Foraminifer - Foraminifera
| Foraminifer | |
|---|---|
 | |
| Canlı Amonyak tepida (Rotaliida) | |
| bilimsel sınıflandırma | |
| Alan adı: | |
| (rütbesiz): | |
| (rütbesiz): | |
| Şube: | |
| Altfilum: | Foraminifer d'Orbigny, 1826 |
| Alt bölümler | |
"Monothalamea "
incertae sedis | |
Foraminifer (/fəˌræməˈnɪfərə/; Latince "delik taşıyıcılar" için; gayri resmi olarak adlandırılan "foramlar") tek hücreli organizmalardır, filum veya sınıf nın-nin hareketsiz protistler ayrıntılı akışla karakterize ektoplazma yiyecek toplamak ve diğer kullanımlar için; ve genellikle bir dış kabuk ("Ölçek ") çeşitli form ve materyallerin testleri. Chitin (bazı basit cinslerde bulunur ve Textularia özellikle) en ilkel tür olduğuna inanılmaktadır. Çoğu foraminifer denizeldir ve çoğu deniz tabanı tortusu üzerinde veya içinde yaşar (yani Bentik ), daha küçük bir sayı ise su sütununda çeşitli derinliklerde yüzerken (yani, planktonik ). Tatlı su veya acı koşullardan daha azı bilinmektedir ve küçük alt birimin moleküler analizi yoluyla çok az sayıda (akuatik olmayan) toprak türü tanımlanmıştır. ribozomal DNA.[2][3]
Foraminifera tipik olarak bir Ölçek veya bir veya birden fazla hazneye sahip olabilen kabuk, bazıları yapı olarak oldukça ayrıntılı hale gelir.[4] Bu kabuklar genellikle şunlardan yapılır: kalsiyum karbonat (CaCO
3) veya yapıştırılmış tortu parçacıkları. Her ikisi de yaşayan 50.000'den fazla tür tanınmaktadır (10.000)[5] ve fosil (40,000).[6][7] Genellikle 1 mm'den küçüktürler, ancak bazıları çok daha büyüktür. en büyük türler 20 cm'ye kadar ulaşır.[8]
Modern bilimsel İngilizcede terim foraminifera hem tekil hem de çoğuldur (kelimenin ne olduğuna bakılmaksızın) Latince türetme) ve bir veya daha fazla örnek veya taksonu tanımlamak için kullanılır: tekil veya çoğul olarak kullanımı bağlamdan belirlenmelidir. Foraminifera, grubu tanımlamak için sıklıkla gayri resmi olarak kullanılır ve bu durumlarda genellikle küçük harflidir.[9]
Çalışma tarihi
Foraminifer için bilinen en eski atıf, Herodot, MÖ 5. yüzyılda onları oluşturan kayayı oluşturan Büyük Giza Piramidi. Bunlar bugün cinsin temsilcileri olarak kabul edilmektedir. Nummulitler. Strabo MÖ 1. yüzyılda aynı foraminiferleri not etti ve bunların kalıntıları olduğunu öne sürdü. mercimek piramitleri inşa eden işçiler tarafından bırakıldı.[10]
Robert Hooke 1665 kitabında açıklandığı ve gösterildiği gibi mikroskop altında bir foraminifer gözlemledi Mikrografi:
Birkaç küçük ve tek Büyüteç'i deniyordum ve rasgele bir beyaz Kum paketini izliyordum, tahıllardan birinin tam olarak şekillendiğini ve bir Kabuk istediğini algıladığımda [...] Her şeyi görüyorum daha iyi bir Mikroskopla ve her iki tarafta ve kenarlarda, düz spiral Kabuklu küçük bir Su Salyangozunun Kabuğuna benzemek için bulundu [...][11]
Antonie van Leeuwenhoek 1700'deki foraminifer testleri tanımlanmış ve gösterilmiş, bunları küçük kırışıklıklar olarak tanımlamıştır; onun illüstrasyonu olarak tanınabilir Elphidium.[12] Erken işçiler foraminiferleri cins içinde sınıflandırdılar Nautilus, belli olanlara benzerliklerini not ederek kafadanbacaklılar. Tarafından tanındı Lorenz Spengler 1781'de foraminiferin septada delikler vardı, bu da sonunda gruba adını verecekti.[13] Spengler ayrıca foraminifer septasının nautili septanın tersi olduğunu ve bir sinir tüpünden yoksun olduklarını belirtti.[14]

Alcide d'Orbigny, 1826 çalışmasında, onları bir dakika grubu olarak gördü kafadanbacaklılar ve onların garip morfolojisine dikkat çekerek, sözde ayağı dokunaçlar olarak yorumladı ve yüksek oranda azalmış (gerçekte, yok) kafayı not etti.[15] Gruba isim verdi foraminifèresveya "delik taşıyıcılar", çünkü grubun üyeleri, kabuklarındaki bölmeler arasındaki bölmelerde deliklere sahipti. Nautili veya ammonitler.[9]
Foraminiferin protozoan doğası ilk olarak Dujardin 1835'te.[13] Kısa bir süre sonra, 1852'de, d'Orbigny, test şekline göre sınıflandırdığı 72 foraminifer cinsini tanıyan bir sınıflandırma şeması üretti - bu, meslektaşlarından ciddi eleştiriler alan bir şema.[12]
H.B. Brady 1884 tarihli monografide, foraminifer buluntuları Challenger sefer. Brady, stratigrafik aralığa çok az bakılmaksızın 29 alt aileye sahip 10 aileyi tanımıştır; taksonomisi, birden çok farklı karakterin taksonomik grupları ayırması gerektiği fikrini vurguladı ve bu nedenle, birbirine yakın ve kalkerli cinsler yerleştirildi.
Bu genel sınıflandırma şeması şu tarihe kadar kalacaktır: Cushman 1920'lerin sonunda çalışmak. Cushman, duvar kompozisyonunu foraminiferlerin sınıflandırılmasında en önemli tek özellik olarak gördü; sınıflandırması geniş çapta kabul gördü, ancak aynı zamanda meslektaşlarından "biyolojik olarak sağlam olmadığı" için eleştiri aldı.
Cushman'ın planı yine de, test duvarının mikro yapısına dayalı olarak bugün hala kullanılan genel gruplara foraminiferleri yerleştiren Tappan ve Loeblich'in 1964 sınıflandırmasına kadar baskın sınıflandırma şeması olarak kaldı.[12] Bu gruplar, üst düzey sınıflandırmanın farklı şemalarına göre çeşitli şekillerde hareket ettirilmiştir. Pawlowski'nin (2013) moleküler sistematiği kullanımı genellikle Tappan ve Loeblich'in gruplarını doğruladı, bazıları polifilik veya parafiletik olarak bulundu; bu çalışma aynı zamanda başlıca foraminifer grupları arasındaki üst düzey ilişkilerin belirlenmesine de yardımcı olmuştur.[16]
Taksonomi
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Pawlowski ve ark.'nın ardından Foraminifera'nın Filogeni. 2013.[16] Monotalamid düzenleri Astrorhizida ve Allogromiida her ikisi de parafiletiktir. |
Foraminifera'nın taksonomik konumu 1854'teki Schultze'den bu yana değişmiştir.[17] sipariş olarak anılan Foraminiferida. Loeblich ve Tappan (1992) Foraminifera'yı bir sınıf olarak yeniden kaydetti[18] şimdi yaygın olarak kabul edildiği gibi.
Foraminiferler tipik olarak Protozoa,[19][20][21] veya benzer Protoctista'da veya Protist krallık.[22][23] Öncelikle aşağıdakilere dayanan ikna edici kanıtlar moleküler filogenetik, Protozoa içinde büyük bir gruba ait oldukları için vardır. Rhizaria.[19] Rhizaria üyeleri arasındaki evrimsel ilişkilerin tanınmasından önce, Foraminiferler genellikle diğerleriyle gruplandırılmıştı. amoeboidler Granuloreticulosa sınıfında filum Rhizopodea (veya Sarcodina) olarak.
Rhizaria sorunludur, çünkü bunlar yerleşik bir grup kullanmaktan ziyade genellikle "süper grup" olarak adlandırılır. taksonomik sıralama gibi filum. Cavalier-Smith, Rhizaria'yı Protozoa krallığı içindeki bir infra-krallık olarak tanımlar.[19]
Bazı taksonomiler Foraminiferleri kendilerine ait bir filuma yerleştirerek, onları yerleştirildikleri amipli Sarcodina ile aynı seviyeye getirdiler.
Morfolojik bağıntılar tarafından henüz desteklenmese de, moleküler veriler Foraminifera'nın Cercozoa ve Radyolarya her ikisi de kompleks kabuklu amoeboidler içerir; bu üç grup Rhizaria'yı oluşturur.[20] Bununla birlikte, foramların diğer gruplarla ve birbirleriyle tam ilişkileri hala tam olarak açık değildir. Foraminiferler ile yakından ilgilidir testate amip.[24]
| Mikhalevich 2013'ten taksonomi[25] |
|---|
* Foraminifer d'Orbigny 1826
|
Anatomi

Çoğu foraminiferin en çarpıcı yönü sert kabukları veya testleridir. Bunlar birden fazla bölmeden birinden oluşabilir ve protein, tortu partikülleri, kalsit, aragonit veya (bir durumda) silisten oluşabilir.[18] Bazı foraminiferlerde testler tamamen eksiktir.[26] Diğer kabuk salgılayan organizmalardan farklı olarak, örneğin yumuşakçalar veya mercanlar foraminifer testleri, hücre zarı, içinde protoplazma. Hücrenin organelleri, testin bölme (ler) i içinde yer alır ve testin delik (ler) i, materyalin psödopodiden iç hücreye ve geri aktarılmasına izin verir.[27]
Foraminiferal hücre, granüler endoplazma ve şeffaf ektoplazmaya bölünmüştür. yalancı net, tek bir açıklıktan veya testteki birçok delikten ortaya çıkabilir. Bireysel yalancı ayaklılar karakteristik olarak her iki yönde akan küçük granüllere sahiptir.[28] Foraminiferler sahip olmak açısından benzersizdir granüloretiküloz psödopodi; yani psödopodileri mikroskop altında granüler görünür; bu psödopodiler genellikle uzar ve bölünebilir ve yeniden birleşebilir. Bunlar hücrenin ihtiyaçlarına uyacak şekilde genişletilebilir ve geri çekilebilir. Sözde ayaklılar, hareket, sabitleme, boşaltım, test yapımı ve diyatomlar veya bakteri gibi küçük organizmalardan oluşan yiyecekleri yakalamak için kullanılır.[29][27]
Testlerin yanı sıra, foraminiferal hücreler bir hücre iskeleti diğer amoeboidlerde görülen yapı olmadan gevşek bir şekilde düzenlenmiş mikrotübüllerin Foramlar, uzun psödopodinin hızlı oluşumuna ve geri çekilmesine izin vererek, mikrotübülleri hızlı bir şekilde birleştirmek ve sökmek için özel hücresel mekanizmalar geliştirmiştir.[18]
Gamont'ta (cinsel form), foraminiferler genellikle yalnızca tek bir çekirdeğe sahipken, agamont (aseksüel form) birden fazla çekirdeğe sahip olma eğilimindedir. En azından bazı türlerde, çekirdekler dimorfiktir ve somatik çekirdekler, üretici çekirdeklerden üç kat daha fazla protein ve RNA içerir. Bununla birlikte, nükleer anatomi oldukça çeşitli görünüyor.[30] Çekirdekler, çok odacıklı türlerde mutlaka bir odayla sınırlı değildir. Çekirdekler küresel olabilir veya çok sayıda lob içerebilir. Çekirdekler tipik olarak 30-50 um çapındadır.[31]
Bazı foraminifer türlerinin hücrelerinde büyük, boş vakuoller bulunur; bunların kesin amacı belirsizdir, ancak bir nitrat rezervuarı olarak işlev gördükleri öne sürülmüştür.[31]
Mitokondri hücre içinde eşit olarak dağılır, ancak bazı türlerde gözeneklerin altında ve hücrenin dış kenarı çevresinde yoğunlaşırlar. Bunun düşük oksijenli ortamlara bir adaptasyon olduğu varsayılmıştır. [31]
Birkaç tür zenofor alışılmadık derecede yüksek konsantrasyonlara sahip olduğu bulunmuştur. Radyoaktif İzotoplar hücrelerinde, ökaryotların en yüksekleri arasındadır. Bunun amacı bilinmemektedir.[32]
Ekoloji
Modern Foraminiferler öncelikle deniz organizmalarıdır, ancak yaşayan bireyler acı tatlı suda bulunmuştur.[28] ve hatta karasal habitatlar.[3] Türlerin çoğu Bentik ve diğer 40 morfospesiler planktonik.[29] Genetik olarak farklı birçok tür morfolojik olarak ayırt edilemez olabileceğinden, bu sayı gerçek çeşitliliğin yalnızca bir bölümünü temsil edebilir.[33]
Bentik foraminiferler tipik olarak ince taneli çökeltilerde bulunur ve burada aktif olarak katmanlar arasında hareket ederler; ancak pek çok tür sert kaya alt tabakalarında, deniz yosunlarına bağlanmış veya tortu yüzeyinin üzerinde bulunur.[18]
Planktonik foraminiferlerin çoğu, rotaliida içinde bir soy olan globigerinina'da bulunur.[16] Bununla birlikte, en az bir başka mevcut rotaliid soy, Neogallitellia, bağımsız olarak planktonik bir yaşam tarzı geliştirmiş gibi görünüyor.[34][35] Ayrıca, bazı Jurassic fosil foraminiferlerinin de bağımsız olarak planktonik bir yaşam tarzı geliştirmiş olabileceği ve Robertinida'nın üyeleri olabileceği öne sürüldü.[36]
Bir dizi foramın tek hücreli yosun gibi endosymbionts gibi çeşitli soylardan yeşil alg, kırmızı yosun, altın yosun, diyatomlar, ve Dinoflagellatlar.[29] Bunlar miksotrofik foraminiferler, besin açısından fakir okyanus sularında özellikle yaygındır.[37] Bazı foramlar kleptoplastik, tutma kloroplastlar yutulan alglerden davranışa fotosentez.[38]
Çoğu foraminifer heterotrofiktir, daha küçük organizmalar ve organik madde tüketir; bazı küçük türler özel besleyicilerdir Fitodetrit diğerleri ise diatom tüketme konusunda uzmanlaşmıştır. Bazı bentik foramlar, psöodopodiyi kullanarak kendilerini tortu ve organik parçacıkların içine hapsetmek için besleme kistleri oluşturur.[18] Bazı foraminiferler gibi küçük hayvanları avlar. kopepodlar veya Cumaceans; hatta bazı foramlar, avlarının testlerinde delikler açarak diğer foramlardan daha önce gelir.[39] Bir grubun, ksenofiyoforların testlerinde bakterileri yetiştirmeleri önerildi.[40] Süspansiyon besleme grupta da yaygındır ve en azından bazı türler çözünmüş organik karbon.[18]
Birkaç foram türü parazit, süngerler, yumuşakçalar, mercanlar ve hatta diğer foraminiferler. Parazitik stratejiler değişiklik gösterir; Bazıları ektoparazit gibi davranarak konakçıdan yiyecek çalmak için psödopodiyi kullanır, diğerleri ise yumuşak dokusuyla beslenmek için ev sahibinin kabuğu veya vücut duvarından geçerler.[18]
Foraminiferler, omurgasızlar, balıklar, kıyı kuşları ve diğer foraminiferler dahil olmak üzere bir dizi büyük organizma tarafından yenir. Bununla birlikte, bazı durumlarda avcıların, organizmaların kendisinden çok foram kabuklarından gelen kalsiyumla daha fazla ilgilenebilecekleri öne sürülmüştür. Birçok su salyangoz türünün seçici olarak foraminifer üzerinden beslendiği, hatta çoğu kez tek tek türleri tercih ettiği bilinmektedir.[41]
Bazı bentik foraminiferlerin hayatta kalabildiği bulunmuştur. anoksik 24 saatin üzerindeki koşullar, seçici olabileceklerini gösterir. anaerobik solunum. Bu, tortu-su arayüzüne yakın değişen oksijen koşullarında hayatta kalmak için bir adaptasyon olarak yorumlanıyor.[42]
Foraminiferler, okyanusun en derin kısımlarında bulunur. Mariana Çukuru, I dahil ederek Challenger Deep, bilinen en derin kısım. Bu derinliklerde karbonat telafi derinliği Testlerin kalsiyum karbonatı aşırı basınçtan dolayı suda çözünür. Challenger Deep'de bulunan Foraminifera bu nedenle karbonat testine sahip değil, bunun yerine organik materyalden birine sahip.[43]
Üreme
Genelleştirilmiş foraminiferal yaşam döngüsü, aşağıdakiler arasında bir değişim içerir: haploid ve diploid nesiller, form olarak çoğunlukla benzer olsalar da.[17][44] Haploid veya gamont başlangıçta tek bir çekirdek ve çok sayıda üretmek için böler gametler, genellikle iki kamçı. Diploid veya agamont, çok çekirdekli, ve sonra mayoz yeni gamonts üretmek için bölünür. Birden çok tur eşeysiz üreme bentik formlarda cinsel nesiller arasında nadir değildir.[28]

Foraminifera üreme döngüleri ile ilişkili morfolojik dimorfizm sergiler. Gamont veya cinsel olarak üreyen haploid formu, megalosferik-Yani onun prolokulusveya birinci oda orantılı olarak büyüktür. Gamont aynı zamanda Bir form. Gamontlar, tipik olarak daha büyük proloküllere sahip olmalarına rağmen, genellikle agamontlardan daha küçük genel test çapına sahiptir.
Olgunluğa ulaştıktan sonra gamont, mitoz aynı zamanda haploid olan binlerce gamet üretmek için. Bu gametlerin hepsinde tam bir set var organeller ve testten zarar görmeden ortama atılır. Gametler farklı değildir sperm ve Yumurta ve bir türden herhangi iki gamet genellikle birbirini dölleyebilir.

İki gamet birleştiğinde, diploid, çok çekirdekli bir hücre oluştururlar. agamont, veya B form. Gamont'un aksine agamont, mikrosferikorantılı olarak küçük bir birinci bölme, ancak tipik olarak daha fazla bölme ile daha büyük genel çapa sahip. Agamont, eşeysiz üreme foraminiferin evresi; yetişkinliğe ulaştıktan sonra, protoplazma testi tamamen boşa çıkarır ve testini böler. sitoplazma mayotik olarak üzerinden çoklu fisyon bir dizi haploid yavru oluşturmak için. Bu yavrular daha sonra dağılmadan önce megalosferik ilk odalarını oluşturmaya başlar.
Bazı durumlarda haploid genç, başka bir megalosferik, haploid yavru üretmek için eşeysiz olarak çoğalan megalosferik bir forma olgunlaşabilir. Bu durumda, ilk megalosferik form, Schizont veya Bir1 biçim, ikincisi ise gamont veya Bir2 form.

Daha soğuk ve derin suda olgunlaşma ve üreme daha yavaş gerçekleşir; bu koşullar ayrıca foramların daha da büyümesine neden olur. Bir formlar her zaman olduğundan çok daha fazla görünür B iki gametin birbiriyle karşılaşma ve başarılı bir şekilde birleşme olasılığının azalması nedeniyle formlar.[45][27]
Üreme modunda varyasyonlar
Farklı foraminifer gruplarında üreme stratejilerinde yüksek derecede çeşitlilik vardır.
İçinde tek gözlü türler, Bir form ve B form hala mevcut. Multiloküler foramların mikrosferik morfunda olduğu gibi, eşeysiz üreme B biçim cinsel olarak yeniden üretilenden daha büyüktür Bir form.
Ailedeki foramlar Spirillinidae kamçılı yerine amipli gametlere sahiptir. Bu gruptaki üremenin diğer yönleri genellikle diğer foram gruplarınınkine benzer.
Kireçli spirilinid Patellina corrugata diğer foraminiferlerin çoğundan biraz farklı bir üreme stratejisine sahiptir. Eşeysiz üreme B form tüm hücreyi çevreleyen bir kist üretir; daha sonra bu kist içinde bölünür ve genç hücreler, kendi testlerinin ilk odasını oluşturmak için ebeveynin testinin kalsitini yamyam eder. Bunlar Bir formlar olgunlaştıktan sonra dokuz kişiye kadar gruplar halinde toplanır; daha sonra tüm grup etrafında koruyucu bir kist oluştururlar. Gametogenez çok az sayıda gamet üreterek bu kist içinde oluşur. B larvalar kistin içinde üretilir; Hücrelere bağlı olmayan herhangi bir çekirdek, gelişmekte olan larvalar için besin olarak tüketilir. Patellina içinde Bir form bildirildi ikievcikli cinsiyetler "artı" ve "eksi" olarak anılır; bu cinsiyetler çekirdek sayısı bakımından farklılık gösterir, "artı" formu üç çekirdeğe ve "eksi" formu dört çekirdeğe sahiptir. B form yine daha büyük Bir form.[27][45][39]
Testler

Foraminiferal testler içindeki organizmayı korumaya hizmet eder. Genel olarak sert ve dayanıklı yapıları nedeniyle (diğer protistlere kıyasla), foraminifer testleri grup hakkında önemli bir bilimsel bilgi kaynağıdır.
Testte sitoplazmanın dışarıya uzanmasına izin veren açıklıklara açıklık denir.[46] birincil Dışa açılan açıklık, bunlarla sınırlı olmamak üzere, yuvarlak, hilal şeklinde, yarık şekilli, başlıklı, ışıma (yıldız şeklinde), dendritik (dallanma) dahil olmak üzere farklı türlerde birçok farklı şekil alır. Bazı foraminiferlerde "dişli", flanşlı veya dudaklı birincil açıklıklar bulunur. Yalnızca bir veya birden fazla birincil açıklık olabilir; çoklu mevcut olduğunda, kümelenmiş veya ekvatoral olabilirler. Birincil açıklığa ek olarak, birçok foraminiferde tamamlayıcı açıklıklar. Bunlar, kalıntı açıklıklar (daha önceki bir büyüme aşamasından geçmiş birincil açıklıklar) veya benzersiz yapılar olarak oluşabilir.
Test şekli farklı foraminiferler arasında oldukça değişkendir; tek odacıklı (tek gözlü) veya çok odacıklı (çok gözlü) olabilirler. Multiloküler formlarda, organizma büyüdükçe yeni odalar eklenir. Spiralli, seri ve miliolin dahil olmak üzere hem tek gözlü hem de çok gözlü formlarda çok çeşitli test morfolojileri bulunur.[27]
Birçok foraminifer, megalosferik ve mikrosferik bireylerle testlerinde dimorfizm sergiler. Bu isimler, tam organizmanın boyutuna atıfta bulunarak alınmamalıdır; daha ziyade, ilk odanın boyutuna atıfta bulunurlar veya prolokulus.

Fosil olarak yapılan testler, Ediacaran dönem[47] ve birçok deniz tortusu öncelikle bunlardan oluşur. Örneğin Mısır piramitlerini oluşturan kireçtaşının neredeyse tamamı nummulitik bentik Foraminifera.[48] Resif Foraminifera'nın yılda yaklaşık 43 milyon ton kalsiyum karbonat ürettiği tahmin edilmektedir.[49]
Genetik çalışmalar çıplak amipi tanımladı Retikülomiks ve tuhaf ksenofoforlar foraminiferans olarak testler olmadan. Diğer birkaç amoeboid, retiküloz psödopodlar üretir ve daha önce foramlarla Granuloreticulosa olarak sınıflandırılırdı, ancak bu artık doğal bir grup olarak kabul edilmiyor ve çoğu artık Cercozoa arasına yerleştiriliyor.[50]
Test bileşimi
Onların şekli ve bileşimi testler foramların tanımlanması ve sınıflandırılması için birincil araçlardır. Çoğu kireçli test salgılar. kalsiyum karbonat.[28] Kireçli testler aşağıdakilerden oluşabilir: aragonit veya kalsit türlere bağlı olarak; kalsit testleri olanlar arasında, test yüksek veya düşük oranda magnezyum ikamesi içerebilir.[12] Test, bazen fosil örneklerinden geri kazanılabilen organik bir matris içerir.[12]
Bazı çalışmalar yüksek miktarda homoplazi foraminiferde ve ne aglütine olmuş ne de kalkerli foraminiferler monofiletik gruplar oluşturmaz.[16]
Yumuşak testler
Bazı foramlarda testler organik materyalden, tipik olarak protein tektin. Tektin duvarları, yüzeye gevşek bir şekilde yapışmış tortu parçacıklarına sahip olabilir.[27] Foram Retikülomiks tamamen bir testten yoksundur, sadece membranöz bir hücre duvarına sahiptir.[26] Organik duvarlı foramlar geleneksel olarak "allogromiidler" olarak gruplandırılmıştır; Bununla birlikte, genetik çalışmalar bunun doğal bir grup oluşturmadığını bulmuştur.[16]
Aglütine testler
Diğer foramlar, proteinler (muhtemelen kolajenle ilişkili), kalsiyum karbonat veya kalsiyum karbonatla birbirine yapıştırılmış (aglütine edilmiş) küçük tortu parçalarından yapılmış testlere sahiptir. Demir (III) oksit.[27][51] Geçmişte bu formlar, tek odacıklı "astrorizidler" ve çok odacıklı tekstiliidler olarak bir arada gruplandırıldı. Ancak son genetik araştırmalar, "astrorizidlerin" doğal bir gruplama oluşturmadığını, bunun yerine foram ağacının geniş bir tabanını oluşturduğunu göstermektedir.[16]
Textulariid foraminifera, globothalamea'nın diğer yaşayan üyelerinden farklı olarak, aglütine testlere sahiptir; ancak bu testlerdeki taneler kalsit çimentosu ile çimentolanır. Bu kalsit çimentosu, diğer globotalamalara benzer şekilde küçük (<100 nm) küresel nanogrenlerden oluşur. Bu testler ayrıca birçok gözeneğe sahip olabilir, bu da onları globothalamea ile birleştiren başka bir özelliktir.[36]

Aglutinating foraminifera, kabuklarına hangi parçacıkları dahil ettikleri konusunda seçici olabilir. Bazı türler belirli boyutlarda ve türlerde kaya parçacıkları tercih eder; diğer türler belirli biyolojik materyallere tercih edilir. Bazı foraminifer türlerinin tercihli olarak aglütine olduğu bilinmektedir. kokolitler testlerini oluşturmak için; diğerleri tercihen kullanır ekinoderm tabaklar, diyatomlar, hatta diğer foraminiferans 'testleri.[52]
Foraminifer Spiculosiphon tercihen silika yapışır sünger dikenler organik bir çimento kullanarak; "sapı" üzerinde uzun sivri uçları ve "ampulü" üzerinde kısaltılmış sivri uçları kullanarak, şekle karşı da güçlü bir seçicilik gösterir. İğneleri hem deniz tabanından yukarı kaldırmanın hem de deniz tabanının erişimini uzatmanın bir yolu olarak kullandığı düşünülmektedir. psödopodi avı yakalamak için.[51]
Aglütine testleri ksenofoforlar herhangi bir foraminiferin en büyüğüdür ve çapı 20 cm'ye kadar ulaşır. "Yabancı cisimlerin taşıyıcısı" anlamına gelen "ksenofiyofor" adı, bu aglütinasyon alışkanlığını ifade eder. Xenophyophores, daha büyük çakıllardan ve daha ince siltlerden kaçınarak 63 ve 500 um arasında tortu tanelerini seçici olarak alır; Partikül tipi tercihen sülfidleri, oksitleri, volkanik camı ve özellikle daha küçük foraminifer testlerini içerdiğinden, tortu tipi, partiküllerin aglütine olduğu güçlü bir faktör gibi görünmektedir. 1.5 cm çapındaki Xenophyophores hiçbir test yapılmadan tamamen çıplak olarak kaydedilmiştir.[53]

Kalkerli testler
Kalkerli testlere sahip foraminiferlerden birkaç farklı kalsit kristal yapısı bulunur.
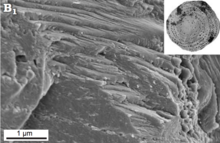
Porselen duvarlar, Miliolida. Bunlar, sıralı bir dış ve iç kalsit astarıyla organize edilmiş yüksek magnezyumlu kalsitten oluşur ("extrados" ve "Intrados", sırasıyla) ve kalın bir merkez tabaka (" porselen ") oluşturan rastgele yönlendirilmiş iğne şeklindeki kalsit kristalleri. Organik bir iç kaplama da mevcuttur. Dış yüzey çukurlu bir yapıya sahip olabilir, ancak deliklerle delinmemiş." Cornuspirid "miliolidler görünüşe göre herhangi bir ekstradan yoksundur.[54][55][36]"Monokristal" bir test yapısı, geleneksel olarak, Spirillinida. Bununla birlikte, bu testler hala tam olarak anlaşılmamış ve yeterince tanımlanmamıştır. Bazı "monokristalli" spirilinidlerin, taramalı elektron mikroskobu ile gözlemlendiğinde, aslında çok küçük kristallerden oluşan bir mozaikten oluşan testlere sahip oldukları bulunmuştur. SEM gözlemi Patellina sp. gerçekten monokristalli bir testin, belirgin bölünme yüzleri ile gerçekten mevcut olabileceğini önermektedir.[36]

Lagenid testler onlarca mikrometre uzunluğa ulaşabilen "elyaf demetleri" nden oluşur; her "demet" tek bir kalsit kristalinden oluşur, enine kesiti üçgen şeklindedir ve merkezinde bir gözenek vardır (test biriktirme eseri olduğu düşünülmektedir). Elyaf demetlerinin "koni" yapısına bağlı bir dahili organik katman da vardır. Kristalin yapı diğer kalkerli foraminiferlerden önemli ölçüde farklılık gösterdiğinden, kalkerli testin ayrı bir evrimini temsil ettiği düşünülmektedir. Göz kapaklarının tam mineralizasyon süreci belirsizliğini koruyor.[55]

Rotaliid testler "hiyalin" olarak tanımlanır. Düşük ila yüksek magnezyum kalsit "nanogren" lerinden oluşurlar. C eksenleri testin dış yüzeyine dik. Ayrıca bu nanograinler, satırlar, sütunlar veya demetler gibi daha yüksek seviyeli yapılara sahip olabilir.[36] Test duvarı karakteristik olarak çift katlıdır (iki katmanlı) ve küçük gözeneklerle baştan sona deliklidir. Test duvarının dış kalsit tabakası "dış tabaka" olarak adlandırılırken, iç kalsit tabakası "iç kaplama" olarak adlandırılır; bu, testin altındaki organik iç astar ile karıştırılmamalıdır. Dış tabaka ve iç astar arasına, ikisini ayıran bir protein tabakası olan "medyan tabaka" sıkıştırılmıştır. Orta katman oldukça değişkendir; türlere bağlı olarak iyi tanımlanabilirken diğerlerinde keskin bir şekilde tasvir edilmemiştir. Bazı cinsler, orta katman içinde tortu parçacıkları içerebilir.[27][56][55]

Carterinids cins dahil Carterina ve Zaninettia, sınıflandırmalarını uzun süre karmaşık hale getiren benzersiz bir kristal yapı testine sahiptir. Bu cinsteki test, organik bir matris ile birbirine bağlanmış ve organik madde "kabarcıklarını" içeren düşük magnezyumlu kalsit uçlarından oluşur; bu, bazı araştırmacıların testin birleştirilmesi gerektiği sonucuna varmasına neden oldu. Bununla birlikte, yaşam çalışmaları, aglütinasyonu bulmada başarısız oldu ve aslında cins, tortu parçacıklarının birikmediği yapay substrat üzerinde keşfedildi.[57] 2014'te yapılan bir genetik çalışma, karterinidlerin Globothalamea içinde bağımsız bir soy olduğunu buldu ve spikül şekli, numuneler arasında tutarlı bir şekilde farklılık gösterdiğinden, spiküllerin salgılanması fikrini destekledi. Carterina ve Zaninettia aynı bölgeden toplandı (oval Carterina, yuvarlatılmış dikdörtgen Zaninettia).[58]
Şimdi soyu tükenmiş Fusulinidler geleneksel olarak homojen mikrogranüler kristallerin test edilmesinde, tercih edilen yönelim olmadan ve hemen hemen hiç çimento içermeyen benzersiz kabul edilmiştir. Bununla birlikte, 2017 yılında yapılan bir araştırma, mikrogranüler yapının aslında fosillerin diyajenetik değişiminin sonucu olduğunu ve değiştirilmemiş fusulinid testlerinin bunun yerine hiyalin yapısına sahip olduğunu buldu. Bu, grubun, Globothalamea.[59]
Robertinids Sahip olmak aragonitik delikli testler; bunlar, nanograinlerden oluştukları için rotaliidlerin testlerine benzerler, ancak, kompozisyon ve iyi organize edilmiş sütunlu alanlara sahip olmaları bakımından farklılık gösterirler. İlk planktonik foramlar aragonitik testlere sahip olduğundan, bunun Robertinida'daki planktonik yaşam tarzının ayrı bir evrimini temsil edebileceği öne sürülmüştür. Globigerinans.[36]
Hyalin aragonitik testleri de mevcuttur. Involutinida.[55]
Silika testleri
Bir cins, Miliamellus opalinden yapılmış deliksiz bir testi vardır silika.[18] Tipik miliolidlerin testlerine benzer şekil ve yapıdadır; test, bir dahili ve harici organik katmanın yanı sıra uzun çubuklardan oluşan bir orta silika katmanından oluşur. Bu silika tabakası ayrıca dış, orta ve iç alt birimlere bölünmüştür; dış ve iç alt birimlerin her biri yaklaşık 0.2 um kalınlığındadır ve uzun eksenleri test yüzeyine paralel olan alt paralel silika çubuk tabakalarından oluşur. Orta alt birim yaklaşık 18 um kalınlığındadır ve açık alanda organik bileşen içermeyen üç boyutlu bir silika çubuklar kafesinden oluşur. İnce yapı, miliolidlerden farklıdır, çubuklar ortalama olarak iki kat daha uzun ve ortalama olarak iki kat daha kalın olmalıdır. Miliamellus are hollow rather than solid, and of course in having a silica test rather than calcite.[60]

Test wall construction
When a secreted test is present, walls of foraminiferal tests may be either nonlamellar veya katmanlı.
Nonlamellar walls are found in some foraminifera, such as Carterinida, Spirillinida, ve Miliolida. In these forms, the secretion of a new chamber is not associated with any further deposition over previous chambers. As such there is no associated layering of calcite layers on the test.[56]
In foraminifera with lamellar walls, the deposition of a new chamber is accompanied by the deposition of a layer over previously-formed chambers. This layer may cover all previous chambers, or it may cover only some of them. These layers are known as secondary lamellae.
Foraminifera with lamellar walls can be further broken down into those with monolamellar walls and those with bilamellar duvarlar. Monolamellar foraminifera secrete test walls which consist of a single layer, while those of bilamellar foraminifera are double-layered with an organic "median layer", sometimes containing sediment particles. In the case of bilamellar foraminifera, the outer layer is referred to as the "outer lamella" whilst the inner layer is referred to as the "inner lining". Monolamellar forams include the Lagenida, while bilamellar forms include the Rotaliida (including the major planktonic subgroup, the Globigerinina ).[56]
Bilamellar test walls can be further divided into those with septal flaps (a layer of test wall covering the previously-secreted septum) and those lacking septal flaps. Septal flaps are not known to be present in any foraminifera other than those with bilamellar walls.
The presence of a septal flap is often, though not always, associated with the presence of an interlocular space. As the name suggests, this is a small space located between chambers; it may be open and form part of the outer surface of the test, or it may be enclosed to form a void. The layer enclosing the void is formed from different parts of the lamellae in different genera, suggesting an independent evolution of enclosed interlocular spaces in order to strengthen the test.[56]
Evrimsel tarih
Molecular clocks indicate that the crown-group of foraminifera likely evolved during the Neoproterozoik, between 900 and 650 million years ago; this timing is consistent with Neoproterozoic fossils of the closely related filose amoebae. As fossils of foraminifera have not been found prior to the very end of the Ediacaran, it is likely that most of these Proterozoic forms did not have hard-shelled tests.[61][62]
Due to their non-mineralised tests, "allogromiids " have no fossil record.[61]

Gizemli vendozoans of the Ediacaran period have been suggested to represent fossil xenophyophores.[63] Ancak, keşfi diagenetically-altered C27 steroller associated with the remains of Dickinsonia cast doubt on this identification and suggest it may instead be an animal.[64] Other researchers have suggested that the elusive trace fossil Paleodictyon ve Onun akraba may represent a fossil xenophyophore[65] and noted the similarity of the extant xenophyophore Occultammina to the fossil;[66] however, modern examples of Paleodictyon have not been able to clear up the issue and the trace may alternately represent a burrow or a glass sponge.[67] Supporting this notion is the similar habitat of living xenophyophores to the inferred habitat of fossil graphoglyptids; however, the large size and regularity of many graphoglyptids as well as the apparent absence of xenophyae in their fossils casts doubt on the possibility.[66] As of 2017 no definite xenophyophore fossils have been found.[68]
Test-bearing foraminifera have an excellent fossil record throughout the Fanerozoik eon. The earliest known definite foraminifera appear in the fossil record towards the very end of the Ediacaran; these forms all have agglutinated tests and are unilocular. These include forms like Platysolenites ve Spirosolenites.[69][47]
Single-chambered foraminifera continued to diversity throughout the Cambrian. Some commonly encountered forms include Ammodiscus, Glomospira, Psammosphera, ve Turritellella; these species are all agglutinated. They make up part of the Ammodiscina, bir soy spirillinids that still contains modern forms.[70][16] Later spirillinids would evolve multilocularity and calcitic tests, with the first such forms appearing during the Triyas; the group saw little effects on diversity due to the K-Pg extinction.[71]
The earliest multi-chambered foraminifera are agglutinated species, and appear in the fossil record during the middle Kambriyen dönem. Due to their poor preservation they cannot be positively assigned to any major foram group.[70]

The earliest known calcareous-walled foraminifera are the Fusulinids, which appear in the fossil record during the Llandoverian epoch of the early Silüriyen. The earliest of these were microscopic, planispirally coiled, and evolute; later forms evolved a diversity of shapes including lenticular, globular, and perhaps most famously, elongated rice-shaped forms. Later species of fusulinids grew to much larger size, with some forms reaching 5 cm in length; reportedly, some specimens reach up to 14 cm in length, making them among the largest foraminifera extant or extinct. Fusulinids are the earliest lineage of foraminifera thought to have evolved symbiosis with photosynthetic organisms. Fossils of fusulinids have been found on all continents except Antarktika; they reached their greatest diversity during the Visean çağ Karbonifer. The group then gradually declined in diversity until finally going extinct during the Permo-Triyas yok oluş olayı.[27][71][72]
Esnasında Turnaisiyen epoch of the Carboniferous, Miliolid foraminifera first appeared in the fossil record, having diverged from the spirillinids içinde Tubothalamea. Miliolids suffered about 50% casualties during both the Permo-Triassic and K-Pg extinctions but survived to the present day. Some fossil miliolids reached up to 2 cm in diameter.[71]

Bilinen en eski Lagenid fossils appear during the Moscovian epoch of the Carboniferous. Seeing little effect due to the Permo-Triassic or K-Pg extinctions, the group diversified through time. Secondarily unilocular taxa evolved during the Jurassic and Cretaceous.
En erken Involutinid fossils appear during the Permian; the lineage diversified throughout the Mesozoic of Eurasia before apparently vanishing from the fossil record following the Cenomanian-Turonian Ocean Anoxic Event. The extant group planispirillinidae has been referred to the involutinida, but this remains the subject of debate.[73][71]
Robertinida first appear in the fossil record during the Anisiyen epoch of the Triassic. The group remained at low diversity throughout its fossil history; all living representatives belong to the Robertinidae, which first appeared during the Paleosen.[71]
The first definite Rotaliid fossils do not appear in the fossil record until the Pliensbakiyen epoch of the Jurassic, following the Triassic-Jurassic event.[74] Diversity of the group remained low until the aftermath of the Cenomanian-Turonian event, after which the group saw a rapid diversification. Of this group, the planktonic Globigerinina —the first known group of planktonic forams—first appears in the aftermath of the Toarcian Turnover; the group saw heavy losses during both the K-Pg extinction and the Eocene-Oligocene extinction, but remains extant and diverse to this day.[71] An additional evolution of planktonic lifestyle occurred in the Miocene or Pliocene, when the rotaliid Neogallitellia independently evolved a planktonic lifestyle.[34][35]
Paleontological applications


Dying planktonic Foraminifera continuously rain down on the sea floor in vast numbers, their mineralized tests preserved as fossils in the accumulating tortu. Beginning in the 1960s, and largely under the auspices of the Deep Sea Drilling, Ocean Drilling, and International Ocean Drilling Programmes, as well as for the purposes of oil exploration, advanced deep-sea drilling techniques have been bringing up sediment cores bearing Foraminifera fossils.[75] The effectively unlimited supply of these fossil tests and the relatively high-precision age-control models available for cores has produced an exceptionally high-quality planktonic Foraminifera fossil record dating back to the mid-Jurassic, and presents an unparalleled record for scientists testing and documenting the evolutionary process.[75] The exceptional quality of the fossil record has allowed an impressively detailed picture of species inter-relationships to be developed on the basis of fossils, in many cases subsequently validated independently through molecular genetic studies on extant specimens[76]
Because certain types of foraminifera are found only in certain environments, their fossils can be used to figure out the kind of environment under which ancient marine sediments were deposited; conditions such as salinity, depth, oxygenic conditions, and light conditions can be determined from the different habitat preferences of various species of forams. This allows workers to track changing climates and environmental conditions over time by aggregating information about the foraminifera present.[77]
In other cases, the relative proportion of planktonic to benthic foraminifera fossils found in a rock can be used as a proxy for the depth of a given locality when the rocks were being deposited.[78]
Foraminifera have significant application in the field of biyostratigrafi. Due to their small size and hard shells, foraminifera may be preserved in great abundance and with high quality of preservation; due to their complex morphology, individual species are easily recognizable. Foraminifera species in the fossil record have limited ranges between the species' first evolution and their disappearance; stratigraphers have worked out the successive changes in foram assemblages throughout much of the Fanerozoik. As such, the assemblage of foraminifera within a given locality can be analyzed and compared to known dates of appearance and disappearance in order to narrow down the age of the rocks. This allows paleontologists to interpret the age of tortul kayaçlar ne zaman radyometrik tarihleme uygulanamaz.[79] This application of foraminifera was discovered by Alva C. Ellisor 1920'de.[80]
Calcareous fossil foraminifera are formed from elements found in the ancient seas where they lived. Thus, they are very useful in paleoklimatoloji ve paleookşinografi. They can be used, as a climate vekil, to reconstruct past climate by examining the kararlı izotop ratios and trace element content of the shells (tests). Global temperature and ice volume can be revealed by the isotopes of oxygen, and the history of the karbon döngüsü and oceanic productivity by examining the stable isotope ratios of carbon;[81] görmek δ18O ve δ13C. The concentration of trace elements, like magnezyum (Mg),[82] lityum (Li)[83] ve bor (B),[84] also hold a wealth of information about global temperature cycles, continental weathering, and the role of the ocean in the global carbon cycle. Geographic patterns seen in the fossil records of planktonic forams are also used to reconstruct ancient okyanus akıntıları.
Modern kullanımlar
petrol endüstrisi relies heavily on mikrofosiller such as forams to find potential hydrocarbon deposits.[85]

For the same reasons they make useful biostratigraphic markers, living foraminiferal assemblages have been used as biyoindikatörler in coastal environments, including indicators of coral reef health. Because calcium carbonate is susceptible to dissolution in acidic conditions, foraminifera may be particularly affected by changing climate and okyanus asitlenmesi.

Foraminifera have many uses in petrol arama and are used routinely to interpret the ages and paleoenvironments of sedimentary strata in oil wells.[86] Agglutinated fossil foraminifera buried deeply in sedimentary basins can be used to estimate thermal maturity, which is a key factor for petroleum generation. Foraminiferal Colouration Index[87] (FCI) is used to quantify colour changes and estimate burial temperature. FCI data is particularly useful in the early stages of petroleum generation (about 100 °C).
Foraminifera can also be used in arkeoloji içinde provenancing of some stone raw material types. Some stone types, such as kireçtaşı, are commonly found to contain fossilised foraminifera. The types and concentrations of these fossils within a sample of stone can be used to match that sample to a source known to contain the same "fossil signature".[88]
Fotoğraf Galerisi

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Pag Island, Adriatic Sea -60 m, field width 5.5 mm

Foraminifera of Indian Ocean, south-eastern coast of Bali, field width 5.5 mm

Foraminifera of Indian Ocean, south-eastern coast of Bali, field width 5.5 mm

Foraminifera of Indian Ocean, south-eastern coast of Bali, field width 5.5 mm

Foraminifera in Ngapali, Myanmar, field width 5.22 mm

Foraminifer Heterostegina depressa, field width 4.4 mm









Referanslar
- ^ Parfrey, Laura Wegener; Lahr, Daniel J. G.; Knoll, Andrew H.; Katz, Laura A. (16 August 2011). "Estimating the timing of early eukaryotic diversification with multigene molecular clocks". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 108 (33): 13624–13629. Bibcode:2011PNAS..10813624P. doi:10.1073/pnas.1110633108. PMC 3158185. PMID 21810989.
- ^ Giere, Olav (2009). Meiobenthology: the microscopic motile fauna of aquatic sediments (2. baskı). Berlin: Springer. ISBN 978-3540686576.
- ^ a b Lejzerowicz, Franck; Pawlowski, Jan; Fraissinet-Tachet, Laurence; Marmeisse, Roland (1 September 2010). "Molecular evidence for widespread occurrence of Foraminifera in soils". Çevresel Mikrobiyoloji. 12 (9): 2518–26. doi:10.1111/j.1462-2920.2010.02225.x. PMID 20406290.
- ^ Kennett, J.P.; Srinivasan, M.S. (1983). Neogene planktonic foraminifera: a phylogenetic atlas. Hutchinson Ross. ISBN 978-0-87933-070-5.
- ^ Ald, S.M. et al. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ^ Pawlowski, J., Lejzerowicz, F., & Esling, P. (2014). Next-generation environmental diversity surveys of foraminifera: preparing the future. The Biological Bulletin, 227(2), 93-106.
- ^ "World Foraminifera Database".
- ^ Marshall M (3 February 2010). "Zoologger: 'Living beach ball' is giant single cell". Yeni Bilim Adamı.
- ^ a b Lipps JH, Finger KL, Walker SE (October 2011). "What Should We call the Foraminifera" (PDF). Journal of Foraminiferal Research. 41 (4): 309–313. doi:10.2113/gsjfr.41.4.309. Alındı 10 Nisan 2018.
- ^ "Foraminifera | Fossil Focus | Time | Discovering Geology | British Geological Survey (BGS)". www.bgs.ac.uk. Alındı 20 Temmuz 2020.
- ^ "Micrographia, or, Some physiological descriptions of minute bodies made by magnifying glasses ?with observations and inquiries thereupon /by R. Hooke ... : Hooke, Robert, : Free Download, Borrow, and Streaming". İnternet Arşivi. Alındı 20 Temmuz 2020.
- ^ a b c d e Sen Gupta, Barun K. (2003), Sen Gupta, Barun K. (ed.), Modern Foraminifera, Springer Netherlands, pp. 7–36, doi:10.1007/0-306-48104-9_2, ISBN 978-0-306-48104-8 Eksik veya boş
| title =(Yardım);| bölüm =yok sayıldı (Yardım) - ^ a b BOUDAGHER-FADEL, MARCELLE K. (2018), "Biology and Evolutionary History of Larger Benthic Foraminifera", Evolution and Geological Significance of Larger Benthic Foraminifera (2 ed.), UCL Press, pp. 1–44, ISBN 978-1-911576-94-5, JSTOR j.ctvqhsq3.3
- ^ Hansen, H. (1 January 1981). "On Lorentz Spengler and a neotype for the foraminifer Calcarina spengleri". Alıntı dergisi gerektirir
| günlük =(Yardım) - ^ d'Orbigny, Alcide (1826). "Tableau Méthodique de la Classe des Céphalopodes". Annales des Sciences Naturelles, Paris (Série 1). 7: 245–314 – via Biodiversity Heritage Library.
- ^ a b c d e f g Pawlowski, Jan; Holzmann, Maria; Tyszka, Jarosław (1 April 2013). "New supraordinal classification of Foraminifera: Molecules meet morphology". Deniz Mikropaleontolojisi. 100: 1–10. Bibcode:2013MarMP.100....1P. doi:10.1016/j.marmicro.2013.04.002. ISSN 0377-8398.
- ^ a b Loeblich Jr, A.R.; Tappan, H. (1964). "Foraminiferida". Part C, Protista 2. Treatise on Invertebrate Paleontology. Amerika Jeoloji Derneği. pp. C55–C786. ISBN 978-0-8137-3003-5.
- ^ a b c d e f g h Sen Gupta, Barun K. (2002). Modern Foraminifera. Springer. s. 16. ISBN 978-1-4020-0598-5.
- ^ a b c Cavalier-Smith, T (2004). "Only Six Kingdoms of Life" (PDF). Bildiriler. Biyolojik Bilimler. 271 (1545): 1251–62. doi:10.1098 / rspb.2004.2705. PMC 1691724. PMID 15306349.
- ^ a b Cavalier-Smith, T (2003). "Protist phylogeny and the high-level classification of Protozoa". Avrupa Protistoloji Dergisi. 34 (4): 338–348. doi:10.1078/0932-4739-00002.
- ^ Tolweb Cercozoa
- ^ European Register of Marine Species
- ^ eForams-taxonomy Arşivlendi 3 Ekim 2011 Wayback Makinesi
- ^ Testate amoebae as environmental indicators (PDF), dan arşivlendi orijinal (PDF) 27 Kasım 2016'da, alındı 27 Kasım 2016
- ^ Mikhalevich, V.I. (2013). "New insight into the systematics and evolution of the foraminifera". Mikropaleontoloji. 59 (6): 493–527.
- ^ a b Pawlowski, Jan; Bolivar, Ignacio; Fahrni, Jose F.; Vargas, Colomban De; Bowser, Samuel S. (1999). "Molecular Evidence That Reticulomyxa Filosa Is A Freshwater Naked Foraminifer". Ökaryotik Mikrobiyoloji Dergisi. 46 (6): 612–617. doi:10.1111/j.1550-7408.1999.tb05137.x. ISSN 1550-7408. PMID 10568034. S2CID 36497475.
- ^ a b c d e f g h ben Saraswati, Pratul Kumar; Srinivasan, M. S. (2016), Saraswati, Pratul Kumar; Srinivasan, M.S. (eds.), "Calcareous-Walled Microfossils", Micropaleontology: Principles and Applications, Springer International Publishing, pp. 81–119, doi:10.1007/978-3-319-14574-7_6, ISBN 978-3-319-14574-7
- ^ a b c d Sen Gupta, Barun K. (1982). "Ecology of benthic Foraminifera". In Broadhead, T.W. (ed.). Foraminifera: notes for a short course organized by M.A. Buzas and B.K. Sen Gupta. Studies in Geology. 6. University of Tennessee, Dept. of Geological Sciences. pp. 37–50. ISBN 978-0910249058. OCLC 9276403.
- ^ a b c Hemleben, C .; Anderson, O.R .; Spindler, M. (1989). Modern Planktonik Foraminifer. Springer-Verlag. ISBN 978-3-540-96815-3.
- ^ Grell, K. G. (1 January 1979). "Cytogenetic systems and evolution in foraminifera". The Journal of Foraminiferal Research. 9 (1): 1–13. doi:10.2113/gsjfr.9.1.1. ISSN 0096-1191.
- ^ a b c Lekieffre, Charlotte; Bernhard, Joan M.; Mabilleau, Guillaume; Filipsson, Helena L.; Meibom, Anders; Geslin, Emmanuelle (1 January 2018). "An overview of cellular ultrastructure in benthic foraminifera: New observations of rotalid species in the context of existing literature". Deniz Mikropaleontolojisi. 138: 12–32. doi:10.1016/j.marmicro.2017.10.005. ISSN 0377-8398.
- ^ Domanov, M. M. (July 2015). "Natural 226Ra and 232Th radionuclides in xenophyophores of the Pacific Ocean". Jeokimya Uluslararası. 53 (7): 664–669. doi:10.1134/S0016702915070034. ISSN 0016-7029. S2CID 127121951.
- ^ Kucera, M.; Darling, K.F. (Nisan 2002). "Cryptic species of planktonic foraminifera: their effect on palaeoceanographic reconstructions". Philos Trans Royal Soc A. 360 (1793): 695–718. Bibcode:2002RSPTA.360..695K. doi:10.1098/rsta.2001.0962. PMID 12804300. S2CID 21279683.
- ^ a b Ujiié, Yurika; Kimoto, Katsunori; Pawlowski, Jan (December 2008). "Molecular evidence for an independent origin of modern triserial planktonic foraminifera from benthic ancestors". Deniz Mikropaleontolojisi. 69 (3–4): 334–340. doi:10.1016/j.marmicro.2008.09.003.
- ^ a b Özdikmen, Hüseyin (June 2009). "Substitute names for some unicellular animal taxa (Protozoa" (PDF). Munis Entomology & Zoology. 4 (1): 233–256.
- ^ a b c d e f Dubicka, Zofia (2019). "Chamber arrangement versus wall structure in the high-rank phylogenetic classification of Foraminifera". Acta Palaeontologica Polonica. 64. doi:10.4202/app.00564.2018. ISSN 0567-7920.
- ^ Mikrobiyal Ekolojideki Gelişmeler, Cilt 11
- ^ Bernhard, J. M .; Bowser, S.M. (1999). "Dysoxic sedimentlerin Bentik Foraminiferleri: kloroplast sekestrasyonu ve fonksiyonel morfoloji". Yer Bilimi Yorumları. 46 (1): 149–165. Bibcode:1999 ESRv ... 46..149B. doi:10.1016 / S0012-8252 (99) 00017-3.
- ^ a b Goldstein, Susan T. (2003), Sen Gupta, Barun K. (ed.), Modern Foraminifera, Springer Netherlands, pp. 37–55, doi:10.1007/0-306-48104-9_3, ISBN 978-0-306-48104-8 Eksik veya boş
| title =(Yardım);| bölüm =yok sayıldı (Yardım) - ^ Laureillard, J; Méjanelle, L; Sibuet, M (2004). "Use of lipids to study the trophic ecology of deep-sea xenophyophores". Deniz Ekolojisi İlerleme Serisi. 270: 129–140. doi:10.3354/meps270129. ISSN 0171-8630.
- ^ Culver, Stephen J .; Lipps, Jere H. (2003), Kelley, Patricia H.; Kowalewski, Michał; Hansen, Thor A. (eds.), "Predation on and by Foraminifera", Predator—Prey Interactions in the Fossil Record, Boston, MA: Springer US, pp. 7–32, doi:10.1007/978-1-4615-0161-9_2, ISBN 978-1-4613-4947-1, alındı 30 Eylül 2020
- ^ Moodley, L.; Hess, C. (1 August 1992). "Tolerance of Infaunal Benthic Foraminifera for Low and High Oxygen Concentrations". Biyolojik Bülten. 183 (1): 94–98. doi:10.2307/1542410. ISSN 0006-3185. JSTOR 1542410. PMID 29304574.
- ^ Gooday, A.J.; Todo, Y.; Uematsu, K.; Kitazato, H. (July 2008). "New organic-walled Foraminifera (Protista) from the ocean's deepest point, the Challenger Deep (western Pacific Ocean)". Linnean Society'nin Zooloji Dergisi. 153 (3): 399–423. doi:10.1111/j.1096-3642.2008.00393.x.
- ^ Moore, R.C.; Lalicker, A.G.; Fischer, C.G. (1952). "Ch 2 Foraminifera and Radiolaria". Invertebrate Fossils. McGraw-Hill. OCLC 547380.
- ^ a b Haynes, J. R. (18 June 1981). Foraminifer. Springer. ISBN 978-1-349-05397-1.
- ^ Lana, C (2001). "Cretaceous Carterina (Foraminifera)". Deniz Mikropaleontolojisi. 41 (1–2): 97–102. Bibcode:2001MarMP..41...97L. doi:10.1016/S0377-8398(00)00050-5.
- ^ a b Kontorovich, A. E.; Varlamov, A. I.; Grazhdankin, D. V.; Karlova, G. A.; Klets, A. G .; Kontorovich, V. A.; Saraev, S. V.; Terleev, A. A.; Belyaev, S. Yu.; Varaksina, I. V.; Efimov, A. S. (1 December 2008). "A section of Vendian in the east of West Siberian Plate (based on data from the Borehole Vostok 3)". Rus Jeolojisi ve Jeofiziği. 49 (12): 932–939. Bibcode:2008RuGG...49..932K. doi:10.1016/j.rgg.2008.06.012. ISSN 1068-7971.
- ^ Foraminifera: Çalışma Tarihi, University College London, retrieved 20 September 2007
- ^ Langer, M. R.; Silk, M. T. B.; Lipps, J. H. (1997). "Global ocean carbonate and carbon dioxide production: The role of reef Foraminifera". Journal of Foraminiferal Research. 27 (4): 271–277. doi:10.2113/gsjfr.27.4.271.
- ^ Adl, S. M.; Simpson, A. G. B.; Farmer, M. A.; Anderson; et al. (2005). "The new higher level classification of Eukaryotes with emphasis on the taxonomy of Protists". Ökaryotik Mikrobiyoloji Dergisi. 52 (5): 399–451. doi:10.1111 / j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- ^ a b Maldonado, Manuel; López-Acosta, María; Sitjà, Cèlia; Aguilar, Ricardo; García, Silvia; Vacelet, Jean (10 June 2013). "A giant foraminifer that converges to the feeding strategy of carnivorous sponges: Spiculosiphon oceana sp. nov. (Foraminifera, Astrorhizida)". Zootaxa. 3669 (4): 571–584. doi:10.11646/zootaxa.3669.4.9. hdl:10261/92975. ISSN 1175-5334. PMID 26312358.
- ^ Thomsen, Erik; Rasmussen, Tine L. (1 July 2008). "Coccolith-Agglutinating Foraminifera from the Early Cretaceous and How They Constructed Their Tests". Journal of Foraminiferal Research. 38 (3): 193–214. doi:10.2113/gsjfr.38.3.193. ISSN 0096-1191.
- ^ Levin, Lisa A.; Thomas, Cynthia L. (1 December 1988). "The ecology of xenophyophores (Protista) on eastern Pacific seamounts". Derin Deniz Araştırmaları Bölüm A. Oşinografik Araştırma Makaleleri. 35 (12): 2003–2027. Bibcode:1988DSRA...35.2003L. doi:10.1016/0198-0149(88)90122-7. ISSN 0198-0149.
- ^ Jain, Sreepat (2020), Jain, Sreepat (ed.), "Benthic Foraminifera", Fundamentals of Invertebrate Palaeontology: Microfossils, Springer Geology, New Delhi: Springer India, pp. 171–192, doi:10.1007/978-81-322-3962-8_9, ISBN 978-81-322-3962-8
- ^ a b c d Dubicka, Zofia; Owocki, Krzysztof; Gloc, Michał (1 April 2018). "Micro- and Nanostructures of Calcareous Foraminiferal Tests: Insight from Representatives of Miliolida, Rotaliida and Lagenida". Journal of Foraminiferal Research. 48 (2): 142–155. doi:10.2113/gsjfr.48.2.142. ISSN 0096-1191.
- ^ a b c d Hansen, Hans Jørgen (2003), Sen Gupta, Barun K. (ed.), "Shell construction in modern calcareous Foraminifera", Modern Foraminifera, Dordrecht: Springer Netherlands, pp. 57–70, doi:10.1007/0-306-48104-9_4, ISBN 978-0-306-48104-8
- ^ Machado, Altair; Barros, Facelucia (8 January 2013). "The occurrence of Carterina spiculotesta (Carter, 1877) on an artificial substrate". Kontrol Listesi. 9 (4): 813–814. doi:10.15560/9.4.813. ISSN 1809-127X.
- ^ Pawlowski, Jan; Holzmann, Maria; Debenay, Jean-Pierre (1 October 2014). "Molecular Phylogeny of Carterina Spiculotesta and Related Species from New Caledonia". Journal of Foraminiferal Research. 44 (4): 440–450. doi:10.2113/gsjfr.44.4.440. ISSN 0096-1191.
- ^ Dubicka, Zofia; Gorzelak, Przemysław (9 November 2017). "Unlocking the biomineralization style and affinity of Paleozoic fusulinid foraminifera". Bilimsel Raporlar. 7 (1): 15218. Bibcode:2017NatSR...715218D. doi:10.1038/s41598-017-15666-1. ISSN 2045-2322. PMC 5680253. PMID 29123221.
- ^ Resig, J; Lowenstam, H; Echols, R; Weiner, S (1980). "An extant opaline foraminifer: test ultrastructure, mineralogy, and taxonomy". Special Publications of the Cushman Foundation for Foraminiferal Research. 19: 205–214.
- ^ a b Pawlowski, Jan; Holzmann, Maria; Berney, Cédric; Fahrni, José; Gooday, Andrew J.; Cedhagen, Tomas; Habura, Andrea; Bowser, Samuel S. (30 September 2003). "The evolution of early Foraminifera". Ulusal Bilimler Akademisi Bildiriler Kitabı. 100 (20): 11494–11498. Bibcode:2003PNAS..10011494P. doi:10.1073/pnas.2035132100. ISSN 0027-8424. PMC 208786. PMID 14504394.
- ^ Groussin, Mathieu; Pawlowski, Jan; Yang, Ziheng (1 October 2011). "Bayesian relaxed clock estimation of divergence times in foraminifera". Moleküler Filogenetik ve Evrim. 61 (1): 157–166. doi:10.1016/j.ympev.2011.06.008. ISSN 1055-7903. PMID 21723398.
- ^ Seilacher, A. (1 January 2007). "The nature of vendobionts". Jeoloji Topluluğu, Londra, Özel Yayınlar. 286 (1): 387–397. Bibcode:2007GSLSP.286..387S. doi:10.1144/SP286.28. ISSN 0305-8719. S2CID 128619251.
- ^ Bobrovskiy, Ilya; Hope, Janet M.; Ivantsov, Andrey; Nettersheim, Benjamin J.; Hallmann, Christian; Brocks, Jochen J. (21 September 2018). "Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals". Bilim. 361 (6408): 1246–1249. Bibcode:2018Sci...361.1246B. doi:10.1126/science.aat7228. ISSN 0036-8075. PMID 30237355.
- ^ Swinbanks, D. D. (1 October 1982). "Piaeodicton: The Traces of Infaunal Xenophyophores?". Bilim. 218 (4567): 47–49. Bibcode:1982Sci...218...47S. doi:10.1126/science.218.4567.47. ISSN 0036-8075. PMID 17776707. S2CID 28690086.
- ^ a b Levin, Lisa A. (1994). "Paleoecology and Ecology of Xenophyophores". PALAIOS. 9 (1): 32–41. Bibcode:1994Palai...9...32L. doi:10.2307/3515076. ISSN 0883-1351. JSTOR 3515076.
- ^ Rona, Peter A.; Seilacher, Adolf; de Vargas, Colomban; Gooday, Andrew J.; Bernhard, Joan M.; Bowser, Sam; Vetriani, Costantino; Wirsen, Carl O.; Mullineaux, Lauren; Sherrell, Robert; Frederick Grassle, J. (1 September 2009). "Paleodictyon nodosum: A living fossil on the deep-sea floor". Derin Deniz Araştırmaları Bölüm II: Oşinografide Güncel Çalışmalar. Marine Benthic Ecology and Biodiversity: A Compilation of Recent Advances in Honor of J. Frederick Grassle. 56 (19): 1700–1712. Bibcode:2009DSRII..56.1700R. doi:10.1016/j.dsr2.2009.05.015. ISSN 0967-0645.
- ^ Gooday, Andrew J; Holzmann, Maria; Caulle, Clémence; Goineau, Aurélie; Kamenskaya, Olga; Weber, Alexandra A. -T.; Pawlowski, Jan (1 March 2017). "Giant protists (xenophyophores, Foraminifera) are exceptionally diverse in parts of the abyssal eastern Pacific licensed for polymetallic nodule exploration". Biyolojik Koruma. 207: 106–116. doi:10.1016/j.biocon.2017.01.006. ISSN 0006-3207.
- ^ McIlroy, Duncan; Green, O. R.; Brasier, M. D. (2001). "Palaeobiology and evolution of the earliest agglutinated Foraminifera: Platysolenites, Spirosolenites and related forms". Lethaia. 34 (1): 13–29. doi:10.1080/002411601300068170. ISSN 1502-3931.
- ^ a b Scott, David B.; Medioli, Franco; Braund, Regan (1 June 2003). "Foraminifera from the Cambrian of Nova Scotia: The oldest multichambered foraminifera". Mikropaleontoloji. 49 (2): 109–126. doi:10.2113/49.2.109. ISSN 1937-2795.
- ^ a b c d e f Tappan, Helen; Loeblich, Alfred R. (1988). "Foraminiferal Evolution, Diversification, and Extinction". Paleontoloji Dergisi. 62 (5): 695–714. ISSN 0022-3360. JSTOR 1305391.
- ^ "Fusulinids | GeoKansas". geokansas.ku.edu. Alındı 16 Mayıs 2020.
- ^ Czaplewski, John J. "PBDB Navigator". paleobiodb.org. Alındı 16 Mayıs 2020.
- ^ Gräfe, K.U. (2005). "Benthic foraminifers and palaeoenvironment in the Lower and Middle Jurassic of the Western Basque-Cantabrian Basin (Northern Spain)". İber Jeolojisi Dergisi. 31 (2): 217–233. S2CID 55664447.
- ^ a b "Nature debates".
- ^ Journal bioinformatics and biology insights, Using the Multiple Analysis Approach to Reconstruct Phylogenetic Relationships among Planktonic Foraminifera from Highly Divergent and Length-polymorphic SSU rDNA Sequences
- ^ Gebhardt, Holger (1 February 1997). "Cenomanian to Turonian foraminifera from Ashaka (NE Nigeria): quantitative analysis and palaeoenvironmental interpretation". Kretase Araştırmaları. 18 (1): 17–36. doi:10.1006/cres.1996.0047. ISSN 0195-6671.
- ^ Báldi, Katalin; Benkovics, László; Sztanó, Orsolya (1 May 2002). "Badenian (Middle Miocene) basin development in SW Hungary: subsidence history based on quantitative paleobathymetry of foraminifera". Uluslararası Yer Bilimleri Dergisi. 91 (3): 490–504. Bibcode:2002IJEaS..91..490B. doi:10.1007/s005310100226. ISSN 1437-3262. S2CID 129296067.
- ^ Australia, c=AU;o=Australia Government;ou=Geoscience (15 May 2014). "Biostratigraphy". www.ga.gov.au. Alındı 20 Temmuz 2020.
- ^ Cushman, Joseph A.; Ellisor, Alva C. (1 January 1945). "The Foraminiferal Fauna of the Anahuac Formation". Paleontoloji Dergisi. 19 (6): 545–572. JSTOR 1299203.
- ^ Zachos, J.C.; Pagani, M .; Sloan, L.; Thomas, E .; Billups, K. (2001). "Trends, Rhythms, and Aberrations in Global Climate, 65 Ma to Present". Bilim. 292 (5517): 686–693. Bibcode:2001Sci...292..686Z. doi:10.1126/science.1059412. PMID 11326091. S2CID 2365991.
- ^ Branson, Oscar; Redfern, Simon A.T.; Tyliszczak, Tolek; Sadekov, Aleksey; Langer, Gerald; Kimoto, Katsunori; Elderfield, Henry (December 2013). "The coordination of Mg in foraminiferal calcite". Dünya ve Gezegen Bilimi Mektupları. 383: 134–141. Bibcode:2013E&PSL.383..134B. doi:10.1016/j.epsl.2013.09.037.
- ^ Misra, S.; Froelich, P. N. (26 January 2012). "Lithium Isotope History of Cenozoic Seawater: Changes in Silicate Weathering and Reverse Weathering". Bilim. 335 (6070): 818–823. Bibcode:2012Sci...335..818M. doi:10.1126/science.1214697. PMID 22282473. S2CID 42591236.
- ^ Hemming, N.G.; Hanson, G.N. (Ocak 1992). "Boron isotopic composition and concentration in modern marine carbonates". Geochimica et Cosmochimica Açta. 56 (1): 537–543. Bibcode:1992GeCoA..56..537H. doi:10.1016/0016-7037(92)90151-8.
- ^ Boardman, R.S .; Cheetham, A.H .; Rowell, A.J. (1987). Fosil Omurgasızlar. Wiley. ISBN 978-0865423022.
- ^ Jones, R.W. (1996). Petrol aramalarında mikropaleontoloji. Clarendon Press. ISBN 978-0-19-854091-5.
- ^ McNeil, D.H .; Yayıncı, D.R .; Snowdon, L.R. (1996). Fosil Foraminiferlerde Renk Değişimi, Termal Olgunluk ve Gömü Diyajenez. Kanada Bülteni Jeolojik Araştırması. 499. Kanada Jeolojik Araştırması. ISBN 978-0-660-16451-9.
- ^ Wilkinson, Ian P .; Williams, Mark; Young, Jeremy R .; Cook, Samantha R .; Fulford, Michael G .; Lott, Graham K. (1 Ağustos 2008). "Mikrofosillerin Silchester'deki Roma mozaiklerinin üretiminde kullanılan tebeşirlerin menşeini değerlendirmede uygulanması". Arkeolojik Bilimler Dergisi. 35 (8): 2415–2422. doi:10.1016 / j.jas.2008.03.010. ISSN 0305-4403.
Dış bağlantılar
- Genel bilgi
- California Üniversitesi Paleontoloji Müzesi web sitesinde bir Foraminifer'e Giriş
- Güney Florida Üniversitesi'ndeki araştırmacılar bir sistem geliştirdi mercan resif ortamlarını izlemek için Foraminifera kullanma
- University College London's mikropaleontoloji sitesi birçok yüksek kaliteli dahil olmak üzere Foraminifera'ya genel bir bakış sunar SEM'ler
- Foraminifer araştırmasında kullanılan resimli terimler sözlüğü OA e-dergisinde yayınlanan Lukas Hottinger'ın sözlüğüdür "Carnets de Géologie - Jeoloji Üzerine Defterler"
- Foraminifer hakkında bilgi Martin Langer'in Mikropaleontoloji Sayfası
- Bentik Foraminifer bilgileri 2005 Urbino Paleoklimatoloji Yaz Okulu'ndan
- Çevrimiçi flip-kitaplar
- Foraminifer araştırmasında kullanılan resimli terimler sözlüğü tarafından Lukas Hottinger (yayınlanan birinin alternatif versiyonu "Carnets de Géologie - Jeoloji Üzerine Defterler" )
- Kaynaklar
- pforams @ mikrotax - planktonik foraminiferlerin taksonomisini detaylandıran çevrimiçi bir veritabanı
- yıldız * kum projesi (parçası mikroskop ) Foraminifera hakkında ortak bir bilgi veritabanıdır.
- 3B modeller forams, tarafından oluşturulan X-ışını tomografisi
- KRONOLAR vardır birkaç Foraminifera kaynağı dahil takson arama sayfası ve bir mikro-paleo bölümü NB Bu içeriğin çoğu artık pforams @ mikrotax web sitesinde bulunmaktadır
- eForams Foraminifer'e ve foraminifer kabuklarının modellenmesine odaklanan bir web sitesidir
- Foraminifera Galerisi Cins ve bölgeye göre yakın tarihli ve fosil Foraminiferlerin resimli kataloğu
- "Foraminifer". NCBI Taksonomi Tarayıcısı. 29178.