Tyrannosaurus - Tyrannosaurus
| Tyrannosaurus | |
|---|---|
 | |
| Yeniden yapılanma T. rex tip numune (CM 9380) Carnegie Doğa Tarihi Müzesi | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Aile: | †Tyrannosauridae |
| Alt aile: | †Tyrannosaurinae |
| Cins: | †Tyrannosaurus Osborn, 1905 |
| Türler | |
| †Tyrannosaurus rex Osborn, 1905 | |
| Diğer Türler | |
| |
| Eş anlamlı | |
Cins eşanlamlı
Tür eşanlamlı
| |
Tyrannosaurus[nb 1] bir cins nın-nin Coelurosaurian Theropod Dinozor. Türler Tyrannosaurus rex (Rex anlamı "kral" Latince ), genellikle denir T. rex veya halk dilinde T-Rex, büyük theropodların en iyi temsil edilenlerinden biridir. Tyrannosaurus şimdi batıda yaşadı Kuzey Amerika, o zamanlar ada kıtası olarak bilinen yerde Laramidia. Tyrannosaurus diğerlerinden çok daha geniş bir menzile sahipti tyrannosaurids. Fosiller çeşitli bulunur Kaya oluşumları ile çıkmak Maastrihtiyen yaş üst Kretase dönem, 68 - 66milyon yıl önce. Tiranozorların bilinen son üyesidir ve son olmayanlar arasındakuş dinozorların daha önce var olması Kretase-Paleojen nesli tükenme olayı.
Diğer tyrannosaurids gibi, Tyrannosaurus bir iki ayaklı etobur uzun, ağır bir kuyrukla dengelenmiş devasa bir kafatası ile. Büyük ve güçlü arka bacaklarına göre, Tyrannosaurus ön ayaklar kısaydı, ancak boyutlarına göre alışılmadık derecede güçlüydü ve iki tırnaklı rakamı vardı. En eksiksiz numune, 12,3 metreye (40 fit) kadar uzunluktadır. T. rex Kalçalarda 12,3 m (40 ft), 3,96 m (13 ft) uzunluğa kadar ve en modern tahminlere göre 8,4 metrik ton (9,3 kısa ton) ile 14 metrik ton (15,4 kısa ton) arasında büyüyebilir. ağırlık. Diğer theropodlar rekabet etse veya aşsa da Tyrannosaurus rex içinde boyut Hala bilinen en büyük kara avcıları arasındadır ve tüm kara hayvanları arasında en güçlü ısırık kuvvetini uyguladığı tahmin edilmektedir. Çevresindeki en büyük etobur, Tyrannosaurus rex büyük ihtimalle bir uç yırtıcı avlanmak hadrosaurlar, zırhlı otçullar gibi Ceratopsia'lılar ve Ankylosaurlar ve muhtemelen Sauropodlar. Bazı uzmanlar dinozorun öncelikle bir çöpçü. Olup olmadığı sorusu Tyrannosaurus apeks bir yırtıcı mıydı yoksa saf bir çöpçü, en uzun tartışmalar arasındaydı paleontoloji. Bugün çoğu paleontolog bunu kabul ediyor Tyrannosaurus hem aktif bir avcı hem de bir çöpçüydü.
Örnekleri Tyrannosaurus rex neredeyse tamamlanmış iskeletlerden bazılarını içerir. Yumuşak doku ve proteinler bu örneklerden en az birinde rapor edilmiştir. Fosil materyalinin bolluğu, yaşam öyküsü ve biyomekanik. Beslenme alışkanlıkları, fizyoloji ve potansiyel hız Tyrannosaurus rex birkaç tartışma konusudur. Onun taksonomi bazı bilim adamlarının düşündüğü gibi tartışmalı Tarbosaurus bataar Asya'dan ikinci olmak Tyrannosaurus diğerleri korurken türler Tarbosaurus ayrı bir cinstir. Kuzey Amerika tyrannosauridlerinin diğer birkaç cinsi de eşanlamlı ile Tyrannosaurus.
Arketipik theropod olarak, Tyrannosaurus 20. yüzyılın başlarından beri en iyi bilinen dinozorlardan biri olmuştur ve film, reklam, posta pulları ve diğer birçok medyada yer almıştır.
Araştırma tarihi
İlk buluntular

Şimdi belgelenen dişlerden dişler Tyrannosaurus rex tarafından 1874'te bulundu Arthur Gölleri yakın Altın, Colorado. 1890'ların başında, John Bell Hatcher doğuda postkraniyal unsurlar toplandı Wyoming. Fosillerin büyük türlerden olduğuna inanılıyordu Ornitomimus grandis (şimdi Deinodon ) ama şimdi kabul ediliyor T. rex kalır.[2]
1892'de, Edward Drinker Cope büyük dinozorun iki omur parçası buldu. Cope, parçaların bir "agathaumid" e ait olduğuna inanıyordu (Ceratopsid ) dinozor ve onlara isim verdi Manospondylus gigasKemikte bulduğu kan damarları için sayısız açıklığa atıfta bulunarak "dev gözenekli omur" anlamına geliyor.[2] M. gigas Kalıntılar, 1907'de Hatcher tarafından bir ceratopsidden ziyade bir theropodun kalıntıları olarak tanımlandı.[3]
Henry Fairfield Osborn arasındaki benzerliği fark etti Manospondylus gigas ve T. rex 1917 gibi erken bir tarihte, bu sırada ikinci omur kaybolmuştu. Parçalı yapısı nedeniyle Manospondylus omurlar, Osborn iki cinsi eşanlamlı hale getirmedi, bunun yerine eski cinsin belirsiz olduğunu düşünüyordu.[4] Haziran 2000'de Black Hills Enstitüsü yaklaşık% 10 oranında bulundu Tyrannosaurus iskelet (BHI 6248) orijinal olabilecek bir sitede M. gigas yerellik.[5]
İskelet keşfi ve adlandırma

Barnum Brown asistan küratör Amerikan Doğa Tarihi Müzesi, ilk kısmi iskeleti buldu T. rex 1900'de Wyoming'in doğusunda. Brown, başka bir kısmi iskelet buldu. Hell Creek Oluşumu Yaklaşık 34 fosilleşmiş kemik içeren, 1902'de Montana'da.[6] O sırada Brown şunları söyledi: "Taş Ocağı No. 1 uyluk kemiği, kasık kemiği, humerus, üç omur ve tarafından tanımlanmamış büyük bir Etçil Dinozorun iki belirlenmemiş kemiğini içeriyor. Bataklık.... daha önce hiç böyle bir şey görmedim Kretase ".[7] Henry Fairfield Osborn, başkanı Amerikan Doğa Tarihi Müzesi, adı ikinci iskelet T. rex Genel ad, Yunan kelimeler τύραννος (Tyrannos, "tiran" anlamına gelir) ve σαῦρος (Sauros, "kertenkele" anlamına gelir). Osborn kullandı Latince kelime Rex, belirli ad için "kral" anlamına gelir. Dolu iki terimli bu nedenle, "zalim kertenkele kral" veya "Kral Zorba Kertenkele" anlamına gelir ve hayvanın büyüklüğünü ve zamanın diğer türleri üzerinde algılanan hakimiyetini vurgular.[6]

Osborn diğer örneği adlandırdı Dynamosaurus imperiosus 1905'te bir makalede.[6] 1906'da Osborn, iki iskeletin aynı türden olduğunu fark etti ve Tyrannosaurus tercih edilen isim olarak.[8] Orijinal Dinamozor malzeme koleksiyonlarında bulunur Doğal Tarih Müzesi, Londra.[9] 1941'de T. rex tip numune satıldı Carnegie Doğa Tarihi Müzesi Pittsburgh, Pennsylvania'da 7.000 dolara.[7] Dinamozor Daha sonra, Andrew McDonald ve meslektaşlarının 2018 yılında başka bir tyrannosaurid türü açıklamasıyla onurlandırılacaktı, Dinamoterror hanedanları McDonald's'ın "çocukluk favorisi" olduğu için adı 1905 ismine referans olarak seçilmiş.[10]
1910'lardan 1950'lerin sonuna kadar Barnum'un keşifleri, Tyrannosaurusolarak Büyük çöküntü ve savaşlar birçok paleontoloğu sahanın dışında tuttu.[5]
Dirilen ilgi

1960'lardan başlayarak, yeni bir ilgi vardı TyrannosaurusBatı Kuzey Amerika'dan 42 iskeletin (kemik sayımına göre% 5-80 tamamlanmış) geri kazanılmasıyla sonuçlandı.[5] 1967'de Dr. William MacMannis, kemik sayımına göre% 15 oranında tamamlanmış ve yeniden yapılandırılmış kafatasına sahip "MOR 008" adlı iskeleti bulmuş ve geri kazanmıştır. Rockies Müzesi. 1990'larda, bugüne kadar bulunan en eksiksiz iskeletlerden ikisi de dahil olmak üzere, önceki yılların neredeyse iki katı kadar çok buluntu ile çok sayıda keşif gördü: Dava açmak ve Stan.[5]
Sue Hendrickson, bir amatör paleontolog, en eksiksiz (yaklaşık% 85) ve en büyüğü keşfetti Tyrannosaurus iskelet Hell Creek Oluşumu keşfeden adıyla anılan Sue örneği, mülkiyeti konusunda yasal bir savaşın hedefi oldu. 1997'de dava, asıl arazi sahibi Maurice Williams lehine sonuçlandı. Fosil koleksiyonu, Saha Doğa Tarihi Müzesi 7,6 milyon dolarlık açık artırmada, onu bugüne kadarki en pahalı dinozor iskeleti yapıyor. Field Doğa Tarihi Müzesi personeli, 1998'den 1999'a kadar 25.000 saatin üzerinde kayayı kemiklerden çıkarmak için harcadı.[11] Kemikler daha sonra New Jersey Montajın yapıldığı yer, ardından son montaj için Chicago'ya geri gönderildi. Takılı iskelet, Field Doğa Tarihi Müzesi'nde 17 Mayıs 2000'de halka açıldı. Bu örneğin fosilleşmiş kemikleri üzerinde yapılan bir araştırma, Sue'nun 19 yaşında tam boyuta ulaştığını ve bilinen herhangi bir tiranozorun tahmini en uzun ömrü olan 28 yaşında öldüğünü gösterdi.[12]

Bir diğeri TyrannosaurusStan (BHI 3033) takma adı, amatör paleontolog Stan Sacrison'un onuruna, 1992'de Hell Creek Formasyonundan kurtarıldı. Stan, bulunan en eksiksiz ikinci iskelettir ve toplamın% 70'ini temsil eden 199 kemik bulunmuştur.[13] Bu tiranozor ayrıca, kırık ve iyileşmiş kaburgalar, kırılmış (ve iyileşmiş) bir boyun ve kafasının arkasında, yaklaşık bir boyunda büyük bir delik dahil olmak üzere birçok kemik patolojisine sahipti. Tyrannosaurus diş.[14]
1998'de Bucky Derflinger, Bucky's ayak parmakları yerin üzerinde açığa çıkarak, o zamanlar 20 yaşında olan Derflinger'i Tyrannosaurus. Örnek, 3.0 metre (10 ft) boyunda ve 11 metre (35 ft) uzunluğunda genç bir yetişkindi. Bucky ilk Tyrannosaurus korunmuş bulunan bulundu furcula (salıncak). Bucky, kalıcı olarak şurada görüntüleniyor: Indianapolis Çocuk Müzesi.[15]

2000 yazında, ekipler tarafından Jack Horner beş keşfedildi Tyrannosaurus yakınındaki iskeletler Fort Peck Rezervuarı.[16] 2001 yılında, bir gencin% 50 tam iskeleti Tyrannosaurus Hell Creek Formasyonu'nda bir ekip tarafından keşfedildi. Burpee Doğa Tarihi Müzesi. Jane (BMRP 2002.4.1) olarak adlandırılan bulgunun bilinen ilk iskelet olduğu düşünülüyordu. pigme tyrannosaurid, Nanotyrannus, ancak sonraki araştırmalar, bunun daha büyük olasılıkla bir çocuk olduğunu ortaya koydu Tyrannosaurusve bilinen en eksiksiz çocuk örneği;[17] Jane, Burpee Doğa Tarihi Müzesi'nde sergileniyor.[18] 2002 yılında, amatör koleksiyoncular Dan Wells ve Don Wyrick tarafından keşfedilen Wyrex adlı bir iskeletin 114 kemiği vardı ve% 38'i tamamlandı. Kazı, 2004 yılında, Black Hills Enstitüsü ilk canlıyla internet üzerinden Tyrannosaurus günlük raporlar, fotoğraflar ve videolar sağlayan kazı.[5]
2006 yılında Montana Eyalet Üniversitesi en büyüğüne sahip olduğunu ortaya çıkardı Tyrannosaurus kafatası henüz keşfedildi (MOR 008 adlı bir örnekten), 5 fit (152 cm) uzunluğunda.[19] Daha sonraki karşılaştırmalar, en uzun başlığın 136,5 santimetre (53,7 inç) (LACM 23844 örneğinden) ve en geniş başın 90,2 santimetre (35,5 inç) (Sue'dan) olduğunu gösterdi.[20]
Ayak izi

İki izole fosilleşmiş ayak izi geçici olarak atandı T. rex. İlk keşfedildi Philmont Scout Çiftliği, New Mexico, 1983'te Amerikalı jeolog Charles Pillmore tarafından. Başlangıçta bir hadrosaurid, ayak izinin incelenmesi, içinde bilinmeyen büyük bir 'topuk' ortaya çıkardı. ornitopod dinozor izleri ve ne olabileceğinin izleri halluks, tiranozor ayağının çiy çiçeği benzeri dördüncü rakamı. Ayak izi olarak yayınlandı Ichnogenus Tyrannosauripus pillmorei 1994 yılında Martin Lockley ve Adrian Hunt. Lockley ve Hunt, parkurun büyük olasılıkla bir T. rex, bu onu bu türün bilinen ilk ayak izi yapar. Parkur, bir zamanlar bitki örtüsüyle kaplı çamur düzlüğünde yapılmıştı. 83 santimetre (33 inç) uzunluğunda ve 71 santimetre (28 inç) genişliğindedir.[21]
Bir tarafından yapılmış olabilecek ikinci bir ayak izi Tyrannosaurus ilk olarak 2007 yılında İngiliz paleontolog Phil Manning tarafından rapor edilmiştir. Hell Creek Oluşumu Montana. Bu ikinci parça, Lockley ve Hunt tarafından açıklanan pistten daha kısa olan 72 santimetre (28 inç) uzunluğundadır. Parçanın yapıp yapmadığı Tyrannosaurus yine de belirsiz Tyrannosaurus ve Nanotyrannus Hell Creek Formasyonunda var olduğu bilinen tek büyük theropodlardır.[22][23]
Wyoming, Glenrock'ta, Maastrihtiyen Geç Kretase evresi ve Lance Formasyonu Scott Persons, Phil Currie ve meslektaşları tarafından 2016 yılında tanımlanmıştır ve bir çocuğa ait olduğuna inanılır. T. rex veya şüpheli tyrannosaurid Nanotyrannus lancensis. Ölçümlerden ve ayak izlerinin pozisyonlarına göre, hayvanın saatte 2,8 ila 5 mil yürüme hızında gittiğine ve 1,56 m (5,1 ft) ila 2,06 m (6,8 m) arasında bir kalça yüksekliğine sahip olduğu tahmin edildi. ft).[24][25][26] 2017'de hız tahminlerini% 50-80 artıran bir takip raporu yayınlandı.[27]
Açıklama
Boyut
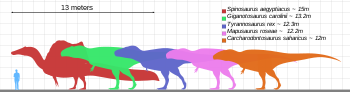
T. rex tüm zamanların en büyük kara etoburlarından biriydi. Takma adı verilen en büyük ve en eksiksiz örneklerden biri Dava açmak (FMNH PR2081), Saha Doğa Tarihi Müzesi. Sue 12,3-12,8 metre (40-42 ft) uzunluğunda ölçüldü,[28][29] kalçalarda 3,66 metre (12 ft) uzunluğundaydı,[30] ve en son araştırmalara göre, çeşitli teknikler kullanılarak, 8.4 metrik ton (9.3 kısa ton) ile 14 metrik ton (15.4 kısa ton) arasında olduğu tahmin edilmektedir.[29][31] Scotty (RSM P2523.8) lakaplı bir örnek Kraliyet Saskatchewan Müzesi, 13 m (43 ft) uzunluğunda olduğu bildirilmektedir. Bir kütle tahmin tekniğinin kullanılması çevre Scotty, ağırlık olarak 8.8 metrik ton (9.7 kısa ton) ile bilinen en büyük örnek olarak tahmin edildi.[32][33]
Her yetişkin değil Tyrannosaurus Elde edilen numune kadar büyük. Tarihsel olarak ortalama yetişkin kütle tahminleri, 4,5 metrik ton (5,0 kısa ton) kadar düşük olan yıllar içinde büyük farklılıklar göstermiştir.[34][35] 7,2 metrik tondan (7,9 kısa ton) fazla,[36] En modern tahminler 5.4 metrik ton (6.0 kısa ton) ile 8.0 metrik ton (8.8 kısa ton) arasında değişiyor.[29][37][38][39][40]
İskelet

Bilinen en büyük T. rex kafatası 1.52 metre (5 ft) uzunluğundadır.[30] Büyük Fenestrae Kafatasındaki (açıklıklar), tüm etçil theropodlarda olduğu gibi ağırlığı azalttı. Diğer açılardan Tyrannosauruskafatası büyük olmayanlardan önemli ölçüde farklıydı.Tyrannosaurid theropods. Arkada son derece genişti ancak dar bir burnu vardı, alışılmadık derecede iyi dürbün görüşü.[41][42] Kafatası kemikleri çok büyüktü ve burun delikleri ve diğer bazı kemikler kaynaşarak aralarındaki hareketi engelledi; ama çoğu pnömatik hale getirilmiş (küçük hava boşluklarından oluşan bir "bal peteği" içeriyordu) ve dolayısıyla daha hafif. Bunlar ve diğer kafatasını güçlendiren özellikler, Tyrannosaurid tüm tiranozorid olmayanları kolayca geride bırakan, giderek daha güçlü bir ısırığa doğru eğilim.[43][44][45] Üst çenenin ucu U şeklindeydi (çoğu tyrannosauroid etoburun V şeklinde üst çeneleri vardı), bu da bir tiranozorun tek bir ısırıkla çıkarabileceği doku ve kemik miktarını artırdı, ancak ön taraftaki baskıyı da artırdı. diş.[46]

Dişleri T. rex işaretli görüntülendi heterodonluk (şekil farklılıkları).[47][48] premaksiller üst çenenin ön tarafında her tarafta dört tane olan dişler sıkıca paketlenmişti, Denine kesit şeklinde, arka yüzeyinde güçlendirici sırtlar bulunan, kesici (uçları keski benzeri bıçaklardı) ve geriye doğru kıvrılmıştı. Dşeklindeki enine kesit, güçlendirici sırtlar ve geriye doğru eğim, dişlerin kırılma riskini azalttı. Tyrannosaurus biraz ve çekti. Kalan dişler, hançerlerden ziyade "öldürücü muzlar" gibi sağlamdı, daha geniş aralıklıydı ve aynı zamanda güçlendirici çıkıntılara sahipti.[49] Üst çenede olanlar, olgun bireylerde her iki tarafta on iki,[47] arka hariç, alt çenenin muadillerinden daha büyüktü. Şimdiye kadar bulunan en büyüğünün, hayvan canlıyken kökü dahil 30,5 santimetre (12 inç) uzunluğunda olduğu tahmin ediliyor ve bu, onu şimdiye kadar bulunan herhangi bir etobur dinozorun en büyük dişi yapıyor.[50] Alt çene sağlamdı. Önü diş kemiği on üç diş taşıyordu. Diş sırasının arkasında, alt çene belirgin şekilde daha uzun hale geldi.[47] Üst ve alt çeneleri Tyrannosaurus, birçok dinozorunki gibi, çok sayıda foramina veya kemikte küçük delikler. Bu foramina için timsah benzeri bir duyu sistemi gibi çeşitli işlevler önerilmiştir.[51] veya kanıtı ağız dışı yapılar pullar veya potansiyel olarak dudaklar gibi.[52][53][54]
Omurga nın-nin Tyrannosaurus on boyun omuru, on üç sırt omuru ve beş sakral omurdan oluşuyordu. Kuyruk omurlarının sayısı bilinmemektedir ve bireyler arasında pekala farklılık gösterebilirdi, ancak muhtemelen en az kırk numaradır. Sue'ya bu türden kırk yedi kuyruk omuru takıldı.[47] Boyun T. rex diğer theropodlarınki gibi doğal bir S-şekilli eğri oluşturdu. Bunlarla karşılaştırıldığında, muazzam başı desteklemek için son derece kısa, derin ve kaslıydı. İkinci omur, eksen özellikle kısaydı. Kalan boyun omurları zayıf bir şekilde opistocoelözdü, yani omur gövdesinin dışbükey bir ön kısmı ve içbükey bir arka. Omurga gövdelerinde tek pleurocoels vardı, pnömatik çöküntüler hava keseciklerinin, yanlarında.[47] Gövdenin omur gövdeleri sağlamdı ancak dar bir bele sahipti. Alt tarafları sallanmıştı. Ön taraflar derin bir dikey çukur ile içbükeydi. Büyük pleurocoelleri vardı. Sinir dikenlerinin güçlü tendonların bağlanması için çok sert ön ve arka tarafları vardı. Sakral omurlar hem omur gövdelerinde hem de sinir dikenlerinde birbirine kaynaşmıştı. Pnömatik hale getirildiler. Pelvise enine süreçler ve sakral kaburgalarla bağlandılar. Kuyruk, masif baş ve gövdeyi dengelemek ve masifler için alan sağlamak için ağır ve orta derecede uzundu. lokomotor kaslar uyluk kemiklerine bağlı. On üçüncü kuyruk omuru, derin kuyruk tabanı ile orta kuyruk arasındaki geçiş noktasını oluşturdu ve bu, oldukça uzun ön eklemlenme süreçleriyle sertleşti. Gövdenin alt tarafı on sekiz veya on dokuz çift parçalı göbek kaburga ile kaplıydı.[47]

omuz kuşağı tüm ön ayaklardan daha uzundu. Kürek kemiği dar bir şafta sahipti, ancak üst ucunda istisnai bir şekilde genişledi. Uzun bir öne çıkıntıyla bağlanmıştır. korakoid, yuvarlaktı. Her iki omuz bıçağı da küçük bir furcula. Eşleştirilmiş göğüs kemikleri muhtemelen şunlardan yapılmıştır: kıkırdak sadece.[47]

Ön ayaklar veya kol çok kısaydı. Üst kol kemiği olan humerus kısa ama sağlamdı. Olağanüstü yuvarlak başlı dar bir üst uca sahipti. Alt kol kemikleri, ulna ve yarıçap, humerustan çok daha kısa olan düz elemanlardı. İkinci metakarpal Normalde theropodlarda bunun tersi doğruyken, birincisinden daha uzun ve daha genişti. Ön ayakların sadece iki pençeli parmağı vardı,[47] ek bir atel benzeri küçük üçüncü ile birlikte metakarpal üçüncü bir rakamın kalıntısını temsil eder.[55]
leğen kemiği büyük bir yapıydı. Üst kemiği, ilium hem çok uzun hem de yüksekti ve arka bacak kasları için geniş bir bağlantı alanı sağlıyordu. Ön kasık kemiği elemanın tüm şaftından daha uzun, muazzam bir kasık botuyla sona erdi. Arka ischium ince ve düzdü, eğik bir şekilde arkaya ve aşağıya işaret ediyordu.[47]
Kolların aksine, herhangi bir theropodun vücut boyutuyla orantılı olarak arka ayaklar en uzun kollar arasındaydı. Ayakta metatars "arctometatarsalian" idi, yani üçüncü metatarsalın ayak bileğine yakın kısmının sıkıştığı anlamına geliyordu. Üçüncü metatars da son derece kıvrımlıydı.[47] Hayvanın muazzam kütlesini telafi etmek için, iskelet boyunca birçok kemik oyulmuş ve önemli bir güç kaybı olmaksızın ağırlığı azaltılmıştır.[47]
Sınıflandırma

Tyrannosaurus ... tip üst ailenin cinsi Tyrannosauroidea, aile Tyrannosauridae ve Tyrannosaurinae alt ailesi; başka bir deyişle, paleontologların aynı gruba başka türleri dahil edip etmemeye karar verdikleri standarttır. Tyrannosaurine alt ailesinin diğer üyeleri arasında Kuzey Amerika Daspletosaurus ve Asya Tarbosaurus,[17][56] ikisi de ara sıra ile eşanlamlı hale gelmiştir Tyrannosaurus.[57] Tyrannosauridlerin bir zamanlar genellikle daha önceki büyük theropodların torunları olduğu düşünülüyordu. megalozorlar ve karnozorlar, ancak daha yakın zamanda, genellikle daha küçük olanla yeniden sınıflandırılmışlardır. Coelurosaurs.[46]

1955'te Sovyet paleontolog Evgeny Maleev yeni bir tür adlandırdı, Tyrannosaurus bataar, şuradan Moğolistan.[58] 1965'e gelindiğinde, bu türün adı değiştirildi Tarbosaurus bataar.[59] Yeniden adlandırmaya rağmen, birçok filogenetik analizler bulundu Tarbosaurus bataar olmak kardeş takson nın-nin T. rex,[56] ve genellikle bir Asya türü olarak kabul edilmiştir. Tyrannosaurus.[46][60][61] Tyrannosaurid'in keşfi Lythronax ayrıca şunu gösterir Tarbosaurus ve Tyrannosaurus yakın akraba, diğer Asyalı tiranozorlarla bir kuşak oluşturuyor Zhuchengtyrannus, ile Lythronax onların kardeş taksonu olmak.[62][63] Steve Brusatte, Thomas Carr ve meslektaşları tarafından 2016'da yapılan bir başka araştırma da şunu gösteriyor: Tyrannosaurus Asya'dan gelen bir göçmen olmasının yanı sıra olası bir soyundan Tarbosaurus.[64]
2001 yılında, çeşitli tyrannosaurid dişler ve bir metatarsal, yakınlardaki bir taş ocağında ortaya çıkarıldı. Zhucheng Çin, Çinli paleontolog tarafından atandı Hu Chengzhi yeni dikilene Tyrannosaurus zhuchengensis. Bununla birlikte, yakın bir bölgede, yeni kurulan tyrannosaurid cinsine bir sağ maksilla ve sol çene kemiği atandı. Zhuchengtyrannus 2011'de ve mümkün T. zhuchengensis dır-dir eşanlamlı ile Zhuchengtyrannus. Her halükârda, T. zhuchengensis olarak kabul edilir nomen dubium holotip eksik olduğu için tanı Tyrannosaurinae seviyesinin altındaki özellikler.[65]
Aşağıda, Tyrannosauridae'nin kladogramı bulunmaktadır. Filogenetik analiz Loewen ve arkadaşları tarafından 2013 yılında yapılmıştır.[62]
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Nanotyrannus

Aynı oluşumlarda bulunan diğer tyrannosaurid fosilleri T. rex başlangıçta ayrı taksonlar olarak sınıflandırıldılar: Aublysodon ve Albertosaurus megagracilis,[57] ikincisi adlandırılıyor Dinotyrannus megagracilis 1995'te.[66] Bu fosiller artık evrensel olarak gençliğe ait kabul ediliyor T. rex.[67] Montana'dan 60 santimetre (2.0 ft) uzunluğunda küçük ama neredeyse tamamlanmış bir kafatası bir istisna olabilir. Bu kafatası, CMNH 7541, başlangıçta bir tür olarak sınıflandırıldı Gorgosaurus (G. lancensis) tarafından Charles W. Gilmore 1946'da.[68] 1988'de örnek yeniden tanımlandı Robert T. Bakker, Phil Currie ve o zamanlar Cleveland Doğa Tarihi Müzesi'nde paleontoloji küratörü olan Michael Williams, orijinal örneğin barındırıldığı ve şimdi sergileniyor. İlk araştırmaları, kafatası kemiklerinin kaynaştığını ve bu nedenle yetişkin bir örneği temsil ettiğini gösterdi. Bunun ışığında, Bakker ve meslektaşları kafatasını adlı yeni bir cinse atadı. Nanotyrannus (görünüşte küçük yetişkin boyutu için "cüce zorba" anlamına gelir). Öldüğünde numunenin yaklaşık 5,2 metre (17 ft) uzunluğunda olduğu tahmin edilmektedir.[69] Ancak 1999 yılında, Thomas Carr Örneğin, Carr ve diğer birçok paleontoloğun bir çocuk olduğunu düşünmesine yol açan bir genç olduğunu ortaya çıkardı. T. rex bireysel.[70][71]

2001 yılında, daha eksiksiz bir genç tiranozor (takma adı "Jane ", katalog numarası BMRP 2002.4.1), orijinali ile aynı türe ait Nanotyrannus örnek, ortaya çıkarıldı. Bu keşif, tiranozorlar üzerine şu konulara odaklanan bir konferans başlattı: Nanotyrannus geçerlilik Burpee Doğa Tarihi Müzesi Daha önce bazı paleontologlar N. lancensis Currie ve Williams da dahil olmak üzere geçerli bir türdü, "Jane" keşfini Nanotyrannus aslında bir çocuktu T. rex.[72][73][74] Peter Larson hipotezini desteklemeye devam etti N. lancensis Her iki çenede iki diş daha olması gibi kafatası özelliklerine dayanan ayrı ama yakından ilişkili bir türdü. T. rex; üçüncü metakarpalde falankslar ile orantılı olarak daha büyük eller ve farklı salıncak tanımlanmamış bir örnekte anatomi. Ayrıca şunu da savundu Stygivenatorgenellikle çocuk olarak kabul edilir T. rex, daha genç olabilir Nanotyrannus örnek.[75][76] Daha sonra yapılan araştırmalar, diğer tyrannosauridlerin Gorgosaurus ayrıca büyüme sırasında diş sayısında azalma yaşandı,[70] ve bu cinsteki aynı yaş grubundaki bireyler arasındaki diş sayısındaki eşitsizlik göz önüne alındığında ve Tyrannosaurusbu özelliğin nedeni de olabilir bireysel varyasyon.[71] 2013 yılında Carr, tüm farklılıkların Nanotyrannus bireysel veya ontogenetik olarak değişken özellikler veya ürünler olduğu ortaya çıktı. kemiklerin bozulması.[77]

2016 yılında, Persons ve Currie tarafından uzuv oranlarının analizi önerildi Nanotyrannus farklı yetenek seviyelerine sahip olan örnekler, potansiyel olarak onu T. rex.[78] Bununla birlikte, paleontolog Manabu Sakomoto, bu sonucun düşük seviyeden etkilenebileceğini söyledi. örnek boyut ve tutarsızlık taksonomik ayrımı yansıtmaz.[79] Joshua Schmerge, 2016 yılında NanotyrannusBMRP 2002.4.1'in kafatasında bir diş oluğu da dahil olmak üzere kafatası özelliklerine dayalı geçerlilik. Schmerge'e göre, bu özellik, T. rex ve sadece içinde bulundu Dryptosaurus ve albertosaurinler, bu öneriyor Nanotyrannus Albertosaurinae içinde farklı bir taksondur.[80] Aynı yıl, Carr ve meslektaşları bunun açıklığa kavuşturmak için yeterli olmadığını belirttiler. Nanotyrannustiranozoroidler arasında ortak ve ontogenetik olarak değişken bir özellik olan geçerlilik veya sınıflandırma.[81]
Holly Woodward ve meslektaşları tarafından yapılan bir 2020 çalışması, atıfta bulunulan örnekleri gösterdi. Nanotyrannus hepsi ontogenetik olarak olgunlaşmamıştı ve bu örneklerin ait olduğu olası bulundu. T. rex.[82] Aynı yıl, Carr hakkında bir makale yayınladı. T. RexCMNH 7541'in taksonun beklenen ontogenetik varyasyonuna uyduğunu ve diğer örneklerde bulunan juvenil özelliklerini gösterdiğini bulan büyüme geçmişi. Kafatası 80 cm'den (31 inç) küçük olan 13 yaşından küçük bir çocuk olarak sınıflandırıldı. Carr, potansiyel filogenetik önemi olan karakterlerin yaş boyunca büyümeyle aynı oranda azaldığını belirterek, incelenen 44 örnek arasında önemli bir cinsel veya filogenetik varyasyon fark edilmedi.[83] Gazetenin sonuçlarını tartışan Carr, bunun nasıl olduğunu anlattı "Nanotyrannus"Örnekler, en küçük yavrular ve alt yetişkinler arasında sürekli bir büyüme geçişi oluşturdu, örneklerin gruplanarak gruplanacağı ayrı bir takson olsaydı beklenenin aksine Tyrannosaurus. Carr, "nanomorfların" birbirine pek benzemediği ve bunun yerine büyüme serilerinde önemli bir köprü oluşturduğu sonucuna vardı. T. rex gençlerin sığ kafatasından, tam gelişmiş yetişkinlerde görülen derin kafatasına kadar derin bir değişimin başlangıcını yakalar. "[84]
Paleobiyoloji
Hayat hikayesi

Birkaç örneğin genç olarak tanımlanması T. rex bilim adamlarının belgelemesine izin verdi ontogenetik türlerdeki değişiklikler, yaşam süresini tahmin edin ve hayvanların ne kadar hızlı büyüyeceğini belirleyin. Bilinen en küçük birey (LACM 28471, "Jordan theropod") sadece 30 kg (66 lb) ağırlığında olduğu tahmin edilirken, en büyüğü, örneğin FMNH PR2081 (Sue) büyük olasılıkla yaklaşık 5.650 kg (12.460 lb) ağırlığındaydı. Histolojik analizi T. rex kemikler, LACM 28471'in öldüğünde sadece 2 yaşında olduğunu, Sue'nun ise türler için maksimuma yakın olabilecek bir yaş olduğunu gösterdi.[37]
Histoloji ayrıca diğer örneklerin yaşının da belirlenmesine izin verdi. Büyüme eğrileri, farklı örneklerin yaşları, kütleleri ile birlikte bir grafik üzerine çizildiğinde geliştirilebilir. Bir T. rex Büyüme eğrisi S şeklindedir ve yavrular, vücut büyüklüğünün önemli ölçüde artmaya başladığı yaklaşık 14 yaşına kadar 1.800 kg (4.000 lb) altında kalmıştır. Bu hızlı büyüme evresinde bir genç T. rex önümüzdeki dört yıl boyunca yılda ortalama 600 kg (1.300 lb) kazanacaktı. 18 yaşında, eğri tekrar düzleşir ve büyümenin dramatik bir şekilde yavaşladığını gösterir. Örneğin, yalnızca 600 kg (1.300 lb) 28 yaşındaki Sue'yu 22 yaşındaki Kanada örneğinden ayırdı (RTMP 81.12.1).[37] Farklı işçiler tarafından yapılan 2004 histolojik bir çalışma, bu sonuçları doğrulayarak, hızlı büyümenin yaklaşık 16 yaşında yavaşlamaya başladığını bulmuştur.[85]
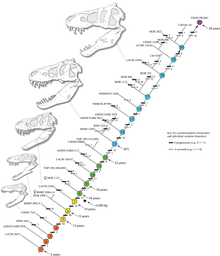
Hutchinson ve meslektaşlarının 2011'de yaptığı bir çalışma, genel olarak önceki tahmin yöntemlerini doğruladı, ancak en yüksek büyüme oranları tahminleri önemli ölçüde daha yüksek; "üstel aşamada T. rex için maksimum büyüme oranlarının 1790 kg / yıl" olduğunu buldu.[29] Bu sonuçlar önceki tahminlerden çok daha yüksek olmasına rağmen, yazarlar bu sonuçların gerçek büyüme oranı ile büyüklüğünde bir hayvandan beklenebilecek olan arasındaki büyük farkı önemli ölçüde azalttığını belirtti.[29] Büyüme hamlesinin sonunda büyüme hızındaki ani değişiklik, fiziksel olgunluğa işaret edebilir, bu hipotez, medüller doku keşfiyle desteklenen bir hipotezdir. uyluk 16 ila 20 yaşındaki birinin T. rex Montana'dan (DAHA 1125, B-rex olarak da bilinir). Medüller doku sadece yumurtlama sırasında dişi kuşlarda bulunur, bu da B-rex'in üreme çağında olduğunu gösterir.[86] Daha fazla çalışma, bu örnek için 18 yaşında olduğunu gösteriyor.[87] 2016'da nihayet Mary Higby Schweitzer ve Lindsay Zanno ve meslektaşları tarafından MOR 1125'in femurundaki yumuşak dokunun medüller doku olduğu doğrulandı. Bu aynı zamanda numunenin dişi kimliğini de doğruladı. İçerisindeki medüller kemik dokusunun keşfi Tyrannosaurus Medüller dokunun kimyasal yapısı şüphe götürmez olduğundan, gelecekteki incelemelerde diğer dinozor türlerinin cinsiyetini belirlemede değerli olabilir.[88] Diğer tyrannosauridler, daha düşük yetişkin boyutlarına karşılık gelen daha düşük büyüme oranlarına rağmen, son derece benzer büyüme eğrileri sergiler.[89]
Dergi için Woodward ve meslektaşları tarafından 2020'de yayınlanan ek bir çalışma Bilim Gelişmeleri çocukluktan yetişkine büyümeleri sırasında, Tyrannosaurus gıda eksikliği gibi çevresel faktörlere karşı koymak için büyümesini yavaşlatabiliyordu. Illinois'deki Burpee Müzesi'nde barındırılan 13 ila 15 yaşları arasındaki iki çocuk örneğine odaklanan çalışma, olgunlaşma oranının Tyrannosaurus kaynak bolluğuna bağlıydı. Bu çalışma aynı zamanda böyle değişen ortamlarda Tyrannosaurus kaynak bolluğu açısından her yıl değişen bir ortama özellikle çok uygundur, bu da diğer orta büyüklükteki yırtıcı hayvanların bu tür sert koşullarda hayatta kalmakta zorluk çekebileceklerini ve genç ve yetişkin tiranozorlar arasındaki niş bölünmeyi açıklamakta zorlandıklarını ima ediyordu. Çalışma ayrıca şunu göstermektedir: Tyrannosaurus ve şüpheli cins Nanotyrannus incelenen iki örneğin kemiklerindeki büyüme halkalarının analizi nedeniyle eş anlamlıdır.[90][91]
Bilinenlerin yarısından fazlası T. rex Örnekler, cinsel olgunluğa ulaştıktan sonra altı yıl içinde ölmüş gibi görünüyor; bu, diğer tiranozorlarda ve günümüzde bazı büyük, uzun ömürlü kuşlarda ve memelilerde de görülen bir model. Bu türler, yüksek bebek ölüm oranları ve ardından gençler arasında nispeten düşük ölüm oranları ile karakterize edilir. Cinsel olgunluktan sonra, kısmen üremenin stresine bağlı olarak ölüm tekrar artar. Bir çalışma, gençlerin nadir olmasının T. rex fosiller kısmen düşük çocuk ölüm oranlarından kaynaklanmaktadır; hayvanlar bu yaşlarda çok sayıda ölmüyorlardı ve bu yüzden de fosilleşmemişlerdi. Bu nadirlik, aynı zamanda eksikliğinden de kaynaklanıyor olabilir. fosil kaydı ya da fosil toplayıcıların daha büyük, daha muhteşem örneklere eğilimine.[89] Thomas Holtz Jr., 2013'te verdiği bir konferansta, dinozorların "hızlı yaşadıklarını ve genç öldüklerini" öne sürerken, memelilerin uzun ömürleri vardır çünkü üremeleri daha uzun sürer.[92] Gregory S.Paul da şunu yazıyor: Tyrannosaurus çabuk çoğaldı ve genç yaşta öldü, ancak kısa ömürlerini yaşadıkları tehlikeli yaşamlara bağlıyor.[93]
Deri ve olası ipliksi tüylenme

Keşfi tüylü dinozorlar olup olmadığı ve ne ölçüde olduğu konusunda tartışmaya yol açtı. Tyrannosaurus tüylü olabilir.[94][95] Yaygın olarak öncüleri olarak tanınan ipliksi yapılar tüyler, küçük gövdeli bazal tyrannosauroid'de rapor edilmiştir. Dilong paradoksu Erken Kretase'den Yixian Formasyonu 2004 yılında Çin.[96] Çünkü bütüncül O dönemde bilinen daha büyük tyrannosauroidlerin izlenimleri, ölçekler, okuyan araştırmacılar Dilong Yalıtım tüylerinin, yüzey / hacim oranlarının daha küçük olması nedeniyle daha büyük türler tarafından kaybedilmiş olabileceğini tahmin etti.[96] Dev türlerin sonraki keşfi Yutyrannus huali, yine Yixian'dan, bazı büyük tiranozoroidlerin bile vücutlarının çoğunu kaplayan tüylere sahip olduklarını göstererek, boyutla ilgili bir özellik oldukları hipotezine şüphe uyandırdı.[97] 2017'de yapılan bir çalışma, tiranozorların bilinen deri izlenimlerini inceledi. Tyrannosaurus kuyruk, kalça ve boyundaki mozaik pul parçalarını koruyan "Wyrex" (BHI 6230) lakaplı örnek.[5] Çalışma, büyük tiranozorların tüy örtüsünün, örneğin Tyrannosaurus varsa, gövdenin üst tarafı ile sınırlıydı.[94]
2016'da yayınlanan bir konferans özeti, theropodların Tyrannosaurus üst dişlerinin görüldüğü gibi çıplak dişler yerine dudaklarla kapatılması timsahlar. Bu, varlığına dayanıyordu emaye Çalışmaya göre sulu kalması gereken, timsahlar gibi suda yaşayan hayvanların karşılaşmadığı bir sorun.[53] 2017 tarihli bir analitik çalışma, tiranozorların dudakları yerine burunlarında büyük, düz pullara sahip olduğunu öne sürdü.[51][98] Bununla birlikte, dudak fikrini desteklediği eleştiriler olmuştur. Timsahların gerçekte düz pulları yok, aksine, tiranozorların hummocky buruşukluğunu gözlemleyerek ve bunu mevcut kertenkelelerle karşılaştırarak, tiranozorların timsah benzeri bir cilt yerine skuamoz pullara sahip olduğunu gördüler.[99][100]
Cinsel dimorfizm

Bilinen örneklerin sayısı arttıkça, bilim adamları bireyler arasındaki farklılıkları analiz etmeye başladılar ve iki farklı vücut tipi gibi görünen şeyleri keşfettiler. morflar, diğer bazı theropod türlerine benzer. Bu morflardan biri daha sağlam bir şekilde inşa edildiğinden, 'sağlam' morf, diğeri 'zarif '. Birkaç morfolojik iki morfla ilişkili farklılıklar analiz etmek için kullanıldı cinsel dimorfizm içinde T. rex'sağlam' morf ile genellikle dişi olduğu öne sürülür. Örneğin, leğen kemiği birkaç 'sağlam' numuneden, belki de geçişine izin vermek için daha geniş görünüyordu. yumurtalar.[101] Ayrıca, 'sağlam' morfolojinin azalmış bir şerit İlk kuyruk omurunda, ayrıca görünürde yumurtaların dışarı çıkmasına izin vermek için üreme organları yanlışlıkla rapor edildiği gibi timsahlar.[102]
Son yıllarda, cinsel dimorfizmin kanıtları zayıfladı. 2005 yılında yapılan bir çalışma, timsah şevron anatomisindeki cinsel dimorfizm iddialarının hatalı olduğunu, bu da benzer dimorfizmin varlığı konusunda şüphe uyandırdığını bildirdi. T. rex cinsiyetler.[103] Son derece sağlam bir birey olan Sue'nun ilk kuyruk omurunda tam boyutlu bir şivron keşfedildi, bu da bu özelliğin iki şekli ayırt etmek için zaten kullanılamayacağını gösteriyor. Gibi T. rex örnekler bulundu Saskatchewan -e Yeni Meksika, bireyler arasındaki farklılıklar, cinsel dimorfizmden çok coğrafi farklılığın göstergesi olabilir. Farklılıklar yaşla ilgili olabilir, 'sağlam' bireyler daha yaşlı hayvanlar olabilir.[47]
Sadece tek T. rex specimen has been conclusively shown to belong to a specific sex. Examination of B-rex demonstrated the preservation of yumuşak doku within several bones. Some of this tissue has been identified as a medullary tissue, a specialized tissue grown only in modern birds as a source of kalsiyum üretimi için yumurta kabuğu sırasında yumurtlama. As only female birds lay eggs, medullary tissue is only found naturally in females, although males are capable of producing it when injected with female reproductive hormonlar sevmek estrojen. This strongly suggests that B-rex was female, and that she died during ovulation.[86] Recent research has shown that medullary tissue is never found in crocodiles, which are thought to be the closest living relatives of dinosaurs, aside from birds. The shared presence of medullary tissue in birds and theropod dinosaurs is further evidence of the close evrimsel ikisi arasındaki ilişki.[104]
Duruş

Birçok gibi iki ayaklı dinozorlar T. rex was historically depicted as a 'living tripod', with the body at 45 degrees or less from the vertical and the tail dragging along the ground, similar to a kanguru. This concept dates from Joseph Leidy 's 1865 reconstruction of Hadrosaurus, the first to depict a dinosaur in a bipedal posture.[105] In 1915, convinced that the creature stood upright, Henry Fairfield Osborn, former president of the American Museum of Natural History, further reinforced the notion in unveiling the first complete T. rex skeleton arranged this way. It stood in an upright pose for 77 years, until it was dismantled in 1992.[106]
By 1970, scientists realized this pose was incorrect and could not have been maintained by a living animal, as it would have resulted in the çıkık or weakening of several eklemler, including the hips and the articulation between the head and the omurga.[107] The inaccurate AMNH mount inspired similar depictions in many films and paintings (such as Rudolph Zallinger 's famous mural The Age of Reptiles içinde Yale Üniversitesi 's Peabody Doğa Tarihi Müzesi )[108] until the 1990s, when films such as Jurassic Park introduced a more accurate posture to the general public.[109] Modern representations in museums, art, and film show T. rex with its body approximately parallel to the ground with the tail extended behind the body to balance the head.[110]
To sit down, Tyrannosaurus may have settled its weight backwards and rested its weight on a pubic boot, the wide expansion at the end of the pubis in some dinosaurs. With its weight rested on the pelvis, it may have been free to move the hindlimbs. Getting back up again might have involved some stabilization from the diminutive forelimbs.[111][107] The latter known as Newman's pushup theory has been debated. Her şeye rağmen, Tyrannosaurus was probably able to get up if it fell, which only would have required placing the limbs below the center of gravity, with the tail as an effective counterbalance.[112]
Silâh

Ne zaman T. rex was first discovered, the humerus was the only element of the forelimb known.[6] For the initial mounted skeleton as seen by the public in 1915, Osborn substituted longer, three-fingered forelimbs like those of Allosaurus.[4] Bir yıl önce, Lawrence Lambe described the short, two-fingered forelimbs of the closely related Gorgosaurus.[113] This strongly suggested that T. rex had similar forelimbs, but this hipotez was not confirmed until the first complete T. rex forelimbs were identified in 1989, belonging to MOR 555 (the "Wankel rex").[114][115] The remains of Sue also include complete forelimbs.[47] T. rex arms are very small relative to overall body size, measuring only 1 meter (3.3 ft) long, and some scholars have labelled them as körelmiş. The bones show large areas for kas attachment, indicating considerable strength. This was recognized as early as 1906 by Osborn, who speculated that the forelimbs may have been used to grasp a mate during çiftleşme.[8] It has also been suggested that the forelimbs were used to assist the animal in rising from a prone position.[107]
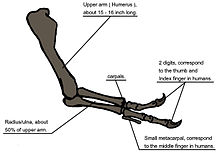
Another possibility is that the forelimbs held struggling prey while it was killed by the tyrannosaur's enormous jaws. This hypothesis may be supported by biyomekanik analizi. T. rex forelimb bones exhibit extremely thick kortikal kemik, which has been interpreted as evidence that they were developed to withstand heavy loads. biceps brachii muscle of an adult T. rex was capable of lifting 199 kilograms (439 lb) by itself; other muscles such as the Brachialis would work along with the biceps to make elbow flexion even more powerful. M. biceps muscle of T. rex was 3.5 times as powerful as the insan eşdeğeri. Bir T. rex forearm had a limited range of motion, with the shoulder and elbow joints allowing only 40 and 45 degrees of motion, respectively. In contrast, the same two joints in Deinonychus allow up to 88 and 130 degrees of motion, respectively, while a human arm can rotate 360 degrees at the shoulder and move through 165 degrees at the elbow. The heavy build of the arm bones, strength of the muscles, and limited range of motion may indicate a system evolved to hold fast despite the stresses of a struggling prey animal. In the first detailed scientific description of Tyrannosaurus forelimbs, paleontologists Kenneth Carpenter and Matt Smith dismissed notions that the forelimbs were useless or that T. rex zorunlu bir çöpçüydü.[116]
According to paleontologist Steven M. Stanley, the 1 metre (3.3 ft) arms of T. rex were used for slashing prey, especially by using its claws to rapidly inflict long, deep gashes to its prey, although this concept is disputed by others believing the arms were used for grasping a sexual partner.[117]
Termoregülasyon

As of 2014, it is not clear if Tyrannosaurus oldu endotermik (“warm-blooded”). Tyrannosaurus, like most dinosaurs, was long thought to have an ektotermik ("cold-blooded") reptilian metabolizma. The idea of dinosaur ectothermy was challenged by scientists like Robert T. Bakker ve John Ostrom in the early years of the "Dinosaur Renaissance ", beginning in the late 1960s.[118][119] T. rex itself was claimed to have been endotermik ("warm-blooded"), implying a very active lifestyle.[35] Since then, several paleontologists have sought to determine the ability of Tyrannosaurus -e düzenlemek its body temperature. Histological evidence of high growth rates in young T. rex, comparable to those of mammals and birds, may support the hypothesis of a high metabolism. Growth curves indicate that, as in mammals and birds, T. rex growth was limited mostly to immature animals, rather than the belirsiz büyüme seen in most other omurgalılar.[85]
Oksijen izotopu ratios in fossilized bone are sometimes used to determine the temperature at which the bone was deposited, as the ratio between certain isotopes correlates with temperature. In one specimen, the isotope ratios in bones from different parts of the body indicated a temperature difference of no more than 4 to 5 °C (7 to 9 °F) between the vertebrae of the torso and the tibia of the lower leg. This small temperature range between the body core and the extremities was claimed by paleontologist Reese Barrick and jeokimyacı William Showers to indicate that T. rex maintained a constant internal body temperature (Homeothermy ) and that it enjoyed a metabolism somewhere between ectothermic reptiles and endothermic mammals.[120] Other scientists have pointed out that the ratio of oxygen isotopes in the fossils today does not necessarily represent the same ratio in the distant past, and may have been altered during or after fossilization (diyajenez ).[121] Barrick and Showers have defended their conclusions in subsequent papers, finding similar results in another theropod dinosaur from a different continent and tens of millions of years earlier in time (Giganotosaurus ).[122] Ornithischian dinosaurs also showed evidence of homeothermy, while Varanid kertenkele from the same formation did not.[123] Bile T. rex does exhibit evidence of homeothermy, it does not necessarily mean that it was endothermic. Such thermoregulation may also be explained by gigantotermi, as in some living Deniz kaplumbağaları.[124][125][126] Similar to contemporary alligators, dorsotemporal fenestra in Tyrannosaurus's skull may have aided thermoregulation.[127]
Yumuşak doku
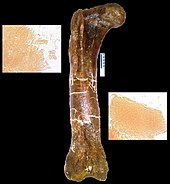
Mart 2005 sayısında Bilim, Mary Higby Schweitzer nın-nin Kuzey Karolina Eyalet Üniversitesi and colleagues announced the recovery of soft tissue from the marrow cavity of a fossilized leg bone from a T. rex. The bone had been intentionally, though reluctantly, broken for shipping and then not preserved in the normal manner, specifically because Schweitzer was hoping to test it for soft tissue.[128] Designated as the Museum of the Rockies specimen 1125, or MOR 1125, the dinosaur was previously excavated from the Hell Creek Oluşumu. Flexible, bifurcating kan damarları and fibrous but elastic kemik matrix tissue were recognized. In addition, microstructures resembling kan hücreleri were found inside the matrix and vessels. The structures bear resemblance to devekuşu blood cells and vessels. Whether an unknown process, distinct from normal fossilization, preserved the material, or the material is original, the researchers do not know, and they are careful not to make any claims about preservation.[129] If it is found to be original material, any surviving proteins may be used as a means of indirectly guessing some of the DNA content of the dinosaurs involved, because each protein is typically created by a specific gene. The absence of previous finds may be the result of people assuming preserved tissue was impossible, therefore not looking. Since the first, two more tyrannosaurs and a hadrosaur have also been found to have such tissue-like structures.[128] Research on some of the tissues involved has suggested that birds are closer relatives to tyrannosaurs than other modern animals.[130]
In studies reported in Bilim in April 2007, Asara and colleagues concluded that seven traces of kolajen proteins detected in purified T. rex bone most closely match those reported in tavuklar, followed by frogs and newts. The discovery of proteins from a creature tens of millions of years old, along with similar traces the team found in a mastodon bone at least 160,000 years old, upends the conventional view of fossils and may shift paleontologists' focus from bone hunting to biochemistry. Until these finds, most scientists presumed that fossilization replaced all living tissue with inert minerals. Paleontologist Hans Larsson of McGill University in Montreal, who was not part of the studies, called the finds "a milestone", and suggested that dinosaurs could "enter the field of molecular biology and really slingshot paleontology into the modern world".[131]
The presumed soft tissue was called into question by Thomas Kaye of the Washington Üniversitesi and his co-authors in 2008. They contend that what was really inside the tyrannosaur bone was slimy biyofilm created by bacteria that coated the voids once occupied by blood vessels and cells.[132] The researchers found that what previously had been identified as remnants of blood cells, because of the presence of iron, were actually framboids, microscopic mineral spheres bearing iron. They found similar spheres in a variety of other fossils from various periods, including an ammonit. In the ammonite they found the spheres in a place where the iron they contain could not have had any relationship to the presence of blood.[133] Schweitzer has strongly criticized Kaye's claims and argues that there is no reported evidence that biofilms can produce branching, hollow tubes like those noted in her study.[134] San Antonio, Schweitzer and colleagues published an analysis in 2011 of what parts of the collagen had been recovered, finding that it was the inner parts of the collagen coil that had been preserved, as would have been expected from a long period of protein degradation.[135] Other research challenges the identification of soft tissue as biofilm and confirms finding "branching, vessel-like structures" from within fossilized bone.[136]
Hız

Scientists have produced a wide range of possible maximum running speeds for Tyrannosaurus, mostly around 11 meters per second (40 km/h; 25 mph), but as low as 5–11 meters per second (18–40 km/h; 11–25 mph) and as high as 20 meters per second (72 km/h; 45 mph). Estimates that Tyrannosaurus had relatively larger leg muscles than any animal alive today but it was so massive that it was not likely to run very fast at all compared to other theropods like Giganotosaurus.[137] Researchers have relied on various estimating techniques because, while there are many izler of large theropods walking, none had the pattern of running.[138]
A 2002 report used a mathematical model (validated by applying it to three living animals: timsahlar, tavuklar, ve insanlar; and eight more species, including emus and ostriches[138]) to gauge the leg muscle mass needed for fast running (over 40 km/h or 25 mph).[137] Scientists who think that Tyrannosaurus was able to run point out that hollow bones and other features that would have lightened its body may have kept adult weight to a mere 4.5 metric tons (5.0 short tons) or so, or that other animals like devekuşları ve atlar with long, flexible legs are able to achieve high speeds through slower but longer strides.[138] Proposed top speeds exceeded 40 kilometers per hour (25 mph) for Tyrannosaurus, but were deemed infeasible because they would require exceptional leg muscles of approximately 40–86% of total body mass. Even moderately fast speeds would have required large leg muscles. If the muscle mass was less, only 18 kilometers per hour (11 mph) for walking or jogging would have been possible.[137] Holtz noted that tyrannosaurids and some closely related groups had significantly longer uzak hindlimb components (shin plus foot plus toes) relative to the femur length than most other theropods, and that tyrannosaurids and their close relatives had a tightly interlocked metatars (foot bones).[139] The third metatarsal was squeezed between the second and fourth metatarsals to form a single unit called an arctometatarsus. This ankle feature may have helped the animal to run more efficiently.[140] Together, these leg features allowed Tyrannosaurus to transmit locomotory forces from the foot to the lower leg more effectively than in earlier theropods.[139]

Additionally, a 2020 study indicates that Tyrannosaurus and other tyrannosaurids were exceptionally efficient walkers. Studies by Dececchi et al., compared the leg proportions, body mass, and the gaits of more than 70 species of theropod dinosaurs including Tyrannosaurus ve akrabaları. The research team then applied a variety of methods to estimate each dinosaur's top speed when running as well as how much energy each dinosaur expended while moving at more relaxed speeds such as when walking. Among smaller to medium-sized species such as dromaeosaurids, longer legs appear to be an adaptation for faster running, in line with previous results by other researchers. But for theropods weighing over 1,000 kg (2,200 lb), top running speed is limited by body size, so longer legs instead were found to have correlated with low-energy walking. The results further indicate that smaller theropods evolved long legs as a means to both aid in hunting and escape from larger predators while larger theropods that evolved long legs did so to reduce the energy costs and increase foraging efficiency, as they were freed from the demands of predation pressure due to their role as apex predators. Compared to more basal groups of theropods in the study, tyrannosaurs like Tyrannosaurus itself showed a marked increase in foraging efficiency due to reduced energy expenditures during hunting or scavenging. This in turn likely resulted in tyrannosaurs having a reduced need for hunting forays and requiring less food to sustain themselves as a result. Additionally, the research, in conjunction with studies that show tyrannosaurs were more agile than other large bodied-theropods, indicates they were quite well-adapted to a long-distance stalking approach followed by a quick burst of speed to go for the kill. Analogies can be noted between tyrannosaurids and modern wolves as a result, supported by evidence that at least some tyrannosaurids were hunting in group settings.[141][142]
A 2017 study estimated the top running speed of Tyrannosaurus as 17 mph (27 km/h), speculating that Tyrannosaurus exhausted its energy reserves long before reaching top speed, resulting in a parabola-like relationship between size and speed.[143][144] Another 2017 study hypothesized that an adult Tyrannosaurus was incapable of running due to high skeletal loads. Using a calculated weight estimate of 7 tons, the model showed that speeds above 11 mph (18 km/h) would have probably shattered the leg bones of Tyrannosaurus. The finding may mean that running was also not possible for other giant theropod dinosaurs like Giganotosaurus, Mapusaurus ve Akrokantosaurus.[145]However, studies by Eric Snively and colleagues, published in 2019 indicate that Tyrannosaurus and other tyrannosaurids were more maneuverable than allosauroids and other theropods of comparable size due to low rotational inertia compared to their body mass combined with large leg muscles. As a result, it is hypothesized that Tyrannosaurus was capable of making relatively quick turns and could likely pivot its body more quickly when close to its prey, or that while turning, the theropod could "pirouette" on a single planted foot while the alternating leg was held out in a suspended swing during pursuit. The results of this study potentially could shed light on how agility could have contributed to the success of tyrannosaurid evolution.[146]
Brain and senses

Tarafından yürütülen bir çalışma Lawrence Witmer and Ryan Ridgely of Ohio University found that Tyrannosaurus shared the heightened sensory abilities of other Coelurosaurs, highlighting relatively rapid and coordinated eye and head movements; an enhanced ability to sense low frequency sounds, which would allow tyrannosaurs to track prey movements from long distances; and an enhanced sense of smell.[147] A study published by Kent Stevens concluded that Tyrannosaurus had keen vision. By applying modified perimetri to facial reconstructions of several dinosaurs including Tyrannosaurus, the study found that Tyrannosaurus had a binocular range of 55 degrees, surpassing that of modern hawks. Stevens estimated that Tyrannosaurus had 13 times the visual acuity of a human and surpassed the visual acuity of an eagle, which is 3.6 times that of a person. Stevens estimated a limiting far point (that is, the distance at which an object can be seen as separate from the horizon) as far as 6 km (3.7 mi) away, which is greater than the 1.6 km (1 mi) that a human can see.[41][42][148]
Thomas Holtz Jr. would note that high depth perception of Tyrannosaurus may have been due to the prey it had to hunt, noting that it had to hunt horned dinosaurs such as Triceratops, armored dinosaurs such as Ankylosaurus, and the duck-billed dinosaurs and their possibly complex social behaviors. He would suggest that this made precision more crucial for Tyrannosaurus enabling it to, "get in, get that blow in and take it down." Tersine, Akrokantosaurus had limited depth perception because they hunted large sauropods, which were relatively rare during the time of Tyrannosaurus.[92]
Tyrannosaurus had very large koku soğanları ve koku alma sinirleri relative to their brain size, the organs responsible for a heightened sense of smell. This suggests that the sense of smell was highly developed, and implies that tyrannosaurs could detect carcasses by scent alone across great distances. The sense of smell in tyrannosaurs may have been comparable to modern akbabalar, which use scent to track carcasses for scavenging. Research on the olfactory bulbs has shown that T. rex had the most highly developed sense of smell of 21 sampled non-avian dinosaur species.[149]

Somewhat unusually among theropods, T. rex had a very long koklea. The length of the cochlea is often related to hearing acuity, or at least the importance of hearing in behavior, implying that hearing was a particularly important sense to tyrannosaurs. Specifically, data suggests that T. rex heard best in the low-frequency range, and that low-frequency sounds were an important part of tyrannosaur behavior.[147] A 2017 study by Thomas Carr and colleagues found that the snout of tyrannosaurids was highly sensitive, based on a high number of small openings in the facial bones of the related Daspletosaurus that contained duyusal nöronlar. The study speculated that tyrannosaurs might have used their sensitive snouts to measure the temperature of their nests and to gently pick-up eggs and hatchlings, as seen in modern crocodylians.[51]
A study by Grant R. Hurlburt, Ryan C. Ridgely and Lawrence Witmer obtained estimates for Encephalization Quotients (EQs), based on reptiles and birds, as well as estimates for the ratio of cerebrum to brain mass. Çalışma şu sonuca vardı: Tyrannosaurus had the relatively largest brain of all adult non-avian dinosaurs with the exception of certain small maniraptoriforms (Bambiraptor, Troodon ve Ornitomimus ). Çalışma şunu buldu Tyrannosaurus 's relative brain size was still within the range of modern reptiles, being at most 2 Standart sapma above the mean of non-avian reptile EQs. The estimates for the ratio of cerebrum mass to brain mass would range from 47.5 to 49.53 percent. According to the study, this is more than the lowest estimates for extant birds (44.6 percent), but still close to the typical ratios of the smallest sexually mature alligators which range from 45.9–47.9 percent.[150] Other studies, such as those by Steve Brusatte, indicate the encephalization quotient of Tyrannosaurus was similar in range (2.0-2.4) to a şempanze (2.2-2.5), though this may be debatable as reptilian and mammalian encephalization quotients are not equivalent.[151]
Sosyal davranış

Suggesting that Tyrannosaurus olabilir pack hunters, Philip J. Currie karşılaştırıldığında T. rex to related species Tarbosaurus bataar ve Albertosaurus lahit, citing fossil evidence that may indicate pack behavior.[152] A find in Güney Dakota nerede üç T. rex skeletons were in close proximity suggested a pack.[153][154] Because available prey such as Triceratops ve Ankylosaurus had significant defenses, it may have been effective for T. rex to hunt in groups.[152]
Currie's pack-hunting hypothesis has been criticized for not having been hakemli, but rather was discussed in a television interview and book called Dino Gangs.[155] The Currie theory for pack hunting by T. rex is based mainly by analogy to a different species, Tarbosaurus bataar, and that the supposed evidence for pack hunting in T. bataar itself had not yet been peer-reviewed. According to scientists assessing the Dino Gangs program, the evidence for pack hunting in Tarbosaurus ve Albertosaurus is weak and based on skeletal remains for which alternate explanations may apply (such as drought or a flood forcing dinosaurs to die together in one place).[155] Fosilleşmiş yol Üst Kretase'den Wapiti Formasyonu kuzeydoğunun Britanya Kolumbiyası, Canada, left by three tyrannosaurids traveling in the same direction, may also indicate packs.[156][157]
Evidence of intraspecific attack were found by Joseph Peterson and his colleagues in the juvenile Tyrannosaurus takma isim Jane. Peterson and his team found that Jane's skull showed healed puncture wounds on the upper jaw and snout which they believe came from another juvenile Tyrannosaurus. Subsequent CT scans of Jane's skull would further confirm the team's hypothesis, showing that the puncture wounds came from a traumatic injury and that there was subsequent healing.[158] The team would also state that Jane's injuries were structurally different from the parasite-induced lesions found in Sue and that Jane's injuries were on her face whereas the parasite that infected Sue caused lesions to the lower jaw.[159]
Besleme stratejileri


Most paleontologists accept that Tyrannosaurus was both an active yırtıcı ve bir çöpçü like most large etobur.[160] By far the largest carnivore in its environment, T. rex büyük ihtimalle bir uç yırtıcı, preying upon hadrosaurlar, armored herbivores like Ceratopsia'lılar ve Ankylosaurlar ve muhtemelen Sauropodlar.[161] A study in 2012 by Karl Bates and Peter Falkingham found that Tyrannosaurus had the most powerful bite of any terrestrial animal that has ever lived, finding an adult Tyrannosaurus could have exerted 35,000 to 57,000 N (7,868 to 12,814 lbf ) of force in the back teeth.[162][163][164] Even higher estimates were made by Mason B. Meers in 2003.[44] This allowed it to crush bones during repetitive biting and fully consume the carcasses of large dinosaurs.[20] Stephan Lautenschlager and colleagues calculated that Tyrannosaurus was capable of a maximum jaw gape of around 80 degrees, a necessary adaptation for a wide range of jaw angles to power the creature's strong bite.[165][166]
A debate exists, however, about whether Tyrannosaurus was primarily a yırtıcı or a pure çöpçü; the debate was assessed in a 1917 study by Lambe which argued Tyrannosaurus was a pure scavenger because the Gorgosaurus teeth showed hardly any wear.[167] This argument may not be valid because theropods replaced their teeth quite rapidly. Ever since the first discovery of Tyrannosaurus most scientists have speculated that it was a predator; like modern large predators it would readily scavenge or steal another predator's kill if it had the opportunity.[168]
Paleontolog Jack Horner has been a major proponent of the view that Tyrannosaurus was not a predator at all but instead was exclusively a scavenger.[114][169][170] He has put forward arguments in the popular literature to support the pure scavenger hypothesis:
- Tyrannosaur arms are short when compared to other known predators. Horner argues that the arms were too short to make the necessary gripping force to hold on to prey.[170]
- Tyrannosaurs had large koku soğanları ve koku alma sinirleri (relative to their brain size). These suggest a highly developed sense of smell which could sniff out carcasses over great distances, as modern akbabalar yapmak. Research on the olfactory bulbs of dinosaurs has shown that Tyrannosaurus had the most highly developed sense of smell of 21 sampled dinosaurs.[149]
- Tyrannosaur teeth could crush bone, and therefore could extract as much food (kemik iliği ) as possible from carcass remnants, usually the least nutritious parts. Karen Chin and colleagues have found bone fragments in coprolites (fossilized feces) that they attribute to tyrannosaurs, but point out that a tyrannosaur's teeth were not well adapted to systematically chewing bone like sırtlanlar do to extract marrow.[171]
- Since at least some of Tyrannosaurus's potential prey could move quickly, evidence that it walked instead of ran could indicate that it was a scavenger.[169] On the other hand, recent analyses suggest that Tyrannosaurus, while slower than large modern terrestrial predators, may well have been fast enough to prey on large hadrosaurlar ve Ceratopsia'lılar.[137][23]
Other evidence suggests hunting behavior in Tyrannosaurus. The eye sockets of tyrannosaurs are positioned so that the eyes would point forward, giving them dürbün görüşü slightly better than that of modern şahinler. It is not obvious why Doğal seçilim would have favored this long-term trend if tyrannosaurs had been pure scavengers, which would not have needed the advanced derinlik algısı o stereoskopik görüş sağlar.[41][42] In modern animals, binocular vision is found mainly in predators.

A skeleton of the hadrosaurid Edmontosaurus Annectens has been described from Montana with healed tyrannosaur-inflicted damage on its tail omur. The fact that the damage seems to have healed suggests that the Edmontosaurus survived a tyrannosaur's attack on a living target, i.e. the tyrannosaur had attempted active predation.[172] Despite the consensus that the tail bites were caused by Tyrannosaurus, there has been some evidence to show that they might have been created by other factors. For example, a 2014 study suggested that the tail injuries might have been due to Edmontosaurus individuals stepping on each other,[173] while another study in 2020 backs up the hypothesis that biomechanical stress is the cause for the tail injuries.[174]. There is also evidence for an aggressive interaction between a Triceratops ve bir Tyrannosaurus in the form of partially healed tyrannosaur tooth marks on a Triceratops brow horn and skuamozal (a bone of the boyun fırfır ); the bitten horn is also broken, with new bone growth after the break. It is not known what the exact nature of the interaction was, though: either animal could have been the aggressor.[175] Beri Triceratops wounds healed, it is most likely that the Triceratops survived the encounter and managed to overcome the Tyrannosaurus. In a battle against a bull Triceratops, Triceratops would likely defend itself by inflicting fatal wounds to the Tyrannosaurus using its sharp horns.[176] Çalışmaları Dava açmak found a broken and healed fibula and tail vertebrae, scarred facial bones and a tooth from another Tyrannosaurus embedded in a neck vertebra, providing evidence for aggressive behavior.[177] Studies on hadrosaur vertebrae from the Hell Creek Formation that were punctured by the teeth of what appears to be a late-stage juvenile Tyrannosaurus indicate that despite lacking the bone-crushing adaptations of the adults, young individuals were still capable of using the same bone-puncturing feeding technique as their adult counterparts.[178]
Tyrannosaurus may have had infectious tükürük used to kill its prey, as proposed by William Abler in 1992. Abler observed that the tırtıllar (tiny protuberances) on the cutting edges of the teeth are closely spaced, enclosing little chambers. These chambers might have trapped pieces of carcass with bacteria, giving Tyrannosaurus a deadly, infectious bite much like the Komodo Ejderhası was thought to have.[179][180] Jack Horner and Don Lessem, in a 1993 popular book, questioned Abler's hypothesis, arguing that Tyrannosaurus's tooth serrations as more like cubes in shape than the serrations on a Komodo monitor's teeth, which are rounded.[114]:214–215
Tyrannosaurus, and most other theropods, probably primarily processed carcasses with lateral shakes of the head, like crocodilians. The head was not as maneuverable as the skulls of allosauroidler, due to flat joints of the neck vertebrae.[181]
Patoloji

In 2001, Bruce Rothschild and others published a study examining evidence for Gerilme kırıkları ve tendon avülsiyonları içinde Theropod dinozorlar ve davranışlarının etkileri. Since stress fractures are caused by repeated trauma rather than singular events they are more likely to be caused by regular behavior than other types of injuries. Of the 81 Tyrannosaurus foot bones examined in the study one was found to have a stress fracture, while none of the 10 hand bones were found to have stress fractures. Araştırmacılar tendon avülsiyonlarını yalnızca Tyrannosaurus ve Allosaurus. Bir avülsiyon yaralanması, Sue'nun kol kemiğinde bir boşluk bıraktı. T. rex, görünüşe göre kökeninde bulunuyor deltoid veya teres majör kaslar. Avülsiyon yaralanmalarının varlığı, her ikisinde de ön ayaklar ve omuz ile sınırlıdır. Tyrannosaurus ve Allosaurus theropodların kuşlarınkinden daha karmaşık ve işlevsel olarak farklı bir kas yapısına sahip olabileceğini düşündürmektedir. Araştırmacılar, Sue'nun tendon avülsiyonunun muhtemelen mücadele eden avlardan elde edildiği sonucuna vardı. Genel olarak stres kırıkları ve tendon avülsiyonlarının mevcudiyeti, zorunlu temizlemeden ziyade "çok aktif" bir avcı temelli diyet için kanıt sağlar.[182]
2009 yılında yapılan bir araştırma, birkaç numunenin kafataslarındaki pürüzsüz kenarlı deliklerin nedeninin Trichomonas yaygın olarak enfekte olan benzeri parazitler kuşlar. "Sue" ve MOR 980 ("Peck's Rex") dahil olmak üzere ciddi şekilde enfekte olmuş kişiler, bu nedenle beslenme giderek zorlaştıktan sonra açlıktan ölmüş olabilirler. Daha önce bu delikler bakteriyel kemik enfeksiyonu ile açıklanmıştı. Aktinomikoz veya tür içi saldırılar yoluyla.[183]
Bir çalışma Tyrannosaurus Aynı cinse atfedilebilen kemiklerde diş izleri olan örnekler, kanıt olarak sunuldu. yamyamlık.[184] Diş izleri humerus, ayak kemikleri ve metatarsallar, başka bir kişiyle çatışmanın yol açtığı yaralardan ziyade fırsatçı temizlemeyi gösterebilir. T. rex.[184][185] Diğer tyrannosaurids yamyamlık da yapmış olabilir.[184]
Paleoekoloji

Tyrannosaurus olarak adlandırılan dönemde yaşadı Lancian faunal evre (Maastrihtiyen yaş) Geç Kretase'nin sonunda. Tyrannosaurus aralığında Kanada kuzeyde en azından New Mexico'nun güneyinde Laramidia.[5] Bu süre içinde Triceratops sıradağlarının kuzey kesimindeki en büyük otoburdu. Titanosaurian Sauropod Alamosaurus güney bölgesinde "hakim" oldu. Tyrannosaurus İç ve kıyı subtropikal ve yarı kurak ovalar dahil olmak üzere farklı ekosistemlerde kalıntılar keşfedilmiştir.

Birkaç önemli Tyrannosaurus kalıntılar bulundu Hell Creek Oluşumu. Maastrihtiyen döneminde bu bölge subtropikal, sıcak ve nemli bir iklime sahip. Flora çoğunlukla şunlardan oluşuyordu: anjiyospermler ama aynı zamanda şafak sekoya gibi ağaçları da (Metasequoia ) ve Araucaria. Tyrannosaurus bu ekosistemi paylaştı Ceratopsia'lılar Leptoceratops, Torosaurus, ve Triceratops, hadrosaurid Edmontosaurus annectens, Parksosaurid Thescelosaurus, Ankylosaurlar Ankylosaurus ve Denversaurus, pachycephalosaurs Pachycephalosaurus ve Sphaerotholus ve theropodlar Ornitomimus, Struthiomimus, Acheroraptor, Dakotaraptor, Pectinodon ve Anzu.[186]
İle başka bir oluşum Tyrannosaurus kalıntılar Lance Formasyonu Wyoming. Bu bir Bayou günümüz Körfez Kıyısı'na benzer bir ortam. Fauna Hell Creek'e çok benziyordu, ancak Struthiomimus akrabasını değiştirmek Ornitomimus. Küçük ceratopsian Leptoceratops bölgede de yaşıyordu.[187]
Güney bölgesinde Tyrannosaurus titanosaur ile birlikte yaşadı Alamosaurus, ceratopsianlar Torosaurus, Bravoceratops ve Ojoceratops bir türden oluşan hadrosaurlar Edmontosaurus, Kritosaurus ve olası bir tür Gryposaurus nodozor Glyptodontopelta, oviraptorid Ojoraptosaurus, theropodların olası türleri Troodon ve Richardoestesia, ve pterosaur Quetzalcoatlus.[188] Muhtemel geri çekilmenin ardından bölgede yarı kurak iç düzlüklerin hakim olduğu düşünülmektedir. Batı İç Denizyolu küresel deniz seviyeleri düşerken.[189]
Tyrannosaurus Meksika'da da yaşamış olabilir Lomas Coloradas Sonora'da oluşum. İskelet kanıtı eksik olsa da, fosil yatağından dökülen ve kırılan altı diş, diğer theropod cinsleriyle iyice karşılaştırıldı ve görünüşe göre Tyrannosaurus. Doğruysa, kanıt aralığı gösterir Tyrannosaurus muhtemelen daha önce düşünülenden daha kapsamlıydı.[190] Tiranozorların aslında Kretase döneminin sonundan önce Kuzey Amerika'ya göç eden Asyalı türler olması muhtemeldir.[191]
Kültürel önem

İlk kez 1905'te tanımlandığından beri, T. rex en çok tanınan dinozor türü haline geldi. popüler kültür. Genel halk tarafından tam bilimsel adıyla bilinen tek dinozordur (iki terimli isim ) ve bilimsel kısaltma T. rex ayrıca geniş bir kullanım alanına sahiptir.[47] Robert T. Bakker bunu not eder Dinozor Heresies "" gibi bir isim "T. rex"Sadece dil için karşı konulamaz."[35]
Notlar
- ^ Telaffuz edildi /tɪˌrænəˈsɔːrəs,taɪ-/Yunanca "zorba kertenkele" anlamına gelir Tyrannos (τύραννος), "zorba" ve Sauros (σαῦρος), "kertenkele"[1]
Referanslar
- ^ "Tyrannosaurus". Çevrimiçi Etimoloji Sözlüğü.
- ^ a b Breithaupt, B. H .; Southwell, E. H .; Matthews, N.A. (15 Ekim 2005). "100 yıllık bir kutlamada Tyrannosaurus rex: Manospondylus gigas, Ornitomimus grandis, ve Dynamosaurus imperiosus, İlk Keşifler Tyrannosaurus rex batıda". Programlı Özetler; 2005 Salt Lake City Yıllık Toplantısı. Amerika Jeoloji Topluluğu. 37 (7): 406. Arşivlenen orijinal 30 Mayıs 2012. Alındı 8 Ekim 2008.
- ^ Hatcher, J.B. (1907). "Ceratopsia". Amerika Birleşik Devletleri Jeolojik Araştırması Monografları. 49: 113–114.
- ^ a b Osborn, H.F. (1917). "İskelet uyarlamaları Ornitoestler, Struthiomimus, Tyrannosaurus". Amerikan Doğa Tarihi Müzesi Bülteni. 35 (43): 733–771. hdl:2246/1334.
- ^ a b c d e f g Larson, N.L. (2008). "Yüz yıllık Tyrannosaurus rex: the skeletons ". Larson, P .; Carpenter, K. (editörler). Tyrannosaurus rex, Zalim Kral. Bloomington, IN: Indiana University Press. pp.1 –55. ISBN 978-0-253-35087-9.
- ^ a b c d Osborn, H.F. (1905). "Tyrannosaurus ve diğer Kretase etobur dinozorları ". AMNH Bülteni. 21 (14): 259–265. hdl:2246/1464. Erişim tarihi: Ekim 6, 2008.
- ^ a b Dingus, L .; Norell, M. (3 Mayıs 2010). Barnum Brown: Keşfeden Adam Tyrannosaurus rex. California Üniversitesi Yayınları. pp.90, 124. ISBN 978-0-520-94552-4.
- ^ a b Osborn, H.F.; Brown, B. (1906). "Tyrannosaurus, Üst Kretase etobur dinozor ". AMNH Bülteni. 22 (16): 281–296. hdl:2246/1473.
- ^ Breithaupt, B. H .; Southwell, E. H .; Matthews, N.A. (2006). Lucas, S. G .; Sullivan, R. M. (editörler). "Dynamosaurus imperiosus ve en eski keşifler Tyrannosaurus rex Wyoming ve Batı'da " (PDF). New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 35: 258.
Orijinal iskeleti Dynamosaurus imperiosus (AMNH 5866 / BM R7995), diğerleriyle birlikte T. rex malzeme (AMNH 973, 5027 ve 5881 parçaları dahil), 1960 yılında British Museum of Natural History'ye (şimdi The Natural History Museum) satıldı. Bu materyal, bu dinozorun ilginç bir "yarı-monte" sergisinde kullanıldı. Londra. Şu anda materyal araştırma koleksiyonlarında bulunuyor.
- ^ McDonald, A. T .; Wolfe, D. G .; Dooley Jr, A.C. (2018). "New Mexico'daki Üst Kretase Menefee Formasyonundan yeni bir tyrannosaurid (Dinosauria: Theropoda)". PeerJ. 6: 6: e5749. doi:10.7717 / peerj.5749. PMC 6183510. PMID 30324024.
- ^ "Sue'nun kemiklerini hazırlamak". Saha Müzesi'nde Sue. Saha Müzesi. 2007. Alındı 24 Ekim 2014.
- ^ Erickson, G .; Makovicky, P. J .; Currie, P. J .; Norell, M .; Yerby, S .; Brochu, C.A. (26 Mayıs 2004). Tyrannosaurid dinozorların "Gigantizm ve yaşam öyküsü parametreleri". Doğa. 430 (7001): 772–775. Bibcode:2004Natur.430..772E. doi:10.1038 / nature02699. PMID 15306807. S2CID 4404887.
- ^ "Stan". Manchester Üniversitesi. 18 Eylül 2010. Arşivlenen orijinal 18 Eylül 2010.
- ^ Beşli, S. (2000). "Jurassic Farce". Tyrannosaurus Dava açmak. W. H. Freeman ve Şirketi, New York. pp.121 –122. ISBN 978-0-7167-4017-9.
- ^ "Bucky The Teenage T. Rex ile Tanışın". Indianapolis Çocuk Müzesi. 7 Temmuz 2014. Arşivlendi 27 Aralık 2014 tarihinde orjinalinden. Alındı 2 Aralık 2019.
- ^ "Dig, beş T. rex örneği çıkarıyor". BBC haberleri. 10 Ekim 2000. Alındı 13 Aralık, 2008.
- ^ a b Currie, P. J .; Hurum, J. H .; Sabath, K. (2003). "Tyrannosaurid dinozorlarda kafatası yapısı ve evrimi" (PDF). Acta Palaeontologica Polonica. 48 (2): 227–234. Alındı 8 Ekim 2008.
- ^ Black, Riley (28 Ekim 2015). "Küçük terör: Tartışmalı dinozor türleri sadece garip bir ara Tyrannosaurus". Smithsonian Dergisi. Alındı 10 Aralık 2018.
- ^ "Müze dünyanın en büyük T-rex kafatası". 2006. Arşivlenen orijinal 14 Nisan 2006. Alındı 7 Nisan 2006.
- ^ a b Gignac, P. M .; Erickson, G.M. (2017). "Aşırı osteofajinin arkasındaki biyomekanik Tyrannosaurus rex". Bilimsel Raporlar. 7 (1): 2012. Bibcode:2017NatSR ... 7.2012G. doi:10.1038 / s41598-017-02161-w. PMC 5435714. PMID 28515439.
- ^ Lockley, M. G .; Hunt, A.P. (1994). "Dev theropod dinozorunun izi Tyrannosaurus Kretase / Tersiyer sınırına yakın, kuzey New Mexico ". Ichnos. 3 (3): 213–218. doi:10.1080/10420949409386390.
- ^ "Hell Creek Formasyonundan (Üst Kretase) Gelen Olası Bir Tyrannosaurid Yolu, Montana, Amerika Birleşik Devletleri". Ulusal Tarih Haberleri Müzesi. 2007. Arşivlenen orijinal 14 Aralık 2007. Alındı 18 Aralık 2007.
- ^ a b Manning, P. L .; Ott, C .; Falkingham, P.L. (2009). "Hell Creek Formasyonundan (Geç Kretase) ilk tyrannosaurid izi, Montana, ABD". PALAIOS. 23 (10): 645–647. Bibcode:2008Palai..23..645M. doi:10.2110 / palo.2008.p08-030r. S2CID 129985735.
- ^ Smith, S. D .; Persons, W. S .; Xing, L. (2016). Glenrock, Lance Formation (Maastrichtian), Wyoming'de "Tyrannosaur" bir yol ". Kretase Araştırmaları. 61 (1): 1–4. doi:10.1016 / j.cretres.2015.12.020.
- ^ Perkins, S. (2016). "Muhtemelen bir T. rex". Paleontoloji. doi:10.1126 / science.aae0270.
- ^ Walton, T. (2016). "Jurassic Park'tan bildiklerinizi unutun: Hız için, T. rex vuruş velociraptors ". Bugün Amerika. Alındı 13 Mart, 2016.
- ^ Ruiz, J. (2017). "Glenrock'ta bir tyrannosaur trackway, Lance Formasyonu (Maastrichtian), Wyoming" (Smith et al., Kretase Araştırmaları, cilt 61, s. 1–4, 2016) ". Kretase Araştırmaları. 82: 81–82. doi:10.1016 / j.cretres.2017.05.033.
- ^ "Sue Adında Bir T. rex" (PDF). Alan Müzesi. Alan Müzesi. Arşivlenen orijinal (PDF) 18 Ağustos 2016. Alındı 4 Ocak 2019.
- ^ a b c d e Hutchinson, J. R .; Bates, K. T .; Molnar, J .; Allen, V .; Makovicky, P. J. (2011). "Tyrannosaurus rex'te Ekstremite ve Vücut Boyutlarının Hareket, Doğuştan Gelişim ve Büyüme Etkileri ile Hesaplamalı Bir Analizi". PLOS ONE. 6 (10): e26037. Bibcode:2011PLoSO ... 626037H. doi:10.1371 / journal.pone.0026037. PMC 3192160. PMID 22022500.
- ^ a b "Sue Bilgi Sayfası" (PDF). Saha Müzesi'nde Sue. Saha Doğa Tarihi Müzesi. Arşivlenen orijinal (PDF) 18 Ağustos 2016.
- ^ Therrien, F .; Henderson, D.M. (12 Mart 2007). "Theropodum sizinkinden daha büyük… ya da değil: theropodlarda kafatası uzunluğundan vücut boyutunu tahmin etmek". Omurgalı Paleontoloji Dergisi. 27 (1): 108–115. doi:10.1671 / 0272-4634 (2007) 27 [108: MTIBTY] 2.0.CO; 2. ISSN 0272-4634.
- ^ Persons, S. W .; Currie, P. J .; Erickson, G.M. (2020). "Daha Yaşlı ve Olağanüstü Büyüklükte Yetişkin Bir Numune Tyrannosaurus rex". Anatomik Kayıt. 303 (4): 656–672. doi:10.1002 / ar.24118. ISSN 1932-8486. PMID 30897281.
- ^ Lyle, A. (22 Mart 2019). "Paleontologlar en büyük Tyrannosaurus rex hiç keşfedildi ". Folio, Alberta Üniversitesi. Alındı 25 Mart, 2019.
- ^ Anderson, J. F .; Hall-Martin, A. J .; Russell, D. (1985). "Memelilerde, kuşlarda ve dinozorlarda uzun kemik çevresi ve ağırlığı". Zooloji Dergisi. 207 (1): 53–61. doi:10.1111 / j.1469-7998.1985.tb04915.x.
- ^ a b c Bakker, R. T. (1986). Dinozor Heresies. New York: Kensington Yayınları. s.241. ISBN 978-0-688-04287-5. OCLC 13699558.
- ^ Henderson, D.M. (1 Ocak 1999). "Soyu tükenmiş hayvanların kütlelerini ve kütle merkezlerini 3 boyutlu matematiksel dilimleme ile tahmin etme". Paleobiyoloji. 25 (1): 88–106.
- ^ a b c Erickson, G. M .; Makovicky, P. J .; Currie, P. J.; Norell, M. A .; Yerby, S. A .; Brochu, C.A. (2004). "Gigantizm ve tyrannosaurid dinozorların karşılaştırmalı yaşam öyküsü parametreleri". Doğa. 430 (7001): 772–775. Bibcode:2004Natur.430..772E. doi:10.1038 / nature02699. PMID 15306807. S2CID 4404887.
- ^ Farlow, J. O .; Smith, M. B .; Robinson, J.M. (1995). "Vücut kütlesi, kemik 'kuvvet göstergesi' ve cursorial potansiyeli Tyrannosaurus rex". Omurgalı Paleontoloji Dergisi. 15 (4): 713–725. doi:10.1080/02724634.1995.10011257. Arşivlenen orijinal 23 Ekim 2008.
- ^ Seebacher, F. (2001). "Dinozorların allometrik uzunluk-kütle ilişkilerini hesaplamak için yeni bir yöntem" (PDF). Omurgalı Paleontoloji Dergisi. 21 (1): 51–60. CiteSeerX 10.1.1.462.255. doi:10.1671 / 0272-4634 (2001) 021 [0051: ANMTCA] 2.0.CO; 2.
- ^ Christiansen, P .; Fariña, R.A. (2004). "Theropod dinozorlarında toplu tahmin". Tarihsel Biyoloji. 16 (2–4): 85–92. doi:10.1080/08912960412331284313. S2CID 84322349.
- ^ a b c Stevens, Kent A. (Haziran 2006). "Theropod dinozorlarında binoküler görüş". Omurgalı Paleontoloji Dergisi. 26 (2): 321–330. doi:10.1671 / 0272-4634 (2006) 26 [321: BVITD] 2.0.CO; 2.
- ^ a b c Jaffe, E. (1 Temmuz 2006). "Saur Gözlerine Bakış: T. rex vizyon doğanın en iyisiydi ". Bilim Haberleri. 170 (1): 3–4. doi:10.2307/4017288. JSTOR 4017288. Alındı 6 Ekim 2008.
- ^ Snively, E .; Henderson, D. M .; Phillips, D. S. (2006). "Tyrannosaurid dinozorların kaynaşmış ve tonozlu nazalleri: Kafa gücü ve beslenme mekaniği için çıkarımlar" (PDF). Acta Palaeontologica Polonica. 51 (3): 435–454. Alındı 8 Ekim 2008.
- ^ a b Meers, M.B. (Ağustos 2003). "Maksimum ısırma gücü ve av boyutu Tyrannosaurus rex ve beslenme davranışının çıkarımıyla ilişkileri ". Tarihsel Biyoloji. 16 (1): 1–12. doi:10.1080/0891296021000050755. S2CID 86782853.
- ^ Erickson, G. M .; Van Kirk, S. D .; Su, J .; Levenston, M.E .; Caler, W. E .; Carter, D.R. (1996). "İçin ısırma kuvveti tahmini Tyrannosaurus rex dişle işaretlenmiş kemiklerden ". Doğa. 382 (6593): 706–708. Bibcode:1996Natur.382..706E. doi:10.1038 / 382706a0. S2CID 4325859.
- ^ a b c Holtz, T.R. (1994). "Tyrannosauridae'nin Filogenetik Konumu: Theropod Sistematiği için Çıkarımlar". Paleontoloji Dergisi. 68 (5): 1100–1117. doi:10.1017 / S0022336000026706. JSTOR 1306180.
- ^ a b c d e f g h ben j k l m n Brochu, C.R. (2003). "Osteoloji Tyrannosaurus rex: neredeyse eksiksiz bir iskeletten bilgiler ve kafatasının yüksek çözünürlüklü bilgisayarlı tomografik analizi ". Omurgalı Paleontoloji Anıları Derneği. 7: 1–138. doi:10.2307/3889334. JSTOR 3889334.
- ^ Smith, J. B. (1 Aralık 2005). "Heterodonty Tyrannosaurus rex: theropod dişlerin taksonomik ve sistematik faydası için çıkarımlar ". Omurgalı Paleontoloji Dergisi. 25 (4): 865–887. doi:10.1671 / 0272-4634 (2005) 025 [0865: HITRIF] 2.0.CO; 2.
- ^ Douglas, K .; Genç, S. (1998). "Dinozor dedektifleri". Yeni Bilim Adamı. Alındı 16 Ekim 2008.
Bir paleontolog, T. rex'in devasa, kavisli dişlerini unutulmaz bir şekilde 'ölümcül muzlar' olarak tanımladı.
- ^ "Sue'nun hayati istatistikleri". Saha Müzesi'nde Sue. Saha Doğa Tarihi Müzesi. Arşivlenen orijinal 29 Eylül 2007. Alındı 15 Eylül 2007.
- ^ a b c Carr, T. D .; Varricchio, D. J .; Sedlmayr, J. C .; Roberts, E. M .; Moore, J.R. (30 Mart 2017). "Anagenez ve timsah benzeri yüz duyu sistemi kanıtı olan yeni bir tiranozor". Bilimsel Raporlar. 7: 44942. Bibcode:2017NatSR ... 744942C. doi:10.1038 / srep44942. ISSN 2045-2322. PMC 5372470. PMID 28358353.
- ^ Morhardt, Ashley (2009). Dinozor gülümsüyor: Sauropsidlerin premaksilla, maksilla ve diş kemiklerinin dokusu ve morfolojisi, dinozorlarda ekstra oral yapıların güvenilir bir şekilde çıkarılması için osteolojik bağıntılar sağlıyor mu? (Yüksek Lisans tezi). Western Illinois Üniversitesi.
- ^ a b Reisz, R. R .; Larson, D. (2016). "Diş anatomisi ve kafatası uzunluğunun diş boyutuna oranları, theropod dinozorların dudakları olduğu hipotezini desteklemektedir.". 4. Yıllık Toplantısı, 2016, Kanada Omurgalı Paleontolojisi Derneği. ISSN 2292-1389.
- ^ MONTEVIALE KROKODYLİLERİNİN (OLIGOCENE, İTALYA) MORFOLOJİSİ, TAKSONOMİSİ VE FİLOJENETİK İLİŞKİLERİ. 2018. s. 67. Alındı 9 Ekim 2020.
- ^ Lipkin, C .; Carpenter, K. (2008). "Tekrar ön ayaklarına bakarken Tyrannosaurus rex". Carpenter, K .; Larson, P. E. (ed.). Tyrannosaurus rex, Zalim Kral. Bloomington: Indiana University Press. pp.167 –190. ISBN 978-0-253-35087-9.
- ^ a b Holtz, T.R., Jr. (2004). "Tyrannosauroidea". İçinde Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.). Dinozorlar. Berkeley: California Üniversitesi Yayınları. pp.111 –136. ISBN 978-0-520-24209-8.
- ^ a b Paul, Gregory S. (1988). Dünyanın yırtıcı dinozorları: eksiksiz bir resimli rehber. New York: Simon ve Schuster. s.228. ISBN 978-0-671-61946-6. OCLC 18350868.
- ^ Maleev, E.A. (1955). F. J. Alcock tarafından çevrilmiştir. "(Rusça başlık)" [Moğolistan'ın devasa etobur dinozorları] (PDF). Doklady Akademii Nauk SSSR (Rusça). 104 (4): 634–637.
- ^ Rozhdestvensky, A. K. (1965). "Asya dinozorlarındaki büyüme değişiklikleri ve taksonomilerindeki bazı sorunlar". Paleontological Journal. 3: 95–109.
- ^ Carpenter, K. (1992). "Asya ve Kuzey Amerika'nın Tyrannosaurids (Dinosauria)". Mateer, N. J .; Chen, P.-j. (eds.). Deniz dışı Kretase jeolojisinin özellikleri. Pekin: Çin Okyanus Basını. s. 250–268. ISBN 978-7-5027-1463-5. OCLC 28260578.
- ^ Carr, T. D .; Williamson, T. E .; Schwimmer, D.R. (2005). "Alabama'nın Geç Kretase (Orta Kampaniyen) Demopolis Oluşumundan Yeni Bir Tür ve Tyrannosauroid Türü". Omurgalı Paleontoloji Dergisi. 25 (1): 119–143. doi:10.1671 / 0272-4634 (2005) 025 [0119: ANGASO] 2.0.CO; 2.
- ^ a b Loewen, M. A .; Irmis, R. B .; Sertich, J. J. W .; Currie, P. J.; Sampson, S. D. (2013). Evans, D. C (ed.). "Tyrant Dinozor Evrimi Geç Kretase Okyanuslarının Yükselişini ve Düşüşünü İzliyor". PLoS ONE. 8 (11): e79420. Bibcode:2013PLoSO ... 879420L. doi:10.1371 / journal.pone.0079420. PMC 3819173. PMID 24223179.
- ^ Vergano, D. (7 Kasım 2013). "Yeni Bulunan" Gore Kralı "Dinozor T. Rex'ten Önce Yönetti". National Geographic. Alındı 10 Kasım 2017.
- ^ Geggel, L. (29 Şubat 2016). "T. Rex Muhtemelen İstilacı Bir Türdü ". Canlı Bilim. Alındı 10 Kasım 2017.
- ^ Hone, D. W. E .; Wang, K .; Sullivan, C .; Zhao, X .; Chen, S .; Li, D .; Ji, S .; Ji, Q .; Xu, X. (2011). "Çin'in Üst Kretase'sinden yeni, büyük bir tyrannosaurine theropod". Kretase Araştırmaları. 32 (4): 495–503. doi:10.1016 / j.cretres.2011.03.005.
- ^ Olshevsky, G. (1995). "Tiranozorların kökeni ve evrimi". Kyoryugaku Saizensen [Dino Frontline]. 9–10: 92–119.
- ^ Carr, T. D .; Williamson, T. E. (2004). "Batı Kuzey Amerika'dan geç Maastrihtiyen Tyrannosauridae (Dinosauria: Theropoda) çeşitliliği". Linnean Society'nin Zooloji Dergisi. 142 (4): 479–523. doi:10.1111 / j.1096-3642.2004.00130.x.
- ^ Gilmore, C.W. (1946). "Montana'nın Lance Formasyonundan yeni bir etobur dinozor". Smithsonian Çeşitli Koleksiyonları. 106: 1–19.
- ^ Bakker, R.T .; Williams, M .; Currie, P.J. (1988). "Nanotyrannus, Montana'nın en son Kretase döneminden yeni bir cüce tyrannosaur cinsi. " Hunteria. 1: 1–30.
- ^ a b Carr, T.D. (1999). "Tyrannosauridae'de (Dinosauria, Coelurosauria) kraniyofasiyal ontogenez". Omurgalı Paleontoloji Dergisi. 19 (3): 497–520. doi:10.1080/02724634.1999.10011161.
- ^ a b Tsuihiji, T .; Watabe, M .; Tsogtbaatar, K .; Tsubamoto, T .; Barsbold, R .; Suzuki, S .; Lee, A.H .; Ridgely, R.C .; Kawahara, Y .; Witmer, L.M. (2011). "Kraniyal osteolojisi Tarbosaurus bataar Bugin Tsav, Moğolistan Nemegt Formasyonundan (Üst Kretase) ". Omurgalı Paleontoloji Dergisi. 31 (3): 497–517. doi:10.1080/02724634.2011.557116. S2CID 15369707.
- ^ Currie, P.J. (2003a). "Kanada, Alberta'nın Geç Kretase'sinden tyrannosaurid dinozorların kraniyal anatomisi". Acta Palaeontologica Polonica. 48: 191–226.
- ^ Currie, Henderson, Horner ve Williams (2005). "Tiranozor dişleri, diş pozisyonları ve taksonomik durumu hakkında Nanotyrannus lancensis"Tyrannosauridae'nin kökeni, sistematiği ve paleobiyolojisi" nde, Burpee Doğa Tarihi Müzesi ve Northern Illinois Üniversitesi tarafından ortaklaşa düzenlenen bir sempozyum.
- ^ Henderson (2005). "Nano No More: Cüce zorbanın ölümü." Burpee Doğa Tarihi Müzesi ve Northern Illinois Üniversitesi'nin ortaklaşa ev sahipliği yaptığı "Tyrannosauridae'nin kökeni, sistematiği ve paleobiyolojisi" adlı sempozyumda.
- ^ Larson (2005). "Bir dava Nanotyrannus. "Burpee Doğa Tarihi Müzesi ve Northern Illinois Üniversitesi'nin ortaklaşa ev sahipliği yaptığı" Tyrannosauridae'nin kökeni, sistematiği ve paleobiyolojisi "adlı sempozyumda.
- ^ Larson P (2013), "Nanotyrannus Lancensis'in geçerliliği (Theropoda, Lancian - Kuzey Amerika'nın Yukarı Maastrihtiyarı", Omurgalı Paleontoloji Derneği: 73. yıllık toplantısı, Programlı Özetler, s. 159.
- ^ D. Carr, Thomas (15 Eylül 2013). ""NANOTİRANNUS GERÇEKTEN GERÇEK DEĞİLDİR"". Tyrannosauroidea Merkez. Alındı 28 Mayıs 2019.
- ^ Persons, W. S .; Currie, P.J. (2016). "Etobur dinozorlarda cursorial uzuv oranlarını puanlama yaklaşımı ve allometriyi açıklama girişimi". Bilimsel Raporlar. 6. 19828. Bibcode:2016NatSR ... 619828P. doi:10.1038 / srep19828. PMC 4728391. PMID 26813782.
- ^ "Arka uzuv oranları Nanotyrannus'un geçerliliğini desteklemiyor". mambobob-raptorsnest.blogspot.com.
- ^ Schmerge, Joshua D .; Rothschild, Bruce M. (2016). "Theropod dinozorlarının diş oluğunun dağılımı: Theropod soyoluşuna ilişkin çıkarımlar ve Nanotyrannus Bakker ve diğerleri, 1988 cinsinin geçerliliği". Kretase Araştırmaları. 61: 26–33. doi:10.1016 / J.CRETRES.2015.12.016. S2CID 85736236.
- ^ Brusatte, Stephen L .; Carr, Thomas D .; Williamson, Thomas E .; Holtz, Thomas R .; Hone, David W.E .; Williams, Scott A. (2016). "Dentary oluk morfolojisi, 'Nanotyrannus'u, geçerli bir tyrannosauroid dinozor taksonu olarak ayırt etmez. Yorum:" Theropod dinozorların diş oluğunun dağılımı: Theropod soyoluşu ve Nanotyrannus cinsinin geçerliliği için çıkarımlar Bakker ve diğerleri, 1988"" (PDF). Kretase Araştırmaları. 65: 232–237. doi:10.1016 / J.CRETRES.2016.02.007. S2CID 56090258.
- ^ Woodward, Holly N .; Tremaine, Katie; Williams, Scott A .; Zanno, Lindsay E .; Horner, John R .; Myhrvold Nathan (2020). "Tyrannosaurus rex'i büyütmek: Osteohistoloji cüce" Nanotyrannus "u çürütür ve gençlikte ontogenetik niş bölünmesini destekler Tyrannosaurus". Bilim Gelişmeleri. 6 (1): eaax6250. Bibcode:2020SciA .... 6.6250W. doi:10.1126 / sciadv.aax6250. ISSN 2375-2548. PMC 6938697. PMID 31911944.
- ^ Carr, T.D. (2020). "Yüksek çözünürlüklü büyüme serisi Tyrannosaurus rex birden fazla kanıttan elde edildi. PeerJ. 8: e9192. doi:10.7717 / peerj.9192.
- ^ Carr, T.D. (5 Haziran 2020). "Tyrannosaurus rex'in yüksek çözünürlüklü büyüme serisi, birden fazla kanıt dizisinden elde edilmiştir - Yazar Dr. Thomas D. Carr yeni çalışmasını tartışıyor". PeerJblog. Alındı 10 Haziran, 2020.
- ^ a b Horner, J. R .; Padian, K. (2004). "Tyrannosaurus rex'in yaş ve büyüme dinamikleri". Bildiriler: Biyolojik Bilimler. 271 (1551): 1875–80. doi:10.1098 / rspb.2004.2829. PMC 1691809. PMID 15347508. Alındı 5 Ekim 2008.
- ^ a b Schweitzer, M. H .; Wittmeyer, J. L .; Horner, J.R. (2005). "Ratitlerde cinsiyete özgü üreme dokusu ve Tyrannosaurus rex". Bilim. 308 (5727): 1456–60. Bibcode:2005Sci ... 308.1456S. doi:10.1126 / science.1112158. PMID 15933198. S2CID 30264554.
- ^ Lee, A. H .; Werning, S. (2008). "Büyüyen dinozorlardaki cinsel olgunluk sürüngen büyüme modellerine uymuyor". Ulusal Bilimler Akademisi Bildiriler Kitabı. 105 (2): 582–587. Bibcode:2008PNAS..105..582L. doi:10.1073 / pnas.0708903105. PMC 2206579. PMID 18195356.
- ^ Schweitzer, M. H .; Zheng, W .; Zanno, L .; Werning, S .; Sugiyama, T. (2016). "Kimya, Tyrannosaurus rex'te cinsiyete özgü üreme dokusunun tanımlanmasını destekler". Bilimsel Raporlar. 6 (23099): 23099. Bibcode:2016NatSR ... 623099S. doi:10.1038 / srep23099. PMC 4791554. PMID 26975806.
- ^ a b Erickson, G. M .; Currie, P. J .; Inouye, B. D .; Winn, A. A. (2006). "Tyrannosaur yaşam tabloları: nonavian dinozor popülasyon biyolojisi örneği". Bilim. 313 (5784): 213–7. Bibcode:2006Sci ... 313..213E. doi:10.1126 / science.1125721. PMID 16840697. S2CID 34191607.
- ^ Woodward, Holly N; Tremaine, Katie; Williams, Scott A; Zanno, Lindsay E; Horner, John R; Myhrvold, Nathan (1 Ocak 2020). "Tyrannosaurus rex'in büyümesi: Osteohistoloji cüce" Nanotyrannus "u çürütür ve genç Tyrannosaurus'ta ontogenetik niş bölünmesini destekler". Bilim Gelişmeleri. 6 (1): eaax6250. Bibcode:2020SciA .... 6.6250W. doi:10.1126 / sciadv.aax6250. PMC 6938697. PMID 31911944.
- ^ Greshko, Michael (1 Ocak 2020). "Bu zarif, yırtıcı dinozorlar gerçekten genç T. rex.". National Geographic. Alındı 2 Ocak, 2020.
- ^ a b Holtz, T. R., Jr (19 Mart 2013) [8 Mart 2013'te yapılan konuşma]. Tyrannosaurus rex'in Hayatı ve Zamanları, Dr. Thomas Holtz ile (Ders). Seattle, WA: Burke Doğa Tarihi ve Kültür Müzesi. Alındı 12 Ekim 2013.
- ^ Paul, G. S. (2008). "Bölüm 18: Kretase Kuzey Amerika ve Asya'nın Devasa Tyrannosaurid Süper Yırtıcılarının Olağanüstü Yaşam Tarzı ve Alışkanlıkları". Larson, P. L .; Carpenter, K. (editörler). Tyrannosaurus, Zalim Kral. Indiana University Press. s. 307–345. ISBN 978-0-253-35087-9. Alındı 14 Eylül 2013.
- ^ a b Bell, P. R .; Campione, N. E .; Kişiler IV, W. S .; Currie, P. J .; Larson, P. L .; Tanke, D. H .; Bakker, R.T. (2017). "Tyrannosauroid bütünlüğü, devlik ve tüy evriminin çelişkili modellerini ortaya çıkarıyor". Biyoloji Mektupları. 13 (6): 20170092. doi:10.1098 / rsbl.2017.0092. PMC 5493735. PMID 28592520.
- ^ Farago, J. (7 Mart 2019). "T. Rex'i Görmediğin Gibi: Tüylü". New York Times. Alındı 7 Mart, 2019.
- ^ a b Xing, X .; Norell, M. A .; Kuang, X .; Wang, X .; Zhao, Q .; Jia, C. (7 Ekim 2004). "Çin'den gelen bazal tyrannosauroidler ve tyrannosauroidlerdeki protofeathers için kanıtlar". Doğa. 431 (7009): 680–684. Bibcode:2004Natur.431..680X. doi:10.1038 / nature02855. PMID 15470426. S2CID 4381777.
- ^ Xing, X .; Wang, K .; Zhang; Ma, Q .; Xing, L .; Sullivan, C .; Hu, D .; Cheng, S .; Wang, S. (5 Nisan 2012). "Çin'in Alt Kretase'sinden dev bir tüylü dinozor" (PDF). Doğa. 484 (7392): 92–95. Bibcode:2012Natur.484 ... 92X. doi:10.1038 / nature10906. PMID 22481363. S2CID 29689629. Arşivlenen orijinal (PDF) 17 Nisan 2012.
- ^ Naish, D. "Büyük Yırtıcı Bir Dinozorun Hassas Yüzü". Tetrapod Zooloji. Scientific American Blog Ağı. Alındı 5 Aralık 2018.
- ^ "MONTEVIALE KROKODYLİLERİNİN (OLIGOCENE, İTALYA) MORFOLOJİSİ, TAKSONOMİSİ VE FİLOJENETİK İLİŞKİLERİ. S. 67". Yıllık Omurgalı Paleontolojisi ve Karşılaştırmalı Anatomi Sempozyumu. Alındı 9 Ekim 2020.
- ^ Milinkovitch, Michel; Manukyan, Liana; Debry, Adrien; Di-Poi, Nicolas; Martin, Samuel; Singh, Daljit; Lambert, Dominique; Zwicker, Matthias (4 Ocak 2013). "Timsah Baş Ölçekleri Gelişimsel Birimler Değildir, Fiziksel Çatlamadan Ortaya Çıkar". Bilim. 339 (6115): 78–81. Bibcode:2013Sci ... 339 ... 78M. doi:10.1126 / science.1226265. PMID 23196908. S2CID 6859452. Alındı 9 Ekim 2020.
- ^ Carpenter, K. (1992). "Varyasyon Tyrannosaurus rex". Carpenter, K .; Currie, P. J. (editörler). Dinozor Sistematiği: Yaklaşımlar ve Perspektifler. Cambridge: Cambridge University Press. s. 141–145. ISBN 978-0-521-43810-0.
- ^ Larson, P.L. (1994). "Tyrannosaurus sex ". Rosenberg, G. D .; Wolberg, D. L. (editörler). Dino Festivali. 7. Paleontoloji Derneği Özel Yayınları. s. 139–155.
- ^ Erickson, G. M .; Kristopher, L. A .; Larson, P. (2005). "Androjen rex - timsahların ve kuş olmayan dinozorların cinsiyetini belirlemek için köşeli çift ayraçların faydası". Zooloji (Jena, Almanya). 108 (4): 277–86. doi:10.1016 / j.zool.2005.08.001. PMID 16351976.
- ^ Schweitzer, M. H .; Elsey, R. M .; Dacke, C. G .; Horner, J. R .; Lamm, E.T. (2007). "Yumurtlayan timsah yap (Timsah mississippiensis) archosaurs medüller kemik oluşturur mu? ". Kemik. 40 (4): 1152–8. doi:10.1016 / j.bone.2006.10.029. PMID 17223615.
- ^ Leidy, J. (1865). "Amerika Birleşik Devletleri'nin Kretase oluşumlarının soyu tükenmiş sürüngenleri üzerine anı". Smithsonian'ın Bilgiye Katkıları. 14: 1–135.
- ^ "Tyrannosaurus". Amerikan Doğa Tarihi Müzesi. Arşivlenen orijinal 8 Aralık 2008. Alındı 16 Ekim 2008.
- ^ a b c Newman, B.H. (1970). "Et yiyen duruş ve yürüyüş Tyrannosaurus". Linnean Society Biyolojik Dergisi. 2 (2): 119–123. doi:10.1111 / j.1095-8312.1970.tb01707.x.
- ^ "Sürüngenler Çağı Duvar". Yale Üniversitesi. 2008. Alındı 16 Ekim 2008.
- ^ Ross, R. M .; Duggan-Haas, D .; Allmon, W. D. (2013). "Duruş Tyrannosaurus rex: Öğrenci Görüşleri Neden Bilimin Gerisinde Kalmaktadır? ". Jeoloji Eğitimi Dergisi. 61 (1): 145–160. Bibcode:2013JGeEd..61..145R. doi:10.5408/11-259.1. S2CID 162343784.
- ^ "Tyrannosaurus Rex: Artık bir tripod değil ". American Association for the Advancement of Science.
- ^ Stevens, Kent A. (2011). "Tyrannosaurus rex -" Rex, otur"". ix.cs.uoregon.edu. Alındı 26 Temmuz 2020.
- ^ "T. rex düştüyse, küçücük kolları ve alçak ağırlık merkezi göz önüne alındığında nasıl ayağa kalktı?". Bilimsel amerikalı. Alındı 26 Temmuz 2020.
- ^ Lambe, L.M. (1914). "Alberta'daki Belly River Formasyonundan yeni bir etobur dinozor cinsi ve türü hakkında, Stephanosaurus marginatus aynı ufuktan ". Ottawa Doğa Uzmanı. 27: 129–135.
- ^ a b c Horner, J. R.; Lessem, D. (1993). Tam T. rex. New York City: Simon ve Schuster. ISBN 978-0-671-74185-3.
- ^ "T. Rex'e Yeni Bir Bakış | Smithsonian Ulusal Doğa Tarihi Müzesi". 13 Nisan 2020. Arşivlenen orijinal 13 Nisan 2020. Alındı 13 Nisan 2020.
- ^ Carpenter, K.; Smith, M. (2001). "Ön ayak Osteolojisi ve Biyomekaniği Tyrannosaurus rex". İçinde Tanke, D. H.; Carpenter, K. (eds.). Mesozoik omurgalı yaşamı. Bloomington: Indiana University Press. pp.90–116. ISBN 978-0-253-33907-2.
- ^ Pickrell, J. (2 Kasım 2017). "T. Rex'in Küçük Kolları Kötü Silahlar Olabilir". National Geographic. Alındı 10 Aralık 2018.
- ^ Bakker, R. T. (1968). "Dinozorların üstünlüğü" (PDF). Keşif. 3 (2): 11–12. Arşivlenen orijinal (PDF) 9 Eylül 2006 tarihinde. Alındı 7 Ekim 2008.
- ^ Bakker, R. T. (1972). "Dinozorlarda endoterminin anatomik ve ekolojik kanıtı" (PDF). Doğa. 238 (5359): 81–85. Bibcode:1972Natur.238 ... 81B. doi:10.1038 / 238081a0. S2CID 4176132. Arşivlenen orijinal (PDF) 9 Eylül 2006 tarihinde. Alındı 7 Ekim 2008.
- ^ Barrick, R. E .; Duşlar, W. J. (1994). "Termofizyolojisi Tyrannosaurus rex: Oksijen İzotoplarından Kanıt ". Bilim. 265 (5169): 222–224. Bibcode:1994Sci ... 265..222B. doi:10.1126 / science.265.5169.222. PMID 17750663. S2CID 39392327.
- ^ Trueman, C .; Chenery, C .; Eberth, D. A .; Spiro, B. (2003). "Dinozorların kemiklerinin oksijen izotop bileşimi üzerindeki diyajenetik etkiler ve karasal ve deniz çökeltilerinden elde edilen diğer omurgalılar". Jeoloji Topluluğu Dergisi. 160 (6): 895–901. Bibcode:2003JGSoc.160..895T. doi:10.1144/0016-764903-019. S2CID 130658189.
- ^ Barrick, R. E .; Duşlar, W. J. (1999). "Termofizyoloji ve biyolojisi Giganotosaurus: ile karşılaştırıldığında Tyrannosaurus". Paleontoloji Electronica. 2 (2). Alındı 7 Ekim 2008.
- ^ Barrick, R. E .; Stoskopf, M. K .; Duşlar, W. J. (1999). "Dinozor kemiklerindeki oksijen izotopları". Farlow, J. O .; Brett-Surman, M. K. (editörler). Komple Dinozor. Bloomington: Indiana University Press. sayfa 474–490. ISBN 978-0-253-21313-6.
- ^ Paladino, F. V .; Spotila, J. R .; Dodson, P. (1999). "Devler için bir plan: büyük dinozorların fizyolojisinin modellenmesi". Farlow, J. O .; Brett-Surman, M. K. (editörler). Komple Dinozor. Bloomington: Indiana University Press. sayfa 491–504. ISBN 978-0-253-21313-6.
- ^ Chinsamy, A .; Hillenius, W. J. (2004). "Nonavian dinozorların fizyolojisi". Weishampel, D. B .; Dodson, P .; Osmólska, H. (editörler). Dinozorlar. Berkeley: California Üniversitesi Yayınları. pp.643 –659. ISBN 978-0-520-24209-8.
- ^ Seymour, R. S. (5 Temmuz 2013). "Memelilere Karşı Büyük Timsahlarda Maksimal Aerobik ve Anaerobik Güç Üretimi: Dinozor Gigantothermy için Çıkarımlar". PLOS ONE. 8 (7): e69361. Bibcode:2013PLoSO ... 869361S. doi:10.1371 / journal.pone.0069361. ISSN 1932-6203. PMC 3702618. PMID 23861968.
- ^ "T. Rex kafasında bir klima vardı, çalışma gösteriyor". Günlük Bilim. Alındı 6 Eylül 2019.
- ^ a b Alanlar, H. (2006). "Dinozor Şok Edici". Smithsonian Dergisi. Alındı 2 Ekim 2008.
- ^ Schweitzer, M.H.; Wittmeyer, J. L .; Horner, J. R .; Toporski, J. K. (2005). "Yumuşak doku damarları ve hücresel koruma Tyrannosaurus rex". Bilim. 307 (5717): 1952–5. Bibcode:2005Sci ... 307.1952S. doi:10.1126 / science.1108397. PMID 15790853. S2CID 30456613.
- ^ Rincon, P. (12 Nisan 2007). "Protein, T. rex'i tavuklara bağlar". BBC haberleri. Alındı 2 Ekim 2008.
- ^ Vergano, D. (13 Nisan 2007). "Dünün T. Rex bugünün tavuğu". Bugün Amerika. Alındı 8 Ekim 2008.
- ^ Kaye, T. G .; Gaugler, G .; Sawlowicz, Z. (2008). Stepanova, A. (ed.). "Bakteriyel Biyofilmler Olarak Yorumlanan Dinosaurian Yumuşak Dokular". PLOS ONE. 3 (7): e2808. Bibcode:2008PLoSO ... 3.2808K. doi:10.1371 / journal.pone.0002808. PMC 2483347. PMID 18665236.
- ^ "Dinozor Yumuşak Dokularının Hala Hayatta Kaldığı Düşüncesine Yeni Araştırma Zorlukları" (Basın bülteni). Newswise. 24 Temmuz 2008. Alındı 8 Ekim 2008.
- ^ "Araştırmacılar Tartışması: Dinozor Dokusu veya Bakteriyel Balçık Korunmuş mu?" (Basın bülteni). Keşfedin. 30 Temmuz 2008. Alındı 4 Eylül 2008.
- ^ San Antonio, J. D .; Schweitzer, M. H .; Jensen, S. T .; Kalluri, R .; Buckley, M .; Orgel, J.P.R.O. (2011). Van Veen, H.W. (ed.). "Dinozor Peptitleri Proteinlerin Hayatta Kalma Mekanizmalarını Öneriyor". PLOS ONE. 6 (6): e20381. Bibcode:2011PLoSO ... 620381S. doi:10.1371 / journal.pone.0020381. PMC 3110760. PMID 21687667.
- ^ Peterson, J. E .; Lenczewski, M.E .; Scherer, R.P. (12 Ekim 2010). "Mikrobiyal Biyofilmlerin Fosil ve Ayakta Olmayan Archosaurlarda Birincil Yumuşak Dokuların Korunmasına Etkisi". PLOS ONE. 5 (10): e13334. Bibcode:2010PLoSO ... 513334P. doi:10.1371 / journal.pone.0013334. PMC 2953520. PMID 20967227.
Korunan organik kalıntıların mikrobiyal biyofilm olarak yorumlanması pek olası değildir
- ^ a b c d Hutchinson, J. R .; Garcia, M. (2002). "Tyrannosaurus hızlı koşucu değildi ". Doğa. 415 (6875): 1018–21. Bibcode:2002Natur.415.1018H. doi:10.1038 / 4151018a. PMID 11875567. S2CID 4389633.
- ^ a b c Hutchinson, J.R. (2004). "İki Ayaklı Koşu Yeteneğinin Biyomekanik Modellemesi ve Duyarlılık Analizi. II. Soyu Tükenmiş Taksonlar" (PDF). Morfoloji Dergisi. 262 (1): 441–461. doi:10.1002 / jmor.10240. PMID 15352202. S2CID 15677774. Arşivlenen orijinal (PDF) 31 Ekim 2008.
- ^ a b Holtz, T.R. (1 Mayıs 1996). "Coelurosauria'nın (Dinosauria; Theropoda) filogenetik taksonomisi". Paleontoloji Dergisi. 70 (3): 536–538. doi:10.1017 / S0022336000038506. Alındı 3 Ekim 2008.
- ^ Benton, M. (2014). Omurgalı Paleontoloji (4. baskı). s. 193. ISBN 978-1118407554.
- ^ Dececchi, T. Alexander; Mloszewska, Aleksandra M .; Jr, Thomas R. Holtz; Habib, Michael B .; Larsson, Hans C.E. (13 Mayıs 2020). "Hızlı ve tutumlu: Farklı lokomotori stratejileri, theropod dinozorlarda uzuvların uzamasını sağlar". PLOS ONE. 15 (5): e0223698. Bibcode:2020PLoSO..1523698D. doi:10.1371 / journal.pone.0223698. PMC 7220109. PMID 32401793.
- ^ "T. rex, düşük hızlarda süper verimli bir yürüyüşçüydü". EurekAlert!.
- ^ "Neden Tyrannosaurus yavaş bir koşucuydu ve neden en büyüğü her zaman en hızlı değil ". Günlük Bilim. 17 Temmuz 2017. Alındı 10 Kasım 2017.
- ^ Hirt, M.R .; Jetz, W .; Rall, B. C .; Brose, U. (2017). "Genel bir ölçeklendirme yasası, en büyük hayvanların neden en hızlı olmadığını ortaya koyuyor". Doğa Ekolojisi ve Evrimi. 1 (8): 1116–1122. doi:10.1038 / s41559-017-0241-4. PMID 29046579. S2CID 425473.
- ^ Satıcılar, W. I .; Pond, S. B .; Brassey, C. A .; Manning, P. L .; Bates, K. T. (18 Temmuz 2017). "Koşma yeteneklerini araştırmak Tyrannosaurus rex stres kısıtlamalı çok gövdeli dinamik analiz kullanma ". PeerJ. 5: e3420. doi:10.7717 / peerj.3420. ISSN 2167-8359. PMC 5518979. PMID 28740745.
- ^ Cotton, J. R .; Hartman, S. A .; Currie, P. J .; Witmer, L. M .; Russell, A. P .; Holtz, T.R. Jr; Burns, M. E .; Sailing, L. A .; Mallison, H .; Henderson, D. M .; O’Brien, H .; Snively, E. (21 Şubat 2019). "Daha düşük dönme ataleti ve daha büyük bacak kasları, diğer büyük theropodlara göre tyrannosauridlerde daha hızlı dönüşler olduğunu gösterir". PeerJ. 7: e6432. doi:10.7717 / peerj.6432. PMC 6387760. PMID 30809441.
- ^ a b Witmer, L. M .; Ridgely, R.C. (2009). "Tyrannosaurların (Dinosauria, Theropoda) Beyin, Braincase ve Kulak Bölgelerine İlişkin Yeni Bilgiler, Duyusal Organizasyon ve Davranış için Çıkarımlar". Anatomik Kayıt. 292 (9): 1266–1296. doi:10.1002 / ar.20983. PMID 19711459. S2CID 17978731.
- ^ Stevens, K.A. (1 Nisan 2011). "Theropod Dinozorlarının Binoküler Vizyonu". Alındı 19 Temmuz 2013.
- ^ a b "T. Rex beyin çalışması rafine bir 'burnu ortaya çıkarır'". Calgary Herald. 28 Ekim 2008. Alındı 29 Ekim 2008.
- ^ Hurlburt, G. S .; Ridgely, R. C .; Witmer, L. M. (5 Temmuz 2013). "Tyrannosaurid dinozorlarda göreceli beyin ve serebrum boyutu: mevcut timsahlarda beyin-endokast nicel ilişkilerini kullanan bir analiz". Parrish, M. J .; Molnar, R.E .; Currie, P. J .; Koppelhus, E. B. (editörler). Tyrannosaurid Paleobiyolojisi. Indiana University Press. s. 134–154. ISBN 978-0-253-00947-0. Alındı 20 Ekim 2013.
- ^ Brusatten, Steve (2018). Dinozorların Yükselişi ve Düşüşü. New York, New York: HarperCollins Yayıncıları. s. 219. ISBN 9780062490438.
- ^ a b "Dino Çeteleri". Discovery Channel. 22 Haziran 2011. Arşivlendi orijinal 19 Ocak 2012. Alındı 19 Ocak 2012.
- ^ Collins, N. (22 Haziran 2011). "Tyrannosaurus Rex 'paketler halinde avlandı'". Telgraf. Alındı 23 Mart, 2014.
- ^ Wallis, P. (11 Haziran 2012). "Op-Ed: T. Rex paket avcıları mı? Korkunç, ancak gerçek olması muhtemel". Digitaljournal.com. Alındı 23 Aralık 2015.
- ^ a b Switek, B. (25 Temmuz 2011). "Bir grup kemik, kana susamış tiranozorlar çetesi yapmaz". Gardiyan. Alındı Haziran 21, 2015.
- ^ Sample, I. (23 Temmuz 2014). "Araştırmacılar, tiranozorların paketler halinde avlandığına dair ilk işareti buldu". Gardiyan. Alındı 28 Temmuz 2014.
- ^ McCrea, R.T. (2014). "Bir 'Tyrannosaurs Dehşeti': Tyrannosauridlerin İlk Yolları ve Tyrannosauridae'deki Çirkinlik ve Patolojinin Kanıtı". PLOS ONE. 9 (7): e103613. Bibcode:2014PLoSO ... 9j3613M. doi:10.1371 / journal.pone.0103613. PMC 4108409. PMID 25054328.
- ^ Peterson, J. E .; Henderson, M. D .; Sherer, R. P .; Vittore, C. P. (2009). "Juvenil Tyrannosaurid'i Yüz Isırma ve Davranışsal Etkileri". PALAIOS. 24 (11): 780–784. Bibcode:2009Palai..24..780P. doi:10.2110 / palo.2009.p09-056r. S2CID 85602478. Arşivlenen orijinal 11 Ağustos 2013.
- ^ Parisi, T. (2 Kasım 2009). "T. rex NIU bilim adamlarının korkunç gençleri: Genç tiranozorlar birbirlerine karşı ciddi bir savaş yaptılar". Northern Illinois Üniversitesi. Arşivlenen orijinal 14 Ağustos 2013. Alındı 10 Ağustos 2013.
- ^ "Öldürme Zamanı T. rex Çöpçü "Tartışması"". National Geographic. 16 Temmuz 2013. Arşivlendi orijinal 12 Temmuz 2018.
- ^ Black, Riley (13 Nisan 2012). "Ne zaman Tyrannosaurus Chomped Sauropod'lar ". Smithsonian Dergisi. Smithsonian Media. Alındı 24 Ağustos 2013.
- ^ Siyah, Riley (2012). " Tyrannosaurus rexTehlikeli ve Ölümcül Isırık ". Smithsonian Dergisi. Smithsonian Enstitüsü.
- ^ Bates, K. T .; Falkingham, P.L. (29 Şubat 2012). "Şuradaki maksimum ısırık performansını tahmin etme Tyrannosaurus rex çoklu cisim dinamiklerini kullanma ". Biyolojik Mektuplar. 8 (4): 660–664. doi:10.1098 / rsbl.2012.0056. PMC 3391458. PMID 22378742.
- ^ Scully, C. (2002). Oxford Uygulamalı Diş Bilimleri El Kitabı. Oxford University Press. s.156. ISBN 978-0-19-851096-3.
- ^ Lautenschlager, Stephan (4 Kasım 2015). "Theropod dinozorlarında kafatası kas-iskelet sistemi kısıtlamalarının tahmin edilmesi". Royal Society Açık Bilim. 2 (11): 150495. Bibcode:2015RSOS .... 250495L. doi:10.1098 / rsos.150495. PMC 4680622. PMID 26716007.
- ^ "Sizi birlikte yemeniz daha iyi mi? Dinozorların çeneleri beslenmeyi nasıl etkiledi?". Günlük Bilim. 3 Kasım 2015. Arşivlendi orijinal 4 Kasım 2015. Alındı 14 Eylül 2018.
- ^ Lambe, L.B. (1917). "Kretase Thopodous dinozoru Gorgosaurus". Kanada Jeolojik Araştırmasının Anıları. 100: 1–84. doi:10.4095/101672.
- ^ Farlow, J. O .; Holtz (2002). "Dinozorlardaki yırtıcı hayvanların fosil kayıtları" (PDF). Kowalewski, M .; Kelley, P.H. (editörler). Predasyonun Fosil Kayıtları. Paleontoloji Derneği Makaleleri. 8. T. R. Jr. s. 251–266. Arşivlenen orijinal (PDF) 31 Ekim 2008.
- ^ a b Horner, J.R. (1994). "Biftek bıçakları, boncuk gözlü gözler ve minik kollar (bir portresi Tyrannosaurus çöpçü olarak) ". Paleontoloji Derneği Özel Yayını. 7: 157–164. doi:10.1017 / S2475262200009497.
- ^ a b Amos, J. (31 Temmuz 2003). "Bilim / Doğa: T. rex yargılanıyor ". BBC haberleri. Alındı 23 Aralık 2015.
- ^ Chin, K .; Tokaryk, T. T .; Erickson, G. M .; Calk, L. C. (18 Haziran 1998). "Büyük boyutlu bir theropod koprolit". Doğa. 393 (6686): 680–682. Bibcode:1998Natur.393..680C. doi:10.1038/31461. S2CID 4343329. Özeti Monastersky, R. (20 Haziran 1998). "Kıçından kepçe almak T. rex". Bilim Haberleri. 153 (25): 391. doi:10.2307/4010364. JSTOR 4010364. Arşivlenen orijinal 11 Mayıs 2013.
- ^ Carpenter, K. (1998). "Theropod dinozorların yırtıcı davranışlarının kanıtı". Gaia. 15: 135–144. Arşivlenen orijinal 17 Kasım 2007. Alındı 5 Aralık 2007.
- ^ Eberth, David A .; Evans, David C., editörler. (2015). "Kanada, Alberta'dan Geç Kretase Hadrosauridae'de Paleopatoloji". Hadrosaurlar. Indiana University Press. s. 540–571. ISBN 978-0-253-01390-3. ön baskı
- ^ Siviero, ART V .; Marka, Leonard R .; Cooper, Allen M .; Hayes, William K .; Rega, Elizabeth; Siviero, Bethania C.T. (2020). "Monodominant Kemikli, Lance Formasyonundan (Maastrichtian), Wyoming ABD'den Edmontosaurus Annectens'te Kuyruk İçi Hareketliliğin Etkileri Olan İskelet Travması". PALAIOS. 35 (4): 201–214. Bibcode:2020Palai..35..201S. doi:10.2110 / palo.2019.079. S2CID 218503493.
- ^ Happ, J .; Carpenter, K. (2008). "Bire bir karşılaşmada yırtıcı-av davranışının analizi Tyrannosaurus rex ve Triceratops". Carpenter, K .; Larson, P. E. (ed.). Tyrannosaurus rex, Tyrant King (Geçmişin Hayatı). Bloomington: Indiana University Press. s. 355–368. ISBN 978-0-253-35087-9.
- ^ Dodson, P. (1996). Boynuzlu Dinozorlar. Princeton University Press. s.19.
- ^ Tanke, D.H.; Currie, P. J. (1998). "Theropod dinozorlarda kafa ısırma davranışı: paleopatolojik kanıtlar" (PDF). Gaia (15): 167–184. ISSN 0871-5424. Arşivlenen orijinal (PDF) 27 Şubat 2008.
- ^ Peterson, J. E .; Daus, K.N. (4 Mart 2019). "Yavruya atfedilebilen beslenme izleri Tyrannosaurus rex ontogenetik beslenme eğilimleri hakkında fikir verir ". PeerJ. 7: e6573. doi:10.7717 / peerj.6573. ISSN 2167-8359. PMC 6404657. PMID 30863686.
- ^ Abler, W.L. (1992). "Tyrannosaurid dinozorların tırtıklı dişleri ve diğer hayvanlarda ısıran yapılar". Paleobiyoloji. 18 (2): 161–183. doi:10.1017 / S0094837300013956.
- ^ Goldstein, E. J. C .; Tyrrell, K. L .; Citron, D. M .; Cox, C. R .; Recchio, I. M .; Okimoto, B .; Bryja, J .; Fry, B.G. (1 Haziran 2013). "16 esir Komodo ejderhasından gelen tükürük ve diş etinin anaerobik ve aerobik bakteriyolojisi (Varanus komodoensis): "zehir olarak bakteri" modeli "için yeni çıkarımlar. Hayvanat Bahçesi ve Yaban Hayatı Tıbbı Dergisi. 44 (2): 262–272. doi:10.1638 / 2012-0022R.1. ISSN 1042-7260. PMID 23805543. S2CID 9932073.
- ^ Snively, E .; Cotton, J. R .; Ridgely, R .; Witmer, L.M. (2013). "Baş ve boyun fonksiyonunun çok gövdeli dinamik modeli Allosaurus (Dinosauria, Theropoda) ". Paleontoloji Electronica. 16 (2). doi:10.26879/338.
- ^ Rothschild, B .; Tanke, D. H .; Ford, T. L. (2001). "Aktiviteye ipucu olarak Theropod stres kırıkları ve tendon avülsiyonları". Tanke, D. H .; Carpenter, K. (editörler). Mesozoik Omurgalı Yaşamı. Indiana University Press. s. 331–336.
- ^ Wolff, E. D. S .; Salisbury, S. W .; Horner, J. R .; Varricchi, D. J. (2009). Hansen, D. M. (ed.). "Yaygın Kuş Enfeksiyonu Zalim Dinozorları Rahatsız Etti". PLOS ONE. 4 (9): e7288. Bibcode:2009PLoSO ... 4.7288W. doi:10.1371 / journal.pone.0007288. PMC 2748709. PMID 19789646.
- ^ a b c Longrich, N. R .; Horner, J. R .; Erickson, G. M .; Currie, P.J. (2010). "Yamyamlık Tyrannosaurus rex". PLOS ONE. 5 (10): e13419. Bibcode:2010PLoSO ... 513419L. doi:10.1371 / journal.pone.0013419. PMC 2955550. PMID 20976177.
- ^ Perkins, S. (29 Ekim 2015). "Tyrannosaurlar muhtemelen yamyamdı (Yorum)". Bilim. Alındı 2 Kasım, 2015.
- ^ Estes, R .; Berberyan, P. (1970). "Montana'dan geç Kretase omurgalı topluluğunun paleoekolojisi". Breviora. 343: 1–35.
- ^ Derstler, K. (1994). "Doğu Wyoming'deki Mızrak Formasyonunun Dinozorları". Nelson, G. E. (ed.). Wyoming Dinozorları. Wyoming Jeoloji Derneği Kılavuz Kitabı, 44. Yıllık Saha Konferansı. Wyoming Jeoloji Derneği. s. 127–146.
- ^ Weishampel, D. B .; Barrett, P. M .; Coria, R. A .; Loeuff, J. L .; Xu, X .; Zhao, X .; Sahni, A .; Gomani, E. M. P .; Noto, C.R. (2004). "Dinozor Dağılımı". Weishampel, D. B .; Dodson, P .; Osmólska, H. (editörler). Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. pp.574 –588. ISBN 978-0-520-24209-8.
- ^ Jasinski, S. E .; Sullivan, R. M .; Lucas, S. G. (2011). "Üst Kretase Ojo Alamo Formasyonundan Alamo Wash yerel faunasının taksonomik bileşimi (Naashoibito Üyesi) San Juan Havzası, New Mexico". Bülten. 53: 216–271.
- ^ Serrano-Brañas, C. I .; Torres-Rodrígueza, E .; Luna, P.C. R .; González, I .; González-León, C. (2014). "Lomas Coloradas Formasyonundan Tyrannosaurid dişler, Cabullona Group (Üst Kretase) Sonora, México". Kretase Araştırmaları. 49: 163–171. doi:10.1016 / j.cretres.2014.02.018.
- ^ Brusatte, S. L .; Carr, T. D. (2016). "Tyrannosauroid dinozorların soyoluşu ve evrimsel tarihi". Bilimsel Raporlar. 6: 20252. Bibcode:2016NatSR ... 620252B. doi:10.1038 / srep20252. PMC 4735739. PMID 26830019.
daha fazla okuma
- Farlow, J. O .; Gatesy, S. M .; Holtz, T.R., Jr .; Hutchinson, J. R .; Robinson, J.M. (2000). "Theropod Locomotion". Amerikalı Zoolog. 40 (4): 640–663. doi:10.1093 / icb / 40.4.640. JSTOR 3884284.
Dış bağlantılar
- The University of Edinburgh Lecture Dr Stephen Brusatte - Tyrannosaur Discoveries 20 Şubat 2015
- Tiranozor soy ağacındaki 28 tür, ne zaman ve nerede yaşadılar? Stephen Brusatte Thomas Carr 2016













