Görsel korteks - Visual cortex
| Görsel korteks | |
|---|---|
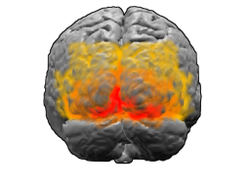 Beynin arkadan görünümü. Kırmızı = Brodmann alan 17 (birincil görsel korteks); turuncu = alan 18; sarı = alan 19 | |
 Yandan gösterilen beyin sola dönük. Yukarıda: dışarıdan görünüm, alttan: ortadan kes. Turuncu = Brodmann alan 17 (birincil görsel korteks) | |
| Detaylar | |
| Tanımlayıcılar | |
| Latince | Korteks görsel |
| MeSH | D014793 |
| NeuroLex İD | nlx_143552 |
| FMA | 242644 |
| Nöroanatominin anatomik terimleri | |
görsel korteks of beyin alanı beyin zarı bu süreçler görsel bilgi. İçinde bulunur oksipital lob. Gelen duyusal girdi gözler içinden geçer yanal genikülat çekirdek içinde talamus ve sonra görsel kortekse ulaşır. Yanal genikülat çekirdekten duyusal girdiyi alan görsel korteksin alanı, görme alanı 1 (V1) olarak da bilinen birincil görsel kortekstir. Brodmann bölgesi 17 veya çizgili korteks. yabancı alanlar 2, 3, 4 ve 5 görsel alanlarından oluşur (V2, V3, V4 ve V5 olarak da bilinir veya Brodmann bölgesi 18 ve tüm Brodmann bölgesi 19 ).[1]
Her ikisi de beynin yarım küreleri görsel bir korteks içerir; sol yarıküredeki görsel korteks sağdan sinyaller alır görsel alan ve sağ yarıküredeki görsel korteks, sol görme alanından sinyaller alır.
Giriş
Birincil görsel korteks (V1), kalkarin fissürü içinde oksipital lob. Her yarım kürenin V1'i bilgileri doğrudan ipsilateralinden alır yanal genikülat çekirdek kontralateral görsel yarım alandan sinyaller alan.
Nöronlar görsel korteks ateşinde aksiyon potansiyalleri görsel uyaranlar kendi içlerinde göründüğünde alıcı alan. Tanım gereği alıcı alan, tüm görsel alan içinde bir eylem potansiyeli ortaya çıkaran bölgedir. Ancak, herhangi bir nöron için, alıcı alanı içindeki bir uyaran alt kümesine en iyi yanıt verebilir. Bu mülk denir nöronal ayarlama. Daha önceki görsel alanlarda, nöronların daha basit ayarları vardır. Örneğin, V1'deki bir nöron, alıcı alanındaki herhangi bir dikey uyarana ateş edebilir. Daha yüksek görsel alanlarda, nöronların karmaşık ayarları vardır. Örneğin, alt temporal kortekste (IT), bir nöron, yalnızca alıcı alanında belirli bir yüz göründüğünde ateşlenebilir.
Görsel korteks, kan kaynağını öncelikle kalkarin dalı of posterior serebral arter.
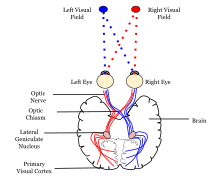
Görsel bilginin sinirsel işlemesinin psikolojik modeli
Ventral-dorsal model

V1, bilgileri ventral akış ve dorsal akış adı verilen iki birincil yola iletir.[2]
- ventral akım V1 ile başlar, V2 görsel alanından, sonra V4 görsel alanından ve alt temporal korteks (BT korteksi). Bazen "Ne Yolu" olarak adlandırılan ventral akış, form tanıma ve nesne temsiliyle ilişkilidir. Ayrıca depolama ile de ilişkilidir. uzun süreli hafıza.
- dorsal akım V1 ile başlar, V2 görsel alanından geçer, ardından dorsomedial bölge (DM / V6) ve medial temporal alan (MT / V5) ve posterior parietal korteks. Bazen "Nerede Yol" veya "Nasıl Yol" olarak adlandırılan dorsal akım, özellikle görsel bilgiler rehberlik etmek için kullanıldığında, hareket, nesne konumlarının temsili ve gözlerin ve kolların kontrolü ile ilişkilidir. Sakkadlar veya ulaşan.
Ventral / dorsal yolların neye karşı nerede hesabı ilk olarak Ungerleider ve Mishkin.[3]
Son zamanlarda, Goodale ve Milner bu fikirleri genişletti ve ventral akışın görsel algı için kritik olduğunu, oysa dorsal akışın yetenekli eylemlerin görsel kontrolüne aracı olduğunu öne sürdü.[4] Gibi görsel yanılsamaların olduğu gösterilmiştir. Ebbinghaus illüzyonu algısal nitelikteki yargıları çarpıtır, ancak özne kavrama gibi bir eylemle karşılık verdiğinde hiçbir bozulma olmaz.[5]
Scharnowski ve Gegenfurtner'daki gibi işler[6] hem eylem hem de algılama sistemlerinin bu tür yanılsamalarla eşit derecede aldatıldığını öne sürer. Bununla birlikte, diğer çalışmalar, kavrama gibi yetenekli eylemlerin resimsel yanılsamalardan etkilenmediği fikrine güçlü bir destek sağlar.[7][8] ve eylem / algı ayrışmasının, serebral korteksteki dorsal ve ventral görsel yollar arasındaki işlevsel işbölümünü karakterize etmek için yararlı bir yol olduğunu öne sürmektedir.[9]
Birincil görsel korteks (V1)
Bu bölüm için ek alıntılara ihtiyaç var doğrulama. (Kasım 2016) (Bu şablon mesajını nasıl ve ne zaman kaldıracağınızı öğrenin) |


birincil görsel korteks beyinde en çok çalışılan görsel alandır. Memelilerde oksipital lobun arka kutbunda yer alır ve en basit, en erken kortikal görme alanıdır. Statik ve hareketli nesnelerle ilgili bilgileri işlemek için oldukça uzmanlaşmıştır ve şu alanlarda mükemmeldir: desen tanıma.[açıklama gerekli ]
Görsel sistemdeki işlevi veya evresi ile tanımlanan birincil görsel korteks yaklaşık olarak çizgili korteks, Ayrıca şöyle bilinir Brodmann bölgesi 17, anatomik konumu ile tanımlanan. "Çizgili korteks" adı, Gennari hattı çıplak gözle görülebilen belirgin bir şerit[10] temsil eden miyelinli aksonlar -den yan genikülat gövde katman 4'te sona eriyor gri madde.
Birincil görsel korteks, işlevsel olarak farklı altı katmana bölünmüştür ve 1'den 6'ya kadar etiketlenmiştir. yanal genikülat çekirdek (LGN), ayrıca 4A, 4B, 4Cα ve 4Cβ etiketli 4 katmana bölünmüştür. Sublamina 4Cα çoğunlukla magnoselüler LGN'den giriş, katman 4Cβ ise parvoselüler yollar.[11]
Her yarım küredeki yetişkin insan birincil görsel korteksindeki ortalama nöron sayısının yaklaşık 140 milyon olduğu tahmin edilmektedir.[12]
Fonksiyon
[alakalı? ]
Bu makale çoğu okuyucunun anlayamayacağı kadar teknik olabilir. Lütfen geliştirmeye yardım et -e uzman olmayanlar için anlaşılır hale getirinteknik detayları kaldırmadan. (Eylül 2016) (Bu şablon mesajını nasıl ve ne zaman kaldıracağınızı öğrenin) |
Korteksteki görsel işlemenin ilk aşaması V1 olarak adlandırılır. Primatlarda, V1 bir belirginlik haritası (önemli olanı vurgular) olarak bilinen dikkat değişimlerine rehberlik etmek için görsel girdilerden bakış kayması.[13]Bunu, görsel girdileri milyonlarca nörondan sinirsel ateşleme hızlarına dönüştürerek yapar, öyle ki en yüksek ateşleyen nöron tarafından işaretlenen görsel konumlar, bakış kaymasını çeken en dikkat çekici konumdur. V1'in atış hızları, üstün kollikulus (beynin ortasında) bakış değişimlerine rehberlik etmek için V1 etkinliklerini okur.
V1, vizyondaki uzamsal bilginin çok iyi tanımlanmış bir haritasına sahiptir. Örneğin, insanlarda, kalkarin sulkus (oksipital lobda) alt yarısına güçlü yanıt verir görsel alan (merkezin altında) ve kalkarinin alt bankası görme alanının üst yarısına kadar. Konsept olarak bu retinotopik haritalama, görsel görüntünün retina V1'e. V1'deki belirli bir konum ile öznel görsel alan arasındaki ilişki çok kesindir: hatta Kör noktalar V1 ile eşlenir. Evrim açısından, bu yazışma çok basittir ve V1'e sahip çoğu hayvanda bulunur. İnsanlarda ve hayvanlarda fovea (koniler retinada), V1'in büyük bir kısmı görme alanının küçük, merkezi kısmıyla eşlenir, bu fenomen kortikal büyütme.[14] Belki de doğru uzaysal kodlama amacıyla, V1'deki nöronlar en küçük alıcı alan herhangi bir görsel korteks mikroskobik bölgenin boyutu.
V1 nöronlarının ayarlama özellikleri (nöronların yanıt verdiği şey) zamanla büyük ölçüde farklılık gösterir. Zamanın erken dönemlerinde (40 ms ve daha fazla) bireysel V1 nöronları küçük bir uyarı setine güçlü bir şekilde uyum sağlar. Yani, nöronal yanıtlar görseldeki küçük değişiklikleri ayırt edebilir. yönelimler, uzaysal frekanslar ve renkler. Ayrıca, insanlarda ve hayvanlarda bireysel V1 nöronları dürbün görüşü göz baskınlığına sahiptir, yani iki gözden birine uyum sağlar. V1'de ve genel olarak birincil duyu korteksinde, benzer ayarlama özelliklerine sahip nöronlar, kortikal sütunlar. David Hubel ve Torsten Wiesel iki ayar özelliği için kortikal sütunların klasik buz küpü organizasyon modelini önerdi: göz baskınlığı ve yönelim. Bununla birlikte, bu model, nöronların ayarlandığı renk, uzaysal frekans ve diğer birçok özelliği barındıramaz.[kaynak belirtilmeli ]. V1'deki tüm bu kortikal sütunların tam organizasyonu, güncel araştırmaların sıcak bir konusu olmaya devam ediyor. Bu fonksiyonun matematiksel modellemesi karşılaştırılmıştır. Gabor dönüşümleri.
Daha sonra (100 ms sonra), V1'deki nöronlar sahnenin daha küresel organizasyonuna da duyarlıdır (Lamme & Roelfsema, 2000).[15] Bu yanıt özellikleri muhtemelen tekrarlayan geri bildirim işleme (daha yüksek seviyeli kortikal alanların alt kademeli kortikal alanlar üzerindeki etkisi) ve yanal bağlantılar piramidal nöronlar (Hupe ve diğerleri 1998). İleri beslemeli bağlantılar esas olarak itici güç olsa da, geri bildirim bağlantıları çoğunlukla etkilerinde düzenleyicidir (Angelucci ve diğerleri, 2003; Hupe ve diğerleri, 2001). Kanıtlar, daha büyük ve daha karmaşık alıcı alanlara sahip V4, BT veya MT gibi daha üst düzey alanlardan kaynaklanan geri bildirimlerin, bağlamsal veya daha karmaşık alıcı alanları hesaba katarak V1 yanıtlarını değiştirebileceğini ve şekillendirebileceğini göstermektedir. ekstra klasik alıcı alan etkiler (Guo ve diğerleri, 2007; Huang ve diğerleri, 2007; Sillito ve diğerleri, 2006).
V1'e aktarılan görsel bilgiler, uzamsal (veya optik) görüntü açısından kodlanmamıştır[kaynak belirtilmeli ] daha çok şöyle tanımlanır: Kenar algılama. Örnek olarak, yarı tarafı siyah ve yarı beyazı içeren bir görüntü için, siyah ve beyaz arasındaki bölme çizgisi en güçlü yerel kontrasta (yani kenar algılama) sahiptir ve kodlanırken, az sayıda nöron parlaklık bilgisini kodlar (siyah veya beyaz se). Bilgi daha sonraki görsel alanlara aktarıldıkça, giderek artan şekilde yerel olmayan frekans / faz sinyalleri olarak kodlanır. Kortikal görsel işlemenin bu erken aşamalarında, görsel bilginin uzamsal konumunun yerel kontrast kodlaması (kenar algılama) arasında iyi korunduğuna dikkat edin.
V1'deki basit hücrelerin aksiyomatik olarak belirlenmiş fonksiyonel modelleri Lindeberg tarafından belirlenmiştir.[16][17] zamansal alan üzerindeki nedensel olmayan veya zamana bağlı ölçek-uzay çekirdeklerinin zamansal türevleri ile kombinasyon halinde uzamsal alan üzerindeki afin Gauss çekirdeklerinin yönlü türevleri açısından (bkz. alıcı alanların aksiyomatik teorisi ). Spesifik olarak, bu teorinin her ikisinin de, DeAngelis ve diğerleri tarafından gerçekleştirilen biyolojik alıcı alan ölçümleri ile iyi niteliksel uyumu olan alıcı alanlar hakkında tahminlere yol açtığı gösterilmiştir.[18][19] ve doğal görüntü dönüşümleri altında kovaryans ve değişmezlik özellikleri dahil matematiksel alıcı alan modelinin iyi teorik özelliklerini garanti eder.[20][alakalı? ]
V1'in boyutlarındaki farklılıkların da yanılsama algısı.[21]
V2
Görsel alan V2veya ikincil görsel korteks, olarak da adlandırılır prestriate korteks,[22] görsel korteksteki ikinci büyük alan ve içindeki ilk bölgedir. görsel ilişki alanı. Güçlü alır ileri besleme V1'den bağlantılar (doğrudan ve pulvinar aracılığıyla) ve V3, V4 ve V5'e güçlü bağlantılar gönderir. Aynı zamanda güçlü gönderir geri bildirim V1 bağlantıları[kaynak belirtilmeli ].
Anatomi açısından, V2 dört kadrana bölünmüştür, sırt ve karın sol ve sağdaki temsil yarım küreler. Bu dört bölge birlikte, görsel dünyanın eksiksiz bir haritasını sağlar. V2, V1 ile ortak birçok özelliğe sahiptir: Hücreler, yönlendirme, uzamsal frekans ve renk gibi basit özelliklere ayarlanmıştır. Birçok V2 nöronunun tepkileri, aynı zamanda daha karmaşık özelliklerle de modüle edilir. yanıltıcı konturlar,[23][24] binoküler uyumsuzluk,[25] ve uyaranın şeklin mi yoksa zeminin bir parçası mı olduğu.[26][27] Son araştırmalar, V2 hücrelerinin az miktarda dikkat modülasyonu gösterdiğini (V1'den fazla, V4'ten az), orta derecede karmaşık modeller için ayarlandığını ve tek bir alıcı alan içinde farklı alt bölgelerdeki çoklu yönelimlerle yönlendirilebileceğini göstermiştir.
Tüm ventral görsel-hipokampal akışın görsel hafıza için önemli olduğu tartışılmaktadır.[28] Bu teori, baskın olandan farklı olarak, nesne tanıma belleğindeki (ORM) değişikliklerin, görsel kortekslerin ventral akışı içinde oldukça birbirine bağlı bir alan olan V2'deki manipülasyondan kaynaklanabileceğini öngörür. Maymun beyninde, bu alan birincil görsel korteksten (V1) güçlü ileri beslemeli bağlantılar alır ve diğer ikincil görsel kortekslere (V3, V4 ve V5) güçlü projeksiyonlar gönderir.[29][30] Primatlarda bu alandaki nöronların çoğu, yönelim, uzaysal frekans, boyut, renk ve şekil gibi basit görsel özelliklere göre ayarlanmıştır.[24][31][32] Anatomik çalışmalar, görsel bilgi işlemede alan V2'nin 3. katmanını içerir. Katman 3'ün aksine, görsel korteksin 6. katmanı birçok nöron türünden oluşur ve bunların görsel uyaranlara tepkileri daha karmaşıktır.
Yakın zamanda yapılan bir çalışmada, V2 korteksinin 6. Katman hücrelerinin, Nesne Tanıma Belleğinin depolanmasında ve kısa süreli nesne anılarının uzun süreli anılara dönüştürülmesinde çok önemli bir rol oynadığı bulundu.[33]
Alan V3 dahil üçüncü görsel korteks
Dönem üçüncü görsel kompleks V2'nin hemen önünde bulunan ve adı verilen bölgeyi içeren korteks bölgesini ifade eder görsel alan V3 insanlarda. "Karmaşık" isimlendirme, bazı araştırmacıların V2'nin önünde bulunan korteksin iki veya üç işlevsel alt bölüm içerebileceğini öne sürmesiyle, V3 alanının tam kapsamı ile ilgili bazı tartışmaların hala mevcut olması gerçeğiyle doğrulanmaktadır. Örneğin, David Van Essen ve diğerleri (1986), serebral hemisferin üst kısmında bulunan "ventral V3" ten (veya ventral posterior alan, VP) farklı bir "dorsal V3" varlığını önermişlerdir. beynin alt kısmı. Dorsal ve ventral V3, beynin diğer bölümleriyle farklı bağlantılara sahiptir, çeşitli yöntemlerle boyanmış bölümlerde farklı görünür ve farklı görsel uyaran kombinasyonlarına yanıt veren nöronlar içerir (örneğin, renk seçici nöronlar ventralde daha yaygındır. V3). İnsanlarda V3A ve V3B dahil ek alt bölümler de bildirilmiştir. Bu alt bölümler dorsal V3 yakınında bulunur, ancak V2'ye bitişik değildir.
Dorsal V3, normalde dorsal akışın bir parçası olarak kabul edilir, V2'den ve birincil görsel alandan girdiler alır ve posteriora yansıtır. parietal korteks. Anatomik olarak Brodmann bölgesi 19. FMRI kullanan Braddick, alan V3 / V3A'nın işlenmesinde bir rol oynayabileceğini öne sürmüştür. küresel hareket[34] Diğer çalışmalar, dorsal V3'ü, adı verilen daha geniş bir alanın parçası olarak düşünmeyi tercih etmektedir. dorsomedial bölge (DM), tüm görsel alanın bir temsilini içerir. DM bölgesindeki nöronlar tutarlı hareket görsel alanın geniş bölümlerini kapsayan büyük desenler (Lui ve işbirlikçileri, 2006).
Ventral V3 (VP), birincil görsel alandan çok daha zayıf bağlantılara ve alt temporal korteks. Daha önceki çalışmalar, VP'nin görme alanının sadece üst kısmının (sabitleme noktasının üstünde) bir temsilini içerdiğini öne sürerken, daha yeni çalışmalar bu alanın daha önce takdir edilenden daha kapsamlı olduğunu ve diğer görsel alanlar gibi tam bir görüntü içerebileceğini gösteriyor. görsel sunum. Revize edilmiş, daha kapsamlı VP, Rosa ve Tweedale tarafından ventrolateral arka alan (VLP) olarak adlandırılıyor.[35]
V4
Görsel alan V4 görsel alanlardan biridir. yabancı görsel korteks. İçinde makaklar V2'nin önünde ve arkasında bulunur posterior inferotemporal alan (PIT). En az dört bölgeden oluşur (sol ve sağ V4d, sol ve sağ V4v) ve bazı gruplar rostral ve kaudal alt bölümleri de içerdiğini bildirir. İnsan V4'ün makak kadar geniş olup olmadığı bilinmemektedir. homolog bu bir tartışma konusudur.[36]
V4, üçüncü kortikal alandır. ventral akım, V2'den güçlü ileri besleme girdisi alarak ve PIT. Ayrıca, özellikle merkezi alan için V1'den doğrudan girdi alır. Ek olarak, V5 ile daha zayıf bağlantıları vardır ve dorsal prelunat girus (DP).
V4, ventral akım güçlü dikkat modülasyonu göstermek için. Çoğu araştırma gösteriyor ki seçici dikkat V4'teki ateşleme oranlarını yaklaşık% 20 oranında değiştirebilir. Moran ve Desimone tarafından yazılan ve bu etkileri karakterize eden ufuk açıcı bir makale, görsel korteksin herhangi bir yerinde dikkat etkilerini bulan ilk makaleydi.[37]
V2 gibi, V4 de yönlendirme, uzamsal frekans ve renk için ayarlanmıştır. V2'den farklı olarak V4, basit geometrik şekiller gibi ara karmaşıklıktaki nesne özellikleri için ayarlanmıştır, ancak hiç kimse V4 için ayar alanının tam parametrik bir tanımını geliştirmemiştir. V4 görsel alanı, görüntüdeki alanlar gibi yüzler gibi karmaşık nesneler için ayarlanmamıştır. inferotemporal korteks vardır.
V4'ün ateşleme özellikleri ilk olarak Semir Zeki 1970'lerin sonlarında, bölgeyi de adlandıran. Bundan önce, V4 anatomik açıklamasıyla biliniyordu, prelunate girus. Başlangıçta Zeki, V4'ün amacının renk bilgilerini işlemek olduğunu savundu. 1980'lerin başındaki çalışmalar, V4'ün önceki kortikal alanlar kadar form tanımaya doğrudan dahil olduğunu kanıtladı.[kaynak belirtilmeli ] Bu araştırma, iki akışlı hipotez, ilk kez 1982'de Ungerleider ve Mishkin tarafından sunulmuştur.
Yakın zamanda yapılan çalışmalar, V4'ün uzun vadeli plastisite sergilediğini göstermiştir.[38] uyaran belirginliğini kodlar, ön göz alanları,[39] ve alıcı alanlarının uzamsal profilindeki değişiklikleri dikkatle gösterir.[kaynak belirtilmeli ]
Orta temporal görme alanı (V5)
orta zamansal görsel alan (MT veya V5), ekstra bir görsel korteks bölgesidir. Her ikisinin birkaç türünde Yeni Dünya maymunları ve Eski Dünya maymunları MT alanı yüksek konsantrasyonda yön seçici nöronlar içerir.[40] Primatlarda MT'nin önemli bir rol oynadığı düşünülmektedir. hareket algısı, yerel hareket sinyallerinin küresel algılara entegrasyonu ve bazılarının rehberliği göz hareketleri.[40]
Bağlantılar
MT, çok çeşitli kortikal ve subkortikal beyin alanlarına bağlıdır. Girdisi görsel kortikal alanlardan V1, V2 ve dorsal V3'ten gelir (dorsomedial bölge ),[41][42] konioselüler bölgeleri LGN,[43] ve aşağı pulvinar.[44] MT'ye projeksiyonların paterni, foveal ve çevresel görsel alanların temsilleri arasında biraz değişir; ikincisi, orta hat korteksinde bulunan alanlardan girdiler alır ve retrosplenial bölge.[45]
Standart bir görünüm, V1'in MT'ye "en önemli" girdiyi sağlamasıdır.[40] Bununla birlikte, birkaç çalışma, MT'deki nöronların, V1 yok edildikten veya etkisiz hale getirildikten sonra bile, genellikle yön seçici bir şekilde görsel bilgilere yanıt verebildiğini göstermiştir.[46] Ayrıca, araştırma Semir Zeki ve ortak çalışanlar, belirli türdeki görsel bilgilerin MT'ye daha V1'e ulaşmadan ulaşabileceğini öne sürdü.
MT, ana çıktısını, FST, MST ve V4t (orta zamansal hilal) dahil olmak üzere hemen çevreleyen kortekste bulunan alanlara gönderir. Diğer MT projeksiyonları frontal ve parietal lobların (frontal göz alanı ve lateral intraparietal alan) göz hareketiyle ilgili alanlarını hedefler.
Fonksiyon
İlk çalışmalar elektrofizyolojik MT'deki nöronların özellikleri, hücrelerin büyük bir kısmının ayarlanmış hareketli görsel uyaranların hızına ve yönüne.[47][48]
Lezyon çalışmalar ayrıca MT'nin hareket algılama ve göz hareketlerindeki rolünü desteklemiştir.[49] Nöropsikolojik Hareket göremeyen bir hasta üzerine yapılan çalışmalar, bunun yerine dünyayı bir dizi statik 'çerçevede' görmek, primattaki V5'in insandaki MT ile homolog olduğunu öne sürdü.[50][51]
Bununla birlikte, V1'deki nöronlar hareketin yönüne ve hızına da ayarlandığından, bu erken sonuçlar tam olarak MT'nin V1'in yapamayacağı ne yapabileceği sorusunu gündeme getirdi. Yerel görsel hareket sinyallerini karmaşık nesnelerin küresel hareketine entegre ettiği için bu bölge üzerinde çok çalışma yapılmıştır.[52]Örneğin, lezyon V5, hareketin algılanmasında ve karmaşık uyaranların işlenmesinde eksikliklere yol açar. Karmaşık görsel özelliklerin (çizgi uçları, köşeler) hareketi için seçici olan birçok nöron içerir. Mikro uyarım V5'te bulunan bir nöronun hareketi, hareket algısını etkiler. Örneğin, bir maymunun V5'inde yukarı doğru hareketi tercih eden bir nöron bulursa ve onu bir elektrotla uyarırsa, maymunun 'sol' ve 'sağ' içeren uyaranlarla sunulduğunda 'yukarı' hareket bildirme olasılığı daha yüksektir. 'yukarı doğru' bileşenler olarak.[53]
MT bölgesinde yapılan hesaplamaların kesin şekli konusunda hala çok tartışmalar var.[54] ve bazı araştırmalar, özellik hareketinin aslında V1 gibi görsel sistemin daha düşük seviyelerinde zaten mevcut olduğunu gösteriyor.[55][56]
Işlevsel organizasyon
MT'nin yön sütunlarında düzenlendiği gösterildi.[57] DeAngelis, MT nöronlarının da binoküler eşitsizlik ayarlarına göre düzenlendiğini savundu.[58]
V6
dorsomedial bölge (DM) olarak da bilinir V6, kendi kendine hareketle ilişkili görsel uyaranlara yanıt veriyor gibi görünüyor[59] ve geniş alan uyarımı.[60] V6, ilk olarak tarafından tanımlanan primatların görsel korteksinin bir alt bölümüdür. John Allman ve Jon Kaas 1975'te.[61] V6, sırtın sırt kısmında yer alır. aşırı korteks, beynin ortasındaki derin oluğun yakınında (medial longitudinal fissür ) ve tipik olarak aynı zamanda medial korteksin kısımlarını içerir, örneğin parieto-oksipital sulkus (POS).[60]:7970 DM, tüm görüş alanının topografik olarak organize edilmiş bir temsilini içerir.[60]:7970
V5 ve V6 görsel alanı arasında benzerlikler vardır. ortak marmoset. Her iki alan da doğrudan bağlantı alır. birincil görsel korteks.[60]:7971 Ve ikisinin de yüksek miyelin içerik, genellikle bilginin hızlı iletimi ile ilgili beyin yapılarında bulunan bir özelliktir.[62]
Uzun yıllar boyunca, DM'nin yalnızca Yeni Dünya maymunları. Bununla birlikte, daha yeni araştırmalar, DM'nin de var olduğunu ileri sürdü. Eski Dünya maymunları ve insanlar.[60]:7972 V6'ya bazen parieto-oksipital alan (PO) da denir, ancak yazışmalar kesin değildir.[63][64]
Özellikleri
Bölgenin DM / V6 bölgesindeki nöronlar gece maymunları ve ortak ipek maymunları görsel konturların yönlendirilmesi için son derece keskin bir seçicilik ve görsel alanın büyük kısımlarını kaplayan uzun, kesintisiz çizgiler tercihi dahil olmak üzere benzersiz tepki özelliklerine sahiptir.[65][66]
Bununla birlikte, MT alanı ile karşılaştırıldığında, çok daha küçük bir DM hücresi oranı, görsel modellerin hareket yönü için seçicilik gösterir.[67] MT alanı ile bir diğer önemli fark, DM'deki hücrelerin bir görüntünün düşük uzamsal frekans bileşenlerine uyum sağlaması ve rastgele noktalar alanı gibi dokulu desenlerin hareketine zayıf yanıt vermesidir.[67] Bu yanıt özellikleri, DM ve MT'nin paralel olarak çalışabileceğini, ilki çevreye göre kendi kendine hareketi analiz ettiğini ve ikincisinin arka plana göre tek tek nesnelerin hareketini analiz ettiğini göstermektedir.[67]
Son zamanlarda, insanda geniş açılı akış alanlarına yanıt veren bir alan tanımlanmıştır ve makak alanı V6'nın bir homologu olduğu düşünülmektedir.[68]
Yollar
DM / V6'daki hücrelerin bağlantıları ve yanıt özellikleri, bu alanın bir alt kümedeki anahtar düğüm olduğunu göstermektedir.dorsal akım ', bazıları tarafından' dorsomedial yol 'olarak anılır.[kaynak belirtilmeli ] Bu yolun, postüral reaksiyonlar ve nesnelere yönelik hareketler dahil olmak üzere iskelet motor aktivitesinin kontrolü için önemli olması muhtemeldir.[64] DM'nin ana 'ileri beslemeli' bağlantısı, oksipital ve paryetal loblar (V6A) arasındaki arayüzde, kortekse hemen rostraldir.[kaynak belirtilmeli ] Bu bölge, sırasıyla, frontal lobun kol hareketlerini kontrol eden bölgeleri ile nispeten doğrudan bağlantılara sahiptir. motor öncesi korteks.[kaynak belirtilmeli ]
Ayrıca bakınız
- Kortikal alan
- Kortikal körlük
- Özellik entegrasyon teorisi
- İnsan beynindeki bölgelerin listesi
- Retinotopi
- Görsel işleme
- Görsel özellik dizisi
- Karmaşık hücre
Referanslar
- ^ Mather, George. "Görsel Korteks". Yaşam Bilimleri Okulu: Sussex Üniversitesi. Sussex Üniversitesi. Alındı 6 Mart 2017.
- ^ Braz, José; Pettré, Julien; Richard, Paul; Kerren, Andreas; Linsen, Lars; Battiato, Sebastiano; Imai, Francisco (11 Şubat 2016). "Etkili Nesne Tanıma için Hedeflenen HMAX Modelinde Algoritmik Optimizasyonlar". Bitar'da, Ahmad W .; Mansour, Mohamad M .; Chehab, Ali (ed.). Bilgisayarla Görme, Görüntüleme ve Bilgisayar Grafikleri Teorisi ve Uygulamaları. Berlin, Almanya: Springer. s. 377. ISBN 9783319299716.
- ^ Ungerleider LG, Mishkin M (1982). "İki Kortikal Görsel Sistem". Ingle DJ, Goodale MA, Mansfield RJ (editörler). Görsel Davranış Analizi. Boston: MIT Press. pp.549–586.
- ^ Goodale MA, Milner AD (1992). "Algılama ve eylem için ayrı yollar". Sinirbilimlerindeki Eğilimler. 15 (1): 20–25. CiteSeerX 10.1.1.207.6873. doi:10.1016/0166-2236(92)90344-8. PMID 1374953. S2CID 793980.
- ^ Aglioti S, DeSouza JF, Goodale MA (1995). "Boyut-kontrast illüzyonları gözü aldatır ama eli yanıltmaz". Güncel Biyoloji. 5 (6): 679–85. doi:10.1016 / S0960-9822 (95) 00133-3. PMID 7552179. S2CID 206111613.
- ^ Franz VH, Scharnowski F, Gegenfurtner (2005). "Kavrama üzerindeki yanılsama etkileri dinamik değil geçici olarak sabittir". Deneysel Psikoloji Dergisi: İnsan Algısı ve Performansı. 31 (6): 1359–78. doi:10.1037/0096-1523.31.6.1359. PMID 16366795.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Ganel T, Goodale MA (2003). "Eylemin görsel kontrolü, ancak algının değil, nesne şeklinin analitik olarak işlenmesini gerektirir". Doğa. 426 (6967): 664–7. Bibcode:2003Natur.426..664G. doi:10.1038 / nature02156. PMID 14668865. S2CID 4314969.
- ^ Ganel T, Tanzer M, Goodale MA (2008). "Görsel yanılsamalar bağlamında eylem ve algı arasında çifte ayrışma: gerçek ve yanıltıcı boyutun zıt etkileri". Psikolojik Bilim. 19 (3): 221–5. doi:10.1111 / j.1467-9280.2008.02071.x. PMID 18315792. S2CID 15679825.
- ^ Goodale MA. (2011). "Vizyonu eyleme dönüştürmek". Vizyon Araştırması. 51 (14): 1567–87. doi:10.1016 / j.visres.2010.07.027. PMID 20691202.
- ^ Glickstein M., Rizzolatti G. Francesco Gennari ve serebral korteksin yapısı Nörobilimlerde Eğilimler, Cilt 7, Sayı 12, 464-467, 1 Aralık 1984.
- ^ Hubel, D.H., Wiesel, T.N .. Makak maymunda genikülo-kortikal liflerin laminer ve kolumnar dağılımı. Karşılaştırmalı Nöroloji Dergisi, Sayı 146, 421–450, 1972.
- ^ Leuba G; Kraftsik R (1994). "İnsan birincil görsel korteksinin hacim, yüzey tahmini, üç boyutlu şekli ve toplam nöron sayısındaki değişimler, orta yaştan yaşlılığa kadar." Anatomi ve Embriyoloji. 190 (4): 351–366. doi:10.1007 / BF00187293. PMID 7840422. S2CID 28320951.
- ^ Zhaoping, L. 2014, V1 hipotezi - önceden özenli seçim ve segmentasyon için aşağıdan yukarıya bir belirginlik haritası oluşturma, https://www.oxfordscholarship.com/view/10.1093/acprof:oso/9780199564668.001.0001/acprof-9780199564668-chapter-5.
- ^ Barghout Lauren (1999). Periferik ve Foveal Desen Maskeleme Arasındaki Farklar Üzerine. Berkeley, California, ABD: Yüksek Lisans Tezi. U.C. Berkeley.
- ^ Barghout Lauren (2003). Vizyon: Küresel Algısal Bağlam Yerel Kontrast İşlemeyi Nasıl Değiştirir (Doktora Tezi). Bilgisayarla görme tekniklerini içerecek şekilde güncellendi. Scholar's Press. ISBN 978-3-639-70962-9.
- ^ Lindeberg, T. (2013). "Görsel alıcı alanların hesaplamalı bir teorisi". Biyolojik Sibernetik. 107 (6): 589–635. doi:10.1007 / s00422-013-0569-z. PMC 3840297. PMID 24197240.
- ^ Lindeberg, T. (2016). "Zamana bağlı ve zamanla özyinelemeli uzay-zamansal alıcı alanlar". Matematiksel Görüntüleme ve Görme Dergisi. 55 (1): 50–88. arXiv:1504.02648. doi:10.1007 / s10851-015-0613-9. S2CID 5000571.
- ^ DeAngelis, G. C .; Ohzawa, I .; Freeman, R.D. (1995). "Merkezi görsel yollarda alıcı alan dinamikleri". Sinirbilimlerindeki Eğilimler. 18 (10): 451–457. doi:10.1016 / 0166-2236 (95) 94496-r. PMID 8545912. S2CID 12827601.
- ^ GC DeAngelis ve A. Anzai "Klasik alıcı alanın modern bir görünümü: V1 nöronları tarafından doğrusal ve doğrusal olmayan uzay-zamansal işleme. İçinde: Chalupa, LM, Werner, JS (eds.) The Visual Neurosciences, cilt 1, s. 704–719. MIT Press, Cambridge, 2004.
- ^ Lindeberg, T. (2013). "Alıcı alanlar düzeyinde görsel işlemlerin değişkenliği". PLOS ONE. 8 (7): e66990. arXiv:1210.0754. Bibcode:2013PLoSO ... 866990L. doi:10.1371 / journal.pone.0066990. PMC 3716821. PMID 23894283.
- ^ Schwarzkopf, D Samuel (2011). "İnsan V1'in yüzey alanı, nesne boyutunun öznel deneyimini tahmin ediyor". Doğa Sinirbilim. 14 (1): 28–30. doi:10.1038 / nn.2706. PMC 3012031. PMID 21131954.
- ^ Gazzaniga, Ivry ve Mangun: Bilişsel sinirbilim, 2002
- ^ von der Heydt, R; Peterhans, E; Baumgartner, G (1984). "Yanıltıcı konturlar ve kortikal nöron tepkileri". Bilim. 224 (4654): 1260–62. Bibcode:1984Sci ... 224.1260V. doi:10.1126 / science.6539501. PMID 6539501.
- ^ a b Anzai, A; Peng, X; Van Essen, D.C (2007). "V2 maymun görme alanındaki nöronlar, yönelimlerin kombinasyonlarını kodlar". Doğa Sinirbilim. 10 (10): 1313–21. doi:10.1038 / nn1975. PMID 17873872. S2CID 6796448.
- ^ von der Heydt, R; Zhou, H; Friedman, H.S (2000). "Maymun görsel korteksinde stereoskopik kenarların temsili". Vizyon Araştırması. 40 (15): 1955–67. doi:10.1016 / s0042-6989 (00) 00044-4. PMID 10828464. S2CID 10269181.
- ^ Qiu, F. T; von der Heydt, R (2005). "Görsel kortekste şekil ve zemin: V2 stereoskopik ipuçlarını Gestalt kurallarıyla birleştirir". Nöron. 47 (1): 155–66. doi:10.1016 / j.neuron.2005.05.028. PMC 1564069. PMID 15996555.
- ^ Maruko, ben; ve alt. (2008). "Makak Maymunlarının Görme Alanı 2'de (V2) Eşitsizlik Duyarlılığının Doğum Sonrası Gelişimi". Nörofizyoloji Dergisi. 100 (5): 2486–2495. doi:10.1152 / jn.90397.2008. PMC 2585398. PMID 18753321.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Bussey, T J; Saksida, L. M (2007). "Hafıza, algılama ve ventral görsel-perirhinal-hipokampal akış: kutuların dışında düşünme". Hipokamp. 17 (9): 898–908. doi:10.1002 / hipo.20320. PMID 17636546.
- ^ Stepniewska, I; Kaas, J.H. (1996). "Makak maymunlarında V2 kortikal bağlantılarının topografik modelleri". Karşılaştırmalı Nöroloji Dergisi. 371 (1): 129–152. doi:10.1002 / (SICI) 1096-9861 (19960715) 371: 1 <129 :: AID-CNE8> 3.0.CO; 2-5. PMID 8835723.
- ^ Gattas, R; Sousa, A. P; Mishkin, M; Ungerleider, L. G. (1997). "Makaktaki bölge V2'nin kortikal izdüşümleri". Beyin zarı. 7 (2): 110–129. doi:10.1093 / cercor / 7.2.110. PMID 9087820.
- ^ Hegdé, Jay; Van Essen, D. C (2000). "Primat Görsel Alanı V2'de Karmaşık Şekiller için Seçicilik". Nörobilim Dergisi. 20 (5): RC61. doi:10.1523 / JNEUROSCI.20-05-j0001.2000. PMID 10684908.
- ^ Hegdé, Jay; Van Essen, D. C (2004). "Makak görsel alanı V2'de şekil analizinin zamansal dinamikleri". Nörofizyoloji Dergisi. 92 (5): 3030–3042. doi:10.1152 / jn.00822.2003. PMID 15201315. S2CID 6428310.
- ^ López-Aranda ve diğerleri. (2009). "Nesne Tanıma Belleğinde V2 Görsel Korteksin 6. Katmanının Rolü". Bilim. 325 (5936): 87–89. Bibcode:2009Sci ... 325 ... 87L. doi:10.1126 / science.1170869. PMID 19574389. S2CID 23990759.
- ^ Braddick, OJ, O'Brien, JM; et al. (2001). "Tutarlı görsel harekete duyarlı beyin alanları". Algı. 30 (1): 61–72. doi:10.1068 / p3048. PMID 11257978. S2CID 24081674.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Rosa MG, Tweedale R (2000). "Marmoset maymunlarının lateral ve ventral ekstrastriat kortekslerindeki görsel alanlar". Karşılaştırmalı Nöroloji Dergisi. 422 (4): 621–51. doi:10.1002 / 1096-9861 (20000710) 422: 4 <621 :: AID-CNE10> 3.0.CO; 2-E. PMID 10861530.
- ^ Goddard E; Goddard, Erin; McDonald, J. S .; Solomon, S. G .; Clifford, C.W. G .; et al. (2011). "Renk duyarlılığı, insan V4'ün dorsal bileşenine karşı çıkıyor". Journal of Vision. 11 (4): 3. doi:10.1167/11.4.3. PMID 21467155.
- ^ Moran, J; Desimone, R (1985). "Extrastriate Cortex'te Görsel İşleme Seçici Dikkat Kapısı". Bilim. 229 (4715): 782–4. Bibcode:1985Sci ... 229..782M. CiteSeerX 10.1.1.308.6038. doi:10.1126 / science.4023713. PMID 4023713.
- ^ Schmid, M. C .; Schmiedt, J. T .; Peters, A. J .; Saunders, R. C .; Maier, A .; Leopold, D.A. (27 Kasım 2013). "Görsel Alan V4'te Birincil Görsel Korteks Olmadan Harekete Duyarlı Yanıtlar" (PDF). Nörobilim Dergisi. 33 (48): 18740–18745. doi:10.1523 / JNEUROSCI.3923-13.2013. PMC 3841445. PMID 24285880.
- ^ Moore, Tirin; Armstrong, Katherine M. (2003). "Ön korteksin mikro uyarımı ile görsel sinyallerin seçici geçitlenmesi". Doğa. 421 (6921): 370–373. Bibcode:2003Natur.421..370M. doi:10.1038 / nature01341. PMID 12540901. S2CID 4405385.
- ^ a b c R, Bradley D (2005) doğdu. "MT görme alanının yapısı ve işlevi". Yıllık Nörobilim İncelemesi. 28: 157–89. doi:10.1146 / annurev.neuro.26.041002.131052. PMID 16022593.
- ^ Felleman D, Van Essen D (1991). "Primat serebral korteksinde dağıtılmış hiyerarşik işlem". Beyin zarı. 1 (1): 1–47. doi:10.1093 / cercor / 1.1.1-a. PMID 1822724.
- ^ Ungerleider L, Desimone R (1986). "Makakta görme alanı MT'nin kortikal bağlantıları". Karşılaştırmalı Nöroloji Dergisi. 248 (2): 190–222. doi:10.1002 / cne.902480204. PMID 3722458.
- ^ Sincich L, Park K, Wohlgemuth M, Horton J (2004). "V1'i baypas etme: MT alanına doğrudan bir genikülat girişi". Doğa Sinirbilim. 7 (10): 1123–8. doi:10.1038 / nn1318. PMID 15378066. S2CID 13419990.
- ^ Warner CE, Goldshmit Y, Bourne JA (2010). "Retina afferentleri, pulvinar ve lateral genikülat çekirdeklerdeki orta temporal alanı hedefleyen röle hücreleriyle sinaps olur". Nöroanatomide Sınırlar. 4: 8. doi:10.3389 / nöro.05.008.2010. PMC 2826187. PMID 20179789.
- ^ Palmer SM, Rosa MG (2006). "Uzak periferik görüşte hareketin analizi için kortikal alanlardan oluşan farklı bir anatomik ağ". Avrupa Nörobilim Dergisi. 24 (8): 2389–405. doi:10.1111 / j.1460-9568.2006.05113.x. PMID 17042793.
- ^ Rodman HR, Gross CG, Albright TD (1989). "Makağın MT alanındaki görsel tepki özelliklerinin afferent temeli. I. Çizgili korteks çıkarmanın etkileri". Nörobilim Dergisi. 9 (6): 2033–50. doi:10.1523 / JNEUROSCI.09-06-02033.1989. PMC 6569731. PMID 2723765.
- ^ Dubner R, Zeki S (1971). "Maymundaki üstün temporal sulkusun anatomik olarak tanımlanmış bir bölgesindeki hücrelerin yanıt özellikleri ve alıcı alanları". Beyin Araştırması. 35 (2): 528–32. doi:10.1016 / 0006-8993 (71) 90494-X. PMID 5002708..
- ^ Maunsell J, Van Essen D (1983). "Makak maymunun orta zamansal görsel alanındaki nöronların işlevsel özellikleri. I. Uyaran yönü, hızı ve yönelimi için seçicilik". Nörofizyoloji Dergisi. 49 (5): 1127–47. doi:10.1152 / jn.1983.49.5.1127. PMID 6864242. S2CID 8708245.
- ^ Dursteler M.R .; Wurtz R.H .; Newsome W.T. (1987). "Makak maymunun üst temporal sulkusu içindeki foveal temsil lezyonlarını takip eden yönsel takip kusurları". Nörofizyoloji Dergisi. 57 (5): 1262–87. CiteSeerX 10.1.1.375.8659. doi:10.1152 / jn.1987.57.5.1262. PMID 3585468.
- ^ Hess R.H .; Baker C.L .; Zihl J. (1989). "'Hareket körü' hasta: düşük seviyeli uzaysal ve zamansal filtreler". Nörobilim Dergisi. 9 (5): 1628–40. doi:10.1523 / JNEUROSCI.09-05-01628.1989. PMC 6569833. PMID 2723744.
- ^ Baker C.L. Jr; Hess R.F; Zihl J. (1991). "'Hareket körü' bir hastada artık hareket algısı, sınırlı ömür boyu rastgele nokta uyaranlarıyla değerlendirildi". Nörobilim Dergisi. 11 (2): 454–61. doi:10.1523 / JNEUROSCI.11-02-00454.1991. PMC 6575225. PMID 1992012.
- ^ Movshon, J.A., Adelson, E.H., Gizzi, M.S. ve Newsome, W.T. (1985). Hareketli görsel desenlerin analizi. İçinde: C. Chagas, R. Gattass ve C. Gross (Eds.), Örüntü tanıma mekanizmaları (s. 117–151), Roma: Vatican Press.
- ^ Britten K.H .; van Wezel R.J. (1998). "Kortikal alanın elektriksel mikro uyarımı, maymunlarda algıya yön veren MST sapmaları" Doğa Sinirbilim. 1 (1): 59–63. doi:10.1038/259. PMID 10195110. S2CID 52820462.
- ^ Wilson, H.R .; Ferrera, V.P .; Yo, C. (1992). "İki boyutlu hareket algısı için psikofiziksel olarak motive edilmiş bir model". Görsel Sinirbilim. 9 (1): 79–97. doi:10.1017 / s0952523800006386. PMID 1633129.
- ^ Tinsley, C.J., Webb, B.S., Barraclough, N.E., Vincent, C.J., Parker, A. ve Derrington, A.M. (2003). "V1 nöral tepkilerinin 2B hareketli modellere doğası, marmoset maymunundaki alıcı alan yapısına bağlıdır". Nörofizyoloji Dergisi. 90 (2): 930–7. doi:10.1152 / jn.00708.2002. PMID 12711710.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Paket C.C .; R.T doğumlu; Livingstone M.S. (2003). "Makak görsel kortekste stereo ve hareket etkileşimlerinin iki boyutlu alt yapısı". Nöron. 37 (3): 525–35. doi:10.1016 / s0896-6273 (02) 01187-x. PMID 12575958.
- ^ Albright T (1984). "Makağın MT görsel alanındaki nöronların yön ve yönelim seçiciliği". Nörofizyoloji Dergisi. 52 (6): 1106–30. doi:10.1152 / jn.1984.52.6.1106. PMID 6520628.
- ^ DeAngelis G, Newsome W (1999). "Makak bölgesi MT'de eşitsizlik seçici nöronların organizasyonu". Nörobilim Dergisi. 19 (4): 1398–415. doi:10.1523 / JNEUROSCI.19-04-01398.1999. PMC 6786027. PMID 9952417.
- ^ Cardin, V; Smith, AT (2010). "İnsanın görsel ve vestibüler kortikal bölgelerinin kendi kendine hareketle ilişkili stereoskopik derinlik gradyanlarına duyarlılığı". Beyin zarı. 20 (8): 1964–73. doi:10.1093 / cercor / bhp268. PMC 2901022. PMID 20034998.
- ^ a b c d e Pitzalis vd. (2006). "Geniş Alan Retinotopisi İnsan Kortikal Görme Alanını V6 Tanımlar". Nörobilim Dergisi. 26 (30): 7962–73. doi:10.1523 / jneurosci.0178-06.2006. PMC 6674231. PMID 16870741.
- ^ Allman JM, Kaas JH (1975). "Dorsomedial kortikal görme alanı: baykuş maymunun (Aotus trivirgatus) oksipital lobunda üçüncü bir katman alanı". Beyin Araştırması. 100 (3): 473–487. doi:10.1016/0006-8993(75)90153-5. PMID 811327. S2CID 22980932.
- ^ Sabrina Pitzalis, Patrizia Fattori, and Claudio Galletti (2013 Jan 16) The functional role of the medial motion area V6
- ^ Galletti C, et al. (2005). "The relationship between V6 and PO in macaque extrastriate cortex" (PDF). Avrupa Nörobilim Dergisi. 21 (4): 959–970. CiteSeerX 10.1.1.508.5602. doi:10.1111/j.1460-9568.2005.03911.x. PMID 15787702.
- ^ a b Galletti C, et al. (2003). "Role of the medial parieto-occipital cortex in the control of reaching and grasping movements". Deneysel Beyin Araştırmaları. 153 (2): 158–170. doi:10.1007/s00221-003-1589-z. PMID 14517595. S2CID 1821863.
- ^ Baker JF, et al. (1981). "Visual response properties of neurons in four extrastriate visual areas of the owl monkey (Aotus trivirgatus): a quantitative comparison of medial, dorsomedial, dorsolateral, and middle temporal areas". Nörofizyoloji Dergisi. 45 (3): 397–416. doi:10.1152/jn.1981.45.3.397. PMID 7218008. S2CID 9865958.
- ^ Lui LL, et al. (2006). "Functional response properties of neurons in the dorsomedial visual area of New World monkeys (Callithrix jacchus)". Beyin zarı. 16 (2): 162–177. doi:10.1093/cercor/bhi094. PMID 15858163.
- ^ a b c http://www.fmritools.com/kdb/grey-matter/occipital-lobe/calcarine-visual-cortex/index.html
- ^ Pitzalis, S., Sereno, M.I., Committeri, G., Fattori, P., Galati, G., Patria, F., & Galletti, C. (2010). "Human v6: The medial motion area". Beyin zarı. 20 (2): 411–424. doi:10.1093/cercor/bhp112. PMC 2803738. PMID 19502476.CS1 Maint: yazar parametresini kullanır (bağlantı)
Dış bağlantılar
- The Primary Visual Cortex by Matthew Schmolesky -de Utah Üniversitesi
- Architecture of the Visual Cortex, by David Hubel -de Harvard Üniversitesi
- ancil-415 -de NeuroNames – striate area 17
- ancil-699 -de NeuroNames – Brodmann area 17 in guenon
- Stained brain slice images which include the "visual%20cortex" -de BrainMaps projesi
- Simulator for computational modeling of visual cortex maps at topographica.org