Talamus - Thalamus
| Talamus | |
|---|---|
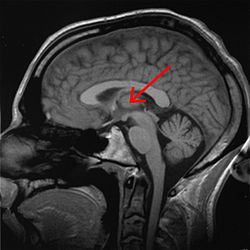 Talamus işaretli (MR enine kesit) | |
 anterolateral görünüm | |
| Detaylar | |
| Parçası | Diensefalon |
| Parçalar | Görmek Talamik çekirdeklerin listesi |
| Arter | Arka serebral arter ve şubeler |
| Tanımlayıcılar | |
| Latince | talamus dorsalis |
| MeSH | D013788 |
| NeuroNames | 300 |
| NeuroLex İD | birnlex_954 |
| TA98 | A14.1.08.101 A14.1.08.601 |
| TA2 | 5678 |
| TE | E5.14.3.4.2.1.8 |
| FMA | 62007 |
| Nöroanatominin anatomik terimleri | |
talamus (kimden Yunan θάλαμος, "bölme")[1] büyük bir kütle gri madde Içinde bulunan sırt bir bölümü diensefalon (bir bölümü ön beyin ). Sinir lifleri talamustan çıkıp ... beyin zarı her yönden hub benzeri bilgi alışverişi. Aktarma gibi çeşitli işlevleri vardır. duyusal dahil olmak üzere sinyaller motor serebral kortekse sinyaller[2][3][sayfa gerekli ] ve düzenlenmesi bilinç, uyku, ve uyanıklık.[4]
Anatomik olarak bir orta çizgi iki yarının (sol ve sağ) simetrik yapısı, omurgalı beyin, serebral korteks ve orta beyin. Sırasında oluşur embriyonik gelişme ilk olarak İsviçre tarafından tanınan diensefalonun ana ürünü olarak embriyolog ve anatomist Wilhelm His Sr. 1893'te.[5]
Anatomi
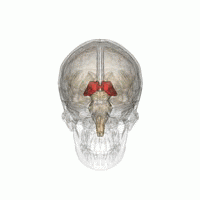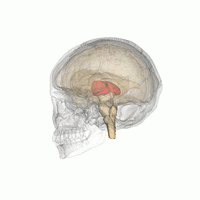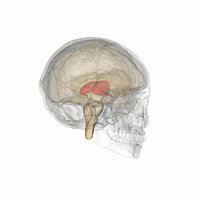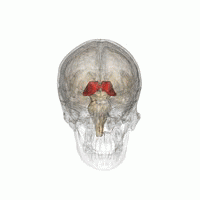
Talamus, içinde bulunan eşleştirilmiş bir gri madde yapısıdır. ön beyin orta beyinden daha üstün, beynin merkezine yakın, sinir lifleri her yönden serebral kortekse çıkıntı yapıyor. Talamusun medial yüzeyi, lateral duvarın üst kısmını oluşturur. üçüncü ventrikül ve karşı talamusun karşılık gelen yüzeyine düzleştirilmiş gri bir bantla bağlanır. intertalamik yapışma. Talamusun yan kısmı filogenetik olarak talamusun (neotalamus) en yeni kısmıdır ve lateral çekirdekleri içerir, pulvinar ve orta ve yanal genikülat çekirdekler.[6][7] Alanları var Beyaz madde dorsal yüzeyi kaplayan stratum zonale dahil talamusta ve dış ve iç medüller lamina. Dış lamina, lateral yüzeyi kaplar ve iç lamina, çekirdekleri ön, medial ve lateral gruplara ayırır.[8]
Kan temini
Talamus, kan kaynağını bir dizi arterden alır: polar arter (arka iletişim arter ), paramedian talamik-subtalamik arterler, inferolateral (talamojenikülat) arterler ve posterior (medial ve lateral) koroidal arterler.[9] Bunların hepsi, posterior serebral arter.[10]
Bazı insanların Percheron arteri Bu, talamusun her iki kısmını beslemek için arka serebral arterden tek bir arteriyel gövdenin çıktığı nadir bir anatomik varyasyondur.
Talamik çekirdekler




Türevleri diensefalon dorsal yerleşimli olanı dahil et epitalamus (esasen Habenula ve ekler) ve perithalamus (pretalamus) içeren Zona incerta ve talamik retiküler çekirdek. Farklı olmaları nedeniyle ontogenetik kökenler, epitalamus ve peritalamus resmen talamustan ayırt edilir. Metatalamus, lateral genikülat ve medial genikülat çekirdeklerden oluşur.
Talamus bir sistem içerir lameller (ondan yapılmış miyelinli lifler ) farklı talamik alt parçaların ayrılması. Diğer alanlar, farklı kümeler ile tanımlanır: nöronlar, benzeri periventriküler çekirdek, intralaminar elemanlar, "çekirdek sınırlayıcılar" ve diğerleri.[11] Talamusun büyük kısmından yapı olarak farklı olan bu son yapılar, birlikte gruplanmıştır. allothalamus aksine izotalamus.[12] Bu ayrım, talamusun genel tanımını basitleştirir.
Bağlantılar
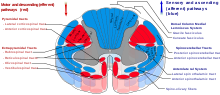
Talamusun hipokampusla birçok bağlantısı vardır. mamillothalamik yol, bu yol şunları içerir: memeliler ve Fornix.[13]
Talamus, beyin zarı aracılığıyla talamokortikal radyasyonlar.[14]
spinotalamik yol omurilikten kaynaklanan duyusal bir yoldur. Talamusa ağrı, sıcaklık, kaşıntı ve Kaba dokunuş. İki ana bölüm vardır: yanal spinotalamik yol, ağrı ve sıcaklığı ileten ve ön (veya ventral) spinotalamik yol, kaba dokunuş ve baskı ileten.
Fonksiyon
Talamusun, genellikle bir röle istasyonu olarak işlev gördüğüne inanılan birden çok işlevi vardır hub, farklı subkortikal alanlar arasında bilgi aktarımı ve beyin zarı.[15] Özellikle, her duyu sistemi ( koku alma sistemi ) duyusal sinyalleri alan ve bunları ilişkili birincil kortikal alana gönderen bir talamik çekirdek içerir.[kaynak belirtilmeli ] Görsel sistem için, örneğin, retina gönderiliyor yanal genikülat çekirdek talamusun görsel korteks içinde oksipital lob.[kaynak belirtilmeli ] Talamusun hem duyusal bilgiyi işlediğine hem de ilettiğine inanılır - birincil duyusal röle alanlarının her biri serebral korteksten güçlü geri besleme bağlantıları alır.[16] Benzer şekilde medial genikulat çekirdek anahtar görevi görür işitsel arasında geçiş alt kollikulus of orta beyin ve birincil işitsel korteks.[kaynak belirtilmeli ] ventral arka çekirdek bir anahtar somatosensoriyel dokunma gönderen röle ve propriyoseptif bilgi birincil somatosensoriyel korteks.[kaynak belirtilmeli ]
Talamus aynı zamanda durumların düzenlenmesinde önemli bir rol oynar. uyku ve uyanıklık.[17] Talamik çekirdekler, serebral korteks ile güçlü karşılıklı bağlantılara sahiptir. talamo-kortiko-talamik devreler ile ilgili olduğuna inanılan bilinç.[18] Talamus, uyarılmayı, farkındalık seviyesini ve aktiviteyi düzenlemede önemli bir rol oynar. Talamustaki hasar kalıcılığa neden olabilir koma.[19]
Talamusun daha öndeki rolü solgun ve zenci bölgeleri Bazal ganglion sistem bozuklukları fark ediliyor, ancak hala tam olarak anlaşılamıyor. Talamusun vestibülere veya tektal işlevler neredeyse yok sayılır. Talamus, basitçe sinyalleri serebral kortekse ileten bir "röle" olarak düşünülmüştür. Daha yeni araştırmalar talamik fonksiyonun daha seçici olduğunu göstermektedir.[20] Talamusun çeşitli bölgelerine birçok farklı işlev bağlıdır. Bu, birçok duyu sistemi için geçerlidir (koku alma sistemi hariç), örneğin işitsel, somatik, içgüdüsel, tat alma ve görsel sistemler lokalize lezyonların spesifik duyu eksikliklerine neden olduğu yerlerde. Talamusun önemli bir rolü, motor ve dil sistemlerini desteklemektir ve bu sistemler için kullanılan devrelerin çoğu paylaşılır. Talamus işlevsel olarak bağlı için hipokamp[21] talamik anterior çekirdeklerde genişletilmiş hipokampal sistemin bir parçası olarak[22] uzamsal bellek ve uzamsal duyusal veriyle ilgili olarak, bunlar insan epizodik belleği ve kemirgen olay belleği için çok önemlidir.[23][24] Meziyo-temporal lobun belirli kısımlarına bağlanan talamik bölgelerin, hatırlama ve aşinalık hafızasının işleyişinin farklılaşmasını sağladığı hipotezi için destek vardır.[13]
Motor kontrol için gerekli nöronal bilgi süreçleri, talamusu bir subkortikal motor merkezi olarak içeren bir ağ olarak önerildi.[25] Primatların beyinlerinin anatomisinin araştırılmasıyla[26] birbirine bağlı dokuların doğası beyincik çoklu motor kortekslere göre, talamusun bazal gangliyonlardan belirli kanalları sağlamada anahtar bir işlevi yerine getirdiğini ve beyincik kortikal motor bölgelerine.[27][28] Bir araştırmada sakkad ve takas karşıtı[29] Üç maymunda motor yanıt, talamik bölgelerin, kazaya karşı göz hareketinin (yani, sunulan bir uyarıcı doğrultusunda gözlerin refleksif sarsılma hareketini inhibe etme yeteneği) oluşumunda rol oynadığı bulundu.[30]
Son araştırmalar, mediodorsal talamusun bilişte daha geniş bir rol oynayabileceğini düşündürmektedir. Spesifik olarak, mediodorsal talamus "sadece korteksteki devrelerin bağlantısını (sinyal gücü) mevcut bağlama uygun olarak güçlendirebilir ve böylece kararların hangi birçok ilişkiyi bağlayarak karmaşık kararlar alma esnekliğine (memeli beyninin) katkıda bulunabilir. zayıf bağlanmış kortikal devrelere bağlıdır. "[31] Araştırmacılar, "MD aktivitesini arttırmanın, farelerin" düşünme "yeteneğini artırdığını keşfetti.[31] Ödülü bulmak için hangi çelişkili duyusal uyaranı izleyeceklerine karar verirken hata oranlarını yüzde 25'ten fazla düşürmek. " [32]
Geliştirme
Talamik kompleks, peritalamustan (veya daha önce ventral talamus olarak da bilinen pretalamustan), orta diensefalik düzenleyiciden (daha sonra oluşan zona limitans intratalamika (ZLI)) ve talamus (dorsal talamus).[33][34] Talamusun gelişimi üç aşamaya ayrılabilir.[35]Talamus, embriyonikten türeyen en büyük yapıdır. diensefalon ön beyin ile beyin arasında yer alan arka kısım.
Erken beyin gelişimi
Sonra sinirlenme anlage pretalam ve talamus nöral tüp içinde indüklenir. Farklı omurgalı model organizmalardan elde edilen veriler, ikisi arasındaki etkileşimin olduğu bir modeli destekler Transkripsiyon faktörleri, Fez ve Otx, belirleyici bir öneme sahiptir. Fez, pretalamusta ifade edilir ve fonksiyonel deneyler, Fez'in pretalam oluşumu için gerekli olduğunu gösterir.[36][37] Posterior olarak, Otx1 ve Otx2, Fez'in ekspresyon alanına bitişiktir ve talamusun düzgün gelişimi için gereklidir.[38][39]
Progenitör alanların oluşumu
Talamik gelişimin erken dönemlerinde iki progenitör alan oluşur, bir kaudal alan ve bir rostral alan. Kaudal alan, yetişkin talamustaki tüm glutamaterjik nöronlara yol açarken, rostral alan yetişkin talamustaki tüm GABAerjik nöronlara yol açar.[40]
Orta diensefalik düzenleyicinin (MDO) oluşumu
Fez ve Otx'in ifade alanları arasındaki arayüzde, orta diensefalik düzenleyici (MDO, aynı zamanda ZLI düzenleyici olarak da adlandırılır) talamik içinde indüklenir. Anlage. MDO, talamustaki merkezi sinyal düzenleyicisidir. Organizatörün eksikliği talamusun yokluğuna yol açar. MDO, geliştirme sırasında ventralden dorsale doğru olgunlaşır. Üyeleri SHH aile ve Wnt ailesi, MDO tarafından yayılan ana temel sinyallerdir.
Organizatör, sinyal merkezi olarak öneminin yanı sıra, projenin morfolojik yapısı içinde olgunlaşır. zona limitans intratalamika (ZLI).
Talamusun olgunlaşması ve parselasyonu
MDO, indüksiyonundan sonra, talamik anajın gelişimini aşağıdaki gibi sinyal molekülleri salgılayarak düzenlemeye başlar. SHH.[41] Farelerde, MDO'daki sinyalleşme işlevi, tamamen yokluğundan dolayı doğrudan ele alınmamıştır. diensefalon SHH mutantlarında.[42]
Civcivlerde yapılan araştırmalar SHH'nin talamik gen indüksiyonu için hem gerekli hem de yeterli olduğunu göstermiştir.[43] İçinde zebra balığı, iki SHH geninin, SHH-a ve SHH-b'nin (önceden twhh olarak tanımlanmıştır) ekspresyonunun MDO bölgesini işaret ettiği ve SHH sinyallemesinin hem pretalamus hem de talamusun moleküler farklılaşması için yeterli olduğu ancak yeterli olmadığı gösterilmiştir. bakım için gerekli ve MDO / alarm plakasından SHH sinyali, ventral Shh sinyalleri verilmezken pretalamik ve talamik bölgenin olgunlaşması için yeterlidir.[44]
SHH'ye maruz kalma talamik nöronların farklılaşmasına yol açar. MDO'dan gelen SHH sinyallemesi, proneural genin posterior-anterior ekspresyon dalgasını indükler Nörojen1 talamusun ana (kaudal) kısmında ve MDO'nun hemen bitişiğindeki rostral talamik hücrelerin kalan dar şeridinde Ascl1 (eski adıyla Mash1) ve pretalamusta.[45][46]
Proneural gen ekspresyonunun bu bölgelendirilmesi, glutamaterjik röle nöronlarının Neurogenin1 + öncülerinden ve GABAerjik inhibe edici nöronların Ascl1 + öncülerinden farklılaşmasına yol açar. Balıklarda, bu alternatif nörotransmiter kaderlerinin seçimi, homolog Her6'nın dinamik ifadesi tarafından kontrol edilir. HES1. Bu tüylü bHLH'nin ifadesi transkripsiyon faktörü Neurogenin'i baskılayan ancak Ascl1 için gerekli olan, kaudal talamustan aşamalı olarak kaybolur, ancak pretalamusta ve rostral talamik hücrelerin şeridinde tutulur. Ek olarak, civciv ve fareler üzerinde yapılan çalışmalar, Shh yolunu bloke etmenin rostral talamus yokluğuna ve kaudal talamusta önemli ölçüde azalmaya yol açtığını göstermiştir. Rostral talamus retiküler çekirdeğe yol açacak ve burada kaudal talamus röle talamusu oluşturacak ve daha da alt bölümlere ayrılacaktır. talamik çekirdekler.[35]
İnsanlarda, hastalığın promotör bölgesinde yaygın bir genetik varyasyon serotonin taşıyıcısı (SERT-uzun ve kısa alel: 5-HTTLPR ) yetişkinlerde talamusun çeşitli bölgelerinin gelişimini etkilediği gösterilmiştir. İki kısa miras kalan insanlar aleller (SERT-ss) daha fazla nörona ve daha büyük bir hacme sahiptir. pulvinar ve muhtemelen talamusun limbik bölgeleri. Talamusun genişlemesi, iki SERT-s alelini miras alan kişilerin neden daha savunmasız olduğuna dair anatomik bir temel sağlar. majör depresyon, travmatik stres bozukluğu sonrası ve intihar.[47]
Klinik önemi
Serebrovasküler bir kaza (inme ) yol açabilir talamik ağrı sendromu,[48] Tek taraflı bir yanma veya ağrı hissi içeren, sıklıkla eşlik eden ruh hali. İkili iskemi Paramedian arter tarafından sağlanan alanın, aşağıdakiler dahil ciddi sorunlara neden olabilir akinetik mutizm ve eşlik edecek okülomotor sorunlar. İlgili bir kavram talamokortikal disritmi. Oklüzyon Percheron arteri iki taraflı talamus enfarktüsüne yol açabilir.
Alkolik Korsakoff sendromu hasardan kaynaklanıyor memeli vücut, mamillothalamic fasciculus veya talamus.[49][50]
Ölümcül ailesel uykusuzluk kalıtsal Prion talamusun dejenerasyonunun meydana geldiği, hastanın yavaş yavaş uyku yeteneğini kaybetmesine ve toplam bir duruma ilerlemesine neden olan hastalık uykusuzluk hastalığı, bu her zaman ölüme yol açar. Aksine, talamustaki hasar, koma.
Ek resimler

Talamusu gösteren insan beyin diseksiyonu.

İnsan talamusu diğer subkortikal yapılarla birlikte cam beyinde.

Talamik çekirdeklerin yanal grubu.

Talamik çekirdeklerin medial grubu.




Ayrıca bakınız
- 5-HT7 reseptörü
- Krista ve Tatiana Hogan - birleşik thalami ile yapışık ikizler
- İnsan beynindeki bölgelerin listesi
- Ventral nükleer grubun motor olmayan bölgesi talamusun
- Primat bazal gangliya sistemi
- Talamik uyarıcı
- Talamotomi
Referanslar
- ^ Harper - indeks & Washington Üniversitesi Fakülte Web Sunucusu & Arama motoru arama sayfası + Perseus Projesi tufts.edu Erişim tarihi: 2012-02-09
- ^ Sherman, S. (2006). "Talamus". Scholarpedia. 1 (9): 1583. Bibcode:2006SchpJ ... 1.1583S. doi:10.4249 / bilim adamı. 1583.
- ^ Sherman, S. Murray; Guillermo, R.W. (2000). Talamusu keşfetmek. Akademik Basın. ISBN 978-0-12-305460-9.
- ^ Gorvett, Zaria. "Einstein'ın ilginç alışkanlıklarından öğrenebilecekleriniz". bbc.com.
- ^ Jones, Edward G, ed. (1985). Talamus - Springer. doi:10.1007/978-1-4615-1749-8. ISBN 978-1-4613-5704-9. S2CID 41337319.
- ^ "NEOTHALAMUS'un Tıbbi Tanımı". www.merriam-webster.com.
- ^ "neotalamus | Oxford Sözlükleri tarafından İngilizce'de neotalamus tanımı". Oxford Sözlükleri | ingilizce.
- ^ Tortora, Gerard; Anagnostakos, Nicholas (1987). Anatomi ve fizyolojinin ilkeleri (5. Harper uluslararası ed.). New York: Harper & Row. s.314. ISBN 978-0060466695.
- ^ Percheron, G. (1982). "Talamusun arteriyel kaynağı". Schaltenbrand'da; Walker, A. E. (editörler). İnsan beyninin stereotaksi. Stuttgart: Thieme. s. 218–32.
- ^ Knipe, H Jones, J vd. Talamus http://radiopaedia.org/articles/thalamus
- ^ Jones Edward G. (2007) "The Thalamus" Cambridge Üni. Basın[sayfa gerekli ]
- ^ Percheron, G. (2003). "Talamus". Paxinos, G .; May, J. (editörler). İnsan sinir sistemi (2. baskı). Amsterdam: Elsevier. s. 592–675.
- ^ a b Carlesimo, GA; Lombardi, MG; Caltagirone, C (2011). "Vasküler talamik amnezi: Yeniden değerlendirme". Nöropsikoloji. 49 (5): 777–89. doi:10.1016 / j.neuropsychologia.2011.01.026. PMID 21255590. S2CID 22002872.
- ^ Washington Üniversitesi (1991). "Talamokortikal radyasyonlar". washington.edu.
- ^ Gazzaniga; İvry; Mangun, Michael, S .; Richard B .; George R. (2014). Bilişsel Sinirbilim - Zihnin Biyolojisi. New York: W.W. Norton. pp.45. ISBN 978-0-393-91348-4.CS1 Maint: birden çok isim: yazarlar listesi (bağlantı)
- ^ "Beynin aracısı olan talamus, duyu iletkeni haline gelir". Chicago Tıp Üniversitesi. Alındı 10 Eylül 2020.
- ^ Steriade, Mircea; Llinás, Rodolfo R. (1988). "Talamusun İşlevsel Durumları ve İlişkili Nöronal Etkileşim". Fizyolojik İncelemeler. 68 (3): 649–742. doi:10.1152 / physrev.1988.68.3.649. PMID 2839857.
- ^ Koma ve Bilinç Bozuklukları ISBN 978-1-447-12439-9 s. 143
- ^ Bilinç Nörolojisi: Bilişsel Sinirbilim ve Nöropatoloji ISBN 978-0-123-74168-4 s. 10
- ^ Leonard, Abigail W. (17 Ağustos 2006). "Beyniniz Bir Bilgisayar Gibi Başlıyor". LiveScience.
- ^ Stein, Thor; Moritz, Çad; Quigley, Michelle; Cordes, Dietmar; Haughton, Victor; Meyerand, Elizabeth (2000). "Fonksiyonel MR Görüntüleme ile Çalışılan Talamus ve Hipokampusta Fonksiyonel Bağlantı". Amerikan Nöroradyoloji Dergisi. 21 (8): 1397–401. PMID 11003270.
- ^ Aggleton, John P .; Brown, Malcolm W. (1999). "Epizodik bellek, amnezi ve hipokampal-anterior talamik eksen" (PDF). Davranış ve Beyin Bilimleri. 22 (3): 425–44, tartışma 444–89. doi:10.1017 / S0140525X99002034. PMID 11301518.
- ^ Aggleton, John P .; O'Mara, Shane M .; Vann, Seralynne D .; Wright, Nick F .; Tsanov, Marian; Erichsen Jonathan T. (2010). "Bellek için hipokampal-anterior talamik yollar: Doğrudan ve dolaylı eylemler ağını ortaya çıkarmak". Avrupa Nörobilim Dergisi. 31 (12): 2292–307. doi:10.1111 / j.1460-9568.2010.07251.x. PMC 2936113. PMID 20550571.
- ^ Burgess, Neil; Maguire, Eleanor A; O'Keefe, John (2002). "İnsan Hipokampusu ve Uzaysal ve Epizodik Bellek". Nöron. 35 (4): 625–41. doi:10.1016 / S0896-6273 (02) 00830-9. PMID 12194864. S2CID 11989085.
- ^ Evarts, E V; Thach, WT (1969). "CNS'nin Motor Mekanizmaları: Serebroserebellar İlişkiler". Yıllık Fizyoloji İncelemesi. 31: 451–98. doi:10.1146 / annurev.ph.31.030169.002315. PMID 4885774.
- ^ Orioli, PJ; Strick, PL (1989). "Motor korteks ve arkuat premotor alan ile serebellar bağlantılar: WGA-HRP'nin retrograd transnöronal taşınmasını kullanan bir analiz". Karşılaştırmalı Nöroloji Dergisi. 288 (4): 612–26. doi:10.1002 / cne.902880408. PMID 2478593. S2CID 27155579.
- ^ Asanuma C, Thach WT, Jones EG (Mayıs 1983). "Maymunda ventral lateral talamik bölgenin sitoarkitektonik tanımı". Beyin Araştırması. 286 (3): 219–35. doi:10.1016/0165-0173(83)90014-0. PMID 6850357. S2CID 25013002.
- ^ Kurata, K (2005). "Motor talamustaki nöronların, maymunlarda kortikal motor alanlara yansıyan aktivite özellikleri ve yeri". Nörofizyoloji Dergisi. 94 (1): 550–66. doi:10.1152 / jn.01034.2004. PMID 15703228.
- ^ http://www.optomotorik.de/blicken/anti-rev.htm[tam alıntı gerekli ]
- ^ Kunimatsu, J; Tanaka, M (2010). "Antisaccades oluşumunda primat motor talamusun rolleri" (PDF). Nörobilim Dergisi. 30 (14): 5108–17. doi:10.1523 / JNEUROSCI.0406-10.2010. PMC 6632795. PMID 20371831.
- ^ a b "Beyin Bölgesi İçin Yeni Rol Keşfedildi". Nörobilim Haberleri. 2017-05-03. Alındı 2017-12-03.
- ^ Schmitt, L. Ian; Wimmer, Ralf D .; Nakajima, Miho; Happ, Michael; Mofakham, Sima; Halassa, Michael M. (11 Mayıs 2017). "Kortikal bağlantının talamik amplifikasyonu dikkatli kontrol sağlar". Doğa. 545 (7653): 219–223. Bibcode:2017Natur.545..219S. doi:10.1038 / nature22073. ISSN 1476-4687. PMC 5570520. PMID 28467827.
- ^ Kuhlenbeck, Hartwig (1937). "Bir kuşun beynindeki (civciv) diensefalik merkezlerin ontogenetik gelişimi ve sürüngen ve memeli diensefalonuyla karşılaştırma". Karşılaştırmalı Nöroloji Dergisi. 66: 23–75. doi:10.1002 / cne.900660103. S2CID 86730019.
- ^ Shimamura, K; Hartigan, DJ; Martinez, S; Puelles, L; Rubenstein, JL (1995). "Ön nöral plakanın ve nöral tüpün uzunlamasına organizasyonu". Geliştirme. 121 (12): 3923–33. PMID 8575293.
- ^ a b Scholpp, Steffen; Lumsden Andrew (2010). "Gelin odası inşa etmek: Talamusun gelişimi". Sinirbilimlerindeki Eğilimler. 33 (8): 373–80. doi:10.1016 / j.tins.2010.05.003. PMC 2954313. PMID 20541814.
- ^ Hirata, T .; Nakazawa, M; Muraoka, O; Nakayama, R; Suda, Y; Hibi, M (2006). "Diensefalon alt bölümlerinin oluşturulmasında çinko parmak genleri Fez ve Fez benzeri işlev görür". Geliştirme. 133 (20): 3993–4004. doi:10.1242 / dev.02585. PMID 16971467.
- ^ Jeong, J.-Y .; Einhorn, Z .; Mathur, P .; Chen, L .; Lee, S .; Kawakami, K .; Guo, S. (2007). "Zebra balığı diensefalonunun korunmuş çinko parmak proteini Fezl tarafından şekillendirilmesi". Geliştirme. 134 (1): 127–36. doi:10.1242 / dev.02705. PMID 17164418.
- ^ Acampora, D; Avantaggiato, V; Tuorto, F; Simeone, A (1997). "Otx gen dozaj gereksinimi yoluyla beyin morfogenezinin genetik kontrolü". Geliştirme. 124 (18): 3639–50. PMID 9342056.
- ^ Scholpp, S .; Foucher, I .; Staudt, N .; Peukert, D .; Lumsden, A .; Houart, C. (2007). "Otx1l, Otx2 ve Irx1b ZLI'yi diensefalonda kurar ve konumlandırır". Geliştirme. 134 (17): 3167–76. doi:10.1242 / dev.001461. PMC 7116068. PMID 17670791.
- ^ Şarkı, Hobeom; Lee, Bumwhee; Pyun, Dohoon; Guimera, Jordi; Oğlu, Youngsook; Yoon, Jaeseung; Baek, Kwanghee; Wurst, Wolfgang; Jeong, Yongsu (2015-02-15). "Ascl1 ve Helt, Dlxs aktivasyonunu baskılayarak talamik nöronal kimliği belirlemek için kombinasyonel olarak hareket eder". Gelişimsel Biyoloji. 398 (2): 280–291. doi:10.1016 / j.ydbio.2014.12.003. ISSN 1095-564X. PMID 25512300.
- ^ Puelles, L; Rubenstein, JL (2003). "Ön beyin gen ifade alanları ve gelişen prosomerik model". Sinirbilimlerindeki Eğilimler. 26 (9): 469–76. doi:10.1016 / S0166-2236 (03) 00234-0. PMID 12948657. S2CID 14658562.
- ^ Ishibashi, M; McMahon, AP (2002). "Bir sonik kirpi bağımlı sinyal rölesi, erken fare embriyosunda diensefalik ve mezensefalik primordianın büyümesini düzenler". Geliştirme. 129 (20): 4807–19. PMID 12361972.
- ^ Kiecker, C; Lumsden, A (2004). "ZLI'den gelen kirpi sinyalleri diensefalik bölgesel kimliği düzenler". Doğa Sinirbilim. 7 (11): 1242–9. doi:10.1038 / nn1338. PMID 15494730. S2CID 29863625.
- ^ Scholpp, S .; Wolf, O; Marka, M; Lumsden, A (2006). "Zona limitans intratalamika'dan gelen kirpi sinyali, zebra balığı diensefalonunun desenini düzenler". Geliştirme. 133 (5): 855–64. doi:10.1242 / dev.02248. PMID 16452095.
- ^ Scholpp, S .; Delogu, A .; Gilthorpe, J .; Peukert, D .; Schindler, S .; Lumsden, A. (2009). "Her6 talamustaki nörojenetik gradyanı ve nöronal kimliği düzenler". Ulusal Bilimler Akademisi Bildiriler Kitabı. 106 (47): 19895–900. Bibcode:2009PNAS..10619895S. doi:10.1073 / pnas.0910894106. PMC 2775703. PMID 19903880.
- ^ Vue, Tou Yia; Bluske, Krista; Alishahi, Amin; Yang, Lin Lin; Koyano-Nakagawa, Naoko; Novitch, Bennett; Nakagawa, Yasushi (2009). "Sonic Hedgehog Sinyali, Farelerde Talamik Progenitör Kimliğini ve Çekirdek Spesifikasyonunu Kontrol Ediyor". Nörobilim Dergisi. 29 (14): 4484–97. doi:10.1523 / JNEUROSCI.0656-09.2009. PMC 2718849. PMID 19357274.
- ^ Young, Keith A .; Holcomb, Leigh A .; Bonkale, Willy L .; Hicks, Paul B .; Yazdani, Umar; Almanca, Dwight C. (2007). "5HTTLPR Polimorfizmi ve Pulvinarın Genişlemesi: Arka Kapının Limbik Sisteme Açılması". Biyolojik Psikiyatri. 61 (6): 813–8. doi:10.1016 / j.biopsych.2006.08.047. PMID 17083920. S2CID 2214561.
- ^ Dejerine, J .; Roussy, G. (1906). "Le sendromu talamik". Revue Neurologique. 14: 521–32.
- ^ Kopelman, MD; Thomson, AD; Guerrini, I; Marshall, EJ (NaN). "Korsakoff sendromu: klinik yönler, psikoloji ve tedavi". Alkol ve Alkolizm (Oxford, Oxfordshire). 44 (2): 148–54. doi:10.1093 / alcalc / agn118. PMID 19151162. Tarih değerlerini kontrol edin:
| tarih =(Yardım) - ^ Rahme, R; Moussa, R; Awada, A; İbrahim, ben; Ali, Y; Maarrawi, J; Rizk, T; Nohra, G; Okais, N; Samaha, E (Nisan 2007). "Sağ hipokampal kanamayı takiben sol talamik enfarktüsten kaynaklanan akut Korsakoff benzeri amnestik sendrom". AJNR. Amerikan Nöroradyoloji Dergisi. 28 (4): 759–60. PMID 17416834.
Dış bağlantılar
| Yetki kontrolü |
|---|