Flagellum - Flagellum
Bu makalenin olması önerildi Bölünmüş başlıklı makalelere Bakteriyel tip kamçı ve Ökaryotik tip kamçı. (Tartışma) (Aralık 2020) |
| Flagellum | |
|---|---|
 Bakteriyel kamçı yapısının yapısı. | |
 SEM kamçılı görüntüsü Chlamydomonas sp. (10000 ×) | |
| Tanımlayıcılar | |
| MeSH | D005407 |
| TH | H1.00.01.1.01032 |
| FMA | 67472 |
| Anatomik terminoloji | |
Bir kamçı (/fləˈdʒɛləm/; çoğul: kamçı) kirpik benzeri bir uzantıdır. vücut hücresi Belli ki bakteri ve ökaryotik olarak adlandırılan hücreler kamçılılar. Bir kamçıda bir veya birkaç kamçı bulunabilir. Bir kamçının birincil işlevi, hareket ama aynı zamanda duyusal olarak da işlev görür organel hücre dışındaki kimyasallara ve sıcaklıklara duyarlıdır.[1][2][3][4] Benzer yapı Archaea aynı şekilde işlev görür, ancak yapısal olarak farklıdır ve Archaellum.[5]
Flagella, yapıdan çok işlevle tanımlanan organellerdir. Flagella büyük ölçüde değişir. Hem prokaryotik hem de ökaryotik flagella yüzme için kullanılabilir, ancak protein bileşimi, yapısı ve itme mekanizması açısından büyük farklılıklar gösterirler. Kelime kamçı içinde Latince anlamına geliyor kırbaç.
Kamçılı bir örnek bakteri ülsere neden olur Helikobakter pilori, mideye ulaşmak için mukus astarından kendisini itmek için birden fazla flagella kullanan epitel.[6] Ökaryotik kamçılı hücrenin bir örneği, memeli sperm hücresi, kendisini dişi üreme kanalında ilerletmek için kamçısını kullanan.[7] Ökaryotik flagella yapısal olarak ökaryotik ile aynıdır kirpikler Bazen işlev veya uzunluğa göre ayrımlar yapılsa da.[8] Fimbriae ve pili aynı zamanda ince eklerdir, ancak farklı işlevlere sahiptir ve genellikle daha küçüktür.
Türler

Şimdiye kadar üç tip flagella ayırt edildi: bakteriyel, arkaya ve ökaryotik.
Bu üç tür arasındaki temel farklar şunlardır:
- Bakteriyel flagella, her biri bir döner motor saat yönünde veya saat yönünün tersine dönebilen tabanında.[10][11][12] Birkaç çeşit bakteri hareketliliği sağlarlar.[13][14]
- Archaeal flagella (Archaella ) yüzeysel olarak bakteriyel kamçıya benzer, ancak birçok ayrıntıda farklıdır vehomolog.[15][16][17]
- Ökaryotik kamçı - hayvan, bitki ve protist hücrelerinkiler - ileri geri hareket eden karmaşık hücresel projeksiyonlardır. Ökaryotik flagella, ökaryotik ile birlikte sınıflandırılır. hareketli kirpikler gibi undulipodia[18] hücresel işlevdeki belirgin dalgalı uzantı rollerini vurgulamak için veya hareketlilik. Birincil kirpikler hareketsizdir ve unlulipodi değildir; Onlar bir .. sahip yapısal olarak farklı 9 + 0 aksonem Yerine 9 + 2 aksonem hem flagella hem de hareketli kirpikler bulunur.
Bakteriyel

Yapı ve kompozisyon
Bakteriyel kamçı, protein kamçı. Şekli 20-nanometre -kalın içi boş tüp. Bu helezoni ve dış zarın hemen dışında keskin bir kıvrımı vardır; bu "kanca", sarmalın ekseninin doğrudan hücreden uzağa bakmasına izin verir. Kanca ve kancanın arasında bir şaft uzanır. bazal vücut hücre zarındaki yatak görevi gören protein halkalarından geçerek. Gram pozitif organizmalar bu bazal vücut halkalarından ikisine sahiptir, biri peptidoglikan katman ve biri hücre zarı. Gram negatif organizmaların böyle dört halkası vardır: L yüzük ile ortak lipopolisakkaritler, P halkası ile ilişkili peptidoglikan katman, M halkası hücre zarı ve S halkası doğrudan plazma zarına bağlanır. Filaman, bir başlık proteini ile biter.[19][20]
Flagellar filament, motor tarafından döndürüldüğünde bakteriyi kancanın içinden iten uzun, sarmal vidadır. Gram negatif dahil olmak üzere incelenen çoğu bakteride Escherichia coli, Salmonella typhimurium, Caulobacter crescentus, ve Vibrio alginolyticus filaman, filaman eksenine yaklaşık olarak paralel 11 protofilamentten oluşur. Her protofilament, bir dizi tandem protein zinciridir. Ancak, Campylobacter jejuni yedi protofilamenti vardır.[21]
Bazal gövdenin bazı türlerle ortak olan birkaç özelliği vardır. salgı gözenekleri merkezlerinde plazma membranı boyunca uzanan içi boş, çubuk benzeri "tıkaç" gibi. Bakteriyel flagella ile bakteriyel salgı sistemi yapıları ve proteinleri arasındaki benzerlikler, bakteriyel flagella'nın tip-üç salgı sistemi.
Motor
Bakteriyel kamçı, döner bir motorla çalıştırılır (Mot kompleksi ) flagellumun iç hücre zarındaki çapa noktasında bulunan proteinden oluşur. Motor tarafından desteklenmektedir proton güdü kuvveti yani bakteri hücre zarı boyunca protonların (hidrojen iyonları) akışıyla konsantrasyon gradyanı hücrenin metabolizması tarafından kurulur (Vibrio türlerin yanal ve kutupsal olmak üzere iki tür flagella vardır ve bazıları bir sodyum iyon pompası yerine Proton pompası[22]). Rotor, protonları zar boyunca taşır ve bu süreçte döndürülür. Rotor tek başına 6.000 ila 17.000 arasında çalışabilir rpm ancak kamçı filament takıldığında genellikle sadece 200 ila 1000 rpm'ye ulaşır. Dönüş yönü, flagellar motor anahtarı rotordaki FliG proteininin pozisyonundaki küçük bir değişikliğin neden olduğu neredeyse anında.[23] Kamçı, son derece enerji verimlidir ve çok az enerji kullanır.[24][güvenilmez kaynak? ] Tork üretimi için kesin mekanizma hala tam olarak anlaşılamamıştır.[25] Flagellar motorun açma-kapama düğmesi olmadığı için, protein epsE, motoru rotordan ayırmak için mekanik bir kavrama olarak kullanılır, böylece kamçı durdurulur ve bakterinin tek bir yerde kalmasına izin verilir.[26]
Flagella'nın silindirik şekli mikroskobik organizmaların hareketine uygundur; bu organizmalar düşük seviyede çalışır Reynolds sayısı, çevreleyen suyun viskozitesinin kütlesinden veya ataletinden çok daha önemli olduğu yerlerde.[27]
Flagella'nın dönme hızı, proton hareket kuvvetinin yoğunluğuna yanıt olarak değişir, bu nedenle belirli hız kontrolü biçimlerine izin verir ve ayrıca bazı bakteri türlerinin boyutlarıyla orantılı olarak dikkate değer hızlara ulaşmalarına izin verir; bazıları saniyede yaklaşık 60 hücre uzunluğuna ulaşır. Böyle bir hızda, bir bakterinin 1 km'yi kaplaması yaklaşık 245 gün sürer; bu yavaş görünse de, ölçek kavramı sunulduğunda perspektif değişir. Makroskopik yaşam formlarına kıyasla, saniyedeki vücut uzunluğu sayısı ile ifade edildiğinde gerçekten çok hızlıdır. Örneğin bir çita, saniyede yalnızca 25 vücut uzunluğuna ulaşır.[28]
Kamçılarının kullanımıyla, E. coli bir vasıtasıyla, cezbedici maddelere doğru ve kovuculardan uzaklaşarak hızla hareket edebilir. önyargılı rastgele yürüyüş kamçı döndürülerek ortaya çıkan 'koşular' ve 'taklalar' ile saat yönünün tersine ve saat yönünde, sırasıyla. İki dönme yönü aynı değildir (kamçı hareketine göre) ve bir moleküler anahtarla seçilir.[29]
Montaj
Flagellar montaj sırasında, flagellum bileşenleri, bazal gövdenin ve yeni oluşan filamanın içi boş çekirdeklerinden geçer. Montaj sırasında, protein bileşenleri taban yerine flagellar uçta eklenir.[30] Laboratuvar ortamındaflagellar filamentler, tek protein olarak saflaştırılmış flagellin içeren bir çözelti içinde kendiliğinden birleşir.[31]
Evrim
Bakteriyel flagellumun en az 10 protein bileşeni, homolog proteinleri paylaşır. tip üç salgı sistemi (T3SS),[32] dolayısıyla biri diğerinden evrimleşmiştir. T3SS, flagellar bir aparatla benzer sayıda bileşene (yaklaşık 25 protein) sahip olduğundan, hangisinin ilk önce evrimleştiğini belirlemek zordur. Bununla birlikte, flagellar sistemi, çeşitli düzenleyiciler ve şaperonlar da dahil olmak üzere genel olarak daha fazla protein içeriyor gibi görünmektedir, bu nedenle flagella'nın bir T3SS'den evrimleştiği iddia edilmiştir. Ancak, aynı zamanda önerildi[33] kamçı ilk önce evrimleşmiş olabilir veya iki yapı paralel olarak gelişmiştir. Erken tek hücreli organizmaların ihtiyacı hareketlilik (mobilite) önce evrim tarafından daha mobil flagella'nın seçileceğini destekler,[33] ancak kamçıdan gelişen T3SS, 'indirgeyici evrim' olarak görülebilir ve sistemden topolojik destek almaz. filogenetik ağaçlar.[34] İki yapının ortak bir atadan ayrı evrimleştiği hipotezi, iki yapı arasındaki protein benzerliklerini ve bunların işlevsel çeşitliliğini açıklar.[35]
Flagella ve akıllı tasarım tartışması
Bazı yazarlar, ancak tüm proteinler yerinde olduğunda düzgün bir şekilde işlev görebileceklerini varsayarak, flagella'nın evrimleşmiş olamayacağını savundu. Başka bir deyişle, flagellar aparat "indirgenemez karmaşık ".[36] Bununla birlikte, birçok protein silinebilir veya mutasyona uğratılabilir ve flagellum, bazen düşük verimlilikle de olsa, yine de çalışır.[37] Ek olarak, flagella'nın bileşimi şaşırtıcı bir şekilde bakteriler arasında çeşitlidir ve birçok protein sadece bazı türlerde bulunurken diğerlerinde bulunmaz.[38] Bu nedenle, flagellar aygıt, evrimsel açıdan açıkça çok esnektir ve protein bileşenlerini mükemmel şekilde kaybedebilir veya kazanabilir. Örneğin, bir dizi mutasyon bulunmuştur. artırmak hareketliliği E. coli.[39] Bakteriyel flagella'nın evrimi için ek kanıtlar arasında, körelmiş kamçı, ara kamçı formları ve flagellar protein sekansları arasındaki benzerlik kalıplarının varlığını, hemen hemen tüm çekirdek kamçı proteinlerinin flagellar olmayan proteinlerle bilinen homolojilere sahip olduğu gözlemini içerir.[32] Ayrıca, basit tekrar eden alt birimlerin kendiliğinden birleşmesi, müteakip sapma ile gen çoğaltması, diğer sistemlerden elementlerin toplanması ('moleküler bricolage') ve rekombinasyon dahil olmak üzere, flagellar evriminde önemli roller oynayan birkaç süreç tanımlanmıştır.[40]
Flagellar düzenleme şemaları

Farklı bakteri türleri, farklı sayıda ve farklı flagella düzenlemelerine sahiptir.
- Monotrichous bakterilerin tek bir flagellumu vardır (ör. Vibrio cholerae ).
- Lophotrichous bakteriler, bakterileri tek bir yönde hareket ettirmek için uyum içinde hareket eden, bakteri yüzeylerinde aynı noktada bulunan birden fazla flagella'ya sahiptir. Çoğu durumda, çoklu kamçı tabanları, hücre zarının özel bir bölgesi ile çevrilidir. kutup organel.[kaynak belirtilmeli ]
- Amphitrichous bakterilerin iki zıt ucunun her birinde tek bir flagellum vardır (bir seferde yalnızca bir flagellum çalışır ve bakterinin, hangi flagellum'un aktif olduğunu değiştirerek, hızla rotasını tersine çevirmesine izin verir).
- Peritrichous bakteriler her yöne doğru çıkıntı yapan flagella'ya sahiptir E. coli).
Bazı büyük biçimlerde Selenomonas, 30'dan fazla bireysel flagella, hücre gövdesinin dışında düzenlenmiştir ve a "fasikül ".
Spiroketler tersine, hücrenin zıt kutuplarından kaynaklanan ve içinde bulunan flagella var Periplazmik boşluk dış zarın kırılmasıyla ve daha yakın zamanda elektron kriyotomografisi mikroskopi.[41][42][43] İpliklerin hücre gövdesine göre dönmesi, tüm bakterinin, normal olarak kamçılı bakterilerin geçişini engelleyecek kadar viskoz malzeme içinde bile, tirbuşon benzeri bir hareketle ilerlemesine neden olur.
Tek renkli bir polar flagellumun saat yönünün tersine dönüşü, tıpkı mantarın içinde hareket eden bir tirbuşon gibi, arkada flagellum ile hücreyi ileri iter. Nitekim, mikroskobik ölçekte su oldukça yapışkan, günlük su deneyimimizden çok farklı.
Flagella solak helislerdir ve yalnızca saat yönünün tersine dönerken demetlenir ve birlikte döndürülür. Bazı rotorlar yön değiştirdiğinde, flagella gevşer ve hücre "yuvarlanmaya" başlar. Kamçıların tümü saat yönünde dönse bile, geometrik nedenlerden ve hidrodinamik nedenlerden dolayı büyük olasılıkla bir demet oluşturmayacaklardır.[44][45] Bu tür bir "yuvarlanma" ara sıra meydana gelebilir ve hücrenin görünüşte yerinde sallanmasına ve hücrenin yeniden yönlenmesine neden olabilir. Bir kamçının saat yönünde dönüşü, hücre için uygun olan kimyasal bileşikler (örneğin gıda) tarafından bastırılır, ancak motor buna oldukça uyumludur. Bu nedenle, uygun bir yönde hareket ederken, kimyasal çekicinin konsantrasyonu artar ve "taklalar" sürekli olarak baskılanır; ancak hücrenin hareket yönü elverişsiz olduğunda (örneğin, kimyasal bir cezbedici maddeden uzaklaştığında), yuvarlanmalar artık baskılanmaz ve hücrenin doğru yöne yeniden yönlendirilme şansı ile çok daha sık meydana gelir.
Bazılarında Vibrio spp. (özellikle Vibrio parahaemolyticus[46]) ve ilgili proteobakteriler gibi Aeromonas enerji için farklı gen kümeleri ve farklı iyon gradyanları kullanan iki flagellar sistem bir arada bulunur. Polar flagella yapısal olarak ifade edilir ve yığın sıvıda hareketlilik sağlarken, yanal flagella, polar flagella dönmeye karşı çok fazla dirençle karşılaştığında ifade edilir.[47][48][49][50][51][52] Bunlar yüzeylerde veya viskoz akışkanlarda kümelenme hareketi sağlar.
Archaeal
Archaellum bazılarının sahip olduğu Archeae yüzeysel olarak bakteriyel kamçıya benzer; 1980'lerde, kaba morfoloji ve davranış temelinde homolog oldukları düşünülüyordu.[53] Hem flagella hem de archaella, hücrenin dışına uzanan filamentlerden oluşur ve hücreyi ilerletmek için döner. Archaeal flagella, merkezi bir kanalı olmayan benzersiz bir yapıya sahiptir. Bakteriyel tip IV'e benzer Pilins arkeal flagellinler (archaellinler), sınıf 3 sinyal peptidleri ile yapılır ve bir tip IV prepilin peptidaz benzeri enzim tarafından işlenir. Archaellinler tipik olarak N-bağlantılı eklenmesi ile modifiye edilir. glikanlar uygun montaj veya işlev için gerekli olan.[4]
1990'lardaki keşifler, arkeal ve bakteriyel kamçı arasında çok sayıda ayrıntılı farklılığı ortaya çıkardı. Bunlar şunları içerir:
- Bakteriyel kamçı, bir akışla motorize edilir. H+ iyonlar (veya ara sıra Na+ iyonlar); archaeal flagella, neredeyse kesinlikle ATP. tork - archaeal flagellumun dönüşüne güç veren motor üreten motor tanımlanmamıştır.
- Bakteriyel hücreler genellikle her biri bağımsız olarak dönen birçok flagellar filamana sahipken, archaeal flagellum, tek bir grup halinde dönen birçok filamentten oluşan bir demetten oluşur.
- Bakteriyel flagella, uçta flagellin alt birimlerinin eklenmesiyle büyür; archaeal flagella, tabana alt birimlerin eklenmesiyle büyür.
- Bakteriyel flagella, archaella'dan daha kalındır ve bakteri filamenti, içinde flagellin alt birimlerinin filamentin içinden akıp ucuna eklenebilecek kadar geniş bir oyuk "tüpüne" sahiptir; arkellum buna izin vermeyecek kadar incedir (12-15 nm).[54]
- Bakteriyel flagella'nın birçok bileşeni, tip III sekresyon sistemleri ancak bakteriyel flagella ve archaella'nın bileşenleri hiçbir sekans benzerliği paylaşmaz. Bunun yerine, archaella'nın bazı bileşenleri, sıralı ve morfolojik benzerliği paylaşır. tip IV pili eylemi ile birleştirilen tip II salgı sistemleri (pili ve protein salgılama sistemlerinin adlandırılması tutarlı değildir).[54]
Bu farklılıklar olabilir[Gelincik kelimeler ] bakteriyel flagella ve archaella'nın klasik bir biyolojik vaka olabileceği anlamına gelir. benzetme veya yakınsak evrim, ziyade homoloji.[55][56] Bununla birlikte, onlarca yıllık iyi duyurulmuş bakteriyel flagella çalışmasına kıyasla (örn. Howard Berg ),[57] Archaella yakın zamanda[ne zaman? ] bilimsel ilgi çekmeye başladı.[kaynak belirtilmeli ]
Ökaryotik




Terminoloji
Bakteriyel flagella ile ökaryotik kirpikler ve flagella arasındaki ayrımı vurgulamayı amaçlayan bazı yazarlar, bu iki ökaryotik yapının adını "undulipodia "(ör. tarafından gönderilen tüm makaleler Margulis 1970'lerden beri)[58] veya her ikisi için "kirpikler" (örneğin, Hülsmann, 1992;[59] Adl ve diğerleri, 2012;[60] çoğu kağıt Cavalier-Smith ), bakteri yapısı için "flagella" nın korunması. Bununla birlikte, bu makalede benimsenen ökaryotlar için "kirpikler" ve "kamçı" terimlerinin ayırt edici kullanımı hala yaygındır (örneğin, Andersen ve diğerleri, 1991;[61] Leadbeater ve diğerleri, 2000).[62]
İç yapı
Ökaryotik kamçı, dokuz kaynaşmış çiftin bir demetidir. mikrotübül iki merkezi tek mikrotüpü çevreleyen ikili. Sözde "9 + 2" yapı, ökaryotik kamçı çekirdeğinin karakteristiğidir. aksonem. Ökaryotik flagellumun tabanında bir bazal vücut, "blefaroplast" veya kinetozom, mikrotübül düzenleme merkezi flagellar mikrotübüller için ve yaklaşık 500 nanometre uzunluğundadır. Bazal cisimler yapısal olarak aynıdır merkezler. Flagellum, hücrenin hücre zarı, böylece kamçının iç kısmına hücrenin sitoplazma.
Morfolojide nispeten sabit olan aksonem ve bazal gövdenin yanı sıra, flagellar aparatın diğer iç yapıları geçiş bölgesi (aksonem ve bazal gövdenin birleştiği yer) ve kök sistemidir (bazal cisimlerden sitoplazmaya uzanan mikrotübüler veya fibriler yapılardır) ), ökaryotların filogenetik ilişkilerinin göstergeleri olarak daha değişken ve kullanışlıdır. Daha nadir görülen diğer yapılar, paraflagellar (veya paraksiyal, paraksonemal) çubuk, R lifi ve S fiberidir.[63]:63–84 Yüzey yapıları için aşağıya bakın.
Mekanizma
Dıştaki 9 ikili mikrotübülün her biri bir çift dynein bitişik mikrotübüle kollar (bir "iç" ve bir "dış" kol); bunlar ATP hidrolizi yoluyla kuvvet üretir. Flagellar aksonem ayrıca şunları içerir: radyal parmaklıklar dış dokuz mikrotübülün her birinden merkez çifte doğru uzanan polipeptit kompleksleri, parmaklığın "başı" içe doğru bakar. Tam işlevi ve etki yöntemi henüz anlaşılmamış olmasına rağmen, radyal kolun flagellar hareketin düzenlenmesinde rol oynadığı düşünülmektedir.
Flagella ve kirpikler
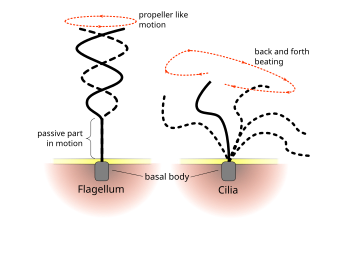
Ökaryotiklerin düzenli vuruş kalıpları kirpikler ve flagella hücresel düzeyde hareket oluşturur. Örnekler, yüzme gibi tek hücrelerin itici gücünden spermatozoa sıvının sabit bir hücre tabakası boyunca taşınmasına, örneğin solunum sistemi. Ökaryotik kamçı ve hareketli kirpikler ultra yapısal olarak özdeş olsa da, iki organelin çarpma şekli farklı olabilir. Flagella durumunda, hareket genellikle düzlemsel ve dalgaya benzer, oysa hareketli kirpikler genellikle bir güç ve iyileşme darbesiyle daha karmaşık üç boyutlu bir hareket gerçekleştirir.[kaynak belirtilmeli ]
Intraflagellar ulaşım
Intraflagellar ulaşım aksonemal alt birimlerin gerçekleştirdiği süreç, transmembran reseptörler ve diğer proteinler, flagellum boyunca yukarı ve aşağı hareket ettirilir, kamçının hem motilite hem de sinyal iletiminde düzgün çalışması için gereklidir.[64]
Evrim ve oluşum
Ökaryotik kamçı veya kirpikler, muhtemelen atadan kalma bir özelliktir,[65] hemen hemen tüm ökaryot gruplarında, nispeten kalıcı bir durum olarak veya kamçılı bir yaşam döngüsü aşaması olarak yaygındır (örn. hayvanat bahçeleri, gametler, hayvanat bahçesi sürekli olarak üretilebilen veya üretilemeyen).[66][67][60]
İlk durum, çok hücreli organizmaların özel hücrelerinde bulunur (örneğin, koanositler nın-nin süngerler veya kirpikli epitel nın-nin metazoanlar ), de olduğu gibi siliatlar ve "kamçılı duruma" (veya "monadoid") sahip birçok ökaryot organizasyon seviyesi ", görmek Flagellata yapay bir grup).
Flagellated yaşam döngüsü aşamaları birçok grupta bulunur, örneğin birçok yeşil alg (zoosporlar ve erkek gametler), Briyofitler (erkek gametler), pteridofitler (erkek gametler), bazıları jimnospermler (sikadlar ve Ginkgo, erkek gamet olarak), merkezli diyatomlar (erkek gametler), kahverengi algler (zoosporlar ve gametler), Oomycetes (eşeysiz zoosporlar ve gametler), hifokititler (zoosporlar), labirentülomisetler (zoosporlar), bazıları apicomplexans (gametler), bazıları radyolar (muhtemelen gametler),[68] foraminiferans (gametler), plazmodioforomisetler (zoosporlar ve gametler), miksogastridler (zoosporlar), metazoanlar (erkek gametler) ve Chytrid mantarlar (zoosporlar ve gametler).
Flagella veya kirpikler, bazı gruplarda, muhtemelen ilkel bir durum olmaktan ziyade bir kayıp nedeniyle tamamen yoktur. Kirpikler kaybı meydana geldi kırmızı yosun, biraz yeşil alg (Zygnematophyceae ), jimnospermler sikadlar hariç ve Ginkgo, anjiyospermler, flama diyatomlar, biraz apicomplexans, biraz amipozoanlar bazılarının sperminde metazoanlar,[69] ve mantarlar (dışında Chytrids ).
Tipoloji
Ökaryotları karakterize etmek için flagella veya cilia ile ilgili bir dizi terim kullanılır.[67][70][63]:60–63[71][72] Mevcut yüzey yapılarına göre flagella şunlar olabilir:
- whiplash flagella (= pürüzsüz, kısaltılmış flagella): kılsız, örn. Opisthokonta
- tüylü kamçı (= tinsel, parıldayan, plöronematik kamçı): kıllarla (= mastigonemler sensu lato), bölünmüştür:
- ince tüylerle (= tübüler olmayan veya basit tüyler): Euglenophyceae, Dinoflagellata, biraz Haptophyceae (Pavlovales )
- sert kıllarla (= tübüler kıllar, retronemler, mastigonemler) sensu stricto), bölünmüştür:
- bipartite kıllar: iki bölgeli. Oluşur Cryptophyceae, Prasinophyceae, ve bazı Heterokonta
- üç bölgeli (= straminipilous) kıllar: üç bölgeli (bir taban, bir boru şekilli şaft ve bir veya daha fazla terminal kıl). Çoğunda görülür Heterokonta
- stichonematic flagella: tek sıra kıl ile
- pantonematik flagella: iki sıra kıl ile
- akronematik: tek, terminal mastigoneme veya flagellar saçlı flagella (örn. bodonidler );[73] bazı yazarlar bu terimi kırbaçla eşanlamlı olarak kullanır
- ölçeklerle: ör. Prasinophyceae
- dikenli: ör., bazıları kahverengi algler
- dalgalı membranlı: ör. kinetoplastidler, biraz parabasalidler
- hortum ile (hücrenin gövde benzeri çıkıntısı): ör., Apusomonadlar, biraz bodonidler[74]
Flagella sayısına göre hücreler şu şekilde olabilir (bazı yazarların "kamçılı" yerine "siliyer" kullandıkları unutulmamalıdır:[60][75]
- yumuşatılmamış: ör. çoğu Opisthokonta
- biflagellated: ör. tümü Dinoflagellata Gametleri Charophyceae, çoğunun Briyofitler ve bazılarının metazoanlar[69]
- üç bayraklı: örneğin, bazılarının gametleri Foraminifer
- dört işaretli: örneğin, bazı Prasinophyceae, Collodictyonidae
- sekiz işaretli: ör. bazı Diplomonada, biraz Prasinophyceae
- çok hücreli: ör. Opalinata, Ciliophora, Stephanopogon, Parabasalida, Hemimastigophora, Caryoblastea, Multicilia, gametler (veya hayvanat bahçeleri ) nın-nin Oedogoniales (Chlorophyta ), biraz pteridofitler ve bazı jimnospermler
Flagella'nın yerleştirildiği yere göre:[76]
- opisthokont: arkaya yerleştirilmiş flagellalı hücreler, örn. Opisthokonta (Vischer, 1945). İçinde Haptophyceae flagella, lateral olarak terminale yerleştirilir, ancak hızlı yüzme sırasında arkaya doğru yönlendirilir.[77]
- akrokont: apikal olarak yerleştirilmiş flagellalı hücreler
- subakrokont: subapikal olarak eklenen flagellalı hücreler
- pleurokont: yanal olarak yerleştirilmiş flagellalı hücreler
Dayak düzenine göre:
- kayma: alt tabaka üzerinde izleyen bir kamçı[74]
- heterodinamik: farklı vurma modellerine sahip flagella (genellikle bir flagellum yiyecek yakalamada, diğeri ise süzülme, demirleme, itme veya "direksiyonda" işlev görür)[78]
- izodinamik: aynı desenlerle atan flagella
Flagellar türüyle ilgili diğer terimler:
- isokont: eşit uzunlukta flagellalı hücreler. Aynı zamanda eskiden Chlorophyta
- anisokont: eşit olmayan uzunlukta flagellalı hücreler, örneğin bazıları Euglenophyceae ve Prasinophyceae
- heterokont: Luther (1899) tarafından ortaya atılan terim Xanthophyceae, eşit olmayan uzunluktaki flagella çifti nedeniyle. Anterior straminipilous flagellum (bir veya iki sıra halinde üçlü mastigonemler ile) ve posterior genellikle pürüzsüz flagelluma sahip hücrelere atıfta bulunarak özel bir anlam kazanmıştır. Ayrıca taksona atıfta bulunmak için kullanılır Heterokonta
- stephanokont: ön ucuna yakın bir flagella taçlı hücreler, örneğin, gametler ve sporlar Oedogoniales bazılarının sporları Bryopsidales. Blackman & Tansley (1902) tarafından ortaya atılan terim Oedogoniales
- akont: kamçı içermeyen hücreler. Ayrıca taksonomik gruplara Aconta veya Akonta olarak atıfta bulunmak için kullanılmıştır: Zygnematophyceae ve Bacillariophyceae (Oltmanns, 1904) veya Rhodophyceae (Christensen, 1962)
Ayrıca bakınız
Referanslar
- ^ Wang Q, Suzuki A, Mariconda S, Porwollik S, Harshey RM (Haziran 2005). "Islaklığı hissetmek: bakteri kamçısı için yeni bir rol". EMBO Dergisi. 24 (11): 2034–42. doi:10.1038 / sj.emboj.7600668. PMC 1142604. PMID 15889148.
- ^ Bardy SL, Ng SY, Jarrell KF (Şubat 2003). "Prokaryotik hareketlilik yapıları". Mikrobiyoloji. 149 (Pt 2): 295–304. doi:10.1099 / mic.0.25948-0. PMID 12624192.
- ^ Silflow CD, Lefebvre PA (Aralık 2001). "Ökaryotik kirpikler ve kamçılıların montajı ve hareketliliği. Chlamydomonas reinhardtii'den dersler". Bitki Fizyolojisi. 127 (4): 1500–7. doi:10.1104 / pp.010807. PMC 1540183. PMID 11743094.
- ^ a b Jarrell K, ed. (2009). Pili ve Flagella: Güncel Araştırma ve Gelecek Eğilimler. Caister Academic Press. ISBN 978-1-904455-48-6.
- ^ Albers SV, Jarrell KF (27 Ocak 2015). "Archaellum: Archaea nasıl yüzer". Mikrobiyolojide Sınırlar. 6: 23. doi:10.3389 / fmicb.2015.00023. PMC 4307647. PMID 25699024.
- ^ Lacy BE, Rosemore J (Ekim 2001). "Helicobacter pylori: ülserler ve daha fazlası: bir çağın başlangıcı". Beslenme Dergisi. 131 (10): 2789S - 2793S. doi:10.1093 / jn / 131.10.2789S. PMID 11584108. Arşivlenen orijinal (özet sayfası) 7 Şubat 2009'da. Alındı 2 Haziran 2008.
- ^ Malo AF, Gomendio M, Garde J, Lang-Lenton B, Soler AJ, Roldan ER (Haziran 2006). "Sperm tasarımı ve sperm işlevi". Biyoloji Mektupları. 2 (2): 246–9. doi:10.1098 / rsbl.2006.0449. PMC 1618917. PMID 17148374.
- ^ Haimo LT, Rosenbaum JL (Aralık 1981). "Kirpikler, kamçı ve mikrotübüller". Hücre Biyolojisi Dergisi. 91 (3 Pt 2): 125s – 130s. doi:10.1083 / jcb.91.3.125s. PMC 2112827. PMID 6459327.
- ^ Streif S, Staudinger WF, Marwan W, Oesterhelt D (2008). "Archaeon Halobacterium salinarum'daki Flagellar rotasyonu ATP'ye bağlıdır". Moleküler Biyoloji Dergisi. 384 (1): 1–8. doi:10.1016 / j.jmb.2008.08.057. PMID 18786541.
- ^ Silverman M, Simon M (Mayıs 1974). "Flagellar rotasyonu ve bakteriyel hareketlilik mekanizması". Doğa. 249 (452): 73–4. Bibcode:1974Natur.249 ... 73S. doi:10.1038 / 249073a0. PMID 4598030. S2CID 10370084.
- ^ Meister GL, Berg HC (1987). "Yüzen bakterilerde kamçı demetlerinin hızlı dönüşü". Doğa. 325 (6105): 637–640. Bibcode:1987Natur.325..637L. doi:10.1038 / 325637a0. S2CID 4242129.
- ^ Berg HC, Anderson RA (Ekim 1973). "Bakteriler kamçı ipliklerini döndürerek yüzerler". Doğa. 245 (5425): 380–2. Bibcode:1973Natur.245..380B. doi:10.1038 / 245380a0. PMID 4593496. S2CID 4173914.
- ^ Jahn TL, Bovee EC (1965). "Mikroorganizmaların hareketi ve hareketi". Mikrobiyolojinin Yıllık İncelemesi. 19: 21–58. doi:10.1146 / annurev.mi.19.100165.000321. PMID 5318439.
- ^ Harshey RM (2003). "Bir yüzeydeki bakteri hareketliliği: ortak bir hedefe giden birçok yol". Mikrobiyolojinin Yıllık İncelemesi. 57: 249–73. doi:10.1146 / annurev.micro.57.030502.091014. PMID 14527279.
- ^ Ng SY, Chaban B, Jarrell KF (2006). "Archaeal flagella, bakteriyel flagella ve tip IV pili: genlerin ve posttranslasyonel modifikasyonların karşılaştırması". Moleküler Mikrobiyoloji ve Biyoteknoloji Dergisi. 11 (3–5): 167–91. doi:10.1159/000094053. PMID 16983194. S2CID 30386932.
- ^ Metlina AL (Kasım 2004). "Prokaryotik hareketlilik organelleri olarak bakteriyel ve arkael kamçı". Biyokimya. Biokhimiia. 69 (11): 1203–12. doi:10.1007 / s10541-005-0065-8. PMID 15627373. S2CID 632440.
- ^ Jarrell K (2009). "Archaeal Flagella ve Pili". Pili ve Flagella: Güncel Araştırma ve Gelecek Trendler. Caister Academic Press. ISBN 978-1-904455-48-6.
- ^ Biyoloji Sözlüğü, 2004, erişim tarihi: 2011-01-01.
- ^ Macnab RM (2003). "Bakteriler flagella'yı nasıl birleştirir". Mikrobiyolojinin Yıllık İncelemesi. 57: 77–100. doi:10.1146 / annurev.micro.57.030502.090832. PMID 12730325.
- ^ Diószeghy Z, Závodszky P, Namba K, Vonderviszt F (Haziran 2004). "HAP2 kapama ile flagellar filamentlerin stabilizasyonu". FEBS Mektupları. 568 (1–3): 105–9. doi:10.1016 / j.febslet.2004.05.029. PMID 15196929. S2CID 33886010.
- ^ Galkin VE, Yu X, Bielnicki J, Heuser J, Ewing CP, Guerry P, Egelman EH (Nisan 2008). "Bakteriyel kamçı filamentler arasında kuaterner yapıların ıraksaması". Bilim. 320 (5874): 382–5. Bibcode:2008Sci ... 320..382G. doi:10.1126 / bilim.1155307. PMID 18420936. S2CID 7702002.
- ^ Atsumi T, McCarter L, Imae Y (Ocak 1992). "Marine Vibrio'nun kutupsal ve yanal flagellar motorları, farklı iyon güdümlü kuvvetler tarafından çalıştırılır. Doğa. 355 (6356): 182–4. Bibcode:1992Natur.355..182A. doi:10.1038 / 355182a0. PMID 1309599. S2CID 4315167.
- ^ Dean T (2 Ağustos 2010). "Doğanın en verimli motoru: flagellar". Avustralyalı Yaşam Bilimcisi.
- ^ Nagata Y (Haziran 2014). "Doğanın nanomotorunun sırlarını açığa çıkarmak". Nikkei Asya İnceleme.
- ^ Mora T, Yu H, Sowa Y, Wingreen NS (Ekim 2009). "Bakteriyel kamçı motorundaki adımlar". PLOS Hesaplamalı Biyoloji. 5 (10): e1000540. arXiv:0904.0438. Bibcode:2009PLSCB ... 5E0540M. doi:10.1371 / journal.pcbi.1000540. PMC 2759076. PMID 19851449.
- ^ Whitfield J (19 Haziran 2008). "Bakteriyel motorların kendi debriyajları vardır". Doğa Haberleri. doi:10.1038 / haber.2008.903. Alındı 17 Mayıs 2017.
- ^ Dusenbery DB (2009). "Bölüm 13". Mikro Ölçekte Yaşamak: Küçük Olmanın Beklenmedik Fiziği. Cambridge: Harvard Üniversitesi Yayınları. ISBN 978-0-674-03116-6.
- ^ Hildebrand M (Kasım 1959). "Çita ve Atın Hareketleri". Journal of Mammalogy. 44 (4): 481–495. doi:10.2307/1376265. JSTOR 1376265. Göre olmasına rağmen Hunter, Luke; Hamman, Dave (2003). Çita. Struik Yayıncılar. s. 37–38.
çitanın kaydedilen en hızlı hızı 110 km / sa (68 mil / sa) idi
- ^ Meadows R (Mayıs 2011). "Bakteriler nasıl vites değiştiriyor?". PLOS Biyoloji. 9 (5): e1001061. doi:10.1371 / journal.pbio.1001061. PMC 3091840. PMID 21572986.
- ^ Minamino T, Imada K, Namba K (Kasım 2008). "Bakteriyel flagellar birleşme için tip III protein ihraç mekanizmaları". Moleküler Biyo Sistemler. 4 (11): 1105–15. doi:10.1039 / b808065h. PMID 18931786.
- ^ Asakura S, Eguchi G, Iino T (Ekim 1964). "In Vitro Bakteriyel Flagella Sulandırılması". Moleküler Biyoloji Dergisi. 10: 42–56. doi:10.1016 / S0022-2836 (64) 80026-7. PMID 14222895.
- ^ a b Pallen MJ, Matzke NJ (Ekim 2006). "Türlerin Kökeni'nden bakteri kamçısının kökenine". Doğa Yorumları. Mikrobiyoloji. 4 (10): 784–90. doi:10.1038 / nrmicro1493. PMID 16953248. S2CID 24057949.
- ^ a b Saier MH (Mart 2004). "Bakteriyel tip III protein salgılama sistemlerinin evrimi". Mikrobiyolojideki Eğilimler. 12 (3): 113–5. doi:10.1016 / j.tim.2004.01.003. PMID 15001186.
- ^ Gophna U, Ron EZ, Graur D (Temmuz 2003). "Bakteriyel tip III sekresyon sistemleri çok eskidir ve çoklu yatay transfer olayları ile gelişmiştir". Gen. 312: 151–63. doi:10.1016 / S0378-1119 (03) 00612-7. PMID 12909351.
- ^ McCann HC, Guttman DS (2008). "Tip III salgı sisteminin evrimi ve bitki-mikrop etkileşimlerinde efektörleri". Yeni Fitolog. 177 (1): 33–47. doi:10.1111 / J.1469-8137.2007.02293.X. PMID 18078471.
- ^ Behe, M. (2007) Evrimin Sınırı. Özgür Basın, New York
- ^ Rajagopala SV, Titz B, Goll J, Parrish JR, Wohlbold K, McKevitt MT, Palzkill T, Mori H, Finley RL, Uetz P (2007). "Bakteriyel hareketliliğin protein ağı". Moleküler Sistem Biyolojisi. 3: 128. doi:10.1038 / msb4100166. PMC 1943423. PMID 17667950.
- ^ Titz B, Rajagopala SV, Ester C, Häuser R, Uetz P (Kasım 2006). "Bakteriyel kamçıda yeni korunmuş montaj faktörü". Bakteriyoloji Dergisi. 188 (21): 7700–6. doi:10.1128 / JB.00820-06. PMC 1636259. PMID 16936039.
- ^ Kakkanat A, Phan MD, Lo AW, Beatson SA, Schembri MA (10 Mayıs 2017). "Escherichia coli ST131'in artmış motilitesiyle ilişkili yeni genler". PLOS ONE. 12 (5): e0176290. Bibcode:2017PLoSO..1276290K. doi:10.1371 / journal.pone.0176290. PMC 5425062. PMID 28489862.
- ^ Pallen MJ, Gophna U (2007). "Bakteriyel kamçı ve Tip III sekresyon: karmaşıklığın evriminde vaka çalışmaları". Genom Dinamiği. 3: 30–47. doi:10.1159/000107602. ISBN 978-3-8055-8340-4. PMID 18753783.
- ^ Izard J, Renken C, Hsieh CE, Desrosiers DC, Dunham-Ems S, La Vake C, Gebhardt LL, Limberger RJ, Cox DL, Marko M, Radolf JD (Aralık 2009). "Kriyo-elektron tomografisi, sifiliz spiroket olan Treponema pallidum'un moleküler yapısını aydınlatıyor". Bakteriyoloji Dergisi. 191 (24): 7566–80. doi:10.1128 / JB.01031-09. PMC 2786590. PMID 19820083.
- ^ Izard J, Hsieh CE, Limberger RJ, Mannella CA, Marko M (Temmuz 2008). "Treponema denticola'nın yerel hücresel mimarisi kriyo-elektron tomografisi ile ortaya çıktı". Yapısal Biyoloji Dergisi. 163 (1): 10–7. doi:10.1016 / j.jsb.2008.03.009. PMC 2519799. PMID 18468917.
- ^ Kudryashev M, Cyrklaff M, Baumeister W, Simon MM, Wallich R, Frischknecht F (Mart 2009). "Patojenik Lyme hastalığı spiroketlerinin karşılaştırmalı kriyo-elektron tomografisi". Moleküler Mikrobiyoloji. 71 (6): 1415–34. doi:10.1111 / j.1365-2958.2009.06613.x. PMID 19210619. S2CID 19650892.
- ^ Kim M, Bird JC, Van Parys AJ, Breuer KS, Powers TR (Aralık 2003). "Bakteriyel flagellar kümelenmenin makroskopik ölçekli bir modeli". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 100 (26): 15481–5. arXiv:cond-mat / 0312562. Bibcode:2003PNAS..10015481K. doi:10.1073 / pnas.2633596100. PMC 307593. PMID 14671319.
- ^ Macnab RM (Ocak 1977). "Demetler halinde dönen bakteri kamçısı: sarmal geometride bir çalışma". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 74 (1): 221–5. Bibcode:1977PNAS ... 74..221M. doi:10.1073 / pnas.74.1.221. PMC 393230. PMID 264676.
- ^ Kim YK, McCarter LL (Temmuz 2000). "Vibrio parahaemolyticus'un kutupsal flagellar gen sisteminin analizi". Bakteriyoloji Dergisi. 182 (13): 3693–704. doi:10.1128 / JB.182.13.3693-3704.2000. PMC 94540. PMID 10850984.
- ^ Atsumi T, Maekawa Y, Yamada T, Kawagishi I, Imae Y, Homma M (Ağustos 1996). "Vibrio alginolyticus'un yanal ve polar kamçılarının yüzmeye etkisi". Bakteriyoloji Dergisi. 178 (16): 5024–6. doi:10.1128 / jb.178.16.5024-5026.1996. PMC 178290. PMID 8759871.
- ^ McCarter LL (2004). "Çift bayraklı sistemler, farklı koşullar altında hareketliliği mümkün kılar". Moleküler Mikrobiyoloji ve Biyoteknoloji Dergisi. 7 (1–2): 18–29. doi:10.1159/000077866. PMID 15170400. S2CID 21963003.
- ^ Merino S, Shaw JG, Tomás JM (Ekim 2006). "Bakteriyel yanal kamçı: uyarılabilir bir kamçı sistemi". FEMS Mikrobiyoloji Mektupları. 263 (2): 127–35. doi:10.1111 / j.1574-6968.2006.00403.x. PMID 16978346.
- ^ Belas R, Simon M, Silverman M (Temmuz 1986). "Vibrio parahaemolyticus'ta lateral flagella gen transkripsiyonunun düzenlenmesi". Bakteriyoloji Dergisi. 167 (1): 210–8. doi:10.1128 / jb.167.1.210-218.1986. PMC 212863. PMID 3013835.
- ^ Kanallar R, Altarriba M, Vilches S, Horsburgh G, Shaw JG, Tomás JM, Merino S (Şubat 2006). "Aeromonas hydrophila AH-3'ün lateral flagellar gen sisteminin analizi". Bakteriyoloji Dergisi. 188 (3): 852–62. doi:10.1128 / JB.188.3.852-862.2006. PMC 1347325. PMID 16428388.
- ^ Kanallar R, Ramirez S, Vilches S, Horsburgh G, Shaw JG, Tomás JM, Merino S (Ocak 2006). "Aeromonas hydrophila'da polar flagellum biyojenezi". Bakteriyoloji Dergisi. 188 (2): 542–55. doi:10.1128 / JB.188.2.542-555.2006. PMC 1347287. PMID 16385045.
- ^ Cavalier-Smith T (1987). "Ökaryotik ve arkebakteriyel hücrelerin kökeni". New York Bilimler Akademisi Yıllıkları. 503 (1): 17–54. Bibcode:1987NYASA.503 ... 17C. doi:10.1111 / j.1749-6632.1987.tb40596.x. PMID 3113314. S2CID 38405158.[kalıcı ölü bağlantı ]
- ^ a b Ghosh A, Albers SV (Ocak 2011). "Archaeal flagellumun montajı ve işlevi". Biyokimya Topluluğu İşlemleri. 39 (1): 64–9. doi:10.1042 / BST0390064. PMID 21265748. S2CID 23810797.
- ^ Thomas NA, Bardy SL, Jarrell KF (Nisan 2001). "Archaeal flagellum: farklı bir prokaryotik hareketlilik yapısı". FEMS Mikrobiyoloji İncelemeleri. 25 (2): 147–74. doi:10.1111 / j.1574-6976.2001.tb00575.x. PMID 11250034.
- ^ "Archaeal flagellum". www.uniprot.org. Alındı 24 Haziran 2019.
- ^ Berg HC (2003). E. coli hareket halinde (1. Aufl. Ed.). New York: Springer. ISBN 9780387008882.
- ^ Taylor FJ (Kasım 2003). "İki krallık sisteminin çöküşü, protistolojinin yükselişi ve Uluslararası Evrimsel Protistoloji Derneği'nin (ISEP) kuruluşu". Uluslararası Sistematik ve Evrimsel Mikrobiyoloji Dergisi. 53 (Pt 6): 1707–14. doi:10.1099 / ijs.0.02587-0. PMID 14657097.
- ^ Hülsmann N (Ağustos 1992). "Undulipodium: Yararsız bir tartışmanın sonu". Avrupa Protistoloji Dergisi. 28 (3): 253–7. doi:10.1016 / s0932-4739 (11) 80231-2. PMID 23195228.
- ^ a b c Adl SM, Simpson AG, Lane CE, Lukeš J, Bass D, Bowser SS, ve diğerleri. (Eylül 2012). "Ökaryotların gözden geçirilmiş sınıflandırması". Ökaryotik Mikrobiyoloji Dergisi. 59 (5): 429–93. doi:10.1111 / j.1550-7408.2012.00644.x. PMC 3483872. PMID 23020233.
- ^ Andersen RA, Barr DJ, Lynn DH, Melkonian M, Moestrup Ø, Sleigh MA (1991). "Protistlerdeki flagellar / siliyer aparatlarla ilişkili hücre iskeleti elemanlarının terminolojisi ve adlandırılması". Protoplazma. 164 (1–3): 1–8. doi:10.1007 / bf01320809. S2CID 40755371.
- ^ Leadbeater, Barry S. C .; Green, John C., eds. (2000). Flagellates: Birlik, Çeşitlilik ve Evrim. Sistematik Derneği Özel Cilt. 59. Taylor ve Francis. ISBN 978-1-4822-6822-5.
- ^ a b Barsanti L, Gualtieri P (2006). Algler: Anatomi, Biyokimya ve Biyoteknoloji. Florida, ABD: CRC Press. ISBN 9780203492598.
- ^ Pazour GJ (Ekim 2004). "İntraflagellar taşıma ve siliyaya bağlı böbrek hastalığı: polikistik böbrek hastalığının siliyer hipotezi". Amerikan Nefroloji Derneği Dergisi. 15 (10): 2528–36. doi:10.1097 / 01.ASN.0000141055.57643.E0. PMID 15466257.
- ^ Yubuki N, Leander BS (Temmuz 2013). "Ökaryot ağacı boyunca mikro tüp düzenleme merkezlerinin evrimi". Bitki Dergisi. 75 (2): 230–44. doi:10.1111 / tpj.12145. PMID 23398214.
- ^ Raven, J.A. (2000). "Kamçılı durum". Leadbeater & Green 2000, s. 27–48. ISBN 9781482268225.
- ^ a b Webster J, Weber R (25 Ocak 2007). "Mantar Sporları". 2007 (3. baskı). Cambridge: Cambridge University Press. s. 23–24. ISBN 9781139461504.
- ^ Lahr DJ, Parfrey LW, Mitchell EA, Katz LA, Lara E (Temmuz 2011). "Amiplerin iffeti: amipli organizmalarda cinsiyet için kanıtların yeniden değerlendirilmesi". Bildiriler. Biyolojik Bilimler. 278 (1715): 2081–90. doi:10.1098 / rspb.2011.0289. PMC 3107637. PMID 21429931.
- ^ a b Austin CR (1995). "İnsan gametlerinin evrimi: spermatozoa.". Grudzinskas JG, Yovich JL (editörler). Gametes: spermatozoon. Cambridge University Press. ISBN 9780521479967.
- ^ South GR, Whittick A (1987). Introduction to Phycology. Oxford: Blackwell Scientific Publications. s. 65. ISBN 9781444314205.
- ^ Dodge JD (1973). The Fine Structure of Algal Cells. Londra: Akademik Basın. pp. 57–79. ISBN 9780323158237.
- ^ Lee RE (2008). Psikoloji (4. baskı). Cambridge University Press. s.7. ISBN 9781139469876.
lee tubular hairs.
- ^ Corliss, J.O .; Lom, J (2000). "An annotated glossary of protozoological terms". In Lee, J.J.; Leedale, G.F.; Bradbury, P. (eds.). An illustrated guide to the protozoa. 2 (2. baskı). Protozoologlar Derneği. pp. 1346–85. ISBN 1891276239.
- ^ a b Jeuck A, Arndt H (November 2013). "A short guide to common heterotrophic flagellates of freshwater habitats based on the morphology of living organisms". Protist. 164 (6): 842–60. doi:10.1016/j.protis.2013.08.003. PMID 24239731.
- ^ Sleigh M (1989). Protozoa and other Protists. Londra: Edward Arnold. s. 98–99. ISBN 9780521428057.
- ^ Sparrow FK (1960). Aquatic phycomycetes (2. baskı). Ann Arbor: Michigan: University of Michigan Press. s.15.
- ^ Hibberd DJ (1976). "The ultrastructure and taxonomy of the Chrysophyceae and Prymnesiophyceae (Haptophyceae): a survey with some new observations on the ultrastructure of the Chrysophyceae". Londra Linnean Derneği Dergisi, Botanik. 72 (2): 55–80. doi:10.1111/j.1095-8339.1976.tb01352.x.
- ^ Sleigh MA (1985). "Origin and evolution of flagellar movement". Cell Motil. 5: 137–138.
daha fazla okuma
- Berg HC (January 2000). "Motile Behavior of Bacteria". Bugün Fizik. 53 (1): 24–29. Bibcode:2000PhT....53a..24B. doi:10.1063/1.882934. Arşivlenen orijinal 15 Nisan 2013.
- Lindemann C (4 April 2008). "Mechanisms of sperm motility". Oakland Üniversitesi. Arşivlenen orijinal 16 Mayıs 2008. Alındı 18 Mayıs 2008.
- Purcell EM (1977). "Life at Low Reynolds Number" (PDF). Amerikan Fizik Dergisi. 45 (1): 3–11. Bibcode:1977AmJPh..45 .... 3P. doi:10.1119/1.10903. hdl:2433/226838. Arşivlenen orijinal (PDF) 5 Haziran 2011'de. Alındı 19 Ekim 2009.
- Matzke NJ (10 November 2003). "Evolution in (Brownian) space: a model for the origin of the bacterial flagellum". talkdesign.org.
Dış bağlantılar

![]() Bu makale şu anda web sitesinde bulunan bir yayından metin içermektedir. kamu malı: Chambers, Ephraim, ed. (1728). Cyclopædia veya Evrensel Sanat ve Bilim Sözlüğü (1. baskı). James ve John Knapton, vd. Eksik veya boş
Bu makale şu anda web sitesinde bulunan bir yayından metin içermektedir. kamu malı: Chambers, Ephraim, ed. (1728). Cyclopædia veya Evrensel Sanat ve Bilim Sözlüğü (1. baskı). James ve John Knapton, vd. Eksik veya boş | title = (Yardım)