Birincil motor korteks - Primary motor cortex
| Birincil motor korteks | |
|---|---|
 Brodmann bölgesi 4 insan beyninin. | |
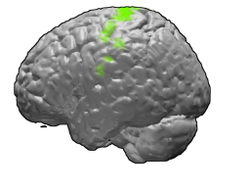
 Yeşil renkte gösterilen birincil motor korteks. | |
| Detaylar | |
| Parçası | Precentral girus |
| Arter | Ön serebral Orta serebral |
| Tanımlayıcılar | |
| Latince | korteks motorius primus |
| NeuroNames | 1910 |
| NeuroLex İD | nlx_143555 |
| FMA | 224854 |
| Nöroanatominin anatomik terimleri | |

birincil motor korteks (Brodmann bölgesi 4 ) insanlarda sırtın arka kısmında bulunan bir beyin bölgesidir. Frontal lob. O birincil bölge of motor sistemi ve dahil olmak üzere diğer motor alanlarıyla birlikte çalışır motor öncesi korteks, tamamlayıcı motor alanı, posterior parietal korteks ve hareketleri planlamak ve yürütmek için birkaç subkortikal beyin bölgesi. Birincil motor korteks, anatomik olarak, korteksin olarak bilinen büyük nöronları içeren bölgesi olarak tanımlanır. Betz hücreleri. Betz hücreleri, diğer kortikal nöronlarla birlikte uzun aksonlar aşağı omurilik -e sinaps omuriliğin internöron devresine ve ayrıca doğrudan omurilikteki kaslara bağlanan alfa motor nöronlarına.
Birincil motor kortekste, motor gösterimi, baş parmağından (serebral yarımkürenin tepesinde) ağza (altta), kortekste adı verilen bir kıvrım boyunca düzenli bir şekilde (ters bir şekilde) düzenlenir. Merkezi oluk. Bununla birlikte, bazı vücut kısımları, kısmen üst üste binen korteks bölgeleri tarafından kontrol edilebilir. Birincil motor korteksin her serebral yarımküresi yalnızca vücudun zıt (kontralateral) tarafının motor temsilini içerir. Bir vücut parçasına ayrılan birincil motor korteks miktarı, vücut yüzeyinin mutlak boyutu ile orantılı değildir, bunun yerine, söz konusu vücut parçası üzerindeki kutanöz motor reseptörlerinin nispi yoğunluğu ile orantılıdır. Vücut kısmındaki kutanöz motor reseptörlerinin yoğunluğu, genellikle o vücut kısmında gerekli olan gerekli hareket hassasiyeti derecesinin göstergesidir. Bu nedenle insan eli ve yüzü bacaklara göre çok daha büyük bir temsile sahiptir.
Birincil motor korteksin keşfi ve diğer motor kortikal alanlarla ilişkisi için ana makaleye bakınız. motor korteks.
Yapısı
İnsan birincil motor korteksi, merkezi sulkusun ön duvarında bulunur. Aynı zamanda sulkusun anteriorundan kısmen precentral girusa uzanır. Önde, birincil motor korteks, precentral girus üzerinde uzanan ve genellikle lateral premotor korteksi oluşturduğu düşünülen bir dizi alanla sınırlanmıştır. Arka tarafta, birincil motor korteks, merkezi sulkusun arka duvarında yer alan birincil somatosensoriyel korteks ile sınırlanmıştır. Ventral olarak birincil motor korteks lateral sulkustaki insular korteks ile sınırlanmıştır. Birincil motor korteks, yarıkürenin tepesine dorsal olarak uzanır ve ardından yarım kürenin medial duvarına doğru devam eder.
Birincil motor korteksin konumu, ayırt edici özelliğin varlığı nedeniyle histolojik incelemede en belirgindir. Betz hücreleri. Birincil motor korteksin V. tabakası dev (70-100 μm ) piramidal nöronlar Betz hücreleridir. Bu nöronlar uzun gönderir aksonlar kontralateral motor çekirdeklerine kafatası sinirleri ve alt motor nöronlar karın boynuzunda omurilik. Bu aksonlar, kortikospinal yol. Betz hücreleri, kortikospinal yolun yalnızca küçük bir yüzdesini oluşturur. Bazı önlemlere göre, omuriliğe yansıyan birincil motor korteks nöronlarının yaklaşık% 10'unu oluştururlar[1] veya omuriliğe yapılan toplam kortikal projeksiyonun yaklaşık% 2-3'ü.[2] Betz hücreleri, korteksin tüm motor çıktısını oluşturmasa da, yine de birincil motor korteks için açık bir belirteç sağlarlar. Betz hücrelerinin varlığı ile karakterize edilen bu korteks bölgesi, Brodmann tarafından 4. bölge olarak adlandırıldı.
Patika
Birincil motor olarak aksonlar serebral boyunca yolculuk Beyaz madde, birbirlerine yaklaşırlar ve arka uzuv of iç kapsül.
Aşağı doğru devam ediyorlar beyin sapı bazılarının kontralateral tarafa geçtikten sonra, kafatası siniri motor çekirdekleri. (Not: birkaç motor lifi sinaps ile alt motor nöronlar aynı tarafında beyin sapı ).
İçinde kontralateral tarafa geçtikten sonra medulla oblongata (piramidal çarpıklık ), aksonlar aşağı doğru hareket eder omurilik olarak yanal kortikospinal yol.
Üst üste geçmeyen lifler beyin sapı ayrı yolculuk ventral kortikospinal yol ve bunların çoğu kontralateral tarafa geçer. omurilik ulaşmadan kısa bir süre önce alt motor nöronlar. Ana kortikospinal yola ek olarak, Motor korteks striatum, hipotalamus, orta beyin ve arka beyin ile talamus bazal gangliya, orta beyin ve medulla dahil olmak üzere diğer kortikal ve subkortikal bölgelere de yansır.[3]
Kortikomotor nöronlar
Kortikomotor nöronlar Omuriliğin ventral boynuzundaki motor nöronlara doğrudan yansıtan birincil korteksteki nöronlardır.[4][5] Kortikomotor nöronların aksonları, birden fazla kasın spinal motor nöronlarında ve spinal internöronlarda sonlanır.[4][5] Primatlara özgüdürler ve işlevlerinin, ayrı ayrı parmakların nispeten bağımsız kontrolü dahil olmak üzere, uzak ekstremitelerin (örneğin eller) uyarlamalı kontrolü olduğu öne sürülmüştür.[5] Kortikomotornöronlar şimdiye kadar yalnızca birincil motor kortekste bulundu ve ikincil motor alanlarında bulunmadı.[5]
Kan temini
Şubeleri orta serebral arter birincil için arteriyel kan kaynağının çoğunu sağlar motor korteks.
Medial yönü (bacak alanları), ön serebral arter.
Fonksiyon
Homunculus
Motor adı verilen bir düzenlemede, birincil motor kortekste farklı vücut parçalarının geniş bir temsili vardır. homunculus (Latince: küçük insan).[6] Bacak bölgesi orta hatta yakın, motor alanının iç kısımlarında medial longitudinal fissür. Birincil motor korteksin yanal, dışbükey tarafı, kalça, gövde, omuz, dirsek, bilek, parmaklar, başparmak, göz kapakları, dudaklar ve çeneye karşılık gelen alanlarda yukarıdan aşağıya düzenlenmiştir. Kol ve el motor alanı en büyüğüdür ve prantral girusun bacak ile yüz bölgesi arasındaki kısmını kaplar.
Bu bölgeler vücuttaki boyutları ile orantılı değildir ve özellikle geniş alanlarla temsil edilen dudaklar, yüz kısımları ve ellerdir. Ampütasyon veya felçten sonra, motor bölgeler vücudun yeni kısımlarını benimsemek için kayabilir.
Talamustan nöral girdi
Birincil motor korteks, farklı talamik çekirdeklerden talamik girdiler alır. Diğerlerinin yanı sıra:
- Ventral lateral çekirdek serebellar afferentler için
- Ventral ön çekirdek bazal ganglion afferentleri için
Alternatif haritalar

Primatların birincil motor korteksinde vücut parçalarının klasik somatotopik düzeninde en az iki değişiklik bildirilmiştir.
İlk olarak, kol temsili bir çekirdek ve çevresel şekilde organize edilebilir. Maymun korteksinde, elin rakamları, birincil motor korteksin arka kenarında bir çekirdek alanda temsil edilir. Bu çekirdek alan, dirsek ve omuz dahil olmak üzere kolun daha proksimal kısımlarının bir temsili ile üç taraftan (dorsal, ön ve ventral taraflarda) çevrelenmiştir.[7][8] İnsanlarda, rakam temsili bilek temsili ile dorsal, anterior ve ventral olarak çevrilidir.[9]
Vücut parçalarının klasik somatotopik düzeninin ikinci bir modifikasyonu, esas olarak insan motor korteksinde incelenen rakamların ve bileğin çift temsilidir. Bir temsil, alan 4p adı verilen arka bölgede, diğeri ise alan 4a adı verilen ön bölgede yer alır. Arka alan herhangi bir duyusal geri bildirim olmaksızın dikkatle etkinleştirilebilir ve hareketlerin başlaması için önemli olduğu öne sürülürken, ön alan duyusal geri bildirime bağlıdır.[10] Hayali parmak hareketleriyle de aktive edilebilir.[11] ve hiçbir gerçek hareket yapmadan konuşmayı dinlemek. Bu ön temsil alanının, karmaşık duyu-motor etkileşimleri içeren hareketleri gerçekleştirmede önemli olduğu öne sürülmüştür.[12] İnsanlarda 4a alanının maymun korteksinde tarif edildiği gibi kaudal premotor korteksin bazı kısımlarına karşılık gelmesi mümkündür.
2009 yılında, iki ayrı evrimsel bölge olduğu bildirildi, biri dış yüzeyde daha eski, diğeri yarıkta bulundu. Daha yaşlı olanı, omurilikteki internöronlar aracılığıyla omurilik motor nöronlarına bağlanır. Yalnızca maymunlarda ve maymunlarda bulunan yenisi, doğrudan omurga motor nöronlarına bağlanır.[13] Doğrudan bağlantılar doğumdan sonra oluşur, dolaylı bağlantılara baskındır ve geliştirebilecekleri devrelerde daha esnektir, bu da karmaşık ince motor becerilerin doğum sonrası öğrenmesine izin verir. "Primat soyunun evrimi sırasında 'yeni' M1 bölgesinin ortaya çıkması, bu nedenle, insan elinin gelişmiş el becerisi için önemli olması muhtemeldir."[14]
Yaygın yanlış anlamalar
İkincil incelemelerde, ders kitaplarında ve popüler materyallerde birincil motor korteksle ilgili bazı yanlış anlamalar yaygındır. En yaygın yanılgılardan üçü burada listelenmiştir.
Gövdenin ayrılmış haritası
Birincil motor korteksle ilgili en yaygın yanılgılardan biri, vücut haritasının temiz bir şekilde ayrılmış olmasıdır. Yine de, bireyselleştirilmiş kasların veya hatta bireyselleştirilmiş vücut parçalarının bir haritası değildir. Harita önemli ölçüde çakışma içeriyor. Bu örtüşme, birincil motor korteksin daha ön bölgelerinde artar. Motor korteks üzerindeki çalışma tarihindeki ana hedeflerden biri, motor kortekste farklı vücut parçalarının ne kadar örtüştüğünü veya ayrıldığını belirlemekti. Bu sorunu ele alan araştırmacılar, el, kol ve omuz haritasının kapsamlı bir örtüşme içerdiğini buldular.[6][8][9][15][16][17][18][19] Kortikal nöronlardan kaslara kesin işlevsel bağlantıyı haritalayan araştırmalar, birincil motor korteksteki tek bir nöronun bile birçok eklemle ilgili birçok kasın aktivitesini etkileyebileceğini göstermektedir.[15] Kediler ve maymunlar üzerinde yapılan deneylerde, hayvanlar karmaşık, koordineli hareketleri öğrendikçe, birincil motor korteksteki harita daha üst üste biniyor ve belli ki birçok kasın kontrolünü entegre etmeyi öğreniyor.[20][21] Maymunlarda, davranışsal bir zaman ölçeğinde motor kortekse elektriksel uyarı uygulandığında, karmaşık, son derece entegre hareketler kavrama şeklindeki el ile uzanmak veya eli ağza götürmek ve ağzı açmak gibi.[22][23] Bu tür kanıtlar, birincil motor korteksin, vücudun kaba bir haritasını içerirken, tek tek kas gruplarının kontrolünü ayırmak yerine kasları anlamlı yollarla bütünleştirmeye katılabileceğini göstermektedir. Daha derin bir organizasyon ilkesinin, vücut parçalarının bir haritasından ziyade davranışsal repertuar içindeki istatistiksel korelasyonların bir haritası olabileceği öne sürülmüştür.[23][24] Hareket repertuarının kısmen ayrı vücut parçalarının eylemlerine bölündüğü ölçüde, harita kaba ve örtüşen bir vücut düzenlemesi içerir.
M1 ve birincil motor korteks
"M1" terimi ve "birincil motor korteks" terimi genellikle birbirinin yerine kullanılır. Bununla birlikte, farklı tarihsel geleneklerden gelirler ve farklı korteks bölümlerine atıfta bulunurlar. Bazı bilim adamları, motor korteksin daha posterior olan bir birincil motor şeride ve daha ön olan bir lateral premotor şeride bölünebileceğini öne sürdü. Bu görüşü orijinal olarak öneren ilk araştırmacılar arasında Campbell,[25] Vogt ve Vogt,[26] Foerster,[27] ve Fulton.[28] Diğerleri motor korteksin bu şekilde bölünemeyeceğini öne sürdü. Bunun yerine, bu ikinci görüşte, sözde birincil motor ve yanal ön motor şeritleri birlikte, M1 olarak adlandırılan tek bir kortikal alan oluşturdu. Yarım kürenin medial duvarındaki ikinci bir motor alanı M2 veya tamamlayıcı motor alanı. Bu görüşün savunucuları arasında Penfield vardı[6] ve Woolsey.[29] Günümüzde birincil motor korteks ile lateral premotor korteks arasındaki ayrım genel olarak kabul edilmektedir. Bununla birlikte, M1 terimi bazen yanlışlıkla birincil motor kortekse atıfta bulunmak için kullanılır. Kesin olarak söylemek gerekirse M1, önceki bazı araştırmacılara göre hem birincil motoru hem de lateral premotor korteksi kapsayan tek bir haritayı ifade eder.
Son ortak yol olarak Betz hücreleri
Betz hücreleri veya birincil motor korteksteki dev piramidal hücreler bazen korteksten omuriliğe giden tek veya ana çıktı olarak yanlış anlaşılır. Bu hata eskidir, en azından 1905'te Campbell'e kadar uzanmaktadır.[25] Yine de Betz hücreleri, korteksten omuriliğe uzanan nöronların yalnızca yaklaşık% 2-3'ünü oluşturur.[2] ve spesifik olarak birincil motor korteksten omuriliğe yansıyan nöronların sadece yaklaşık% 10'u.[1] Dahil olmak üzere bir dizi kortikal alan motor öncesi korteks, tamamlayıcı motor alanı ve hatta birincil somatosensoriyel korteks bile omuriliğe yansıtılır. Betz hücreleri hasar gördüğünde bile, korteks yine de subkortikal motor yapılarla iletişim kurabilir ve hareketi kontrol edebilir. Betz hücrelerinin bulunduğu birincil motor korteks hasar görürse, geçici bir felç oluşur ve diğer kortikal alanlar, açıkça kaybedilen işlevin bir kısmını devralabilir.
Klinik önemi
Precentral girus lezyonları, felç vücudun karşı tarafının (yüz felci, kol bacak monoparezi, hemiparezi ) - görmek üst motor nöron.
Hareket kodlaması
Evarts[30] motor korteksteki her nöronun bir kastaki güce katkıda bulunduğunu öne sürdü. Nöron aktif hale geldikçe, omuriliğe bir sinyal gönderir, sinyal bir motor nörona iletilir, motornöron bir kasa sinyal gönderir ve kas kasılır. Motor korteks nöronunda ne kadar fazla aktivite olursa, o kadar fazla kas gücü olur.
Georgopoulos ve meslektaşları[31][32][33] tek başına kas gücünün çok basit bir açıklama olduğunu öne sürdü. Maymunları çeşitli yönlere ulaşmak için eğittiler ve motor korteksteki nöronların aktivitesini izlediler. Motor korteksteki her bir nöronun belirli bir erişim yönü sırasında maksimum düzeyde aktif olduğunu ve komşu erişim yönlerine daha az yanıt verdiğini buldular. Bu temelde, motor korteksteki nöronların etkilerini "oylayarak" veya etkilerini bir "nüfus kodu ", tam olarak bir erişim yönünü belirtebilir.
Motor korteks nöronlarının bir erişimin yönünü kodladığı önerisi tartışmalı hale geldi. Scott ve Kalaska[34] her bir motor korteks nöronunun, erişim yönünden çok eklem hareketinin ve kas gücünün ayrıntılarıyla daha iyi korelasyon gösterdiğini gösterdi. Schwartz ve meslektaşları[35] motor korteks nöronlarının elin hızı ile iyi korelasyon gösterdiğini gösterdi. Strick ve meslektaşları[36] motor korteksteki bazı nöronların kas kuvveti ile ve bazılarının uzaysal hareket yönü ile ilişkili olarak aktif olduğunu bulmuşlardır. Todorov[37] birçok farklı korelasyonun, birçok hareket parametresinin kas kuvveti ile ilişkili olduğu bir kas kontrolörünün sonucu olduğunu öne sürdü.
Primat motor korteksindeki nöronların omuriliği ve dolayısıyla hareketi kontrol ettiği kod tartışmalıdır.
Kemirgen modelinde, motor korteksin nasıl harekete neden olduğunu anlamada belirli bir ilerleme kaydedilmiştir. Kemirgen motor korteksi, maymun motor korteksi gibi, farklı yaygın eylem türlerini vurgulayan alt bölgeler içerebilir.[38][39] Örneğin, bir bölgenin ritmik kontrolünü vurguladığı görülmektedir. çırpma.[38][40][41] Bu bölgedeki nöronlar, belirli bir subkortikal çekirdeği yansıtır. desen üreteci bıyıkların döngüsel ritmini koordine eder. Bu çekirdek daha sonra bıyıkları kontrol eden kaslara yansıtılır.
Ek resimler

Motor yolu.


Ayrıca bakınız
- Kortikospinal yol
- Motor korteks
- Kortikal homunkulus
- Üst motor nöron
- Brodmann bölgesi
- İnsan beynindeki bölgelerin listesi
Referanslar
- ^ a b Rivara CB, Sherwood CC, Bouras C, Hof PR (2003). "İnsan birincil motor korteksindeki Betz hücrelerinin stereolojik karakterizasyonu ve uzamsal dağılım modelleri". Anatomik Kayıt Bölüm A: Moleküler, Hücresel ve Evrimsel Biyolojide Keşifler. 270 (2): 137–151. doi:10.1002 / ar.a.10015. PMID 12524689.
- ^ a b Lassek, A.M. (1941). "Maymunun piramidal yolu". J. Comp. Neurol. 74 (2): 193–202. doi:10.1002 / cne.900740202. S2CID 83536088.
- ^ "Bölüm görüntüleyici". brainarchitecture.org. Alındı 2020-11-20.
- ^ a b Siegelbaum, Steven A .; Hudspeth, A.J (2013). Sinir biliminin ilkeleri. Kandel, Eric R. (5. baskı). New York. ISBN 9780071390118. OCLC 795553723.
- ^ a b c d Lemon, Roger N. (4 Nisan 2008). "Motor Kontrolde Azalan Yollar". Yıllık Nörobilim İncelemesi. 31 (1): 195–218. doi:10.1146 / annurev.neuro.31.060407.125547. ISSN 0147-006X. PMID 18558853.
- ^ a b c Penfield, W. ve Boldrey, E. (1937). "İnsanın beyin korteksindeki elektriksel uyarımla incelenen somatik motor ve duyusal temsil". Beyin. 60 (4): 389–443. doi:10.1093 / beyin / 60.4.389.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Kwan HC, MacKay WA, Murphy JT, Wong YC (1978). "Uyanık primatlarda prantral korteksin mekansal organizasyonu. II. Motor çıktıları". J. Neurophysiol. 41 (5): 1120–1131. doi:10.1152 / jn.1978.41.5.1120. PMID 100584.
- ^ a b Park, M.C., Belhaj-Saif, A., Gordon, M. ve Cheney, P.D. (2001). "Rhesus makaklarında primer motor korteksin ön ayaklardaki temsilindeki tutarlı özellikler". J. Neurosci. 21 (8): 2784–2792. doi:10.1523 / JNEUROSCI.21-08-02784.2001. PMID 11306630.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ a b Meier, J.D., Aflalo, T.N., Kastner, S. ve Graziano, M.S.A. (2008). "İnsan birincil motor korteksinin karmaşık organizasyonu: Yüksek çözünürlüklü bir fMRI çalışması". J. Neurophysiol. 100 (4): 1800–1812. doi:10.1152 / jn.90531.2008. PMC 2576195. PMID 18684903.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Binkofski F, Fink GR, Geyer S, Buccino G, Gruber O, Shah NJ, Taylor JG, Seitz RJ, Zilles K, Freund HJ (2002). "İnsan birincil motor korteks alanları 4a ve 4b'deki sinirsel aktivite, eyleme gösterilen dikkat ile farklı şekilde modüle edilir". J. Neurophysiol. 88 (1): 514–519. doi:10.1152 / jn.2002.88.1.514. PMID 12091573.
- ^ Nikhil Sharma; Not: Jones; T.A. Marangoz; Jean-Claude Baron (2008). "Motor İmgeleri sırasında BA 4a ve 4p'nin katılımının haritalanması". NeuroImage. 41 (1): 92–99. doi:10.1016 / j.neuroimage.2008.02.009. PMID 18358742. S2CID 8673179.
- ^ Terumitsu M, Ikeda K, Kwee IL ve Nakada, T (2009). "Karmaşık duyusal işlemede birincil motor korteks alanı 4a'nın katılımı: 3.0T fMRI çalışması". NeuroReport. 20 (7): 679–683. doi:10.1097 / WNR.0b013e32832a1820. PMID 19339906. S2CID 23674509.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Rathelot, J.-A .; strick, P. L. (20 Ocak 2009). "Kortiko-motonöronal hücrelere dayalı birincil motor korteksin alt bölümleri". Proc. Natl. Acad. Sci. 106 (3): 918–923. Bibcode:2009PNAS..106..918R. doi:10.1073 / pnas.0808362106. PMC 2621250. PMID 19139417.
- ^ Costandi, Mo (2009). "El becerisinin evrimi". Ulusal Bilimler Akademisi Bildiriler Kitabı. 106 (3): 918–923. doi:10.1073 / pnas.0808362106. PMC 2621250. PMID 19139417. Alındı 29 Kasım 2015.
- ^ a b Cheney, P.D. & Fetz, E.E. (1985). "Tek tek kortikomotonöronal (CM) hücreler ve primatlarda tek intrakortikal mikro uyaranlar tarafından uyandırılan karşılaştırılabilir kas kolaylaştırma modelleri: CM hücrelerinin fonksiyonel grupları için kanıt". J. Neurophysiol. 53 (3): 786–804. doi:10.1152 / jn.1985.53.3.786. PMID 2984354.
- ^ Schieber, M.H. & Hibbard, L.S. (1993). "Motor korteks el bölgesi ne kadar somatotopiktir?". Bilim. 261 (5120): 489–492. Bibcode:1993Sci ... 261..489S. doi:10.1126 / science.8332915. PMID 8332915.
- ^ Rathelot, J.A. & Strick, P.L. (2006). "Makak motor kortekste kas temsili: anatomik bir perspektif". Proc. Natl. Acad. Sci. Amerika Birleşik Devletleri. 103 (21): 8257–8262. Bibcode:2006PNAS..103.8257R. doi:10.1073 / pnas.0602933103. PMC 1461407. PMID 16702556.
- ^ Sanes, J.N., Donoghue, J.P., Thangaraj, V., Edelman, R.R. ve Warach, S. (1995). "İnsan motor korteksindeki el hareketlerini kontrol eden paylaşılan nöral substratlar". Bilim. 268 (5218): 1775–1777. Bibcode:1995Sci ... 268.1775S. doi:10.1126 / science.7792606. PMID 7792606.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Donoghue, J.P., Leibovic, S. ve Sanes, J.N. (1992). "Sincap maymun motor korteksindeki ön ayak bölgesinin organizasyonu: parmak, bilek ve dirsek kaslarının temsili". Tecrübe. Beyin Res. 89 (1): 1–10. doi:10.1007 / bf00228996. PMID 1601087. S2CID 1398462.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Nudo, R.J., Milliken, G.W., Jenkins, W.M. ve Merzenich, M.M. (1996). "Yetişkin sincap maymunlarının birincil motor korteksindeki hareket temsillerinin kullanıma bağlı değişiklikleri". J. Neurosci. 16 (2): 785–807. doi:10.1523 / JNEUROSCI.16-02-00785.1996. PMC 6578638. PMID 8551360.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Martin, J.H., Engber, D. ve Meng, Z. (2005). "Ön ayak kullanımının, kedinin birincil motor korteksinde ön ayak motor temsilinin doğum sonrası gelişimi üzerindeki etkisi". J. Neurophysiol. 93 (5): 2822–2831. doi:10.1152 / jn.01060.2004. PMID 15574795.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Graziano, M.S.A. Taylor, C.S.R. ve Moore, T. (2002). "Ön merkez korteksinin mikro uyarılmasından kaynaklanan karmaşık hareketler". Nöron. 34 (5): 841–851. doi:10.1016 / S0896-6273 (02) 00698-0. PMID 12062029. S2CID 3069873.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ a b Graziano, M.S.A. (2008). Akıllı Hareket Makinesi. Oxford, İngiltere: Oxford University Press.
- ^ Graziano, M.S.A. ve Aflalo, T.N. (2007). "Davranışsal repertuarın korteksle eşleştirilmesi". Nöron. 56 (2): 239–251. doi:10.1016 / j.neuron.2007.09.013. PMID 17964243.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ a b Campbell, A.W. (1905). Serebral Fonksiyonun Lokalizasyonu Üzerine Histolojik Çalışmalar. Cambridge, Massachusetts: Cambridge University Press.
- ^ Vogt, C. ve Vogt, O. (1919). "Ergebnisse unserer Hirnforschung". Journal für Psychologie und Neurologie. 25: 277–462.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Foerster, O (1936). Hughlings Jackson'ın doktrinleri ışığında "insanın motor korteksi". Beyin. 59 (2): 135–159. doi:10.1093 / beyin / 59.2.135.
- ^ Fulton, J (1935). "Motor" ve "premotor" alanların tanımına ilişkin bir not. Beyin. 58 (2): 311–316. doi:10.1093 / beyin / 58.2.311.
- ^ Woolsey, C.N., Settlage, P.H., Meyer, D.R., Sencer, W., Hamuy, T.P. ve Travis, A.M. (1952). "Ön ve" tamamlayıcı "motor alanlarındaki lokalizasyon paterni ve bunların premotor alan kavramı ile ilişkisi". Sinir ve Akıl Hastalıkları Araştırma Derneği. New York, NY: Raven Press. 30: 238–264.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Evarts, E.V. (1968). "Piramidal yol aktivitesinin gönüllü hareket sırasında uygulanan kuvvetle ilişkisi". J. Neurophysiol. 31 (1): 14–27. doi:10.1152 / jn.1968.31.1.14. PMID 4966614.
- ^ Georgopoulos, A.P., Kalaska, J.F., Caminiti, R. ve Massey, J.T. (1982). "Primat motor korteksinde iki boyutlu kol hareketlerinin yönü ile hücre boşalması arasındaki ilişkiler üzerine". J. Neurosci. 2 (11): 1527–1537. doi:10.1523 / JNEUROSCI.02-11-01527.1982. PMC 6564361. PMID 7143039.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Georgopoulos A.P., Kettner, R.E. ve Schwartz, A.B. (1988). "Üç boyutlu uzayda motor korteks ve serbest kol hareketlerini görsel hedeflere primat edin. II. Hareketin yönünün nöronal popülasyon tarafından kodlanması". J. Neurosci. 8 (8): 2928–2937. doi:10.1523 / JNEUROSCI.08-08-02928.1988. PMC 6569382. PMID 3411362.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Georgopoulos A.P., Schwartz, A.B. ve Kettner, R.E. (1986). "Hareket yönünün nöronal popülasyon kodlaması". Bilim. 233 (4771): 1416–1419. Bibcode:1986Sci ... 233.1416G. doi:10.1126 / science.3749885. PMID 3749885.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Scott, S.H. & Kalaska, J.F. (1995). "Benzer el yolları, ancak farklı kol duruşlarıyla hareketlere ulaşma sırasında motor korteks aktivitesindeki değişiklikler". J. Neurophysiol. 73 (6): 2563–2567. doi:10.1152 / jn.1995.73.6.2563. PMID 7666162.
- ^ Moran, D.W. & Schwartz, A.B. (1999). "Ulaşırken hız ve yönün motor kortikal gösterimi". J. Neurophysiol. 82 (5): 2676–2692. doi:10.1152 / jn.1999.82.5.2676. PMID 10561437.
- ^ Kakei, S., Hoffman, D. ve Strick, P (1999). "Birincil motor kortekste kas ve hareket gösterimleri". Bilim. 285 (5436): 2136–2139. CiteSeerX 10.1.1.137.8610. doi:10.1126 / science.285.5436.2136. PMID 10497133.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Todorov, E (2000). "Gönüllü kol hareketlerinde kas aktivasyonunun doğrudan kortikal kontrolü: bir model". Nat Neurosci. 3 (4): 391–398. doi:10.1038/73964. PMID 10725930. S2CID 13996279.
- ^ a b Haiss, F. ve Schwarz, C (2005). "Sıçan birincil motor korteksinin bıyık gösteriminde farklı hareket kontrolü modlarının mekansal ayrımı". J. Neurosci. 25 (6): 1579–1587. doi:10.1523 / JNEUROSCI.3760-04.2005. PMC 6726007. PMID 15703412.
- ^ Ramanathan, D., Conner, J.M. ve Tuszynski, M.H. (2006). "Beyin hasarından sonra işlevin geri kazanılmasıyla ilişkili bir motor kortikal esneklik biçimi". Proc. Natl. Acad. Sci. AMERİKA BİRLEŞİK DEVLETLERİ. 103 (30): 11370–11375. Bibcode:2006PNAS..10311370R. doi:10.1073 / pnas.0601065103. PMC 1544093. PMID 16837575.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Brecht, M., Schneider, M., Sakmann, B. ve Margrie, T.W. (2004). "Sıçan motor korteksindeki tek piramidal hücrelerin uyarılmasıyla uyandırılan bıyık hareketleri". Doğa. 427 (6976): 704–710. Bibcode:2004Natur.427..704B. doi:10.1038 / nature02266. PMID 14973477. S2CID 1105868.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ Cramer, N.P. & Keller, A (2006). "Çırpma yapan merkezi kalıp oluşturucunun kortikal kontrolü". J. Neurophysiol. 96 (1): 209–217. doi:10.1152 / jn.00071.2006. PMC 1764853. PMID 16641387.