Ankylosaurus - Ankylosaurus - Wikipedia
| Ankylosaurus | |
|---|---|
 | |
| Oyuncular Ankylosaurus kafatası (AMNH 5214) önden görünümde, Rockies Müzesi | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Sipariş: | †Ornithischia |
| Aile: | †Ankylosauridae |
| Kabile: | †Ankylosaurini |
| Cins: | †Ankylosaurus Kahverengi, 1908 |
| Türler: | †A. magniventris |
| Binom adı | |
| †Ankylosaurus magniventris Kahverengi, 1908 | |
Ankylosaurus[nb 1] bir cins nın-nin zırhlı dinozor. Onun fosiller bulundu jeolojik oluşumlar en sonuna kadar çıkmak Kretase Periyot, yaklaşık 68–66 milyon yıl önce, Batı Kuzey Amerika'da, onu kuş olmayanların sonuncusu dinozorlar. Tarafından adlandırıldı Barnum Brown 1908'de; cinsteki tek tür A. magniventris. Cins adı "erimiş kertenkele" anlamına gelir ve özel ad "büyük göbek" anlamına gelir. Bugüne kadar bir avuç örnek kazıldı, ancak tam bir iskelet keşfedilmedi. Ankylosauria'nın diğer üyeleri daha kapsamlı fosil materyalleriyle temsil edilse de, Ankylosaurus genellikle kabul edilir arketipik sıradışı özelliklere sahip olmasına rağmen grubunun üyesi.
Muhtemelen en çok bilinen ankilozorid, Ankylosaurus 6 ila 8 metre (20 ila 26 ft) uzunluğunda olduğu ve 4,8 ila 8 metrik ton (5,3 ila 8,8 kısa ton) arasında ağırlığa sahip olduğu tahmin edilmektedir. Öyleydi dört ayaklı, geniş, sağlam bir gövdeye sahip. Geniş, alçak bir kafatasına sahipti, iki boynuzu başın arkasından geriye doğru işaret ediyordu ve bunların altında geriye ve aşağıyı gösteren iki boynuz vardı. Diğer ankylosaurlardan farklı olarak, burun delikleri öne değil yana doğru bakıyordu. Çenelerin ön kısmı bir gagayla kaplıydı ve arkasında küçük, yaprak şeklinde dişler vardı. Boynu kaplayan kemikli yarım halkalarla zırh plakaları veya osteodermlerle kaplıydı ve büyük bir kulüp kuyruğunun ucunda. Kafatasındaki ve vücudun diğer bölümlerindeki kemikler kaynaşarak güçlerini arttırır ve bu özellik cins isminin kaynağıdır.
Ankylosaurus Ankylosauridae ailesinin bir üyesidir ve en yakın akrabaları görünüşe göre Anodontozorlar ve Euoplocephalus. Ankylosaurus Yavaş hareket eden bir hayvan olduğu, gerektiğinde hızlı hareket edebildiği düşünülmektedir. Geniş ağzı, seçici olmadığını gösterir. tarayıcı. Sinüsler ve burun içindeki burun odaları ısı ve su dengesi için olabilir veya seslendirmede rol oynamış olabilir. Kuyruk kulübünün avcılara karşı savunmada veya tür içi mücadele. Ankylosaurus içinde bulundu Hell Creek, Lance, Scollard, Fransız, ve Ferris ancak çevresinde ender görülüyor. Yanında yaşamasına rağmen nodosaurid ankylosaur, aralıkları ve Ekolojik nişler çakışmış gibi görünmüyor ve Ankylosaurus yüksek arazilerde yaşamış olabilir. Ankylosaurus ayrıca dinozorların yanında yaşadı Tyrannosaurus, Triceratops, ve Edmontosaurus.
Keşif tarihi

1906'da bir Amerikan Doğa Tarihi Müzesi Amerikalı liderliğindeki sefer paleontolog Barnum Brown keşfetti tip numune nın-nin Ankylosaurus magniventris (AMNH 5895) içinde Hell Creek Oluşumu Gilbert Creek yakınında, Montana. Örnek (toplayıcı Peter Kaisen tarafından bulundu) bir kafatasının üst kısmı, iki diş, omuz kuşağının bir kısmı, servikal, dorsal ve kaudal omurlar, kaburgalar ve otuzdan fazla osteodermler (zırh plakaları). Bilimsel olarak kahverengi tarif 1908'deki hayvan; cins adı türetilmiştir Yunan kelimeler αγκυλος Ankulos ('bükülmüş' veya 'çarpık'), tıbbi terime atıfta bulunarak ankiloz, kafatası ve vücuttaki kemiklerin füzyonundan kaynaklanan sertlik ve σαυρος Sauros ('kertenkele'). İsim, "kaynaşmış kertenkele", "sert kertenkele" veya "kıvrımlı kertenkele" olarak tercüme edilebilir. türler isim Magniventris türetilmiştir Latince: magnus ('harika ve Latince: Venter ('göbek'), hayvanın vücudunun büyük genişliğine atıfta bulunur.[2][3][4]

1908 açıklamasına eşlik eden iskelet rekonstrüksiyonu, eksik parçaları benzer bir şekilde restore etti. Stegosaurus ve Brown sonucu soyu tükenmiş zırhlı memeliye benzetti Glyptodon.[2] Modern tasvirlerin aksine, Brown'un stegosaur benzeri rekonstrüksiyonu sağlam ön ayakları, kuvvetli kavisli bir sırt, ilium ve pubisten öne doğru çıkıntı yapan sivri uçlu bir pelvis ve kısa, sarkık bir kuyruk gösterdi. kuyruk kulübü o zamanlar bilinmiyordu. Brown ayrıca zırh plakalarını arkadan aşağı doğru paralel sıralar halinde yeniden inşa etti; bu düzenleme tamamen varsayımsaldı. Brown'ın rekonstrüksiyonu oldukça etkili oldu ve onun diyagramına dayalı olarak hayvanın restorasyonları 1980'lerin sonlarında yayınlandı.[5][6][7] Brown'ın 1908 tarihli bir incelemesinde Ankylosaurus Amerikan paleontolog Samuel Wendell Williston iskelet rekonstrüksiyonunu çok az kalıntıya dayandığı için eleştirdi ve Ankylosaurus sadece cinsin eşanlamlısıydı Stegopelta Williston, 1905'te adını vermişti. Williston ayrıca ilgili iskeletin bir iskelet rekonstrüksiyonu olduğunu belirtti. Polacanthus Macar paleontolog tarafından Franz Nopcsa daha iyi bir örnekti Ankylosaurlar hayatta ortaya çıkacaktı.[8] Eşanlamlılık iddiası diğer araştırmacılar tarafından kabul edilmedi ve iki cins artık farklı kabul ediliyor.[9]
Brown, bir kazı yaparken 77 osteoderm toplamıştı. Tyrannosaurus içindeki örnek Lance Formasyonu Wyoming'den 1900'de. Bu osteodermlerden (örnek AMNH 5866) bahsetti. Ankylosaurus ama onların ait olduğunu düşündü Tyrannosaurus yerine. Paleontolog Henry Fairfield Osborn ayrıca bu görüşü, Tyrannosaurus şimdi olduğu gibi örnek eşanlamlı cins Dinamozor Daha yakın zamanda yapılan incelemeler, bunların, Ankylosaurus; Brown onları bazılarıyla karşılaştırmış görünüyor Euoplocephalus Ait olduğu hatalı şekilde kataloglanmış olan osteodermler Ankylosaurus AMNH'de.[10][11]

1910'da Brown liderliğindeki başka bir AMNH keşif gezisi, Ankylosaurus numune (AMNH 5214) içinde Scollard Oluşumu tarafından Red Deer Nehri Alberta, Kanada'da. Bu örnek, tam bir kafatası, çeneler, bu cinsten bilinen ilk ve tek kuyruk klübünün yanı sıra kaburgalar, omurlar, uzuv kemikleri ve zırhı içeriyordu. 1947'de Amerikalı fosil toplayıcıları Charles M. Sternberg ve T. Potter Chamney, 1910 örneğinin bulunduğu yerin 1 kilometre (0,6 mil) kuzeyinde bir kafatası ve çene (örnek CMN 8880, eski adıyla NMC 8880) topladı. Bu bilinen en büyük Ankylosaurus kafatası, ancak yer yer hasar görmüş. Kaudal omurların bir bölümü (örnek CCM V03) 1960'larda Powder Nehri drenaj, Montana, Hell Creek Formasyonunun bir parçası. Bu beş tamamlanmamış örneğe ek olarak, diğer birçok izole osteoderm ve diş bulunmuştur.[12][10]
1990'da Amerikalı paleontolog Walter P. Coombs iki kafatasının dişlerinin atandığına dikkat çekti A. magniventris bazı ayrıntılarda holotip örneğindekilerden farklıydı ve yeni bir türü adlandırmak için "düşünceli bir cazibesini" ifade etmesine rağmen Ankylosaurus bunlar için, türlerdeki varyasyon aralığı tam olarak belgelenmediği için bunu yapmaktan kaçındı. Ayrıca, holotip örneğiyle ilişkili iki dişin, içinde bulundukları gibi, belki de ona ait olmadıkları olasılığını da gündeme getirdi. matris burun odaları içinde.[13] Amerikalı paleontolog Kenneth Carpenter dişlerin ait olduğunu kabul etti A. magniventris 2004'te tüm örneklerin aynı türe ait olduğunu ve diğer ankylosaurların dişlerinin oldukça değişken olduğunu kaydetti.[10]

Bilinenlerin çoğu Ankylosaurus Bazı paleontologlar, Carpenter 2004 yılında cinsi yeniden tanımlayana kadar, numuneler bilimsel olarak uzun uzun tarif edilmemişti. Ankylosaurus oldu arketipik grubunun üyesi ve en iyi bilinen ankylosaur popüler kültür, belki de salonda gösterilen hayvanın yaşam boyu yeniden yapılandırılması nedeniyle 1964 Dünya Fuarı New York'ta.[10] O heykel ve Amerikalı sanatçı Rudolph Zallinger 1947 duvar resmi Sürüngenler Çağı ve daha sonra popüler olan diğer tasvirler Ankylosaurus 1910 yılında bu özelliğin ilk keşfinin ardından bir kuyruk kulübü ile. Aşinalığına rağmen, en yakın akrabalarına göre çok daha az kalıntıyla bilinmektedir. 2017'de Kanadalı paleontologlar Victoria M. Arbor ve Jordan Mallon, daha önce literatürde bahsedilmeyen holotip unsurları (kafatasının parçaları ve boyun yarım halkaları gibi) dahil olmak üzere yeni ankilozor keşifleri ışığında cinsi yeniden tanımladı. Yine de sonuca vardılar Ankylosaurus ikonik ve grubunun en tanınmış üyesidir, ilgili ankilozorlara kıyasla tuhaftı ve bu nedenle grubu temsil etmedi.[12]
Birçok geleneksel popüler tasvir şov Ankylosaurus çömelme duruşunda ve büyük bir kuyruk sopası yere sürüklenirken. Modern rekonstrüksiyonlar, hayvanı daha dik bir uzuv duruşu ile ve kuyruğu yerden tutulmuş olarak gösterir. Aynı şekilde, vücuttan yana doğru çıkıntı yapan büyük dikenler (dikenlerinkine benzer) nodosaurid ankylosaurs) birçok geleneksel tasvirde bulunur, ancak Ankylosaurus kendisi.[5] Zırhı Ankylosaurus sık sık Edmontoni (daha önce Palaeoscincus ); ek olarak Ankylosaurus sivri uçlarla tasvir edilmek, Edmontoni ayrıca bir ile tasvir edilmiştir Ankylosaurus-Amerikan ressamın bir duvar resmi de dahil olmak üzere, kuyruk kulübü (nodosauridlerin sahip olmadığı bir özellik) Charles R. Şövalye 1930'dan itibaren.[12]
Açıklama

Ankylosaurus bilinen en büyüğüydü ankilozorin dinozor ve muhtemelen en büyüğü ankilozorid.[12] 2004 yılında Carpenter, 64.5 santimetre (2.12 ft) uzunluğunda ve 74.5 santimetre (2.44 ft) genişliğindeki bilinen en büyük kafatasına (örnek CMN 8880) sahip bireyin yaklaşık 6.25 metre (20.5 ft) uzunluğunda ve bir kalçaya sahip olduğunu tahmin etti. yaklaşık 1,7 metre (5,6 ft) yükseklik. Bilinen en küçük kafatası (örnek AMNH 5214) 55,5 santimetre (1,82 ft) uzunluğunda ve 64,5 santimetre (2,12 ft) genişliğindedir ve Carpenter, yaklaşık 5,4 metre (18 ft) uzunluğunda ve yaklaşık 1,4 metre (4,6 ft) uzunluğunda olduğunu tahmin etmektedir. kalçalarda.[10] Amerikalı paleontolog Roger B.J. Benson ve arkadaşları, 2014 yılında AMNH 5214'ün ağırlığını 4,78 metrik ton (5,27 kısa ton) olarak tahmin etti.[14]
Arbor ve Mallon, 2017'de daha eksiksiz ankilozorinlerle yapılan karşılaştırmalara dayanarak, CMN 8880 için 7,56 ila 9,99 metre (24,8 ila 32,8 ft) ve AMNH 5214 için 6,02 ila 7,95 metre (19,8 ila 26,1 ft) bir uzunluk tahmin etti. en küçük örneğidir Ankylosauruskafatası hala diğer ankilozorinlerden daha büyüktür. Diğer birkaç ankylosaur'un uzunluğu yaklaşık 6 metreye (20 ft) ulaştı. AMNH 5214'ün omurları diğer ankilozorinlerden önemli ölçüde daha büyük olmadığından, Arbor ve Mallon, büyükler için yaklaşık 10 metrelik (33 ft) üst aralık tahminlerini değerlendirdiler. Ankylosaurus çok uzun ve bunun yerine 8 metre (26 ft) uzunluk önerdi. Arbor ve Mallon, AMNH 5214 için 4,78 metrik ton (5,27 kısa ton) bir ağırlık tahmin etti ve CMN 8880'in ağırlığını geçici olarak 7.95 metrik ton (8.76 kısa ton) olarak tahmin etti.[12]
Kafatası

Bilinen üç Ankylosaurus kafatasları çeşitli ayrıntılarda farklılık gösterir; bunun sonucu olduğu düşünülüyor tafonomi (kalıntıların çürümesi ve fosilleşmesi sırasında meydana gelen değişiklikler) ve bireysel varyasyon. Kafatası alçak ve üçgen şeklindeydi ve uzun olduğundan daha genişti; kafatasının arkası geniş ve alçaktı. Kafatasının geniş bir gaga üzerinde premaksilla. yörüngeler (göz yuvaları) neredeyse yuvarlak ila hafif ovaldi ve kafatası öne doğru sivrildiği için doğrudan yana bakmıyordu. Beyin tabanı, diğer ankilozorinlerde olduğu gibi kısa ve sağlamdı. Yörüngelerin üzerindeki tepeler yukarı doğru birleşti skuamozal boynuzlar (şekilleri "piramidal "), kafatasının arkasından yanları geriye doğru işaret ediyor. İlgili resimde görüldüğü gibi, tepe ve boynuz muhtemelen orijinal olarak ayrı öğelerdi. Pinacosaurus ve Euoplocephalus. Üst boynuzların altında cüce Geriye ve aşağıya dönük boynuzlar mevcuttu. Boynuzlar başlangıçta kafatasına kaynaşmış osteodermler olabilir. ölçek Ankylosaurların kafataslarının yüzeylerindeki benzeri kafatası süslemelerine "Caputegulae"ve sonucuydu yeniden modelleme kafatasının kendisi. Bu yok etti dikişler yetişkin ankilozorlar için yaygın olan kafatası unsurları arasında. Kafatasının kaputegulum modeli, bazı detaylar paylaşılsa da örnekler arasında değişkendir. Caputegulalar, kafatasındaki konumlarına ve Ankylosaurus burun delikleri arasındaki burnun önünde nispeten büyük, altıgen (veya elmas şeklinde) bir burun kaputegülü içerir; bu, her iki tarafında bir loreal kaputegulum, her bir yörüngenin üzerinde bir ön ve arka supraorbital kaputegulum ve ense kaputegulanın bir çıkıntısına sahiptir. kafatasının arkası.[12][10][15]

Burun bölgesi Ankylosaurus ankilozorlar arasında benzersizdi ve akrabalarına kıyasla "aşırı" bir dönüşüm geçirmişti. Burun önden kavisli ve kesikti ve burun delikleri eliptikti ve eğik bir şekilde öne veya yukarı baktıkları bilinen diğer tüm ankilozoridlerden farklı olarak aşağı ve dışa doğru yönlendirilmişlerdi. Ek olarak, burun delikleri önden görünmüyordu çünkü sinüsler premaksilla kemiklerinin kenarlarına, diğer ankilozorlarda görülenden daha büyük ölçüde genişledi. Geniş loreal caputegulalar - burnun kayış benzeri, yan osteodermleri - burun deliklerinin genişlemiş açıklığını tamamen kaplayarak soğanlı bir görünüm verdi. Burun deliklerinde ayrıca intranaryal septum Burun geçişini sinüsten ayıran. Burnun her iki tarafında dördü genişleyen beş sinüs vardı. üst çene kemik. Burun boşlukları (veya odaları) Ankylosaurus uzatılmış ve orta hatta bir septum ile ayrılmış, burnun içini iki aynalı yarıya böler. Burun odalarının iki açıklığı vardı. Choanae (iç burun delikleri) ve hava geçişi ilmeklendi.[10][12] Üst çene yanlara doğru genişledi ve içerideki sinüslerden kaynaklanmış olabilecek bir çıkıntı izlenimi verdi. Çene kemiğinin etli yanakların bağlanma yeri olabilecek bir çıkıntısı vardı; Ornitoschianlarda yanakların varlığı tartışmalıdır, ancak bazı nodozorların ete gömülmüş olabilecek yanak bölgesini kaplayan zırh plakaları vardı.[10]
AMNH 5214 numunesinde 34–35 diş alveolleri maksillada (diş yuvaları). Bu numunenin maksillasındaki diş sıraları yaklaşık 20 santimetre (7,9 inç) uzunluğundadır. Her alveolde bir foramen (açılış) yanına yakın yedek diş görülebilir. Diğer ankylosaurlarla karşılaştırıldığında, çene nın-nin Ankylosaurus uzunluğu ile orantılı olarak düşüktü ve yandan bakıldığında diş sırası kavisli yerine neredeyse düzdü. Çeneler yalnızca en küçük örnekte (AMNH 5214) tamamen korunmuştur ve yaklaşık 41 santimetre (1.35 ft) uzunluğundadır. En büyük numunenin (CMN 8880) eksik mandibulası aynı uzunluktadır. AMNH 5214'ün solunda 35 diş alveolü vardır diş hekimi ve sağda 36, toplamda 71. öncül çene ucunun kemiği henüz bulunamadı.[10] Diğer ankylosaurlar gibi, Ankylosaurus yanlara doğru sıkıştırılmış küçük, filliform (yaprak şekilli) dişlere sahipti.[13] Dişler çoğunlukla geniş olduklarından daha uzundu ve çok küçüktü; kafatasına orantılı büyüklükleri, çenelerinin Ankylosaurus diğer ankylosaurinlerden daha fazla diş barındırabilir. En büyüğünün dişleri Ankylosaurus kafatası, mutlak anlamda en küçük kafatasından daha küçüktür. Diş sırasındaki bazı dişler arkaya doğru kıvrılır ve diş kronları genellikle bir tarafı diğerinden daha yassıdır.[10] Ankylosaurus dişler tanı ve düz kenarlarına göre diğer ankilozoridlerin dişlerinden ayırt edilebilir. dişler büyüktü, sayıları dişin ön kısmında altı ile sekiz arasında ve arkada beş ile yedi arasında değişiyordu.[10][16]
Postkraniyal iskelet

İskeletinin çoğunun yapısı Ankylosaurusçoğu dahil leğen kemiği, kuyruk ve ayaklar hala bilinmiyor.[10] Öyleydi dört ayaklı ve arka bacakları ön ayaklarından daha uzundu.[17] Holotip örneğinde, kürek kemiği (kürek kılıcı) 61.5 santimetre (2.02 ft) uzunluğundadır ve korakoid (kürek kemiğinin alt ucuna bağlı dikdörtgen bir kemik). Ayrıca vardı entezler (bağ dokusu) çeşitli kas ekleri için. humerus AMNH 5214'ün (üst kol kemiği) kısa, çok geniş ve yaklaşık 54 santimetre (1.77 ft) uzunluğundaydı. uyluk (uyluk kemiği), yine AMNH 5214'ten, 67 santimetre (2.20 ft) uzunluğundaydı ve çok sağlamdı. Ayakları Ankylosaurus tam olarak bilinmemekle birlikte, ileri ankilozoridlerde olduğu gibi arka ayakların muhtemelen üç parmağı vardı.[10]
boyun omurları vücuda doğru yüksekliği artan geniş sinir dikenleri vardı. Sinir dikenlerinin ön kısmı, yetişkin dinozorlar arasında yaygın olan iyi gelişmiş entezlere sahipti ve büyük bağlar, bu büyük kafayı desteklemeye yardımcı oldu. sırt omur genişliklerine göre kısa merkez (veya bedenler) vardı ve sinir dikenleri kısa ve dardı. Sırt omurları, sırtın aşağı doğru hareketini sınırlayan sıkıca aralıklıydı. Sinir dikenleri vardı kemikleşmiş (kemiğe döndü) tendonlar bazı omurlarla da örtüşüyordu. Son dört arka omurun kaburgaları, diyapofizler ve parapofizler (kaburgaları omurlarla eklemleyen yapılar) ve göğüs kafesi vücudun bu bölümünde çok genişti. Kaudal omurlar, her iki tarafta da içbükey oldukları anlamına gelen hafif amfikol olan merkezlere sahipti.[10]
Zırh

Önemli bir özelliği Ankylosaurus deriye gömülü osteodermler veya yivler olarak bilinen topuzlardan ve kemik plakalarından oluşan zırhıydı. Bunlar eklemlemede bulunamadı, bu nedenle vücuttaki kesin yerleşimleri bilinmemekle birlikte, ilgili hayvanlara dayanarak çıkarımlar yapılabilir ve çeşitli konfigürasyonlar önerilmiştir. Osteodermlerin çapı 1 santimetre (0.39 inç) ile 35.5 santimetre (1.16 ft) arasında değişiyordu ve şekli değişiyordu. Osteodermler Ankylosaurus genellikle ince cidarlı ve alt tarafı oyuktur. Nazaran Euoplocephalusosteodermler Ankylosaurus daha pürüzsüzdü. Diğer ankylosauridlerde olduğu gibi birçok küçük osteoderm ve kemikçik muhtemelen büyük olanlar arasındaki boşluğu işgal etti. Vücudu kaplayan osteodermler, bir kenarda alçak bir omurga ile çok düzdü. Buna karşılık, nodosaurid Edmontoni osteodermlerinin orta hattında bir kenardan diğerine uzanan yüksek omurgalara sahipti. Ankylosaurus Orta hatta bir omurga ile bazı küçük osteodermler vardı.[12][10]

Diğer ankylosaurids gibi, Ankylosaurus vardı servikal yarım halkalar (boynundaki zırh plakaları), ancak bunlar yalnızca parçalardan biliniyor, bu da onların tam yerleşimlerini belirsiz kılıyor. Carpenter, yukarıdan bakıldığında plakaların eşleştirilmiş olacağını, boyun boyunca ters bir V şekli oluşturacağını ve orta hat boşluğunun muhtemelen harekete izin vermek için küçük kemikçiklerle (yuvarlak kemikli yarıklar) doldurulacağını öne sürdü. Bu zırh kemerinin genişliğinin sadece boyuna sığamayacak kadar geniş olduğuna ve boyun tabanını kaplayıp omuz bölgesine kadar devam ettiğine inanıyordu. Arbor ve Kanadalı paleontolog Philip J. Currie Carpenter'ın 2015'teki yorumuna katılmadı ve holotip numunesinin servikal yarım halka parçalarının Carpenter tarafından önerilen şekilde birbirine uymadığına işaret etti (bunun nedeni kırılma olabilir). Bunun yerine, parçaların, yakından ilişkili olduğu gibi, boynun üst kısmı etrafında iki yarı dairesel zırh plakası oluşturan iki servikal yarım halkanın kalıntılarını temsil ettiğini öne sürdüler. Anodontozorlar ve Euoplocephalus.[10][15] Arbor ve Mallon, bu yarım halkaların şeklini boynun üst kısmındaki "sürekli U-şekilli boyunduruklar" olarak tanımlayarak bu fikri detaylandırdılar ve şunu önerdiler: Ankylosaurus her yarım halkada oval tabanlı altı omurgalı osteoderm vardı.[12]
İkinci servikal yarım halkanın arkasındaki ilk osteodermlerin şekli ilk yarım halkadakilere benzer olacaktı ve arkadaki osteodermlerin çapı muhtemelen arkaya doğru azaldı. En büyük osteodermler, derideki kırışıklıklar ile ayrılmış dört veya beş enine sıra ile muhtemelen vücudun çoğu boyunca enine ve boylamasına sıralar halinde düzenlenmiştir. Kanatlardaki osteodermler muhtemelen arkadakilerden daha kare bir dış çizgiye sahip olacaktı. Yanlarda dört uzunlamasına osteoderm sırası olabilir. Bazı bazal ankilozorların ve birçok nodozorun aksine, ankilozoridlerin kalçalarının üzerinde birlikte kemikleşmiş pelvik kalkanlara sahip oldukları görülmemektedir. Omurgasız bazı osteodermler, kalça bölgesinin üzerine yerleştirilmiş olabilir. Ankylosaurus, de olduğu gibi Euoplocephalus. Ankylosaurus pelvik bölge üzerinde, vücudun geri kalanındakilerden daha küçük olan üç veya dört enine dairesel osteoderm sırasına sahip olabilir. Scolosaurus. Pelvisin yanlarında daha küçük, üçgen osteodermler mevcut olabilir. Düzleştirilmiş, sivri uçlu plakalar, kuyruğun yan tarafındakilere benzer. Saichaniave benzer şekilde dağıtılmış olabilir Ankylosaurus. Oval omurgalı osteodermler kuyruğun üst tarafına veya uzuvların yan tarafına yerleştirilmiş olabilir. Sıkıştırılmış, üçgen osteodermler ile bulundu Ankylosaurus örnekler pelvisin veya kuyruğun yanlarına yerleştirilmiş olabilir. Ovoid, omurgalı ve gözyaşı damlası şeklindeki osteodermler, Ankylosaurusve ön ayaklara yerleştirilmiş olabilir, örneğin Pinacosaurus, ancak arka bacaklarda osteodermlerin olup olmadığı bilinmemektedir.[10][12]

Kuyruk kulübü (veya kuyruk topuzu) Ankylosaurus orta hatta bir sıra küçük osteoderm ve ucunda iki küçük osteoderm olmak üzere iki büyük osteodermden oluşuyordu; bu osteodermler son kuyruk omurunu gizledi. Sadece AMNH 5214 örneğinin kuyruk kulübü bilindiğinden, bireyler arasındaki varyasyon aralığı bilinmemektedir. AMNH 5214'ün kuyruk kulübü 60 santimetre (2.0 ft) uzunluğunda, 49 santimetre (1.61 ft) genişliğinde ve 19 santimetre (7.5 inç) yüksekliğindedir. En büyük numunenin kulübü 57 santimetre (1.87 ft) genişliğinde olabilir. Kuyruk kulübü Ankylosaurus yukarıdan bakıldığında yarım daire şeklindeydi, yukarıdan bakıldığında Euoplocephalus ve Scolosaurus ama sivri kulüp osteodermlerinin aksine Anodontozorlar ya da dar, uzun kulübü Dyoplosaurus. Son yedi kuyruk omuru, kuyruk kulübünün "sapını" oluşturdu. Bu omurlar temas halindeydi, aralarında kıkırdak yoktu ve bazen birlikte kemikleşmişlerdi, bu da onları hareketsiz kılıyordu. Kuyruk klübünün önündeki omurlara yapışan kemikleşmiş tendonlar ve bu özellikler birlikte onu güçlendirmeye yardımcı oldu. Kilitli zigapofizler (eklem süreçleri) ve sap omurlarının sinir dikenleri, yukarıdan bakıldığında U şeklindeyken, diğer çoğu ankilozoridinkiler V şeklindedir, bu da Ankylosaurus daha geniş olmak. Daha büyük genişlik, kuyruğun Ankylosaurus vücut uzunluğuna göre diğer ankilozoridlerden daha kısaydı veya aynı oranlara sahipti, ancak daha küçük bir sopayla.[12][10][18]
Sınıflandırma
Brown düşünüldü Ankylosaurus o kadar belirgin ki bunu o yaptı tip cins yeni aile, Ankylosauridae, masif, üçgen kafatasları, kısa boyunlar, sert sırtlar, geniş gövdeler ve osteodermlerle temsil edilir. Ayrıca sınıflandırdı Palaeoscincus (sadece dişlerden bilinir) ve Euoplocephalus (daha sonra sadece kısmi bir kafatası ve osteodermlerden bilinir) ailenin bir parçası olarak. Kalıntıların parçalı durumundan dolayı Brown, birbirlerinden tam olarak ayırt edemedi. Euoplocephalus ve Ankylosaurus. Ailenin sadece birkaç eksik üyesiyle karşılaştırma yapmak için grubun toplumun bir parçası olduğuna inanıyordu. alttakım Stegosauria.[2] 1923'te Osborn adını icat etti Ankilosauri, böylece ankilozoridleri kendi alt sıralarına yerleştirir.[19]


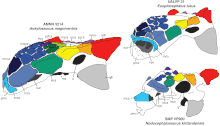
Ankylosauria ve Stegosauria artık sınıf içinde birlikte gruplandırılmıştır. Thyreophora. Bu grup ilk olarak Sinemurian yaş ve 135 milyon yıl hayatta kaldı. Maastrihtiyen. Yaygındı ve çok çeşitli ortamlarda yaşadılar.[10][6] Daha eksiksiz örnekler ve yeni cinsler keşfedildikçe, ankilosauri'nin birbiriyle ilişkili olduğuna dair teoriler daha karmaşık hale geldi ve hipotezler çalışmalar arasında sıklıkla değişti. Ankylosauridae'ye ek olarak, Ankylosauria, Nodosauridae ailelerine bölünmüştür ve bazen Polacanthidae (bu ailelerin kuyruk kulüpleri yoktu).[20] Ankylosaurus parçası olarak kabul edilir alt aile Ankylosaurinae (üyelerine ankilozorinler denir) Ankylosauridae içinde.[20] Ankylosaurus en yakından ilgili gibi görünüyor Anodontozorlar ve Euoplocephalus.[21] Aşağıdaki kladogram 2015'e dayanıyor Filogenetik analiz Arbor ve Currie tarafından yönetilen Ankylosaurinae:[15]
| Ankylosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Dan beri Ankylosaurus ve Asya cinsleri ile gruplanmış diğer Geç Kretase Kuzey Amerika ankilozoridleri (bir kabile yazarların adı Ankylosaurini ), Arbor ve Currie daha önceki Kuzey Amerika ankilozoridlerinin son zamanlarda neslinin tükendiğini öne sürdü. Albiyen veya Senomaniyen Orta Çağlar Kretase. Ankylosaurids daha sonra Kuzey Amerika'yı Asya'dan yeniden kolonileştirdi. Kampaniyen veya Turoniyen Geç Kretase çağları ve orada çeşitlendirilmiş yine, gibi cinslere yol açar Ankylosaurus, Anodontozorlar, ve Euoplocephalus. Bu, 30 milyon yıllık bir boşluğu açıklıyor. fosil bu çağlar arasında Kuzey Amerika ankilozoridlerinin kaydı.[15]
Paleobiyoloji
Besleme

Diğerleri gibi ornithischians, Ankylosaurus oldu otçul. Geniş ağzı, seçici olmayan düşükAraştır kırpma,[10] bazı ilgili cinslerde görüldüğü ölçüde olmasa da, özellikle Euoplocephalus.[12][22] Ankylosaurlar beslenmemiş olabilir lifli ve odunsu bitkiler sert yapraklar ve etli meyveler dahil olmak üzere çeşitli diyetleri olmuş olabilir.[23] Ankylosaurus Muhtemelen bolca beslendi eğrelti otları ve düşük büyüyen çalılar. Varsayalım ki endotermik, Ankylosaurus kuru bitki örtüsü miktarına benzer şekilde, günde 60 kilogram (130 lb) eğrelti otu tüketirdi. fil tüketirdi. Beslenme gereksinimleri daha etkili bir şekilde karşılanabilirdi eğer Ankylosaurus küçük, sivri uçlu dişleri ve gagasının şekli, örneğin Euoplocephalus. Küçük dişlerin kullanım için uyarlanmış olabileceği bazı omurgasızlar da ek besin sağlayabilirdi.[12]
Fosilleri Ankylosaurus dişler, nodosaurid ankylosaurlarda olduğu gibi, tacın ucundan ziyade taç yüzünde aşınma gösterir.[10] 1982'de Carpenter bebeğe atfedildi Ankylosaurus Lance ve Hell Creek Formasyonlarından kaynaklanan ve sırasıyla 3.2 ila 3.3 milimetre (0.13 ila 0.13 inç) uzunluğunda olan iki çok küçük diş. Daha küçük olan diş çok aşınmış, bu da Carpenter'ın genel olarak ankilozoridlerin ya da en azından gençlerin yiyeceklerini tamamen yutmadığını, ancak bir çeşit çiğneme kullandığını öne sürmesine neden oldu.[16] Yetişkinlerden beri Ankylosaurus yiyeceklerini çok az çiğnemiş olsaydı, gün içinde yiyecek toplamak için bir filden daha az zaman harcardı.[12] Göğüs kafesinin genişliğine bağlı olarak, çiğnenmemiş yiyeceklerin sindirimi şu şekilde kolaylaştırılmış olabilir: arka bağırsak fermantasyonu Genişlemiş hallerinde birkaç odacığa sahip modern otçul kertenkelelerde olduğu gibi kolon.[10]

1969'da Avusturyalı paleontolog Georg Haas Ankylosaur kafataslarının büyüklüğüne rağmen, ilişkili kas yapısının nispeten zayıf olduğu sonucuna vardı. Ayrıca çene hareketinin yukarı ve aşağı hareketlerle sınırlı olduğunu düşünüyordu. Haas, bundan çıkarım yaparak, ankilozorların nispeten yumuşak, aşındırıcı olmayan bitki örtüsünü yediğini öne sürdü.[24] Daha sonra araştırma Euoplocephalus bu hayvanlarda ileri ve yan çene hareketinin mümkün olduğunu, kafatasının önemli kuvvetlere dayanabildiğini belirtir.[25] 2016 yılı çalışması diş tıkanıklığı Ankylosaur örneklerinin (dişler arasındaki temas) geriye doğru (palinal) çene hareketi kabiliyetinin olduğunu buldu. bağımsız olarak gelişti farklı ankylosaur'da soylar Geç Kretase Kuzey Amerika ankilozoridleri gibi Ankylosaurus ve Euoplocephalus.[22]
Ankylosaur'un bir örneği Pinacosaurus kas gerginliği belirtileri gösteren büyük paraglossalia'yı (dilde bulunan üçgen kemikler veya kıkırdaklar) korur ve bunun ankilozorların ortak bir özelliği olduğu düşünülmektedir. Örneği inceleyen araştırmacılar, ankylosaurların, dişleri oldukça küçük olduğu ve nispeten yavaş bir oranda değiştirildiği için beslenirken kas dillerine ve hyobranchia'ya (dil kemikleri) büyük ölçüde güvendiklerini öne sürdüler. Biraz modern semenderler benzer dil kemiklerine sahip olmak ve kavrayıcı yiyecek almak için diller.[23] Burun deliklerinin geri çekilmiş konumu Ankylosaurus ile karşılaştırıldı fosforlu (kazma) solucan kertenkeleleri ve kör yılanlar ve muhtemelen yuva yapan bir hayvan olmasa da, burnu Ankylosaurus toprak hareketi davranışını gösterebilir. Bu faktörler ve diğer ornitoschianlara göre ankilozorlarda düşük diş oluşumu oranı, Ankylosaurus olabilir her yerde yaşayan (hem bitki hem de hayvan maddesini yemek). Ayrıca (veya alternatif olarak) kökler için toprağa kazılmış olabilir ve yumrular.[12]
Hava sahaları ve duyular
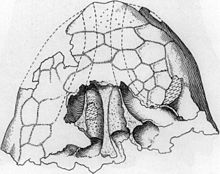
1977'de Polonyalı paleontolog Teresa Maryańska Ankilozorların karmaşık sinüslerinin ve burun boşluklarının kafatasının ağırlığını hafifletmiş olabileceğini, bez veya için bir oda olarak hareket etti ses rezonansı.[10][26] Carpenter, bu hipotezleri reddederek, dört ayaklı hayvanlar sesler çıkarır gırtlak, burun delikleri değil ve boşluklar kafatası hacminin yalnızca küçük bir yüzdesini oluşturduğu için ağırlıktaki azalma minimum düzeydeydi. Ayrıca bir bezi olası olmadığını düşündü ve sinüslerin belirli bir işlevi olmayabileceğini belirtti.[10] Solunum yollarının, solunum yollarının varlığına ve düzenlenmesine bağlı olarak solunan havanın memeli benzeri bir tedavisini gerçekleştirmek için kullanıldığı da öne sürülmüştür. özel kemikler.[26]
Burun pasajlarının 2011 yılı çalışması Euoplocephalus Japon paleontolog Tetsuto Miyashita ve meslektaşları tarafından, bir ısı ve su dengeleme sistemi olarak işlevlerini destekleyerek, geniş kan damarı sistemi ve artan mukoza zarı (modern hayvanlarda ısı ve su değişimi için kullanılır). Araştırmacılar ayrıca, ilmeklerin bir rezonans odası olarak işlev görmesi fikrini de desteklediler. saiga antilop ve döngüsel soluk borusu vinçler ve kuğu. İç kulağın rekonstrüksiyonu, muhtemelen nazal pasajlar tarafından üretilen düşük tonlu rezonant sesler gibi düşük frekanslarda işitmeye adaptasyonu önerir. Döngünün aşağıdakilerle ilgili olma olasılığını tartıştılar koku alma (koku duyusu) koku alma bölgesi ana hava yolunun yanlarına doğru itilir.[27]
Carpenter'a göre burun odalarının şekli Ankylosaurus hava akışının tek yönlü olduğunu (inhalasyon ve ekshalasyon sırasında akciğerler boyunca döngü), ancak arka burun bölmesinde iki yönlü de olabilir, hava koku alma lobları.[10] Ankylosauridlerin genişlemiş koku alma bölgesi, iyi gelişmiş bir koku alma duyusuna işaret eder.[27] Burun deliklerinin geriye doğru çekilmesi, suda yaşayan hayvanlarda ve hortum her iki olasılık da geçerli değildir Ankylosaurusburun delikleri küçülme veya premaksilla uzama eğiliminde olduğundan. Buna ek olarak, geniş bir şekilde ayrılmış burun delikleri, (her bir burun deliğinin farklı yönlerden kokduğu) stereo-olfaksiyona izin vermiş olabilir. geyik, bu özellik hakkında çok az şey bilinmektedir.[12] Yörüngelerinin konumu Ankylosaurus biraz öner stereoskopik görüş.[10]
Uzuv hareketleri

1978'de Coombs tarafından yapılan ankilozor ön ayakları kas yapısının rekonstrüksiyonları, ön ayakların hayvanın ağırlığının çoğunu taşıdığını ve muhtemelen yiyecek toplama için ön ayaklarda yüksek kuvvet dağıtımı için uyarlandığını göstermektedir. Ek olarak, Coombs, ankilozorların toynak benzeri yapısı olmasına rağmen, ankilozorların kazma kabiliyetine sahip olabileceğini öne sürdü. Manus sınırlı fosforsal aktiviteye sahip olacaktı. Ankylosaurlar muhtemelen yavaş hareket eden ve halsiz hayvanlar olmuştu.[28][29] Gerektiğinde hızlı hareket edebilecek olsalar da.[17]
Büyüme
En büyüğünün skuamozal boynuzları Ankylosaurus Numune, en küçük numuneden daha kördür, bu da aynı şekilde Euoplocephalusve bu temsil edebilir ontogenetik varyasyon (büyüme gelişimi ile ilgili).[12] Örneklerinin çalışmaları Pinacosaurus Farklı yaşlardan itibaren, ontogenetik gelişim sırasında, juvenil ankilozorların kaburgalarının omurlarıyla kaynaştığını bulmuşlardır. Ön ayakların sağlamlığı güçlü bir şekilde artarken, arka ayaklar iskeletin geri kalanına göre daha büyük hale gelmedi, bu da kolların ağırlığın çoğunu taşıdığına dair bir kanıt. Servikal yarım halkalarda, altta yatan kemik bandı, onu aynı anda birbirine kaynaşan altta yatan osteodermlere bağlayan büyümeler geliştirdi.[30] Kafatasında, orta kemik plakaları önce burun ve arka kenarda kemikleşmiş, kemikleşme yavaş yavaş orta bölgelere doğru uzanıyordu. Vücudun geri kalanında kemikleşme boyundan geriye kuyruk yönünde ilerledi.[31]
Savunma

Ankylosauridlerin osteodermleri, diğer ankylosaurlara kıyasla inceydi ve rastgele dağıtılmış yastıklarla güçlendirilmiş gibi görünüyordu. kolajen lifler. Yapısal olarak benzer Sharpey lifleri ankilozoridlere özgü bir özellik olan doğrudan kemik dokusuna gömülmüşlerdi. Bu, ankylosauridlere hem hafif hem de oldukça dayanıklı, yırtıcı hayvanların dişlerinin kırılmasına ve delinmesine karşı dirençli bir zırh kaplaması sağlayacaktır.[32] palpebral kemikler gözlerin üzerinde onlar için ek koruma sağlamış olabilir.[33] Carpenter 1982'de ağır damarlanmış zırhın da bir rolü olabileceğini öne sürdü. termoregülasyon modernde olduğu gibi timsahlar.[34]

Kuyruk kulübü Ankylosaurus bir saldırganın kemiklerini kırmaya yetecek kadar darbe üretebilen aktif bir savunma silahı gibi görünüyor. Kuyruğun tendonları kısmen kemikliydi ve çok elastik değildi, bu da bir silah olarak kullanıldığında kulübe büyük bir kuvvet iletilmesine izin veriyordu.[10] Coombs, 1979'da birkaç arka ekstremite kasının kuyruğun sallanmasını kontrol edeceğini ve kulübün şiddetli itişmelerinin, metatarsal kemikler büyük theropodlar.[29] 2009 yılında yapılan bir araştırma, ankilozoridlerin kuyruklarını yanal olarak 100 derece sallayabildiğini tahmin ediyordu. iptal clubs would have had a lowered eylemsizlik momenti and been effective weapons. The study also found that while adult ankylosaurid tail clubs were capable of breaking bones, those of juveniles were not. Despite the feasibility of tail-swinging, the researchers could not determine whether ankylosaurids used their clubs for defense against potential predators, in tür içi mücadele, ya da her ikisi de.[35]
In 1993 Tony Thulborn proposed that the tail club of ankylosaurids primarily acted as a decoy for the head, as he thought the tail too short and inflexible to have an effective reach; the "dummy head" would lure a predator close to the tail, where it could be struck.[36] Carpenter has rejected this idea, as tail club shape is highly variable among ankylosaurids, even in the same genus.[10]
Paleoçevre

Ankylosaurus existed between 68 and 66 million years ago, in the final, or Maastrichtian, stage of the Geç Kretase Dönem. It was among the last dinosaur genera that appeared before the Kretase-Paleojen nesli tükenme olayı. The type specimen is from the Hell Creek Formation of Montana, while other specimens have been found in the Lance and Ferris Formations in Wyoming, the Scollard Formation in Alberta, and the Fransız Formasyonu in Saskatchewan, all of which date to the end of the Cretaceous.[37][38][12]
Fosilleri Ankylosaurus are rare in these sedimanlar, and the distribution of its remains suggests that it was ecologically rare, or restricted to the uplands of the formations rather than the coastal lowlands, where it would have been more likely to fossilize. Another ankylosaur, a nodosaur referred to as Edmontoni sp., is also found in the same formations, but according to Carpenter, the range of the two genera does not seem to have overlapped. Their remains have so far not been found in the same localities, and the nodosaur appears to have inhabited the lowlands. The narrower muzzle of the nodosaur suggests it had a more selective diet than Ankylosaurus, further indicating ecological separation, whether their range overlapped or not.[12][10]
With its low center of gravity, Ankylosaurus would have been unable to knock down trees like modern elephants do. It was also incapable of chewing bark and thus unlikely to have practiced bark stripping. Bir yetişkin olarak, Ankylosaurus does not appear to have congregated in groups (though some ankylosaurs appear to have congregated when young). So, although it was a large herbivore with similar energetic requirements, it is therefore improbable that Ankylosaurus was able to modify the landscape of its ecosystem in the way elephants do; hadrosauridler may instead have had such an "ekosistem mühendisi "rol.[12]
The formations where Ankylosaurus fossils have been found represent different sections of the western shore of the Batı İç Denizyolu dividing western and eastern North America during the Cretaceous, a broad coastal plain extending westward from the seaway to the newly formed kayalık Dağlar. These formations are composed largely of kumtaşı ve çamurtaşı, which have been attributed to taşkın yatağı ortamlar.[39][40][41] The regions where Ankylosaurus and other Late Cretaceous ankylosaurs have been found had a warm subtropikal /ılıman iklim, hangisiydi muson, had occasional rainfall, tropikal fırtınalar, ve Orman yangınları.[22] In the Hell Creek Formation, many types of plants were supported, primarily anjiyospermler, with less common iğne yapraklılar, eğrelti otları ve sikadlar. An abundance of fossil leaves found at dozens of different sites indicates that the area was largely forested by small trees.[42] Ankylosaurus shared its environment with other dinosaurs that included the ceratopsids Triceratops ve Torosaurus, Hipsilofodont Thescelosaurus, the hadrosaurid Edmontosaurus, an indeterminate nodosaur, the pachycephalosaurian Pachycephalosaurus, and the theropods Struthiomimus, Ornitomimus, Pectinodon, ve Tyrannosaurus.[38][43]
Ayrıca bakınız
Referanslar
Notlar
- ^ Telaffuz edildi /ˌæŋkəloʊˈsɔːrəs/ ANG-kə-lo-SAWR-əs[1]
Alıntılar
- ^ "Ankylosaurus". Merriam-Webster Sözlüğü.
- ^ a b c Brown, B. (1908). "The Ankylosauridae, a new family of armored dinosaurs from the Upper Cretaceous". Amerikan Doğa Tarihi Müzesi Bülteni. 24: 187–201. hdl:2246/1435.
- ^ Creisler, B. (July 7, 2003). "Dinosauria Translation and Pronunciation Guide A". Arşivlenen orijinal 18 Ağustos 2010. Alındı 3 Eylül 2010.
- ^ Liddell, H. G.; Scott, R. (1980) [1871]. Yunanca-İngilizce Sözlük (kısaltılmış ed.). Oxford University Press. s.5. ISBN 978-0-19-910207-5.
- ^ a b Glut, D.F. (1997). "Ankylosaurus". Dinozorlar, ansiklopedi. McFarland & Company, Inc. Yayıncılar. pp.141–143. ISBN 978-0-375-82419-7.
- ^ a b Coombs, W. (1978). "The families of the ornithischian dinosaur order Ankylosauria" (PDF). Paleontoloji Dergisi. 21 (1): 143–170. Arşivlenen orijinal (PDF) 8 Temmuz 2015. Alındı 8 Temmuz 2015.
- ^ Naish, D. (2009). The Great Dinosaur Discoveries. London: A & C Black Publishers LTD. s. 58–59. ISBN 978-1408119068.
- ^ Williston, S. W. (1908). "Review: The Ankylosauridae". Amerikan Doğa Uzmanı. 42 (501): 629–30. doi:10.1086/278987. JSTOR 2455817.
- ^ Carpenter, K. (2001). "Chapter 21: Phylogenetic Analysis of the Ankylosauria". In Carpenter, K. (ed.). Zırhlı Dinozorlar. pp. 454–83. ISBN 978-0-253-33964-5.
- ^ a b c d e f g h ben j k l m n Ö p q r s t sen v w x y z aa ab AC Carpenter, K. (2004). "Yeniden Tanımlama Ankylosaurus magniventris Brown 1908 (Ankylosauridae) from the Upper Cretaceous of the Western Interior of North America". Kanada Yer Bilimleri Dergisi. 41 (8): 961–86. Bibcode:2004CaJES..41..961C. doi:10.1139/e04-043.
- ^ Osborn, H. F. (1905). "Tyrannosaurus and other Cretaceous carnivorous dinosaurs". AMNH Bülteni. 21 (14): 259–265. hdl:2246/1464.
- ^ a b c d e f g h ben j k l m n Ö p q r s t Arbour, V.M.; Mallon, J.C. (2017). "Unusual cranial and postcranial anatomy in the archetypal ankylosaur Ankylosaurus magniventris". FACETS. 2 (2): 764–794. doi:10.1139/facets-2017-0063.
- ^ a b Coombs, W. (1990). "Teeth and taxonomy in ankylosaurs". Carpenter, K .; Currie, P. J. (eds.). Dinosaur systematics: Approaches and perspectives. Cambridge University Press. pp. 269–79. ISBN 978-0-521-43810-0.
- ^ Benson, R. B. J .; Campione, N. E .; Carrano, M. T .; Mannion, P. D.; Sullivan, C .; et al. (2014). "Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage". PLOS Biol. 12 (5): e1001853. doi:10.1371 / journal.pbio.1001853. PMC 4011683. PMID 24802911.
- ^ a b c d Arbor, V. M .; Currie, P.J. (2015). "Ankylosaurid dinozorların sistematiği, filogenisi ve paleobiyocoğrafyası". Sistematik Paleontoloji Dergisi. 14 (5): 1–60. doi:10.1080/14772019.2015.1059985. S2CID 214625754.
- ^ a b Carpenter, K. (1982). "Baby dinosaurs from the Late Cretaceous Lance and Hell Creek formations and a description of a new species of theropod". Rocky Dağı Jeolojisi. 20 (2): 123–134.
- ^ a b Coombs, W. P. (1978). "Dinozorlardaki cursorial adaptasyonların teorik yönleri". Biyolojinin Üç Aylık İncelemesi. 53 (4): 393–418. doi:10.1086/410790. S2CID 84505681.
- ^ Arbor, V. M .; Currie, P.J. (2015). "Ankylosaurid dinosaur tail clubs evolved through stepwise acquisition of key features". Anatomi Dergisi. 227 (4): 514–23. doi:10.1111/joa.12363. PMC 4580109. PMID 26332595.
- ^ Osborn, H.F. (1923). "Two Lower Cretaceous dinosaurs of Mongolia". Amerikan Müzesi Novitates. 95: 1–10. hdl:2246/3267.
- ^ a b Thompson, R. S.; Parish, J. C .; Maidment, S.C. R .; Barrett, P. M. (2012). "Ankylosaurian dinozorlarının filogenisi (Ornithischia: Thyreophora)". Sistematik Paleontoloji Dergisi. 10 (2): 301–312. doi:10.1080/14772019.2011.569091. S2CID 86002282.
- ^ Arbour, V.M.; Currie, P.J.; Badamgarav, D. (2014). "The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia". Linnean Society'nin Zooloji Dergisi. 172 (3): 631–652. doi:10.1111/zoj.12185.
- ^ a b c Ősi, A .; Prondvai, E .; Mallon, J.; Bodor, E. R. (2016). "Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornithischia)". Tarihsel Biyoloji. 29 (4): 539–570. doi:10.1080/08912963.2016.1208194. S2CID 55372674.
- ^ a b Hill, R. V.; D'Emic, M. D.; Bever, G. S .; Norell, M. A. (2015). "A complex hyobranchial apparatus in a Cretaceous dinosaur and the antiquity of avian paraglossalia". Linnean Society'nin Zooloji Dergisi. 175 (4): 892–909. doi:10.1111/zoj.12293.
- ^ Haas, G. (1969). "On the jaw musculature of ankylosaurs". Amerikan Müzesi Novitates. 2399: 1–11. hdl:2246/2609.
- ^ Rybczynski, N .; Vickaryous, M. K. (2001). "Chapter 14: Evidence of Complex Jaw Movement in the Late Cretaceous Ankylosaurid, Euoplocephalus tutuş (Dinosauria: Thyreophora) ". K. Carpenter (ed.). Zırhlı Dinozorlar. Indiana University Press. s. 299–317. ISBN 978-0-253-33964-5.
- ^ a b Maryanska, T. (1977). "Ankylosauridae (Dinosauria) from Mongolia" (PDF). Paleontoloji Polonica. 37: 85–151.
- ^ a b Miyashita, T.; Arbour V. M.; Witmer L. M.; Currie, P. J. (2011). "Zırhlı bir dinozorun iç kafatası morfolojisi Euoplocephalus X-ışını bilgisayarlı tomografik rekonstrüksiyon ile desteklendi " (PDF). Anatomi Dergisi. 219 (6): 661–75. doi:10.1111 / j.1469-7580.2011.01427.x. PMC 3237876. PMID 21954840. Arşivlenen orijinal (PDF) 24 Eylül 2015.
- ^ Coombs, W. (1978). "Ankylosauria'nın (Reptilia, Ornithischia) ön ayak kasları". Paleontoloji Dergisi. 52 (3): 642–57. JSTOR 1303969.
- ^ a b Coombs, W. (1979). "Osteology and myology of the hindlimb in the Ankylosauria (Reptillia, Ornithischia)". Paleontoloji Dergisi. 53 (3): 666–84. JSTOR 1304004.
- ^ Burns, M; Tumanova, T; Currie, P (2015). "Postcrania of juvenile Pinacosaurus grangeri (Ornithischia: Ankylosauria) from the Upper Cretaceous Alagteeg Formation, Alag Teeg, Mongolia: implications for ontogenetic allometry in ankylosaurs". Paleontoloji Dergisi. 89 (1): 168–182. doi:10.1017/jpa.2014.14. S2CID 130610291.
- ^ Currie, P. J .; Badamgarav, D .; Koppelhus, E. B .; Sissons, R.; Vickaryous, M. K. (2011). "Hands, feet, and behaviour in Pinacosaurus (Dinosauria: Ankylosauridae)" (PDF). Acta Palaeontologica Polonica. 56 (3): 489–504. doi:10.4202/app.2010.0055. S2CID 129291148.
- ^ Scheyer, T. M .; Sander, P. M. (2004). "Histology of ankylosaur osteoderms: implications for systematics and function". Omurgalı Paleontoloji Dergisi. 24 (4): 874–93. doi:10.1671/0272-4634(2004)024[0874:hoaoif]2.0.co;2. JSTOR 4524782.
- ^ Coombs W. (1972). "Kemikli Göz Kapağı Euoplocephalus (Reptilia, Ornithischia) ". Paleontoloji Dergisi. 46 (5): 637–50. JSTOR 1303019..
- ^ Carpenter, K. (1982). "İskelet ve dermal zırh rekonstrüksiyonu Euoplocephalus tutuş (Ornithischia: Ankylosauridae) Alberta'nın Geç Kretase yaşlı Oldman Formasyonundan ". Kanada Yer Bilimleri Dergisi. 19 (4): 689–97. Bibcode:1982CaJES..19..689C. doi:10.1139 / e82-058.
- ^ Arbor, V.M. (2009). "Estimating impact forces of tail club strikes by ankylosaurid dinosaurs". PLOS ONE. 4 (8): e6738. Bibcode:2009PLoSO ... 4,6738A. doi:10.1371 / journal.pone.0006738. PMC 2726940. PMID 19707581.
- ^ Thulborn, T. (1993). "Mimicry in ankylosaurid dinosaurs". Güney Avustralya Müzesi Kayıtları. 27: 151–58.
- ^ Vickaryous, M. K.; Maryanska, T.; Weishampel, D. B. (2004). "Ankilosauria". Weishampel, D. B .; Dodson, P .; Osmólska, H. (editörler). Dinosauria. California Üniversitesi Yayınları. pp. 363–92. ISBN 978-0-520-24209-8.
- ^ a b Weishampel, D. B.; Barrett, P. M .; Coria, R. A .; Le Loeuff, J .; Xu X .; Zhao X.; Sahni, A .; Gomani, E. M. P.; Noto, C. R. (2004). "Dinosaur Distribution". Weishampel, D. B .; Dodson, P .; Osmolska, H.. (eds.). The Dinosauria (2nd). California Üniversitesi Yayınları. pp. 517–606. ISBN 978-0-520-24209-8.
- ^ Lofgren, D. F. (1997). "Hell Creek Formation". Currie, P.J .; Padian, K. (editörler). Dinozorlar Ansiklopedisi. Akademik Basın. pp. 302–03. ISBN 978-0-12-226810-6.
- ^ Breithaupt, B. H. (1997). "Lance Formation". Currie, P.J .; Padian, K. (editörler). Dinozorlar Ansiklopedisi. Akademik Basın. s. 394–95. ISBN 978-0-12-226810-6.
- ^ Eberth, D. A. (1997). "Edmonton Group". Currie, P. J .; Padian, K. (editörler). Dinozorlar Ansiklopedisi. Akademik Basın. s. 199–204. ISBN 978-0-12-226810-6.
- ^ Johnson, K. R. (1997). "Hell Creek Flora". Currie, P. J .; Padian, K. (editörler). Dinozorlar Ansiklopedisi. Akademik Basın. pp. 300–02. ISBN 978-0-12-226810-6.
- ^ Bigelow, P. "Cretaceous 'Hell Creek Faunal Facies'; Late Maastrichtian". Arşivlendi from the original on December 26, 2009. Alındı 24 Mart 2014.
Dış bağlantılar
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||


| Yetki kontrolü |
|---|