan, Çin'in Üst Jura Shishugou Formasyonundan ". Omurgalı Paleontoloji Dergisi. 36 (1): e1029579. doi:10.1080/02724634.2015.1029579.
- ^ Paul Gregory S. (2010). Princeton dinozorlar için saha rehberi. Princeton University Press. ISBN 9780691137209. OCLC 930852339.[sayfa gerekli ]
- ^ a b Snively E, Cox A (Mart 2008). "Pachycephalosaur kafatasının yapısal mekaniği kafa atma davranışına izin verdi". Paleontoloji Electronica. 11 (1). ISSN 1094-8074.
- ^ Marangoz K (1997). "Pachycephalosaur'larda (Ornithischia: Dinosauria) agonistik davranış: kafa atma davranışına yeni bir bakış" (PDF). Jeolojiye Katkılar. 32 (1): 19–25. Arşivlenen orijinal (PDF) 2016-03-03 tarihinde. Alındı 2011-07-16.
- ^ a b Goodwin MB, Horner JR (Haziran 2004). "Pachycephalosaurların (Ornithischia: Marginocephalia) kraniyal histolojisi, kafa atma davranışıyla tutarsız geçici yapıları ortaya çıkarır". Paleobiyoloji. 30 (2): 253–267. doi:10.1666 / 0094-8373 (2004) 030 <0253: CHOPOM> 2.0.CO; 2.
- ^ Lehman TM (2010). "Batı Teksas'ın San Carlos ve Aguja Formasyonlarından (Üst Kretase) Pachycephalosauridae ve frontoparietal kubbenin gözlemleri". Omurgalı Paleontoloji Dergisi. 30 (3): 786–798. doi:10.1080/02724631003763532. ISSN 0272-4634.
- ^ a b c Longrich NR, Sankey J, Tanke D (2010). "Texacephale langstoni, güney Teksas, ABD'deki üst Campanian Aguja Formasyonundan yeni bir pachycephalosaurid (Dinosauria: Ornithischia) cinsi". Kretase Araştırmaları. 31 (2): 274–284. doi:10.1016 / j.cretres.2009.12.002.
- ^ a b c Peterson JE, Dischler C, Longrich NR (2013). "Kraniyal Patolojilerin Dağılımları Kubbe Başlı Dinozorlarda (Pachycephalosauridae) Kafa Atma Durumuna Dair Kanıt Sağlıyor". PLoS ONE. 8 (7): e68620. Bibcode:2013PLoSO ... 868620P. doi:10.1371 / journal.pone.0068620. PMC 3712952. PMID 23874691.
- ^ Reid, R.E.H. (1997). "Kemiklerin ve dişlerin histolojisi". Currie, P.J .; Padian, K. (editörler). Dinozorlar Ansiklopedisi. San Diego, CA: Academic Press. s. 329–339.
- ^ a b Horner JR, Goodwin MB (2009). "Üst Kretase Dinozorunda Aşırı Kranial Ontogeny Pachycephalosaurus". PLoS ONE. 4 (10): e7626. Bibcode:2009PLoSO ... 4.7626H. doi:10.1371 / journal.pone.0007626. PMC 2762616. PMID 19859556.
- ^ Mallon, Ürdün C; David C Evans; Michael J Ryan; Jason S Anderson (2013). "Kanada, Alberta'daki Dinozor Parkı Formasyonundan (Üst Kampaniyen) otçul dinozorlar arasında beslenme yüksekliği katmanlaşması". BMC Ekolojisi. 13: 14. doi:10.1186/1472-6785-13-14. PMC 3637170. PMID 23557203.
- ^ Naish D, Martill DM (2008). "Büyük Britanya'nın Dinozorları ve Londra Jeoloji Derneği'nin keşiflerindeki rolü: Ornithischia". Jeoloji Topluluğu Dergisi. 165 (3): 613–623. Bibcode:2008JGSoc.165..613N. doi:10.1144/0016-76492007-154.
- ^ Weishampel, David B. (2009). Dinozorlar: kısa bir doğa tarihi. Cambridge University Press. ISBN 9780511479410. OCLC 476234422.[sayfa gerekli ]
- ^ Dieudonné PE, Cruzado-Caballero P, Godefroit P, Tortosa T (2020). "Cerapodan dinozorlarının yeni bir filogeni". Tarihsel Biyoloji: 1–21. doi:10.1080/08912963.2020.1793979.CS1 Maint: ekstra noktalama (bağlantı)
- ^ Maryańska T, Osmólska H (1974). "Pachycephalosauria, ornithischian dinozorlarının yeni bir alt grubu". Palaeontologica Polonica. 30: 45–102.
- ^ Benton, Michael J. (2004). Omurgalı Paleontoloji (3. baskı). Blackwell Publishing. s. 472. ISBN 978-0-632-05637-8.
- ^ Sereno PC (1986). "Kuş kalçalı dinozorların filogenisi (Ornithischia Siparişi)". National Geographic Araştırma. 2: 234–256.
- ^ Sullivan RM (2006). "Pachycephalosauridae'nin (Dinosauria: Ornithischia) taksonomik bir incelemesi" (PDF). New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 35: 347–365. Arşivlenen orijinal (PDF) 2007-09-27 tarihinde.
- ^ Williamson TE, Carr TD (2002). "Batı Kuzey Amerika'dan türetilmiş yeni bir pachycephalosaurian cinsi". Omurgalı Paleontoloji Dergisi. 22 (4): 779–801. doi:10.1671 / 0272-4634 (2002) 022 [0779: ANGODP] 2.0.CO; 2. ISSN 0272-4634.
- ^ Evans DC, Schott RK, Larson DW, Brown CM, Ryan MJ (2013). "En eski Kuzey Amerika pachycephalosaurid ve küçük gövdeli ornitosiyan dinozorların gizli çeşitliliği". Doğa İletişimi. 4. 1828. Bibcode:2013NatCo ... 4.1828E. doi:10.1038 / ncomms2749. PMID 23652016.
Referanslar
- Benton, Michael J (2004). Omurgalı Paleontoloji (3. baskı). Blackwell Publishing. ISBN 978-0-632-05637-8.
- Carpenter, Kenneth (1997). "Pachycephalosaur'larda (Ornithischia: Dinosauria) agonistik davranış: kafa atma davranışına yeni bir bakış" (PDF). Jeolojiye Katkılar. 32 (1): 19–25. Arşivlenen orijinal (PDF) 2016-03-03 tarihinde. Alındı 2011-07-16.
- Colbert, Edwin (1955). Omurgalıların Evrimi. New York: John Wiley.
- Galton, P (1970). "Pachycephalosaurids - dinosaurian koçları". Keşif. 6: 23–32.
- Goodwin, MB; Horner, JR (Haziran 2004). "Pachycephalosaurların (Ornithischia: Marginocephalia) kraniyal histolojisi, kafa atma davranışıyla tutarsız geçici yapıları ortaya çıkarır". Paleobiyoloji. 30 (2): 253–267. doi:10.1666 / 0094-8373 (2004) 030 <0253: CHOPOM> 2.0.CO; 2.
- Horner, JR; Goodwin, MB (2009). Sereno, Paul (ed.). "Üst Kretase Dinozorunda Aşırı Kraniyal Ontogeny Pachycephalosaurus". PLoS ONE. 4 (10): e7626. Bibcode:2009PLoSO ... 4.7626H. doi:10.1371 / journal.pone.0007626. PMC 2762616. PMID 19859556.
- Lehman, TM (2010). "Batı Teksas'ın San Carlos ve Aguja Formasyonlarından (Üst Kretase) Pachycephalosauridae ve frontoparietal kubbenin gözlemleri". Omurgalı Paleontoloji Dergisi. 30 (3): 786–798. doi:10.1080/02724631003763532. ISSN 0272-4634.
- Longrich, NR; Sankey, J; Tanke, D (2010). "Texacephale langstoni, üst Campanian Aguja Formasyonundan yeni bir pachycephalosaurid (Dinosauria: Ornithischia) cinsi, güney Teksas, ABD ". Kretase Araştırmaları. 31 (2): 274–284. doi:10.1016 / j.cretres.2009.12.002.
- Maryańska, T; Osmólska, H (1974). "Pachycephalosauria, ornithischian dinozorlarının yeni bir alt grubu". Palaeontologica Polonica (30): 45–102.
- Sereno, PC (1986). "Kuş kalçalı dinozorların filogenisi (Ornithischia Siparişi)". National Geographic Araştırma. 2: 234–256.
- Snively, E; Cox, A (Mart 2008). "Pachycephalosaur kafatasının yapısal mekaniği kafa atma davranışına izin verdi". Paleontoloji Electronica. 11 (1). ISSN 1094-8074.
- Snively, E; Theodor, JM (2011). Carpenter, Kenneth (ed.). "Pachycephalosaur Stegoceras validum (Ornithischia, Dinosauria) ve Combative Artiodactyls'de Baş Vuruşu Davranışının Ortak İşlevsel İlişkileri". PLoS ONE. 6 (6): e21422. Bibcode:2011PLoSO ... 621422S. doi:10.1371 / journal.pone.0021422. PMC 3125168. PMID 21738658.
- Stokstad, Erik (Kasım 2007). "Omurgalı Paleontoloji Derneği Toplantısı: Azgın Genç Dinozorlar Ayrı Tür Yanılsamasına Neden Oldu mu?". Bilim. 318 (5854): 1236. doi:10.1126 / science.318.5854.1236. PMID 18033861.
- Sues, H-D (1978). "Pakycephalosaurid dinozorlardaki kubbenin işlevsel morfolojisi". Neues Jahrbuch für Geologie und Paläontologie: 459–472.
- Sullivan, RM (2006). "Pachycephalosauridae'nin (Dinosauria: Ornithischia) taksonomik bir incelemesi" (PDF). New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni (35): 347–365. Arşivlenen orijinal (PDF) 2007-09-27 tarihinde.
- Watabe, Mahito; Tsogtbaatar, Khishigjaw; Sullivan, Robert M. (2011). "Baynshire Formasyonundan yeni bir pachycephalosaurid (Senomaniyen-geç Santoniyen), Gobi Çölü, Moğolistan" (PDF). Fosil Kaydı 3. New Mexico Doğa Tarihi ve Bilim Müzesi, Bülten. 53: 489–497. Arşivlenen orijinal (PDF) 2011-09-29 tarihinde.
- Williamson, TE; Carr, TD (2002). "Batı Kuzey Amerika'dan türetilmiş yeni bir pachycephalosaurian cinsi". Omurgalı Paleontoloji Dergisi. 22 (4): 779–801. doi:10.1671 / 0272-4634 (2002) 022 [0779: ANGODP] 2.0.CO; 2. ISSN 0272-4634.
Dış bağlantılar
- TEDx konuşma açık Youtube tarafından Jack Horner şekil değiştiren dinozor kafatasları ve yanlış dinozor sınıflandırması üzerine
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||





