Krill - Krill
| Krill | |
|---|---|
 | |
| Kuzey kril (Meganyctiphanes norvegica) | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Arthropoda |
| Altfilum: | Kabuklular |
| Sınıf: | Malacostraca |
| Üst sipariş: | Eucarida |
| Sipariş: | Euphausiacea Dana, 1852 |
| Aileler ve cinsler | |
| |
Krill küçükler kabuklular of sipariş Euphausiaceave dünyanın tüm okyanuslarında bulunur. "Kril" adı, Norveççe kelime kril, "küçük kızartmak balık ",[1] bu da genellikle balık türlerine atfedilir.
Krill önemli bir tropik seviye bağlantı - alt tarafına yakın besin zinciri. Besleniyorlar fitoplankton ve (daha az ölçüde) Zooplankton ama aynı zamanda birçok büyük hayvan için ana besin kaynağıdır. İçinde Güney okyanus, bir tür, Antarktik kril, Euphausia superba, bir tahmin oluşturur biyokütle yaklaşık 379.000.000 ton[2] en büyük toplam biyokütleye sahip türler arasında yer alıyor. Bu biyokütlenin yarısından fazlası her yıl balinalar, foklar, penguenler, kalamar ve balıklar tarafından yenir. Çoğu kril türü büyük görünür günlük dikey geçişler Böylece gece yüzeye yakın ve gün boyunca daha derin sularda avcılar için yiyecek sağlar.
Krill, ticari olarak Güney Okyanusunda ve Japonya çevresindeki sularda avlanır. Toplam küresel hasat miktarı yılda 150.000–200.000 tondur ve bunun çoğu Scotia Denizi. Krill avının çoğu için kullanılır su kültürü ve akvaryum gibi beslemeler yem içinde balıkçılık veya ilaç endüstrisinde. Japonya, Filipinler ve Rusya'da kril aynı zamanda insan tüketimi için de kullanılmaktadır ve şu şekilde bilinmektedir: Okiami (オ キ ア ミ) Japonyada. Olarak yenirler camarones İspanya ve Filipinler'de. Filipinler'de kril aynı zamanda Alamang ve adı verilen tuzlu bir macun yapmak için kullanılır Bagoong.
Krill ayrıca ana avıdır. balenli balinalar, I dahil ederek Mavi balina.
Taksonomi
Krill büyüklere aittir eklem bacaklı alt filum, Kabuklular. En tanıdık ve en büyük kabuklular grubu, sınıf Malacostraca içerir üst sipariş Eucarida üç sipariş içeren Euphausiacea (kril), Decapoda (karides, karides, ıstakoz, yengeç) ve planktonik Amphionidacea.
Euphausiacea düzeni iki aileler. Daha bol Euphausiidae 10 farklı içerir cins toplam 85 tür ile. Bunlardan cins Euphausia 31 türle en büyüğüdür.[3] Daha az bilinen aile, Bentheuphausiidae, sadece bir tane var Türler, Bentheuphausia ambliyops, bir banyo havası 1.000 m'nin (3.300 ft) altındaki derin sularda yaşayan kril. Günümüze ulaşan en ilkel kril türü olarak kabul edilir.[4]
Ticari Euphausiidae'nin tanınmış türleri krill balıkçılığı Dahil etmek Antarktik kril (Euphausia superba), Pasifik kril (E. pacifica) ve Kuzey kril (Meganyctiphanes norvegica).[5]
Filogeni
| Euphausiacea'nın önerilen soyoluşu[6] | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Morfolojik verilerden elde edilen soyoluş, (♠) isimler,[6] (♣) muhtemelen parafiletik takson Nematobrachion içinde.[6] (♦) sınıfları Casanova'dan (1984) farklıdır,[7] nerede Pseudoeuphausia kız kardeşi Nyctiphanes, Euphausia kız kardeşi Thysanopoda ve Nematobrachion kız kardeşi Stylocheiron. |
2013 itibarıyla[Güncelleme], Euphausiacea düzeninin monofiletik birkaç benzersiz korunmuş morfolojik özellik nedeniyle (otapomorfi ) çıplak ipliksi solungaçları ve ince torakopodlar gibi[8] ve moleküler çalışmalarla.[9][10][11]
Euphausiacea takımının yeri hakkında birçok teori vardır. İlk tanımından beri Thysanopode triküspid tarafından Henri Milne-Edwards 1830'da, biramöz torakopodlarının benzerliği, zoologları sırayla euphausiids ve Mysidacea'yı gruplandırmaya yöneltti. Şizopoda bölünmüş olan Johan Erik Vesti Boas 1883'te iki ayrı sipariş halinde.[12] Sonra, William Thomas Calman (1904), Mysidacea üst siparişte Peracarida ve süper takımdaki euphausiids Eucarida 1930'lara kadar bile Schizopoda emri savunuldu.[8] Daha sonra, Euphausiacea düzeninin şu grupla gruplandırılması önerildi: Penaeidae (karides ailesi) Decapoda'da gelişimsel benzerliklere dayanmaktadır. Robert Gurney ve Isabella Gordon.[13][14] Bu tartışmanın nedeni, krillerin on ayaklıların ve diğer mysidlerin bazı morfolojik özelliklerini paylaşmasıdır.[8]
Moleküler çalışmalar, muhtemelen aşağıdakiler gibi anahtar nadir türlerin azlığı nedeniyle, onları açık bir şekilde gruplamamıştır. Bentheuphausia ambliyops kril ve Amfiyonidler reynaudii Eucarida'da. Bir çalışma, Eucarida'nın monofilini (bazal Mysida ile) destekler,[15] Mysida (Schizopoda) ile başka bir grup Euphausiacea,[10] Yine başka bir grup Euphausiacea ile Hoplocarida.[16]
Zaman çizelgesi
Euphausiacea'ya hiçbir mevcut fosil kesin olarak atanamaz. Bazıları tükenmiş Eumalacostracan takson euphausiaceans olduğu düşünülmüştür. Antrakofoz, Crangopsis - şimdi atandı Aeschronectida (Hoplocarida)[6] - ve Paleomiz.[17] Tüm flört türleşme olaylar tarafından tahmin edildi moleküler saat krill ailesinin son ortak atasını yerleştiren yöntemler, Euphausiidae (sipariş Euphausiacea eksi Bentheuphausia ambliyops) içinde yaşamış olmak Alt Kretase hakkında 130 milyon yıl önce.[10]
Dağıtım
Krill, dünya çapında tüm okyanuslarda görülse de, pek çok tür endemik veya neritik (yani kıyı) dağılımları. Bentheuphausia ambliyops, bir banyo havası türler, vardır kozmopolit dağılım derin deniz yaşam alanı içinde.[18]
Cinsin türleri Thysanoessa ikisinde de meydana gelir Atlantik ve Pasifik okyanuslar.[19] Pasifik ev sahipliği yapmaktadır Euphausia pacifica. Kuzey krili, Atlantik boyunca Akdeniz kuzeye.
Neritik dağılımlara sahip türler, cinsin dört türünü içerir Nyctiphanes.[20] Boyunca bol miktarda bulunurlar yükselen bölgeleri Kaliforniya, Humboldt, Benguela, ve Kanarya mevcut sistemler.[21][22][23] Sadece neritik dağılıma sahip başka bir tür E. crystalorophiasAntarktika kıyı şeridine endemik olan.[24]
Endemik dağılımlara sahip türler şunları içerir: Nyctiphanes capensis, yalnızca Benguela akımında meydana gelen,[20] E. mucronata Humboldt akımında,[25] ve altı Euphausia Güney Okyanusu'na özgü türler.
Antarktika'da yedi tür biliniyor,[26] cinste bir Thysanoessa (T. macrura ) ve altı inç Euphausia. Antarktik kril (Euphausia superba) genellikle 100 m'ye (330 ft) ulaşan derinliklerde yaşar,[27] oysa buz kril (Euphausia crystallorophias ) 4.000 m (13.100 ft) derinliğe ulaşır, ancak bunlar genellikle en fazla 300-600 m (1.000-2.000 ft) derinliklerde yaşarlar.[28] İkisi de şurada bulunur enlemler güneyi 55 ° G, ile E. crystalorophias güneye hakim 74 ° G[29] ve bölgelerinde buz paketi. Bilinen diğer türler Güney okyanus vardır E. frigida, E. longirostris, E. triacantha ve E. vallentini.[30]
Anatomi ve morfoloji
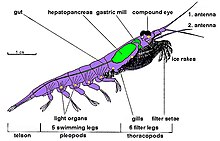

Krill vardır on ayaklı kabuklular ve tüm kabuklular gibi cılız dış iskelet. Standartları var dekapod anatomisi üç vücuttan oluşan parçalar: sefalotoraks ile oluşan baş ve göğüs kaynaşmış pleon on yüzme ayağı taşıyan ve kuyruk pervanesi. Krilin bu dış kabuğu çoğu türde şeffaftır.
Krill özelliği karmaşık Bileşik gözler. Bazı türler, tarama yoluyla farklı aydınlatma koşullarına uyum sağlar pigmentler.[31]
İki tane var anten ve birkaç çift göğüs bacağı denilen pereiopodlar veya Torakopodlar, toraksa bağlı oldukları için bu şekilde adlandırılmıştır. Sayıları cins ve tür arasında değişir. Bu torasik bacaklar arasında beslenme bacakları ve bakım bacakları bulunur.
Krilller dekopodlardır, bu nedenle tüm türlerde beş çift vardır yüzme bacakları "yüzücü" olarak adlandırılan, Istakoz veya tatlı su kereviti.
Çoğu kril yetişkinler kadar yaklaşık 1-2 santimetredir (0.4-0.8 inç). Birkaç tür, 6–15 santimetre (2,4–5,9 inç) düzeyinde büyür. En büyük kril türü, Thysanopoda spinicauda, hayatları açık okyanusun derinliklerinde.[32] Krill, gerçek gibi diğer kabuklulardan kolayca ayırt edilebilir. karides dışarıdan görülerek solungaçlar.[33]
Dışında Bentheuphausia ambliyops, kril biyolüminesan organları olan hayvanlar denilen fotoforlar ışık yayabilir. Işık, bir enzim katalize edilmiş kemilüminesans reaksiyon, burada a lusiferin (bir tür pigment) bir lusiferaz enzim. Araştırmalar, birçok kril türünün lusiferininin floresan tetrapirol benzer ama aynı değil dinoflagellat lusiferin[34] ve kril muhtemelen bu maddeyi kendilerinin üretmediğini, ancak dinoflagellat içeren diyetlerinin bir parçası olarak aldıklarını söylüyor.[35] Krill fotoforları, lenslere ve odaklanma yeteneklerine sahip karmaşık organlardır ve kaslar tarafından döndürülebilir.[36] Bu organların kesin işlevi henüz bilinmemektedir; olasılıklar arasında çiftleşme, sosyal etkileşim veya oryantasyon ve gölgelerini üstteki ortam ışığına karşı telafi etmek için bir tür karşı aydınlatma kamuflajı sayılabilir.[37][38]
Ekoloji

Besleme
Birçok kril Filtre besleyicileri:[22] onların en önde ekler Torakopodlar, yiyeceklerini sudan filtreleyebilecekleri çok ince taraklar oluştururlar. Bu filtreler türlerde çok iyi olabilir (örneğin Euphausia spp.) öncelikle beslenen fitoplankton özellikle diyatomlar, tek hücreli yosun. Krill çoğunlukla her yerde yaşayan,[40] birkaç tür olmasına rağmen etobur, küçük avlamak Zooplankton ve balık larvalar.[41]
Krill, sudaki önemli bir unsurdur. besin zinciri. Krill birincil üretim avlarının küçük alglerle doğrudan beslenemeyen daha büyük hayvanlar tarafından tüketilmeye uygun bir biçime dönüştürülmesini sağlar. Kuzey kril ve diğer bazı türler nispeten küçük bir filtre sepetine sahiptir ve aktif olarak avlanır kopepodlar ve daha büyük zooplankton.[41]
Predasyon
Birçok hayvan kril ile beslenir, daha küçük hayvanlardan balık veya penguenler gibi daha büyük olanlara mühürler ve balenli balinalar.[42]
Bir rahatsızlık ekosistem Krill popülasyonunda bir düşüşe neden olan, geniş kapsamlı etkilere sahip olabilir. Bir kokolitofor çiçek açmak Bering Denizi 1998 yılında,[43] örneğin, etkilenen bölgedeki diyatom konsantrasyonu düştü. Krill daha küçük kokolitoforlarla ve dolayısıyla kril popülasyonuyla (esas olarak E. pacifica) o bölgede keskin bir düşüş yaşadı. Bu da diğer türleri etkiledi: yelkovan nüfus düştü. Olayın bir nedeni olduğu düşünülüyordu Somon o sezon ortaya çıkmadı.[44]
Birkaç tek hücreli endoparasitoidik siliatlar cinsin Collinia kril türlerini enfekte edebilir ve etkilenen popülasyonları tahrip edebilir. Bu tür hastalıklar rapor edildi Thysanoessa inermis Bering Denizi'nde ve ayrıca E. pacifica, Thysanoessa spinifera, ve T. gregaria Kuzey Amerika Pasifik kıyılarının açıklarında.[45][46] Biraz ektoparazitler ailenin Dajidae (epikaridean izopodlar ) kril (ve ayrıca karides ve mysids ); böyle bir parazit Oculophryxus bicaulis, kril üzerinde bulunan Stylocheiron afin ve S. longicorne. Kendini hayvanın gözüne tutturur ve kafasından kan emer; etkilenen hayvanların hiçbiri olgunlaşmadığından, görünüşe göre konakçının üremesini engeller.[47]
İklim değişikliği kril popülasyonları için başka bir tehdit oluşturmaktadır.[48]
Plastikler
Ön araştırma, krilin sindirebileceğini gösteriyor mikroplastikler 5 mm (0,20 inç) çapın altında, onları parçalara ayırır ve daha küçük formda çevreye geri verir.[49]
Yaşam öyküsü ve davranış

Krilin yaşam döngüsü, türden türe küçük ayrıntılara rağmen nispeten iyi anlaşılmıştır.[13][22] Krill yumurtadan çıktıktan sonra birkaç larva aşaması yaşarlar.Nauplius, psödometanauplius, Metanauplius, kaliptoplar, ve Furcilia, her biri alt aşamalara ayrılıyor. Pseudometanauplius aşaması, yumurtalarını yumurtalık bir kese içine bırakan türlere özeldir: "kese yumurtlayanlar" olarak adlandırılanlar. Larvalar büyür ve tüy dökmek geliştikçe tekrar tekrar, sert dış iskelet çok küçük olduğunda yerini alıyor. Küçük hayvanlar büyük olanlara göre daha sık tüy dökerler. Yumurta sarısı vücutlarındaki rezervler larvaları metanauplius evresiyle besler.
Calyptopsis aşamalarına göre farklılaşma bir ağız ve sindirim sistemi geliştirecek kadar ilerlemiştir ve fitoplankton yemeye başlarlar. O zamana kadar yumurta sarısı rezervleri tükenir ve larvalar, fotik bölge, okyanusun alglerin geliştiği üst katmanları. Furcilia aşamaları sırasında, en öndeki bölümlerden başlayarak yüzücü çiftlerinin bulunduğu bölümler eklenir. Her yeni çift yalnızca bir sonraki tüy dökümü sırasında işlevsel hale gelir. Furcilia aşamalarından herhangi biri sırasında eklenen segment sayısı, çevresel koşullara bağlı olarak bir tür içinde bile değişebilir.[50] Son furcilia aşamasından sonra, olgunlaşmamış bir genç, bir yetişkine benzer bir şekilde ortaya çıkar ve daha sonra gelişir. gonadlar ve cinsel olarak olgunlaşır.[51]
Üreme

Türe ve iklime göre değişen çiftleşme mevsimi boyunca, erkek bir sperm çuvalı dişinin genital açıklığında (adı Thelycum). Dişiler birkaç bin yumurta taşıyabilir. yumurtalık Bu, hayvanın vücut kütlesinin üçte birini oluşturabilir.[52] Krill, bir sezonda birden fazla yavruya sahip olabilir ve kuluçka aralıkları günler sırasına göre devam eder.[23][53]
Krill, iki tür yumurtlama mekanizması kullanır.[23] Cinsin 57 türü Bentheuphausia, Euphausia, Meganyctiphanes, Thysanoessa, ve Thysanopoda "yumurtlayan yumurtalar" dır: dişi döllenmiş yumurtaları suya bırakır, burada genellikle batarlar, dağılırlar ve kendi başlarına kalırlar. Bu türler genellikle nauplius 1 aşamasında yumurtadan çıkar, ancak son zamanlarda bazen metanauplius veya hatta calyptopis aşamaları olarak yumurtadan çıktıkları keşfedilmiştir.[54] Diğer cinslerin kalan 29 türü, dişinin yumurtaları yanında taşıdığı ve metanauplii olarak yumurtadan çıkana kadar en arkadaki torakopod çiftlerine bağlı olduğu "keseli yumurtlayanlar" dır. Nematoscelis difficilis nauplius veya psödometanauplius olarak yumurtadan çıkabilir.[55]
Tüy dökme
Tüy dökme, bir numune sert dış iskeletini aştığında meydana gelir. Daha hızlı büyüyen genç hayvanlar, daha yaşlı ve büyük olanlara göre daha sık tüy dökerler. Tüy dökme sıklığı türe göre büyük ölçüde değişir ve tek bir tür içinde bile enlem, su sıcaklığı ve yiyecek bulunabilirliği gibi birçok dış faktöre tabidir. Subtropikal türler Nyctiphanes simplexörneğin, iki ila yedi günlük bir toplam tüyler arası süreye sahiptir: larvalar ortalama dört günde bir tüy dökerken, gençler ve yetişkinler bunu ortalama altı günde bir yapar. İçin E. superba Antarktik denizinde −1 ile 4 ° C (30 ve 39 ° F) arasındaki sıcaklığa bağlı olarak 9 ile 28 gün arasında değişen tüy dökme dönemleri gözlemlenmiştir ve Meganyctiphanes norvegica içinde Kuzey Denizi tüy dökme dönemleri de 9 ila 28 gün arasında değişir, ancak 2.5 ila 15 ° C (36.5 ila 59.0 ° F) arasındaki sıcaklıklarda.[56] E. superba yeterli yiyecek bulunmadığında vücut boyutunu küçültebilir, dış iskeleti çok büyüdüğünde de tüy dökebilir.[57] Benzer bir küçülme de gözlenmiştir. E. pacificaPasifik Okyanusu'nda, anormal derecede yüksek su sıcaklıklarına adaptasyon olarak kutuplardan ılıman bölgelere uzanan bir tür. Büzülme, diğer ılıman bölge kril türleri için de kabul edilmiştir.[58]
Ömür
Bazı yüksek enlemli kril türleri altı yıldan fazla yaşayabilir (ör. Euphausia superba); orta enlem türleri gibi diğerleri Euphausia pacifica, sadece iki yıl yaşa.[5] Subtropikal veya tropikal türlerin ömrü hala daha kısadır, ör. Nyctiphanes simplex, genellikle sadece altı ila sekiz ay yaşar.[59]
Kaynıyor
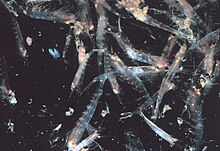
Çoğu kril kaynaşma hayvanlar; Bu tür sürülerin boyutları ve yoğunlukları türlere ve bölgelere göre değişir. İçin Euphausia superbasürüler metreküp başına 10.000 ila 60.000 kişiye ulaşır.[60][61] Sürü, bireyleri seçmek isteyen daha küçük avcıların kafasını karıştıran savunma mekanizmasıdır. 2012'de Gandomi ve Alavi, bir başarılı stokastik algoritma kril sürülerinin davranışını modellemek için. Algoritma üç ana faktöre dayanmaktadır: "(i) diğer bireylerin varlığından kaynaklanan hareket (ii) yiyecek arama aktivitesi ve (iii) rastgele yayılma."[62]
Dikey geçiş

Krill tipik olarak bir günlük dikey göç. Günü daha derinlerde geçirdikleri ve gece yüzeye doğru yükseldikleri varsayılmıştır. Ne kadar derine giderlerse, aktivitelerini o kadar azaltırlar,[63] görünüşe göre avcılarla karşılaşmaları azaltmak ve enerji tasarrufu yapmak için. Krillde yüzme aktivitesi mide dolgunluğuna göre değişir. Yüzeyde beslenen doymuş hayvanlar daha az aktif olarak yüzerler ve bu nedenle karışık tabakanın altına batarlar.[64] Batarken üretirler dışkı bu Antarktika'da bir rolü ima eder karbon döngüsü. Boş mideli Krill daha aktif yüzer ve böylece yüzeye doğru yönelir.
Dikey göç, günde 2-3 kez meydana gelebilir. Bazı türler (ör. Euphausia superba, E. pacifica, E. hanseni, Pseudeuphausia latifrons, ve Thysanoessa spinifera) gün boyunca beslenme ve üreme amaçlı yüzey sürüleri oluştururlar, bu davranış tehlikeli olsa da onları avcılara karşı son derece savunmasız hale getirir.[65]
Kullanarak deneysel çalışmalar Artemia salina bir model olarak, krillerin onlarca metre derinliğindeki gruplar halinde birkaç yüz metrelik dikey göçlerinin, toplu olarak okyanus karışımı üzerinde önemli bir etkiye sahip olmak için yeterli miktarda aşağı doğru su jeti oluşturabileceğini öne sürüyor.[66]
Yoğun sürüler bir aşırı beslenme balıklar, kuşlar ve memeli yırtıcı hayvanlar arasında, özellikle yüzeye yakın. Rahatsız edildiğinde bir sürü dağılır ve hatta bazı bireylerin anında tüy döktüğü gözlemlenmiştir. exuvia arkasında bir yem olarak.[67]
Krill normalde 5–10 cm / s (saniyede 2-3 vücut uzunluğu) hızla yüzer,[68] yüzücülerini tahrik için kullanmak. Büyük göçleri okyanus akıntılarına tabidir. Tehlikede olduklarında, bir kaçış tepkisi aranan ıstakoz - flicking onların kuyruk yapılar, telson ve üropodlar, suda nispeten hızlı bir şekilde geriye doğru hareket ederek, saniyede 10 ila 27 vücut uzunluğu aralığında hızlara ulaşırlar; E. superba yaklaşık 0,8 m / s (3 ft / s) anlamına gelir.[69] Yüzme performansları, birçok araştırmacının yetişkin krilleri şu şekilde sınıflandırmasına neden olmuştur: mikro nektonik yaşam formları, yani (zayıf) akımlara karşı bireysel hareket edebilen küçük hayvanlar. Krill'in larva formları genellikle zooplankton olarak kabul edilir.[70]
Biyojeokimyasal çevrimler

Antarktik kril, bağlamında önemli bir türdür. biyojeokimyasal döngü[71][39] Ve içinde Antarktika besin ağı.[72][73] Yeteneğinden dolayı Güney Okyanusunda önemli bir rol oynar. döngü besinleri ve penguenleri beslemek ve balya ve Mavi balinalar.

İnsan kullanır

Hasat tarihi
Krill, en azından 19. yüzyıldan beri ve muhtemelen daha önce Japonya'da insanlar ve evcil hayvanlar için bir besin kaynağı olarak hasat edilmiştir. Okiami. Büyük ölçekli balıkçılık, 1960'ların sonlarında ve 1970'lerin başlarında gelişti ve şimdi yalnızca Antarktika sularında ve Japonya çevresindeki denizlerde gerçekleşiyor. Tarihsel olarak, en büyük kril balıkçılığı ülkeleri Japonya ve Sovyetler Birliği idi ya da Sovyetler Birliği'nin dağılmasından sonra, Rusya ve Ukrayna.[74] 1983'te yalnızca Güney Okyanusu'nda yaklaşık 528.000 ton olan hasat zirveye ulaştı (bunun% 93'ünü Sovyetler Birliği aldı), şimdi aşırı avlanmaya karşı bir önlem olarak yönetiliyor.[75]
1993'te, iki olay kril avcılığının azalmasına neden oldu: Rusya sektörden çıktı; ve Antarktika Deniz Yaşayan Kaynaklarının Korunması Sözleşmesi (CCAMLR) bir için maksimum yakalama kotalarını tanımladı sürdürülebilir sömürü Antarktika krili. Ekim 2011 tarihli bir incelemeden sonra, Komisyon kotayı değiştirmemeye karar verdi.[76]
Yıllık Antarktika avı, CCAMLR av kotasının kabaca ellide biri olan 100.000 ton civarında sabitlendi.[77] Ana sınırlayıcı faktör muhtemelen siyasi ve yasal sorunların yanı sıra yüksek maliyetlerdi.[78] Japon balıkçılığı 70.000 tonda doydu.[79]
Kril dünya çapında bulunmasına rağmen, Güney Okyanuslarda balıkçılık tercih edilir çünkü kril bu bölgelerde daha "yakalanabilir" ve bol miktarda bulunur. Özellikle Antarktika denizlerinde bozulmamış "temiz ürün" olarak kabul edilirler.[74]
2018'de Antarktika'da faaliyet gösteren hemen hemen her kril balıkçılık şirketinin 2020'den itibaren Antarktika Yarımadası'nın etrafındaki devasa alanlardaki operasyonlarını, penguen üreme kolonilerinin etrafındaki "tampon bölgeler" de dahil olmak üzere terk edeceği açıklandı.[80]
İnsan tüketimi

Toplam olmasına rağmen biyokütle Antarktika krilinin yüzdesi 400 milyon kadar olabilir ton, bunun üzerindeki insan etkisi kilit taşı türleri büyüyor, toplam balıkçılık veriminde% 39 artışla 2010-2014 arasında 294.000 tona yükseliyor.[77] Krill hasadına dahil olan başlıca ülkeler Norveç (2014 yılında toplam avlanmanın% 56'sı), Kore Cumhuriyeti (% 19) ve Çin (18%).[77]
Krill zengin bir kaynaktır protein ve Omega-3 yağlı asitler erken dönemde geliştirilmekte olan 21'inci yüzyıl insan yemeği olarak diyet takviyeleri yağ kapsülleri olarak, çiftlik hayvanları yiyecek ve Evcil Hayvan gıda.[74][76][81] Krill, karidesten biraz daha güçlü bir balık aromasıyla tuzlu tadı verir. Toplu tüketim ve ticari olarak hazırlanmış ürünler için, yenmeyenleri çıkarmak için soyulmaları gerekir. dış iskelet.[81]
2011'de ABD Gıda ve İlaç İdaresi üretilmiş bir itirazsız mektup yayınladı Krill yağı ürün olmak genellikle güvenli olarak kabul edilir (GRAS ) insan tüketimi için.[82]
Krill (ve diğerleri planktonik karides ) en çok tüketilen Güneydoğu Asya, nerede fermente (kabuklar bozulmadan) ve genellikle ince öğütülmüş Karides ezmesi. Tavada kızartılıp beyaz pirinçle birlikte yenebilir veya eklemek için kullanılabilir. Umami çok çeşitli geleneksel yemeklere tatlar.[83][84] Fermantasyon sürecinden elde edilen sıvı da aynı şekilde hasat edilir. balık sosu.[85]
Ayrıca bakınız
 Kabuklular portalı
Kabuklular portalı- Soğuk su karidesi
Referanslar
- ^ "Krill". Çevrimiçi Etimoloji Sözlüğü. Alındı 22 Haziran 2010.
- ^ A. Atkinson; V. Siegel; E.A. Pakhomov; M.J. Jessopp; V. Loeb (2009). "Toplam biyokütlenin ve yıllık Antarktik kril üretiminin yeniden değerlendirilmesi" (PDF). Derin Deniz Araştırmaları Bölüm I. 56 (5): 727–740. Bibcode:2009DSRI ... 56..727A. doi:10.1016 / j.dsr.2008.12.007.
- ^ Volker Siegel (2011). Siegel V (ed.). "Euphausiidae Dana, 1852". Dünya Euphausiacea veritabanı. Dünya Deniz Türleri Kaydı. Alındı 25 Kasım 2011.
- ^ E. Brinton (1962). "Pasifik euphausiidlerinin dağılımı". Boğa. Scripps Inst. Oceanogr. 8 (2): 51–270.
- ^ a b S. Nicol; Y. Endo (1999). "Krill balıkçılık: Geliştirme, yönetim ve ekosistem etkileri". Sucul Yaşam Kaynakları. 12 (2): 105–120. doi:10.1016 / S0990-7440 (99) 80020-5.
- ^ a b c d Andreas Maas; Dieter Waloszek (2001). "Larva gelişimi Euphausia superba Dana, 1852 ve Euphausiacea'nın filogenetik analizi " (PDF). Hidrobiyoloji. 448: 143–169. doi:10.1023 / A: 1017549321961. S2CID 32997380. Arşivlenen orijinal (PDF) 2011-07-18 tarihinde.
- ^ Bernadette Casanova (1984). "Phylogénie des Euphausiacés (Crustacés Eucarides)" [Phylogeny of the Euphausiacea (Crustacea: Eucarida)]. Bulletin du Muséum National d'Histoire Naturelle (Fransızcada). 4: 1077–1089.
- ^ a b c Bernadette Casanova (2003). "Ordre des Euphausiacea Dana, 1852". Crustaceana. 76 (9): 1083–1121. doi:10.1163/156854003322753439. JSTOR 20105650.
- ^ M. Eugenia D'Amato; Gordon W. Harkins; Tulio de Oliveira; Peter R. Teske; Mark J. Gibbons (2008). "Neritik krilin moleküler tarihlemesi ve biyocoğrafyası Nyctiphanes" (PDF). Deniz Biyolojisi. 155 (2): 243–247. doi:10.1007 / s00227-008-1005-0. S2CID 17750015.
- ^ a b c Simon N. Jarman (2001). "Krill'in evrimsel tarihi, nükleer büyük alt birim rDNA dizi analizinden çıkarılmıştır". Linnean Society Biyolojik Dergisi. 73 (2): 199–212. doi:10.1111 / j.1095-8312.2001.tb01357.x.
- ^ Xin Shen; Haiqing Wang; Minxiao Wang; Bin Liu (2011). "Tam mitokondriyal genom dizisi Euphausia pacifica (Malacostraca: Euphausiacea) yeni bir gen düzenini ve alışılmadık tandem tekrarlarını ortaya koyuyor ". Genetik şifre. 54 (11): 911–922. doi:10.1139 / g11-053. PMID 22017501.
- ^ Johan Erik Vesti Boas (1883). "Studien über die Verwandtschaftsbeziehungen der Malacostraken" [Malacostraca ilişkileri üzerine çalışmalar]. Morfoloji Jahrbuch (Almanca'da). 8: 485–579.
- ^ a b Robert Gurney (1942). Decapod Crustacea Larvaları (PDF). Ray Topluluğu.
- ^ Isabella Gordon (1955). "Euphausiacea'nın sistematik konumu". Doğa. 176 (4489): 934. Bibcode:1955Natur.176..934G. doi:10.1038 / 176934a0. S2CID 4225121.
- ^ Trisha Spears, Ronald W. DeBry, Lawrence G. Abele & Katarzyna Chodyl (2005). Boyko, Christopher B. (ed.). "Perakarid monofili ve interordinal filogeni, nükleer küçük alt birim ribozomal DNA dizilerinden çıkarılmıştır (Crustacea: Malacostraca: Peracarida)" (PDF). Washington Biyoloji Derneği Tutanakları. 118 (1): 117–157. doi:10.2988 / 0006-324X (2005) 118 [117: PMAIPI] 2.0.CO; 2.CS1 bakimi: birden çok ad: yazarlar listesi (bağlantı)
- ^ K. Meland; E. Willassen (2007). "Mysidacea" nın (Crustacea) ayrılığı ". Moleküler Filogenetik ve Evrim. 44 (3): 1083–1104. CiteSeerX 10.1.1.653.5935. doi:10.1016 / j.ympev.2007.02.009. PMID 17398121.
- ^ Frederick R. Schram (1986). Kabuklular. Oxford University Press. ISBN 978-0-19-503742-5.
- ^ J. J. Torres; J. J. Childress (1985). "Batipelajik euphausiid'in solunumu ve kimyasal bileşimi Bentheuphausia ambliyops". Deniz Biyolojisi. 87 (3): 267–272. doi:10.1007 / BF00397804. S2CID 84486097.
- ^ Volker Siegel (2011). "Thysanoessa Brandt, 1851 ". WoRMS. Dünya Deniz Türleri Kaydı. Alındı 18 Haziran 2011.
- ^ a b D'Amato, M.E. et al.: "Neritik krilin moleküler tarihlemesi ve biyocoğrafyası Nyctiphanes ", içinde Deniz Biyolojisi cilt. 155, hayır. 2, s. 243-247, Ağustos 2008.
- ^ Volker Siegel (2011). V. Siegel (ed.). "Nyctiphanes Sars, 1883 ". Dünya Euphausiacea veritabanı. Dünya Deniz Türleri Kaydı. Alındı 18 Haziran 2011.
- ^ a b c J. Mauchline; L.R. Fisher (1969). Euphausiids Biyolojisi. Deniz Biyolojisindeki Gelişmeler. 7. Akademik Basın. ISBN 978-7-7708-3615-2.
- ^ a b c Jaime Gómez-Gutiérrez; Carlos J. Robinson (2005). "Embriyonik, erken larva gelişme zamanı, kuluçka mekanizması ve kese yumurtlama euphausiidinin kuluçka dönemi Nyctiphanes simplex Hansen ". Plankton Araştırma Dergisi. 27 (3): 279–295. doi:10.1093 / plankt / fbi003.
- ^ S. N. Jarman; N. G. Elliott; S. Nicol; A. McMinn (2002). "Antarktika kıyı krillerinde genetik farklılaşma Euphausia crystallorophias". Kalıtım. 88 (4): 280–287. doi:10.1038 / sj.hdy.6800041. PMID 11920136.
- ^ R. Escribano; V. Marin; C. Irribarren (2000). "Dağılımı Euphausia mucronata Şili'nin kuzeyindeki Yarımada Mejillones'in yükselen bölgesinde: oksijen minimum katmanının etkisi ". Scientia Marina. 64 (1): 69–77. doi:10.3989 / scimar.2000.64n169.
- ^ P. Brueggeman. "Euphausia crystallorophias". Ross Adası ve McMurdo Sound, Antarktika'ya Sualtı Sahası Rehberi. California Üniversitesi, San Diego.
- ^ "Krill, Euphausia superba". MarineBio.org. Alındı 25 Şubat 2009.
- ^ J.A. Kirkwood (1984). "Güney Okyanusu'nun Euphausiacea Rehberi". ANARE Araştırma Notları. 1: 1–45.
- ^ A. Sala; M. Azzali; A. Russo (2002). "Ross Denizi Krili: Dağılımı, bolluğu ve demografisi Euphausia superba ve Euphausia crystallorophias İtalyan Antarktika Seferi sırasında (Ocak – Şubat 2000) ". Scientia Marina. 66 (2): 123–133. doi:10.3989 / scimar.2002.66n2123.
- ^ G. W. Hosie; M. Fukuchi; S. Kawaguchi (2003). "Güney Okyanusu Sürekli Plankton Kaydedici anketinin geliştirilmesi" (PDF). Oşinografide İlerleme. 58 (2–4): 263–283. Bibcode:2003PrOce..58..263H. doi:10.1016 / j.pocean.2003.08.007.[kalıcı ölü bağlantı ]
- ^ E. Gaten. "Meganyctiphanes norvegica". Leicester Üniversitesi. Arşivlenen orijinal 1 Temmuz 2009. Alındı 25 Şubat 2009.
- ^ E. Brinton (1953). "Thysanopoda spinicauda, yeni bir batipelajik dev euphousid kabuklu, karşılaştırmalı notlarla T. cornuta ve T. egregia". Washington Bilimler Akademisi Dergisi. 43: 408–412.
- ^ "Euphausiacea". Tazmanya Su Ürünleri ve Balıkçılık Enstitüsü. Arşivlenen orijinal 30 Eylül 2009. Alındı 6 Haziran 2010.
- ^ O. Shimomura (1995). "Euphausiid karideslerinin biyolüminesansında yer alan, oldukça kararsız iki bileşen F ve P'nin rolleri". Biyolüminesans ve Kemilüminesans Dergisi. 10 (2): 91–101. doi:10.1002 / biyo.1170100205. PMID 7676855.
- ^ J. C. Dunlap; J. W. Hastings; O. Shimomura (1980). "Uzaktan ilişkili organizmaların ışık yayan sistemleri arasındaki çapraz reaksiyon: yeni tip ışık yayan bileşik". Ulusal Bilimler Akademisi Bildiriler Kitabı. 77 (3): 1394–1397. Bibcode:1980PNAS ... 77.1394D. doi:10.1073 / pnas.77.3.1394. JSTOR 8463. PMC 348501. PMID 16592787.
- ^ P. J. Herring; E. A. Widder (2001). "Plankton ve Nekton'da Biyolüminesans". J. H. Steele'de; S. A. Thorpe; K. K. Turekyan (editörler). Okyanus Bilimi Ansiklopedisi. 1. Akademik Basın, San Diego. pp.308–317. ISBN 978-0-12-227430-5.
- ^ S. M. Lindsay; M. I. Latz (1999). Bazı euphausiid kabuklular tarafından ışıldayan karşı gölgeleme için deneysel kanıtlar. Amerikan Limnoloji ve Oşinografi Derneği (ASLO) Su Bilimleri Toplantısı. Santa Fe.
- ^ Sönke Johnsen (2005). "Kırmızı ve Siyah: biyolüminesans ve derin denizlerdeki hayvanların rengi" (PDF). Bütünleştirici ve Karşılaştırmalı Biyoloji. 4 (2): 234–246. doi:10.1093 / icb / 45.2.234. PMID 21676767. S2CID 247718. Arşivlenen orijinal (PDF) 2005-10-02 tarihinde.
- ^ a b c d Cavan, EL, Belcher, A., Atkinson, A., Hill, SL, Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. ve Steinberg, DK (2019) "Antarktika krilinin biyojeokimyasal döngülerde önemi". Doğa iletişimi, 10(1): 1–13. doi:10.1038 / s41467-019-12668-7.
 Materyal, bir altında bulunan bu kaynaktan kopyalandı. Creative Commons Attribution 4.0 Uluslararası Lisansı.
Materyal, bir altında bulunan bu kaynaktan kopyalandı. Creative Commons Attribution 4.0 Uluslararası Lisansı. - ^ G. C. Cripps; A. Atkinson (2000). "Antarktik krilinde etoburun bir göstergesi olarak yağ asidi bileşimi, Euphausia superba". Kanada Balıkçılık ve Su Bilimleri Dergisi. 57 (S3): 31–37. doi:10.1139 / f00-167.
- ^ a b Olav Saether; Trond Erling Ellingsen; Viggo Mohr (1986). "Kuzey Atlantik krilinin lipidleri" (PDF). Lipid Araştırma Dergisi. 27 (3): 274–285. PMID 3734626.
- ^ M. J. Schramm (10 Ekim 2007). "Tiny Krill: Deniz Ürünleri Zincirindeki Devler". NOAA Ulusal Deniz Koruma Alanı Programı. Alındı 4 Haziran 2010.
- ^ J. Weier (1999). "Değişen akıntılar Bering Denizi'ne yeni bir mavi tonu koyuyor". NOAA Dünya Gözlemevi. Alındı 15 Haziran 2005.
- ^ R. D. Brodeur; G. H. Kruse; P. A. Livingston; G. Walters; J. Ianelli; G. L. Swartzman; M. Stepanenko; T. Wyllie-Echeverria (1998). FOCI Uluslararası Bering Denizindeki Son Koşullar Çalıştayı Taslak Raporu. NOAA. s. 22–26.
- ^ J. Roach (17 Temmuz 2003). "Bilim adamları gizemli krill katilini keşfetti". National Geographic Haberleri.
- ^ J. Gómez-Gutiérrez; W. T. Peterson; A. de Robertis; R.D. Brodeur (2003). "Parazitoid siliatların neden olduğu krillerde kitlesel ölüm". Bilim. 301 (5631): 339. doi:10.1126 / science.1085164. PMID 12869754. S2CID 28471713.
- ^ J. D. Shields; J. Gómez-Gutiérrez (1996). "Oculophryxus bicauliseuphausiid üzerinde yeni bir tür ve decad izopod paraziti türü Stylocheiron afin Hansen ". Uluslararası Parazitoloji Dergisi. 26 (3): 261–268. doi:10.1016/0020-7519(95)00126-3. PMID 8786215.
- ^ Rusty Dornin (6 Temmuz 1997). "Antarktika kril nüfusu azalıyor". CNN. Alındı 18 Haziran 2011.
- ^ Dawson, Amanda L; Kawaguchi, So; Kral Catherine K; Townsend, Kathy A; Kral Robert; Huston, Wilhelmina M; Bengtson Nash, Susan M (2018). "Antarktik krilleri tarafından sindirim parçalanması yoluyla mikroplastikleri nanoplastiklere dönüştürmek". Doğa İletişimi. 9 (1): 1001. Bibcode:2018NatCo ... 9.1001D. doi:10.1038 / s41467-018-03465-9. PMC 5843626. PMID 29520086.
- ^ M. D. Knight (1984). "Güney Kaliforniya Körfezi popülasyonundaki larva morfogenezindeki varyasyon Euphausia pacifica Kıştan Yaza, 1977–1978 " (PDF). CalCOFI Raporu. XXV.
- ^ "Euphausia superba". Tür bilgi formu. Gıda ve Tarım Örgütü. Alındı 4 Haziran 2010.
- ^ R. M. Ross; L. B. Quetin (1986). "Antarktika krilleri ne kadar üretkendir?". BioScience. 36 (4): 264–269. doi:10.2307/1310217. JSTOR 1310217.
- ^ Janine Cuzin-Roudy (2000). "Kuzey krilinde mevsimsel üreme, çoklu yumurtlama ve doğurganlık, Meganyctiphanes norvegicave Antarktika kril, Euphausia superba". Kanada Balıkçılık ve Su Bilimleri Dergisi. 57 (S3): 6-15. doi:10.1139 / f00-165.
- ^ J. Gómez-Gutiérrez (2002). "Laboratuar koşullarında euphausiid türünün yumurtlayan üç yayında yumurtlayan yumurtalarının kuluçka mekanizması ve gecikmiş yumurtadan çıkması". Plankton Araştırma Dergisi. 24 (12): 1265–1276. doi:10.1093 / plankt / 24.12.1265.
- ^ E. Brinton; M. D. Ohman; A. W. Townsend; M. D. Knight; A. L. Bridgeman (2000). Dünya Okyanusu'nun Euphausiids. Dünya Biyoçeşitlilik Veritabanı CD-ROM Serisi, Springer Verlag. ISBN 978-3-540-14673-5.
- ^ F. Buchholz (2003). "Güney ve Kuzey krillerinin fizyolojisi üzerine deneyler, Euphausia superba ve Meganyctiphanes norvegica, with emphasis on moult and growth – a review". Marine and Freshwater Behaviour and Physiology. 36 (4): 229–247. doi:10.1080/10236240310001623376. S2CID 85121989.
- ^ H.-C. Shin; S. Nicol (2002). "Using the relationship between eye diameter and body length to detect the effects of long-term starvation on Antarctic krill Euphausia superba". Deniz Ekolojisi İlerleme Serisi. 239: 157–167. Bibcode:2002MEPS..239..157S. doi:10.3354/meps239157.
- ^ B. Marinovic; M. Mangel (1999). "Krill can shrink as an ecological adaptation to temporarily unfavourable environments" (PDF). Ekoloji Mektupları. 2: 338–343.
- ^ J. G. Gómez (1995). "Distribution patterns, abundance and population dynamics of the euphausiidsNyctiphanes simplex ve Euphausia eximia off the west coast of Baja California, Mexico" (PDF). Deniz Ekolojisi İlerleme Serisi. 119: 63–76. Bibcode:1995MEPS..119...63G. doi:10.3354/meps119063.
- ^ U. Kils; P. Marshall (1995). "Der Krill, wie er schwimmt und frisst – neue Einsichten mit neuen Methoden ("The Antarctic krill – how it swims and feeds – new insights with new methods")". In I. Hempel; G. Hempel (eds.). Biologie der Polarmeere – Erlebnisse und Ergebnisse (Biology of the Polar Oceans Experiences and Results). Fischer Verlag. s. 201–210. ISBN 978-3-334-60950-7.
- ^ R. Piper (2007). Olağanüstü Hayvanlar: Meraklı ve Sıradışı Hayvanların Ansiklopedisi. Greenwood Press. ISBN 978-0-313-33922-6.
- ^ Gandomi, A.H.; Alavi, A.H. (2012). "Krill Herd: A New Bio-Inspired Optimization Algorithm". Doğrusal Olmayan Bilim ve Sayısal Simülasyonda İletişim. 17 (12): 4831–4845. Bibcode:2012CNSNS..17.4831G. doi:10.1016/j.cnsns.2012.05.010.
- ^ J. S. Jaffe; M. D. Ohmann; A. de Robertis (1999). "Sonar estimates of daytime activity levels of Euphausia pacifica in Saanich Inlet" (PDF). Kanada Balıkçılık ve Su Bilimleri Dergisi. 56 (11): 2000–2010. doi:10.1139/cjfas-56-11-2000. Arşivlenen orijinal (PDF) on 2011-07-20.
- ^ Geraint A. Tarling; Magnus L. Johnson (2006). "Satiation gives krill that sinking feeling". Güncel Biyoloji. 16 (3): 83–84. doi:10.1016/j.cub.2006.01.044. PMID 16461267.
- ^ Dan Howard (2001). "Krill" (PDF). In Herman A. Karl; John L. Chin; Edward Ueber; Peter H. Stauffer; James W. Hendley II (eds.). Beyond the Golden Gate – Oceanography, Geology, Biology, and Environmental Issues in the Gulf of the Farallones. Amerika Birleşik Devletleri Jeolojik Araştırması. s. 133–140. Circular 1198. Alındı 8 Ekim 2011.
- ^ Wishart, Skye (July–August 2018). "The krill effect". Yeni Zelanda Coğrafi (152): 24.
- ^ D. Howard. "Krill in Cordell Bank National Marine Sanctuary". Ulusal Okyanus ve Atmosfer İdaresi. Alındı 15 Haziran 2005.
- ^ David A. Demer; Stéphane G. Conti (2005). "New target-strength model indicates more krill in the Southern Ocean". ICES Journal of Marine Science. 62 (1): 25–32. doi:10.1016/j.icesjms.2004.07.027.
- ^ U. Kils (1982). "Swimming behavior, swimming performance and energy balance of Antarctic krill Euphausia superba". BIOMASS Scientific Series 3, BIOMASS Research Series: 1–122.
- ^ S. Nicol; Y. Endo (1997). "Krill Fisheries of the World". FAO Fisheries Technical Paper. 367.
- ^ Ratnarajah, L., Bowie, A.R., Lannuzel, D., Meiners, K.M. and Nicol, S. (2014) "The biogeochemical role of baleen whales and krill in Southern Ocean nutrient cycling". PLOS ONE, 9(12): e114067. doi:10.1371/journal.pone.0114067
- ^ Hopkins, T.L., Ainley, D.G., Torres, J.J., Lancraft, T.M., 1993. Trophic structure in open waters of the Marginal Ice Zone in the Scotia Weddell Confluence region during spring (1983). Polar Biology 13, 389–397.
- ^ Lancraft, T.M., Relsenbichler, K.R., Robinson, B.H., Hopkins, T.L., Torres, J.J., 2004. A krill-dominated micronekton and macrozooplankton community in Croker Passage, Antarctica with an estimate of fish predation. Deep-Sea Research II 51, 2247–2260.
- ^ a b c Grossman, Elizabeth (14 July 2015). "Scientists consider whether krill need to be protected from human over-hunting". Public Radio International (PRI). Alındı 1 Nisan 2017.
- ^ "Krill fisheries and sustainability: Antarctic krill (Euphausia superba)". Commission for the Conservation of Antarctic Marine Living Resources. 23 Nisan 2015. Alındı 1 Nisan 2017.
- ^ a b Schiermeier, Q (2010). "Ecologists fear Antarctic krill crisis". Doğa. 467 (7311): 15. doi:10.1038/467015a. PMID 20811427.
- ^ a b c "Krill – biology, ecology and fishing". Commission for the Conservation of Antarctic Marine Living Resources. 28 Nisan 2015. Alındı 1 Nisan 2017.
- ^ Minturn J. Wright (1987). "The Ownership of Antarctica, its Living and Mineral Resources". Journal of Law and the Environment. 4 (2): 49–78.
- ^ S. Nicol; J. Foster (2003). "Recent trends in the fishery for Antarctic krill". Sucul Yaşam Kaynakları. 16: 42–45. doi:10.1016/S0990-7440(03)00004-4.
- ^ Josh, Gabbatiss (10 July 2018). "Krill fishing industry backs massive Antarctic ocean sanctuary to protect penguins, seals and whales". Bağımsız. Alındı 10 Temmuz 2018.
- ^ a b "Why krill?". Southwest Fisheries Science Center, US National Oceanic and Atmospheric Administration. 22 Kasım 2016. Alındı 1 Nisan 2017.
- ^ Cheeseman MA (22 July 2011). "Krill oil: Agency Response Letter GRAS Notice No. GRN 000371". ABD FDA. Alındı 3 Haziran 2015.
- ^ Omori, M. (1978). "Zooplankton fisheries of the world: A review". Deniz Biyolojisi. 48 (3): 199–205. doi:10.1007/BF00397145. S2CID 86540101.
- ^ Pongsetkul, Jaksuma; Benjakul, Soottawat; Sampavapol, Punnanee; Osako, Kazufumi; Faithong, Nandhsha (17 September 2014). "Chemical composition and physical properties of salted shrimp paste (Kapi) produced in Thailand". International Aquatic Research. 6 (3): 155–166. doi:10.1007/s40071-014-0076-4.
- ^ Abe, Kenji; Suzuki Kenji; Hashimoto, Kanehisa (1979). "Krill'in Balık Sosu Malzemesi Olarak Kullanımı". Nippon Suisan Gakkaishi. 45 (8): 1013–1017. doi:10.2331 / suisan.45.1013.
daha fazla okuma
- Boden, Brian P.; Johnson, Martin W.; Brinton, Edward: "Euphausiacea (Crustacea) of the North Pacific". Bulletin of the Scripps Institution of Oceanography. Volume 6 Number 8, 1955.
- Brinton, Edward: "Euphausiids of Southeast Asian waters". Naga Report volume 4, part 5. La Jolla: University of California, Scripps Institution of Oceanography, 1975.
- Conway, D. V. P.; White, R. G .; Hugues-Dit-Ciles, J.; Galienne, C. P.; Robins, D. B.: Guide to the coastal and surface zooplankton of the South-Western Indian Ocean, Sipariş Euphausiacea, Occasional Publication of the Birleşik Krallık Deniz Biyolojisi Derneği No. 15, Plymouth, UK, 2003.
- Everson, I. (ed.): Krill: biology, ecology and fisheries. Oxford, Blackwell Science; 2000. ISBN 0-632-05565-0.
- Hamner, William M. (May 1984). "Krill — Untapped Bounty From the Sea?". National Geographic. Cilt 165 hayır. 5. pp. 626–642. ISSN 0027-9358. OCLC 643483454.
- Mauchline, J.: Euphausiacea: Yetişkinler, Conseil International pour l'Exploration de la Mer, 1971. Identification sheets for adult krill with many line drawings. PDF file, 2 Mb.
- Mauchline, J.: Euphausiacea: Larvalar, Conseil International pour l'Exploration de la Mer, 1971. Identification sheets for larval stages of krill with many line drawings. PDF file, 3 Mb.
- Tett, P.: The biology of Euphausiids, lecture notes from a 2003 course in Marine Biology itibaren Napier Üniversitesi.
- Tett, P.: Biyolüminesans, lecture notes from the 1999/2000 edition of that same course.
Dış bağlantılar
- Webcam of Krill Aquarium at Australian Antarctic Division
- 'Antarctic Energies' animation by Lisa Roberts



