Dihidrofolat redüktaz - Dihydrofolate reductase
| Dihidrofolat redüktaz | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Tavuk karaciğeri dihidrofolat redüktazın kristal yapısı. PDB girişi 8dfr | |||||||||
| Tanımlayıcılar | |||||||||
| EC numarası | 1.5.1.3 | ||||||||
| CAS numarası | 9002-03-3 | ||||||||
| Veritabanları | |||||||||
| IntEnz | IntEnz görünümü | ||||||||
| BRENDA | BRENDA girişi | ||||||||
| ExPASy | NiceZyme görünümü | ||||||||
| KEGG | KEGG girişi | ||||||||
| MetaCyc | metabolik yol | ||||||||
| PRIAM | profil | ||||||||
| PDB yapılar | RCSB PDB PDBe PDBsum | ||||||||
| Gen ontolojisi | AmiGO / QuickGO | ||||||||
| |||||||||
| Dihidrofolat redüktaz | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Tanımlayıcılar | |||||||||
| Sembol | DHFR_1 | ||||||||
| Pfam | PF00186 | ||||||||
| Pfam klan | CL0387 | ||||||||
| InterPro | IPR001796 | ||||||||
| PROSITE | PDOC00072 | ||||||||
| SCOP2 | 1dhi / Dürbün / SUPFAM | ||||||||
| |||||||||
| R67 dihidrofolat redüktaz | |||||||||
|---|---|---|---|---|---|---|---|---|---|
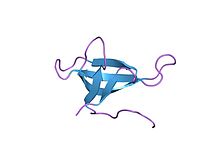 Plazmidle kodlanmış bir dihidrofolat redüktazın yüksek çözünürlüklü yapısı E. coli. PDB girişi 2gqv | |||||||||
| Tanımlayıcılar | |||||||||
| Sembol | DHFR_2 | ||||||||
| Pfam | PF06442 | ||||||||
| InterPro | IPR009159 | ||||||||
| SCOP2 | 1vif / Dürbün / SUPFAM | ||||||||
| |||||||||





Dihidrofolat redüktazveya DHFR, bir enzim azalır dihidrofolik asit -e tetrahidrofolik asit, kullanma NADPH gibi elektron vericisi tetrahidrofolat türlerine dönüştürülebilen kofaktörler 1-karbon transfer kimyasında kullanılır. İnsanlarda DHFR enzimi şu şekilde kodlanır: DHFR gen.[5][6]Kromozom 5'in q11 → q22 bölgesinde bulunur.[7] Bakteriyel Türler farklı DHFR'ye sahip enzimler (diaminoheterosiklik molekülleri bağlama modellerine göre), ancak memeli DHFR'ler oldukça benzerdir.[8]
Yapısı
Merkezi sekiz telli beta kıvrımlı levha ana özelliğini oluşturur polipeptid DHFR'nin omurga katlanması.[9] Bu ipliklerin yedisi paraleldir ve sekizinci paralel olarak uzanır. Dört alfa sarmalları ardışık beta dizilerini bağlayın.[10] Kalıntılar 9 - 24, "Met20" veya "döngü 1" olarak adlandırılır ve diğer döngülerle birlikte, çevreleyen ana alt alanın parçasıdır. aktif site.[11] aktif site yer almaktadır N terminali bir içeren dizinin yarısı korunmuş Pro -Trp dipeptid; triptofan bağlayıcı olduğu gösterilmiştir substrat enzim tarafından.[12]

Bağlı dihidrofolat ve NADPH ile insan DHFR'si

Fonksiyon
Dihidrofolat redüktaz dönüştürür dihidrofolat içine tetrahidrofolat de novo sentezi için gerekli bir proton mekiği pürinler, timidilik asit ve kesin amino asitler. Fonksiyonel dihidrofolat redüktaz geni, kromozom 5'e eşlenirken, birden fazla intronsuz işlenmiş psödojen veya dihidrofolat redüktaz benzeri gen, ayrı kromozomlarda tanımlanmıştır.[13]

DHFR tarafından katalize edilen reaksiyon.

Tetrahidrofolat sentez yolu.


Tüm organizmalarda bulunan DHFR, hücredeki tetrahidrofolat miktarını düzenlemede kritik bir role sahiptir. Tetrahidrofolat ve türevleri aşağıdakiler için gereklidir: pürin ve timidilat hücre proliferasyonu ve hücre büyümesi için önemli olan sentez.[14] DHFR, sentezinde merkezi bir rol oynar. nükleik asit öncülerdir ve DHFR'den tamamen yoksun olan mutant hücrelerin büyümesi için glisin, bir amino asit ve timidine ihtiyaç duyduğu gösterilmiştir.[15] DHFR, tetrahidrobiopterinin dihidrobiopterinden kurtarılmasında rol oynayan bir enzim olarak da gösterilmiştir.[16]
Mekanizma

Genel mekanizma
DHFR, bir hidritin transferini katalize eder. NADPH -e dihidrofolat eşlik eden bir protonasyon ile birlikte tetrahidrofolat.[14] Sonunda, dihidrofolat tetrahidrofolata indirgenir ve NADPH oksitlenerek NADP +. Met20'nin ve aktif bölgenin yakınındaki diğer halkaların yüksek esnekliği, ürünün, tetrahidrofolatın salınmasını teşvik etmede rol oynar. Özellikle Met20 halkası, hidritin NADPH'den dihidrofolata transferini teşvik etmek için NADPH'nin nikotinamid halkasının stabilize edilmesine yardımcı olur.[11]

Bu enzimin mekanizması aşamalı ve kararlı durum tesadüfidir. Spesifik olarak, katalitik reaksiyon, NADPH ve substratın enzimin bağlanma bölgesine bağlanmasıyla başlar, ardından protonasyon ve kofaktör NADPH'den substrata hidrit aktarımı izler. Bununla birlikte, son iki adım aynı geçiş durumunda aynı anda gerçekleşmez.[17][18] Hesaplamalı ve deneysel yaklaşımların kullanıldığı bir çalışmada, Liu ve diğerleri protonasyon aşamasının hidrit transferinden önce geldiği sonucuna varılır.[19]

DHFR'nin enzimatik mekanizmasının, pH'a bağlı olduğu, özellikle hidrit transfer aşamasının, aktif bölgenin elektrostatiği ve kalıntılarının iyonlaşma durumu üzerinde önemli bir etkiye sahip olduğu gösterildiğinden gösterilmiştir.[19] Substrat üzerindeki hedeflenen nitrojenin asitliği, substratın suyla doğrudan temas etmesine rağmen hidrofobik olduğu kanıtlanan enzimin bağlanma bölgesine bağlanmasında önemlidir.[17][20] Asp27, bağlanma yerindeki tek yüklü hidrofilik kalıntıdır ve Asp27 üzerindeki yükün nötralizasyonu enzimin pKa'sını değiştirebilir. Asp27, substratın protonasyonuna yardımcı olarak ve substratı hidrit transferi için uygun konformasyonda kısıtlayarak katalitik mekanizmada kritik bir rol oynar.[21][17][20] Protonasyon adımının, enol tatomerizasyonu ile ilişkili olduğu gösterilmiştir, ancak bu dönüşüm, proton bağışı için uygun kabul edilmemektedir.[18] Protonasyon aşamasına bir su molekülünün dahil olduğu kanıtlanmıştır.[22][23][24] Su molekülünün enzimin aktif bölgesine girişi Met20 döngüsü ile kolaylaştırılır.[25]
DHFR'nin konformasyonel değişiklikleri
DHFR tarafından katalize edilen reaksiyonun katalitik döngüsü beş önemli ara maddeyi içerir: holoenzim (E: NADPH), Michaelis kompleksi (E: NADPH: DHF), üçlü ürün kompleksi (E: NADP+: THF), tetrahidrofolat ikili kompleks (E: THF) ve THF‚NADPH kompleksi (E: NADPH: THF). E: NADPH: THF'den E: NADPH'ye ürün (THF) ayrışma adımı, kararlı durum devri sırasında hız belirleme adımıdır.[21]
Konformasyonel değişiklikler, DHFR'nin katalitik mekanizmasında kritiktir.[26] DHFR'nin Met20 döngüsü, aktif bölgeyi açabilir, kapatabilir veya tıkayabilir.[23][17] Buna bağlı olarak, açık, kapalı ve tıkalı durumlar olarak sınıflandırılan üç farklı biçim Met20'ye atanır. Ek olarak, belirsiz karakterizasyon sonuçları nedeniyle Met20'nin ekstra bozuk konformasyonu tanımlandı.[23] Met20 halkası, nikotinamid halkasının aktif bölgeden kapatıldığı üç ürün bağlama ara ürününde tıkalı konformasyonunda gözlenir. Bu konformasyonel özellik, NADP'nin ikame edilmesinin+ NADPH tarafından ürün ayrıştırmadan önce. Bu nedenle, bir sonraki reaksiyon döngüsü, substratın bağlanması üzerine meydana gelebilir.[21]


R67 DHFR
Eşsiz yapısı ve katalitik özellikleri nedeniyle, R67 DHFR geniş çapta incelenmektedir. R67 DHFR, E. coli kromozomal DHFR ile genetik ve yapısal olarak ilişkisi olmayan bir tip II R-plazmid tarafından kodlanmış DHFR'dir. Çözücüye [boş] maruz kalan tek bir aktif bölge gözeneğine sahip 222 simetrisine sahip bir homotetramerdir.[27] Aktif bölgenin bu simetrisi, enzimin farklı bağlanma modu ile sonuçlanır: Pozitif işbirliğine sahip iki dihidrofolat (DHF) molekülü veya negatif işbirliğine sahip iki NADPH molekülü veya bir substrat artı bir, ancak yalnızca ikincisi katalitik aktivite.[28] E. coli kromozomal DHFR ile karşılaştırıldığında, daha yüksek K'ye sahiptirm dihidrofolat (DHF) ve NADPH'nin bağlanmasında. Çok daha düşük katalitik kinetikler, hidrit transferinin, ürün (THF) salımından çok hız belirleme adımı olduğunu gösterir.[29]
R67 DHFR yapısında, homotetramer bir aktif saha gözeneği oluşturur. Katalitik işlemde, DHF ve NADPH zıt pozisyondan gözeneğe girer. NADPH'nin nikotinamid halkası ile DHF'nin pteridin halkası arasındaki π-π istifleme etkileşimi, aktif bölgedeki iki reaktanı sıkıca bağlar. Bununla birlikte, DHF'nin p-aminobenzoglutamate kuyruğunun esnekliği, geçiş durumunun oluşumunu teşvik edebilen bağlanma üzerine gözlendi.[30]
Klinik önemi
Dihidrofolat redüktaz eksikliği ile bağlantılı megaloblastik anemi.[13] Tedavi ile indirgenmiş folik asit formları. Bu reaksiyonun ürünü olan tetrahidrofolat, insanlarda folatın aktif formu olduğundan, DHFR'nin inhibisyonu fonksiyonel folat eksikliği. DHFR, DNA prekürsör sentezindeki temel rolü nedeniyle inhibisyon için çekici bir farmasötik hedeftir. Trimetoprim, bir antibiyotik Bakteriyel DHFR'yi inhibe ederken metotreksat, bir kemoterapi ajan, memeli DHFR'sini inhibe eder. Ancak, direnç DHFR'nin kendisindeki mutasyonel değişikliklerin bir sonucu olarak bazı ilaçlara karşı gelişmiştir.[31]
DHFR mutasyonları, nadir görülen otozomal resesif doğuştan folat metabolizması hatasına neden olur. megaloblastik anemi, pansitopeni ve şiddetli serebral folat eksikliği ile düzeltilebilir folinik asit takviye.[32][33]
Terapötik uygulamalar
Yapmak için hücreleri hızla bölerek folat gerektiğinden timin bu etki, terapötik avantaj için kullanılabilir.
DHFR, kanser tedavisinde ve bakteriyel enfeksiyonlara karşı potansiyel bir hedef olarak hedeflenebilir. DHFR, bir hücredeki tetrahidrofolat seviyelerinden sorumludur ve DHFR'nin inhibisyonu, kanser ve bakteriyel enfeksiyonların özelliği olan hücrelerin büyümesini ve çoğalmasını sınırlayabilir. Metotreksat, bir rekabetçi engelleyici DHFR, DHFR'yi inhibe eden böyle bir antikanser ilaçtır.[34] Diğer ilaçlar arasında trimetoprim ve pirimetamin. Bu üçü, antitümör ve antimikrobiyal ajanlar olarak yaygın olarak kullanılmaktadır.[35] Genel olarak DHFR'yi ve özellikle de bakteriyel DHFR'leri hedefleyen diğer bileşik sınıfları, diaminopteridinler, diaminotriazinler, diaminopirolokinazinler, stilbenler, kalonlar, deoksibenzoinler gibi sınıflara dahildir.[36]
Trimetoprimin çeşitli Gram pozitif bakteriyel patojenler.[37] Bununla birlikte, trimetoprim ve DHFR'yi hedefleyen diğer ilaçlara direnç, terapötik kullanımlarının başarısını sınırlayan çeşitli mekanizmalar nedeniyle ortaya çıkabilir.[38][39][40] Direnç, DHFR gen amplifikasyonundan kaynaklanabilir, mutasyonlar DHFR'de,[41][42] diğerleri arasında ilaç alımında azalma. Ne olursa olsun, trimetoprim ve sülfametoksazol kombinasyon halinde onlarca yıldır antibakteriyel bir ajan olarak kullanılmaktadır.[37]
Folat büyüme için gereklidir,[43] ve folat metabolizmasının yolu, kanser tedavilerinin geliştirilmesinde bir hedeftir. DHFR böyle bir hedeftir. Bir rejim floroürasil, doksorubisin ve metotreksatın ilerlemiş mide kanseri olan hastalarda sağkalımı uzattığı gösterilmiştir.[44] DHFR inhibitörleri ile ilgili daha fazla araştırma, kanseri tedavi etmek için daha fazla yol sağlayabilir.
Bakterilerin büyümesi ve çoğalması için DHFR'ye de ihtiyacı vardır ve bu nedenle bakteriyel DHFR için seçici olan inhibitörler antibakteriyel maddeler olarak uygulama alanı bulmuşlardır.[37]
Dihidrofolat redüktaz inhibitörleri olarak kullanılan küçük molekül sınıfları arasında diaminokinazolin ve diaminopirolokinazolin,[45] diaminopirimidin, diaminopteridin ve diaminotriazinler.[46]
Potansiyel şarbon tedavisi

Dihidrofolat redüktaz Bacillus anthracis (BaDHFR) bulaşıcı hastalık olan şarbonun tedavisinde doğrulanmış bir ilaç hedefi. BaDHFR daha az hassastır trimetoprim diğer türlerden dihidrofolat redüktazdan daha analoglar Escherichia coli, Staphylococcus aureus, ve Streptococcus pneumoniae. Dört türden dihidrofolat redüktazın yapısal hizalaması, yalnızca BaDHFR'nin kombinasyona sahip olduğunu gösterir. fenilalanin ve tirozin sırasıyla 96 ve 102. pozisyonlarda.
BaDHFR'nin trimetoprim analoglar bu iki kalıntıdan (F96 ve Y102) kaynaklanır ve bunlar aynı zamanda gelişmiş kinetik ve katalitik verimlilik sağlar.[47] Mevcut araştırma, yeni antifolat inhibitörleri için kurşun optimizasyonuna rehberlik etmek için BaDHFR'deki aktif site mutantlarını kullanır.[47]
Araştırma aracı olarak
DHFR, tespit etmek için bir araç olarak kullanılmıştır. protein-protein etkileşimleri içinde protein parçası tamamlama deneyi (PCA).
CHO hücreleri
DHFR eksik CHO hücreleri en yaygın kullanılanlardır hücre çizgisi rekombinant proteinlerin üretimi için. Bu hücreler transfekte Birlikte plazmid taşımak dhfr gen ve rekombinant protein için gen tek bir ifade sistemi ve sonra tabi seçici koşullar timidin eksikliğinde orta. Sadece ilgili gen ile birlikte eksojen DHFR genine sahip hücreler hayatta kalır.
Etkileşimler
Dihidrofolat redüktaz ile etkileşime girdiği gösterilmiştir. GroEL[48] ve Mdm2.[49]
Etkileşimli yol haritası
İlgili makalelere bağlanmak için aşağıdaki genlere, proteinlere ve metabolitlere tıklayın.[§ 1]

- ^ Etkileşimli yol haritası, WikiPathways'de düzenlenebilir: "FloropirimidinActivity_WP1601".
Referanslar
- ^ a b c GRCh38: Ensembl sürümü 89: ENSG00000228716 - Topluluk, Mayıs 2017
- ^ a b c GRCm38: Topluluk sürümü 89: ENSMUSG00000021707 - Topluluk, Mayıs 2017
- ^ "İnsan PubMed Referansı:". Ulusal Biyoteknoloji Bilgi Merkezi, ABD Ulusal Tıp Kütüphanesi.
- ^ "Mouse PubMed Referansı:". Ulusal Biyoteknoloji Bilgi Merkezi, ABD Ulusal Tıp Kütüphanesi.
- ^ Chen MJ, Shimada T, Moulton AD, Harrison M, Nienhuis AW (Aralık 1982). "İntronless insan dihidrofolat redüktaz genleri, işlenmiş RNA moleküllerinden türetilir". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 79 (23): 7435–9. Bibcode:1982PNAS ... 79.7435C. doi:10.1073 / pnas.79.23.7435. PMC 347354. PMID 6961421.
- ^ Chen MJ, Shimada T, Moulton AD, Cline A, Humphries RK, Maizel J, Nienhuis AW (Mart 1984). "Fonksiyonel insan dihidrofolat redüktaz geni". Biyolojik Kimya Dergisi. 259 (6): 3933–43. PMID 6323448.
- ^ Funanage VL, Myoda TT, Moses PA, Cowell HR (Ekim 1984). "İnsan dihidrofolat redüktaz geninin kromozom 5'in q11 ---- q22 bölgesine atanması". Moleküler ve Hücresel Biyoloji. 4 (10): 2010–6. doi:10.1128 / mcb.4.10.2010. PMC 369017. PMID 6504041.
- ^ Smith SL, Patrick P, Stone D, Phillips AW, Burchall JJ (Kasım 1979). "Domuz karaciğeri dihidrofolat redüktaz. Saflaştırma, özellikler ve amino asit dizisi". Biyolojik Kimya Dergisi. 254 (22): 11475–84. PMID 500653.
- ^ Matthews DA, Alden RA, Bolin JT, Freer ST, Hamlin R, Xuong N, Kraut J, Poe M, Williams M, Hoogsteen K (Temmuz 1977). "Dihidrofolat redüktaz: metotreksat ile ikili kompleksin x-ışını yapısı". Bilim. 197 (4302): 452–5. Bibcode:1977Sci ... 197..452M. doi:10.1126 / science.17920. PMID 17920.
- ^ Filman DJ, Bolin JT, Matthews DA, Kraut J (Kasım 1982). "Kristal yapıları Escherichia coli ve Lactobacillus casei 1.7 Å çözünürlükte rafine edilmiş dihidrofolat redüktaz. II. Bağlı NADPH ortamı ve kataliz için çıkarımlar ". Biyolojik Kimya Dergisi. 257 (22): 13663–72. PMID 6815179.
- ^ a b Osborne MJ, Schnell J, Benkovic SJ, Dyson HJ Wright PE (Ağustos 2001). "Dihidrofolat redüktaz komplekslerinde omurga dinamiği: katalitik mekanizmada döngü esnekliğinin rolü". Biyokimya. 40 (33): 9846–59. doi:10.1021 / bi010621k. PMID 11502178.
- ^ Bolin JT, Filman DJ, Matthews DA, Hamlin RC, Kraut J (Kasım 1982). "Kristal yapıları Escherichia coli ve Lactobacillus casei 1.7 Å çözünürlükte rafine edilmiş dihidrofolat redüktaz. I. Metotreksatın genel özellikleri ve bağlanması ". Biyolojik Kimya Dergisi. 257 (22): 13650–62. PMID 6815178.
- ^ a b "Entrez Geni: DHFR dihidrofolat redüktaz".
- ^ a b Schnell JR, Dyson HJ, Wright PE (2004). "Dihidrofolat redüktazın yapısı, dinamikleri ve katalitik işlevi". Biyofizik ve Biyomoleküler Yapının Yıllık Değerlendirmesi. 33 (1): 119–40. doi:10.1146 / annurev.biophys.33.110502.133613. PMID 15139807. S2CID 28611812.
- ^ Urlaub G, Chasin LA (Temmuz 1980). "Dihidrofolat redüktaz aktivitesinden yoksun Çin hamster hücre mutantlarının izolasyonu". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 77 (7): 4216–20. Bibcode:1980PNAS ... 77.4216U. doi:10.1073 / pnas.77.7.4216. PMC 349802. PMID 6933469.
- ^ Crabtree MJ, Tatham AL, Hale AB, Alp NJ, Channon KM (Ekim 2009). "Endotelyal nitrik oksit sentaz bağlanmasının düzenlenmesinde dihidrofolat redüktaz tarafından tetrahidrobiopterin geri dönüşümünün kritik rolü: de novo biopterin sentezinin kurtarma yollarına göre göreceli önemi". Biyolojik Kimya Dergisi. 284 (41): 28128–36. doi:10.1074 / jbc.M109.041483. PMC 2788863. PMID 19666465.
- ^ a b c d Rod TH, Brooks CL (Temmuz 2003). "Dihidrofolat redüktaz, dihidrofolatın protonasyonunu nasıl kolaylaştırır". Amerikan Kimya Derneği Dergisi. 125 (29): 8718–9. doi:10.1021 / ja035272r. PMID 12862454.
- ^ a b Wan Q, Bennett BC, Wilson MA, Kovalevsky A, Langan P, Howell EE, Dealwis C (Aralık 2014). "Nötron ve ultra yüksek çözünürlüklü X-ışını kristalografisi kullanarak dihidrofolat redüktazın katalitik mekanizmasını çözmeye doğru". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 111 (51): 18225–30. Bibcode:2014PNAS..11118225W. doi:10.1073 / pnas.1415856111. PMC 4280638. PMID 25453083.
- ^ a b Liu CT, Francis K, Layfield JP, Huang X, Hammes-Schiffer S, Kohen A, Benkovic SJ (Aralık 2014). "Escherichia coli dihidrofolat redüktaz katalizli proton ve hidrit transferleri: zamansal düzen ve Asp27 ve Tyr100'ün rolleri ". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 111 (51): 18231–6. Bibcode:2014PNAS..11118231L. doi:10.1073 / pnas.1415940111. PMC 4280594. PMID 25453098.
- ^ a b Czekster CM, Vandemeulebroucke A, Blanchard JS (Ocak 2011). "Dihidrofolat redüktazın kinetik ve kimyasal mekanizması Tüberküloz". Biyokimya. 50 (3): 367–75. doi:10.1021 / bi1016843. PMC 3074011. PMID 21138249.
- ^ a b c Fierke CA, Johnson KA, Benkovic SJ (Haziran 1987). "Dihidrofolat redüktaz ile ilişkili kinetik şemanın oluşturulması ve değerlendirilmesi Escherichia coli". Biyokimya. 26 (13): 4085–92. doi:10.1021 / bi00387a052. PMID 3307916.
- ^ Reyes VM, Sawaya MR, Brown KA, Kraut J (Şubat 1995). "İzomorf kristal yapıları Escherichia coli folat, 5-deazafolat ve 5,10-dideazatetrahidrofolat ile komplekslenmiş dihidrofolat redüktaz: mekanik çıkarımlar ". Biyokimya. 34 (8): 2710–23. doi:10.1021 / bi00008a039. PMID 7873554.
- ^ a b c Sawaya MR, Kraut J (Ocak 1997). "Mekanizmasındaki döngü ve alt alan hareketleri Escherichia coli dihidrofolat redüktaz: kristalografik kanıt ". Biyokimya. 36 (3): 586–603. doi:10.1021 / bi962337c. PMID 9012674.
- ^ Chen YQ, Kraut J, Blakley RL, Callender R (Haziran 1994). "Dihidrofolat redüktaza bağlı dihidrofolatın N5'in pKa'sının Raman spektroskopisi ile belirlenmesi: mekanik etkiler". Biyokimya. 33 (23): 7021–6. doi:10.1021 / bi00189a001. PMID 8003467.
- ^ Shrimpton P, Allemann RK (Haziran 2002). "E. coli dihidrofolat redüktazın katalitik döngüsünde suyun rolü". Protein Bilimi. 11 (6): 1442–51. doi:10.1110 / ps.5060102. PMC 2373639. PMID 12021443.
- ^ Antikainen NM, Smiley RD, Benkovic SJ, Hammes GG (Aralık 2005). "Konformasyona bağlı enzim katalizi: dihidrofolat redüktazın tek moleküllü ve geçici kinetik araştırması". Biyokimya. 44 (51): 16835–43. doi:10.1021 / bi051378i. PMID 16363797.
- ^ Narayana N, Matthews DA, Howell EE, Nguyen-huu X (Kasım 1995). "Trimetoprim dirençli bakterilerden plazmidle kodlanmış bir dihidrofolat redüktaz yeni bir D2 simetrik aktif bölgeye sahiptir". Doğa Yapısal Biyoloji. 2 (11): 1018–25. doi:10.1038 / nsb1195-1018. PMID 7583655. S2CID 11914241.
- ^ Bradrick TD, Beechem JM, Howell EE (Eylül 1996). "Bir D2 simetrik protein olan R67 dihidrofolat redüktazın tek aktif gözeneğinde ikili ve üçlü kompleks oluşumu sırasında olağandışı bağlanma stokiyometrileri ve işbirliği gözlemlenir". Biyokimya. 35 (35): 11414–24. doi:10.1021 / bi960205d. PMID 8784197.
- ^ Park H, Zhuang P, Nichols R, Howell EE (Ocak 1997). "R67 dihidrofolat redüktazın mekanik çalışmaları. PH ve H62C mutasyonunun etkileri". Biyolojik Kimya Dergisi. 272 (4): 2252–8. doi:10.1074 / jbc.272.4.2252. PMID 8999931.
- ^ Kamath G, Howell EE, Agarwal PK (Ekim 2010). "Köpeği sallayan kuyruk: R67 dihidrofolat redüktazdaki katalize bakışlar". Biyokimya. 49 (42): 9078–88. doi:10.1021 / bi1007222. PMID 20795731.
- ^ Cowman AF, Lew AM (Kasım 1989). "Antifolat ilaç seçimi, Plasmodium chabaudi'de kromozom 7'nin kopyalanması ve yeniden düzenlenmesiyle sonuçlanır". Moleküler ve Hücresel Biyoloji. 9 (11): 5182–8. doi:10.1128 / mcb.9.11.5182. PMC 363670. PMID 2601715.
- ^ Banka S, Blom HJ, Walter J, Aziz M, Urquhart J, Clouthier CM, vd. (Şubat 2011). "Dihidrofolat redüktaz eksikliğinin neden olduğu doğuştan metabolizma hatasının tanımlanması ve karakterizasyonu". Amerikan İnsan Genetiği Dergisi. 88 (2): 216–25. doi:10.1016 / j.ajhg.2011.01.004. PMC 3035707. PMID 21310276.
- ^ Nyhan WL, Hoffmann GF, Barshop BA (30 Aralık 2011). Kalıtsal Metabolizma Hastalıkları Atlası 3E. CRC Basın. s. 141–. ISBN 978-1-4441-4948-7.
- ^ Li R, Sirawaraporn R, Chitnumsub P, Sirawaraporn W, Wooden J, Athappilly F, Turley S, Hol WG (Ocak 2000). "Üç boyutlu yapısı M. tuberculosis dihidrofolat redüktaz, yeni tüberküloz ilaçlarının tasarımı için fırsatlar ortaya koymaktadır ". Moleküler Biyoloji Dergisi. 295 (2): 307–23. doi:10.1006 / jmbi.1999.3328. PMID 10623528. S2CID 24527344.
- ^ Benkovic SJ, Fierke CA, Naylor AM (Mart 1988). "Dihidrofolat redüktaz mutantları üzerinde yapılan çalışmalardan enzim işlevi hakkında bilgiler". Bilim. 239 (4844): 1105–10. Bibcode:1988Sci ... 239.1105B. doi:10.1126 / science.3125607. PMID 3125607.
- ^ Srinivasan B, Tonddast-Navaei S, Roy A, Zhou H, Skolnick J (Eylül 2018). "Kimyasal uzay Escherichia coli dihidrofolat redüktaz inhibitörleri: Eski böcekler için yeni ilaçları keşfetmeye yönelik yeni yaklaşımlar ". Tıbbi Araştırma İncelemeleri. 39 (2): 684–705. doi:10.1002 / med.21538. PMC 6370515. PMID 30192413.
- ^ a b c Hawser S, Lociuro S, Islam K (Mart 2006). "Antibakteriyel maddeler olarak dihidrofolat redüktaz inhibitörleri". Biyokimyasal Farmakoloji. 71 (7): 941–8. doi:10.1016 / j.bcp.2005.10.052. PMID 16359642.
- ^ Narayana N, Matthews DA, Howell EE, Nguyen-huu X (Kasım 1995). "Trimetoprim dirençli bakterilerden plazmidle kodlanmış bir dihidrofolat redüktaz, yeni bir D2 simetrik aktif bölgeye sahiptir". Doğa Yapısal Biyoloji. 2 (11): 1018–25. doi:10.1038 / nsb1195-1018. PMID 7583655. S2CID 11914241.
- ^ Huennekens FM (Haziran 1996). "Dihidrofolat redüktaz arayışında". Protein Bilimi. 5 (6): 1201–8. doi:10.1002 / pro.5560050626. PMC 2143423. PMID 8762155.
- ^ Banerjee D, Mayer-Kuckuk P, Capiaux G, Budak-Alpdogan T, Gorlick R, Bertino JR (Temmuz 2002). "Dihidrofolat redüktaz ve timidilat sentazı hedefleyen ilaçlara direncin yeni yönleri". Biochimica et Biophysica Açta (BBA) - Hastalığın Moleküler Temeli. 1587 (2–3): 164–73. doi:10.1016 / S0925-4439 (02) 00079-0. PMID 12084458.
- ^ Toprak E, Veres A, Michel JB, Chait R, Hartl DL, Kishony R (Aralık 2011). "Dinamik olarak sürdürülen ilaç seçimi altında antibiyotik direncine giden evrimsel yollar". Doğa Genetiği. 44 (1): 101–5. doi:10.1038 / ng.1034. PMC 3534735. PMID 22179135.
- ^ Rodrigues JV, Bershtein S, Li A, Lozovsky ER, Hartl DL, Shakhnovich EI (Mart 2016). "Biyofiziksel ilkeler, ilaç direncinin uygunluk manzaralarını öngörüyor". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 113 (11): E1470-8. Bibcode:2016PNAS..113E1470R. doi:10.1073 / pnas.1601441113. PMC 4801265. PMID 26929328.
- ^ Bailey SW, Ayling JE (Eylül 2009). "İnsan karaciğerindeki dihidrofolat redüktazın son derece yavaş ve değişken aktivitesi ve bunun yüksek folik asit alımı üzerindeki etkileri". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 106 (36): 15424–9. doi:10.1073 / pnas.0902072106. PMC 2730961. PMID 19706381.
- ^ Murad AM, Santiago FF, Petroianu A, Rocha PR, Rodrigues MA, Rausch M (Temmuz 1993). "İleri mide kanserinde 5-florourasil, doksorubisin ve metotreksat ile modifiye edilmiş tedavi". Kanser. 72 (1): 37–41. doi:10.1002 / 1097-0142 (19930701) 72: 1 <37 :: AID-CNCR2820720109> 3.0.CO; 2-P. PMID 8508427.
- ^ Srinivasan B, Skolnick J (Mayıs 2015). "Yavaş başlangıçlı sıkı bağlanma inhibisyonuna ilişkin bilgiler Escherichia coli dihidrofolat redüktaz: yeni sıkı bağlanma inhibitörleri olarak pirrolo [3,2-f] kinazolin-1,3-diamin ve türevlerinin ayrıntılı mekanik karakterizasyonu ". FEBS Dergisi. 282 (10): 1922–38. doi:10.1111 / Şub.13244. PMC 4445455. PMID 25703118.
- ^ Srinivasan B, Tonddast-Navaei S, Skolnick J (Ekim 2015). "Ligand bağlanma çalışmaları, ön yapı-aktivite ilişkisi ve 1-fenil-6,6-dimetil-1,3,5-triazin-2,4-diamin türevlerinin inhibitörleri olarak ayrıntılı mekanik karakterizasyonu Escherichia coli dihidrofolat redüktaz ". Avrupa Tıbbi Kimya Dergisi. 103: 600–14. doi:10.1016 / j.ejmech.2015.08.021. PMC 4610388. PMID 26414808.
- ^ a b Beierlein JM, Karri NG, Anderson AC (Ekim 2010). "Bacillus anthracis dihidrofolat redüktazın hedeflenen mutasyonları, karmaşık yapı − aktivite ilişkileri". Tıbbi Kimya Dergisi. 53 (20): 7327–36. doi:10.1021 / jm100727t. PMC 3618964. PMID 20882962.
- ^ Mayhew M, da Silva AC, Martin J, Erdjument-Bromage H, Tempst P, Hartl FU (Şubat 1996). "GroEL-GroES şaperonin kompleksinin merkezi boşluğunda protein katlanması". Doğa. 379 (6564): 420–6. Bibcode:1996Natur.379..420M. doi:10.1038 / 379420a0. PMID 8559246. S2CID 4310511.
- ^ Maguire M, Nield PC, Devling T, Jenkins RE, Park BK, Polański R, Vlatković N, Boyd MT (Mayıs 2008). "MDM2, monobikitinasyon yoluyla dihidrofolat redüktaz aktivitesini düzenler". Kanser araştırması. 68 (9): 3232–42. doi:10.1158 / 0008-5472.CAN-07-5271. PMC 3536468. PMID 18451149.
daha fazla okuma
- Joska TM, Anderson AC (Ekim 2006). "Bacillus cereus ve Bacillus anthracis dihidrofolat redüktazın yapı-aktivite ilişkileri: yeni güçlü ilaç liderlerinin tanımlanmasına doğru". Antimikrobiyal Ajanlar ve Kemoterapi. 50 (10): 3435–43. doi:10.1128 / AAC.00386-06. PMC 1610094. PMID 17005826.
- Chan DC, Fu H, Forsch RA, Queener SF, Rosowsky A (Haziran 2005). "Yeni piritreksim analoglarının tasarımı, sentezi ve yan zincirde omega-karboksialkoksi veya omega-karboksi-1-alkinil ikameli diğer diaminopirimidin dihidrofolat redüktaz inhibitörlerinin antifolat aktivitesi". Tıbbi Kimya Dergisi. 48 (13): 4420–31. doi:10.1021 / jm0581718. PMID 15974594.
- Banerjee D, Mayer-Kuckuk P, Capiaux G, Budak-Alpdogan T, Gorlick R, Bertino JR (Temmuz 2002). "Dihidrofolat redüktaz ve timidilat sentazı hedefleyen ilaçlara direncin yeni yönleri". Biochimica et Biophysica Açta (BBA) - Hastalığın Moleküler Temeli. 1587 (2–3): 164–73. doi:10.1016 / S0925-4439 (02) 00079-0. PMID 12084458.
- Stockman BJ, Nirmala NR, Wagner G, Delcamp TJ, DeYarman MT, Freisheim JH (Ocak 1992). "Çözeltide insan dihidrofolat redüktaz için diziye özgü 1H ve 15N rezonans atamaları". Biyokimya. 31 (1): 218–29. doi:10.1021 / bi00116a031. PMID 1731871.
- Beltzer JP, Spiess M (Aralık 1991). "Asialoglikoprotein reseptörünün plazma membranı kaplı veziküllerin beta adaptörüne in vitro bağlanması". EMBO Dergisi. 10 (12): 3735–42. doi:10.1002 / j.1460-2075.1991.tb04942.x. PMC 453108. PMID 1935897.
- Davies JF, Delcamp TJ, Prendergast NJ, Ashford VA, Freisheim JH, Kraut J (Ekim 1990). "Folat ve 5-deazafolat ile kompleks haline getirilmiş rekombinant insan dihidrofolat redüktazın kristal yapıları". Biyokimya. 29 (40): 9467–79. doi:10.1021 / bi00492a021. PMID 2248959.
- Will CL, Dolnick BJ (Aralık 1989). "5-Florourasil, metotreksata dirençli KB hücrelerinde dihidrofolat redüktaz öncü mRNA işlemesini ve / veya nükleer mRNA stabilitesini inhibe eder". Biyolojik Kimya Dergisi. 264 (35): 21413–21. PMID 2592384.
- Masters JN, Attardi G (Mart 1985). "Polisomal RNA haritasında 5 'uçları ana mRNA başlangıç bölgesinin birkaç yüz nükleotid yukarısında bulunan ayrı insan dihidrofolat redüktaz geni transkriptleri". Moleküler ve Hücresel Biyoloji. 5 (3): 493–500. doi:10.1128 / mcb.5.3.493. PMC 366741. PMID 2859520.
- Miszta H, Dabrowski Z, Lanotte M (Kasım 1988). "Kemik iliği stromal hücrelerinde enzimik tetrahidrofolat dehidrojenaz (EC 1.5.1.3) ekspresyonunun in vitro modelleri". Lösemi. 2 (11): 754–9. PMID 3185016.
- Oefner C, D'Arcy A, Winkler FK (Haziran 1988). "Folat ile kompleks oluşturulmuş insan dihidrofolat redüktazın kristal yapısı". Avrupa Biyokimya Dergisi / FEBS. 174 (2): 377–85. doi:10.1111 / j.1432-1033.1988.tb14108.x. PMID 3383852.
- Yang JK, Masters JN, Attardi G (Haziran 1984). "İnsan dihidrofolat redüktaz gen organizasyonu. G + C bakımından zengin 5 'kodlamayan dizinin kapsamlı korunması ve homolog memeli genlerinden güçlü intron boyutu sapması". Moleküler Biyoloji Dergisi. 176 (2): 169–87. doi:10.1016/0022-2836(84)90419-4. PMID 6235374.
- Masters JN, Yang JK, Cellini A, Attardi G (Haziran 1983). "Bir insan dihidrofolat redüktaz psödojeni ve bunun, spesifik haberci RNA'nın çoklu formları ile ilişkisi". Moleküler Biyoloji Dergisi. 167 (1): 23–36. doi:10.1016 / S0022-2836 (83) 80032-1. PMID 6306253.
- Chen MJ, Shimada T, Moulton AD, Cline A, Humphries RK, Maizel J, Nienhuis AW (Mart 1984). "Fonksiyonel insan dihidrofolat redüktaz geni". Biyolojik Kimya Dergisi. 259 (6): 3933–43. PMID 6323448.
- Funanage VL, Myoda TT, Moses PA, Cowell HR (Ekim 1984). "İnsan dihidrofolat redüktaz geninin kromozom 5'in q11 ---- q22 bölgesine atanması". Moleküler ve Hücresel Biyoloji. 4 (10): 2010–6. doi:10.1128 / mcb.4.10.2010. PMC 369017. PMID 6504041.
- Ustalar JN, Attardi G (1983). "İnsan dihidrofolik asit redüktazı kodlayan cDNA'nın nükleotid dizisi". Gen. 21 (1–2): 59–63. doi:10.1016/0378-1119(83)90147-6. PMID 6687716.
- Morandi C, Masters JN, Mottes M, Attardi G (Nisan 1982). "İnsan dihidrofolat redüktaz haberci RNA'nın çoklu formları. Klonlama ve ifade Escherichia coli DNA kodlama dizilerinin ". Moleküler Biyoloji Dergisi. 156 (3): 583–607. doi:10.1016/0022-2836(82)90268-6. PMID 6750132.
- Bonifaci N, Sitia R, Rubartelli A (Eylül 1995). "HIV Tat içeren bir eksojen füzyon proteininin nükleer translokasyonu, açılmayı gerektirir". AIDS. 9 (9): 995–1000. doi:10.1097/00002030-199509000-00003. PMID 8527095. S2CID 8417864.
- Mayhew M, da Silva AC, Martin J, Erdjument-Bromage H, Tempst P, Hartl FU (Şubat 1996). "GroEL-GroES şaperonin kompleksinin merkezi boşluğunda protein katlanması". Doğa. 379 (6564): 420–6. Bibcode:1996Natur.379..420M. doi:10.1038 / 379420a0. PMID 8559246. S2CID 4310511.
- Gross M, Robinson CV, Mayhew M, Hartl FU, Radford SE (Aralık 1996). "GroEL'e bağlı DHFR'de önemli hidrojen değişimi koruması, substrat döngüsünün yinelemeli turları sırasında korunur". Protein Bilimi. 5 (12): 2506–13. doi:10.1002 / pro.5560051213. PMC 2143321. PMID 8976559.
- Schleiff E, Shore GC, Goping IS (Mart 1997). "İnsan mitokondriyal ithalat reseptörü, Tom20p. Tom20-glutatyon S-transferaz ve mitokondriyal öncü proteinler arasındaki spesifik etkileşimleri ortaya çıkarmak için glutatyon kullanımı". FEBS Mektupları. 404 (2–3): 314–8. doi:10.1016 / S0014-5793 (97) 00145-2. PMID 9119086. S2CID 29177508.
- Cody V, Galitsky N, Luft JR, Pangborn W, Rosowsky A, Blakley RL (Kasım 1997). "Nikotinamid adenin dinükleotid fosfat ve çok sıkı bağlanma inhibitörü PT523 ile indirgenmiş insan dihidrofolat redüktaz üçlü komplekslerinin iki bağımsız kristal yapısının karşılaştırılması". Biyokimya. 36 (45): 13897–903. doi:10.1021 / bi971711l. PMID 9374868.
- Vanguri VK, Wang S, Godyna S, Ranganathan S, Liau G (Nisan 2000). "Trombospondin-1, polihistidine yüksek afinite ve özgüllükle bağlanır". Biyokimyasal Dergi. 347 (Pt 2): 469–73. doi:10.1042/0264-6021:3470469. PMC 1220979. PMID 10749676.
Dış bağlantılar
- 1988 Nobel Tıp dersi
- Proteopedia: Dihidrofolat redüktaz
- Mevcut tüm yapısal bilgilere genel bakış PDB için UniProt: P00374 (Dihidrofolat redüktaz) PDBe-KB.
PDB galerisi | |
|---|---|
|