HIV-proteaz inhibitörlerinin keşfi ve geliştirilmesi - Discovery and development of HIV-protease inhibitors
Birçok önemli fizyolojik süreç, proteolitik enzim aktivite ve bir enzim ve onun arasındaki denge olduğunda dramatik sonuçlar olabilir. substratlar rahatsız. Bu ileriye dönük olarak, küçük molekül keşfi ligandlar, sevmek proteaz inhibitörleri katalitik aktiviteleri modüle edebilen, muazzam bir terapötik etkiye sahiptir.[1] Bu nedenle, HIV proteaz terapötik müdahale için en önemli yaklaşımlardan biridir. HIV enfeksiyon[2] ve onların gelişimi, büyük bir başarı olarak kabul edilir. yapı temelli ilaç tasarımı.[3] HIV'e karşı oldukça etkilidirler[4] ve 1990'lardan beri HIV / AIDS için anti-retroviral tedavilerin kilit bir bileşeni olmuştur.AIDS.[5]
Tarih
İnsan immün yetmezlik virüsü (HIV), lentivirüs iki büyük türe sahip olan HIV-1 bu da çoğunluğunun epidemi, ve HIV-2, dağılımı Batı Afrika'da yoğunlaşan yakın bir akraba.[6] HIV enfeksiyon ilk olarak 1981'de San Francisco ve New York'ta tanımlandı.[7] 1985 yılında HIV, edinilmiş immün yetmezlik sendromunun (AIDS) nedensel ajanı olarak tanımlandı ve genetik şifre hemen hazırdı. Bu bilgi, gelişiminin yolunu açtı seçici inhibitörler.[6]
HIV-2, HIV-1'den biraz daha düşük bulaşma riski taşır ve enfeksiyon, AIDS'e daha yavaş ilerleme eğilimindedir.[7] Yaygın kullanımda HIV genellikle HIV-1 anlamına gelir.[8]
HIV-1 proteaz en iyi bilinenlerden biridir aspartik proteazlar ve AIDS tedavisi için çekici bir hedef.[9]
HIV proteazının keşfinden sonra, ilk inhibitörünün pazara ulaşması yalnızca 10 yıl sürdü.[10] Son derece seçici ilk raporlar antagonistler HIV proteazına karşı 1987'de ortaya çıktı. Faz I denemeleri Sakinavir 1989'da başladı ve 1995'te reçeteli kullanım için onaylanan ilk HIV proteaz inhibitörü oldu. Dört ay sonra, diğer iki proteaz inhibitörü, ritonavir ve indinavir, onaylandı.[6] 2009 yılında on proteaz inhibitörü HIV'e karşı tedavi için pazara ulaştı, ancak bir proteaz inhibitörü, amprenavir, 2004 yılında piyasadan çekilmiştir.[6][11]
HIV'in yaşam döngüsü

HIV, adı verilen virüsler sınıfına aittir. retrovirüsler şeklinde genetik bilgi taşıyan RNA. HIV enfekte T hücreleri taşıyan CD4 antijen yüzeylerinde. HIV hedef hücresini enfekte ettiğinde, viral ve hücresel zarların füzyonunu gerektirir.[12] İlk adım, virüsün zarf proteinleri (gp120, gp41) ve hedef hücre üzerindeki spesifik konakçı hücre yüzey reseptörleri (örneğin CD4 reseptörü) arasındaki etkileşimdir. Daha sonra virüs, kemokin Koruyucular CXCR4 veya CCR5 zarf proteinlerinde yapısal değişikliklere neden olur. Bu füzyon, virüsün içinden geçtiği bir gözenek oluşturur. kapsid hücreye girer.[13] Hücreye girdikten sonra, virüsün RNA'sı, DNA ilk viral olarak kodlanan enzim, ters transkriptaz. Viral DNA, çekirdek tarafından hücrenin genetik materyaline entegre edildiği integral ikinci bir viral olarak kodlanmış enzim. Konakçı hücrenin aktivasyonu, transkripsiyon Viral DNA'nın içine mRNA. MRNA daha sonra viral proteinlere çevrilir ve üçüncü viral olarak kodlanmış enzim, yani HIV proteaz, bir viral poliprotein öncüsünü ayrı ayrı olgun proteinlere ayırmak için gereklidir. Viral RNA ve viral proteinler, hücrenin yüzeyinde birleşerek yeni Virionlar. Virionlar tomurcuk hücreden çıkar ve diğer hücreleri enfekte etmek için salınır. Konakçının genetik sisteminin yok edilmesinden viryonların tomurcuklanmasına ve salınmasına kadar bu kapsamlı hücre hasarı nedeniyle enfeksiyon kapmış tüm hücreler sonunda öldürülür.[12]
Hareket mekanizması
HIV yaşam döngüsünde müdahale edilebilecek ve böylece virüsün çoğalmasını durdurabilecek birkaç adım vardır. Çok kritik bir adım, polipeptit öncüllerinin olgun enzimlere ve yapısal proteinlere proteolitik bölünmesidir. katalize edilmiş HIV proteaz tarafından.[12] HIV proteaz inhibitörleri, virüs aspartil proteazının etkisini rekabetçi bir şekilde inhibe eden peptid benzeri kimyasallardır. Bu ilaçlar, virüsün temel yapısal ve enzimatik bileşenlerini içeren HIV Gag ve Pol poliproteinlerinin proteolitik bölünmesini önler. Bu, HIV partiküllerinin olgun bulaşıcı formlarına dönüşmesini engeller.[6]
Proteaz inhibitörleri değişebilir adiposit neden metabolizma lipodistrofi, Ortak yan etki HIV proteaz inhibitörlerinin çoğunun kullanımı ile ilişkilidir. Adiposit inhibisyonu gibi birçok mekanizma önerilmiştir. farklılaşma, trigliserid birikim ve artış lipoliz. Proteaz inhibitörlerinin insülinle uyarılan glukoz alımı üzerindeki etkisini dikkate alan teoriler de lipodistrofik sendromla ilişkilendirilmiştir. Proteaz inhibitörlerinin bir azalmaya neden olması mümkündür. insülin uyarılmış tirozin fosforilasyon IRS-1, insülin sinyallemesinde erken aşamaların inhibisyonunu temsil eder. Azaldı adiponektin salgılanması ve uyarılmış ifadesi interlökin-6 HIV proteaz inhibitörleri ile bağlantılı olarak, insülin ile uyarılan glukoz alımının inhibisyonuna da katkıda bulunabilir.[14]
Tasarım
Proteaz inhibitörleri, geçiş durumu proteazın gerçek substratlar. Bir peptid bağı -NH-CO- içeren bir hidroksietilen grubu (-CH2-CH (OH) -) proteazın parçalanamadığı. HIV proteaz inhibitörleri, aktif site ve aspartil proteazın bilgisinden yararlanılarak rasyonel olarak tasarlandı. aksiyon modu. En umut verici geçiş durumu taklidi, ilk proteaz inhibitörünün keşfine yol açan hidroksietilamindi. Sakinavir. Bu keşfin ardından, diğer HIV proteaz inhibitörleri de aynı prensip kullanılarak tasarlandı.[15]
Bağlayıcı site

HIV proteaz, iki 99 içeren C2-simetrik homodimerik bir enzimdir. amino asit monomerler. Her monomer bir katkıda bulunur aspartik asit kataliz için gerekli olan kalıntı,[6] Asp-25 ve Asp-25´. HIV proteaz, Asp- dizisine sahiptir.Thr -Gly bu, diğer memeli aspartik proteaz enzimleri arasında korunur. Genişletilmiş beta sayfası kanat olarak bilinen monomerler üzerindeki bölge, kısmen substrat bağlanma bölgesini oluşturur ve iki aspartil kalıntısının bir hidrofobik boşluk.[12][16][17] Her esnek kanat üç karakteristik bölge içerir: dışa doğru uzanan yan zincirler (Tanışmak 46, Phe 53), içeriye doğru uzanan hidrofobik zincirler (Ile 47, Ile54) ve glisinden zengin bir bölge (Gly48, 49, 51, 52). İle50 dönüşün ucunda kalır ve enzim bağımsız olduğunda bir su molekülü hidrojen bağları Her monomerde Ile50 omurgasına.[17]
HIV proteazları, hidroliz yüksek sekans seçiciliği ve katalitik yeterliliğe sahip peptit bağları. HIV proteazının mekanizması, aspartik proteaz ailesinin geri kalanıyla birçok özelliği paylaşır, ancak bu enzimin tam ayrıntılı mekanizması tam olarak anlaşılmamıştır.[12] Su molekülü, kanatçıkların açılıp kapanmasında ve enzim ile substrat arasındaki afinitenin artmasında rol oynuyor gibi görünmektedir. Aspartil kalıntıları, peptit bağlarının hidrolizinde rol oynar.[17] Bu enzim için tercih edilen bölünme bölgesi, N terminali prolin kalıntılarının yanı, özellikle fenilalanin ile prolin veya tirozin arasında ve prolin.[6][16]
Geliştirme
İlk HIV proteaz inhibitörü olan sakinavir, bir peptidomimetik hidroksietilamin[6] 1995 yılında piyasaya sürüldü.[18] Bu bir geçiş halidir analog proteazın doğal bir substratının.[6] HIV-1 proteazın, Tyr-Pro veya Phe-Pro dipeptitlerini içeren dizileri klevaj ettiği gözlemi, temel tasarım kriteriydi.[19] Dekahidroizokinolin (DIQ) grubunun eklenmesi, sakuinavirin keşfine yol açan en önemli modifikasyonlardan biriydi. Bu ikame maddesi, inhibitörün konformasyonel özgürlüğünü sınırlandırarak sulu çözünürlüğü ve gücü geliştirir.[20] Saquinavir hem HIV-1 hem de HIV-2'ye karşı etkilidir[5] ve genellikle iyi tolere edilir ancak yüksek serum konsantrasyonuna ulaşılamaz.[11]
Ritonavir bir peptidomimetik HIV proteaz inhibitörü olan 1996 yılında piyasaya sürüldü.[18] C2-simetrisini proteazın bağlanma bölgesine uyacak şekilde tasarlanmıştır.[6] Ritonavir geliştiricileri, Abbott Laboratuvarları, virüse karşı aktif olan ancak zayıf olan bileşiklerle başladı. biyoyararlanım. Bazı iyileştirmeler yapıldı, örneğin terminal fenil kalıntıları çıkarıldı ve piridil gruplar bunun yerine suda çözünürlük eklemek için koydu. Bu iyileştirmelerin nihai ürünü ritonavirdi.[19] Önemli gastrointestinal yan etkiler ve büyük hap yükü, ritonavirin ana dezavantajlarıdır ve bu nedenle tek bir tedavi olarak kullanılmamaktadır.[11] Bununla birlikte, sitokrom P450 enzimi aracılı metabolizmanın güçlü bir inhibitörüdür.[19] ve sadece farmakokinetik güçlendirme için diğer proteaz inhibitörleri ile kombinasyon terapisinde kullanılır.[11]
Indinavir Peptidomimetik bir hidroksietilen HIV proteaz inhibitörü olan 1996 yılında piyasaya çıktı.[6][18] Indinavirin tasarımı moleküler modelleme ve Röntgen kristal yapı inhibe edilmiş enzim kompleksinin. Terminal fenil bileşenleri hidrofobik bağlanmanın artmasına katkıda bulunur. güç.[19] HIV Gag-poliproteininin fenilalanin-prolin bölünme bölgesinin bir analoğudur.[6]
Nelfinavir peptidomimetik olmayan ilk proteaz inhibitörü idi. Oral olarak biyolojik olarak temin edilebilen ve peptidik olmayan bir inhibitör olan nelfinavirin tasarım sürecinde, peptidik inhibitörlerin yinelemeli protein birlikte kristal yapı analizi kullanılmış ve inhibitörlerin kısımları peptidik olmayan ikame ediciler ile değiştirilmiştir.[19] Nelfinavir yeni bir 2-metil-3-hidroksibenzamid grubu içerirken, karboksil terminali saquinavir ile aynı DIQ grubunu içerir.[19] Nelfinavir 1997'de pazarlandı[18] ve endike olan ilk proteaz inhibitörü idi. pediatrik AIDS.[19]
Amprenavir pazara 1999 yılında ulaştı.[18] O bir N,Nikame edilmiş aminosülfonamid peptid olmayan HIV proteaz inhibitörü[6] ve önceki proteaz inhibitörleri ile bazı ortak özellikleri paylaşır. Sakinavire benzer ancak farklı bir çekirdeğe sahiptir. fonksiyonel gruplar her iki ucunda. Bir ucunda tetrahidrofuran karbamat grubu ve diğer uçta eklenmiş bir amid ile bir izobutilfenil sülfonamiddir. Bu yapı, daha az kiral sentezlemeyi kolaylaştıran ve gelişmiş aqueus çözünürlüğü veren merkezler. Bu da daha iyi oral biyoyararlanım sağlar.[19] Ancak, amprenavir, fosamprenavir nedeniyle 2004 yılında piyasadan çekilmiştir. ön ilaç, birçok yönden üstün olduğunu kanıtladı.[6]
Lopinavir 2000 yılında piyasaya sürüldü[18] ve başlangıçta inhibitörün etkileşimlerini azaltmak için tasarlanmıştır. Val HIV-1 proteazın 82'si, sıklıkla mutasyona uğramış içinde ilaca dirençli suşlar virüsün.[19] Peptidomimetik bir HIV proteaz inhibitörüdür[6] ve çekirdeği ritonavirinkiyle aynıdır. 5- yerinetiazolil ritonavir son grubu, lopinavir bir fenoksiasetil grubuna sahiptir ve ritonavirdeki 2-izopropiltiyazolil grubu, amino terminalin altı üyeli bir siklikliğe sahip olduğu modifiye bir valin ile değiştirilmiştir. üre ekli.[19]
Fosamprenavir 2003 yılında piyasaya sürüldü[18] ve hızla ve büyük ölçüde amprenavire metabolize olan bir fosfoester ön ilacıdır.[21] Çözünürlük ve biyoyararlanım amprenavir'den daha iyidir[6] bu da günlük hap yükünün azalmasına neden olur.[22]
Atazanavir 2003 yılında piyasaya sürüldü[18] ve bir azapeptid proteaz inhibitörüdür[18] enzim bağlanma bölgesinin C2 simetrisine uyacak şekilde tasarlanmıştır.[11] Atazanavir, önceki HIV proteaz inhibitörlerinden daha iyi dirençli profiller göstermiştir.[4] Sadece olabileceği için diğer proteaz inhibitörleri arasında benzersizdir. emilmiş asidik bir ortamda.[11]
Tipranavir peptidik olmayan bir HIV-1 proteaz inhibitörüdür[11] 2005 yılında pazara ulaştı.[18] Piyasadaki diğer HIV proteaz inhibitörlerinin aksine, tipranavir peptidik olmayan kumarin şablon ve antiproteaz aktivitesi, yüksek verimli tarama.[23] 5,6-dihidro-4-hidroksi-2-pirron içeren bu sülfonamid, 3-ikame edilmiş kumarin ve dihidropiron taramalarından ortaya çıkmıştır.[24] Çoklu proteaz inhibitörüne dirençli HIV-1'e karşı geniş antiviral aktiviteye sahiptir.[25]
Darunavir 2006 yılında pazara ulaştı[18] ve terminal tetrahidrofuran (THF) grubunda kritik bir değişiklikle amprenavirin peptidik olmayan bir analoğudur. Tek bir THF grubu yerine darunavir, bir bis-THF oluşturmak için bileşikte kaynaşmış iki THF grubu içerir. parça bu onu amprenavirden daha etkili kılar. Bu yapısal değişiklikle birlikte stereokimya bis-THF parçası etrafında, amprenavir için bir direnç geliştiren proteaz ile sürekli bağlanmaya izin veren oryantasyon değişiklikleri sağlar.[26]
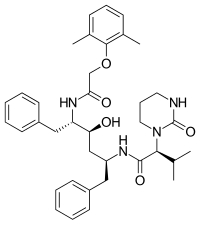
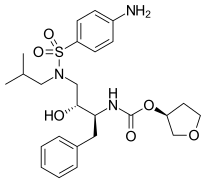
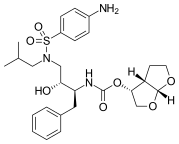
FDA onaylı tüm proteaz inhibitörleri aşağıda listelenmiştir.
 | 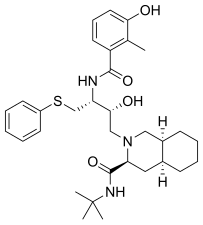 |  |  |
| Saquinavir | Nelfinavir | Ritonavir | Lopinavir |
 |  |  | |
| Amprenavir | Fosamprenavir | Darunavir | |
 |  |  | |
| Indinavir | Atazanavir | Tipranavir |
Yapı-aktivite ilişkisi

Piyasadaki tüm HIV proteaz inhibitörleri, bir kumarin iskelesine dayanan tipranavirin merkezi çekirdeği dışında, bir hidroksietilen iskeleden oluşan bir merkezi çekirdek motifi içerir.[15] HIV proteaz inhibitörleri üzerindeki çok önemli bir grup, hidroksil ile bir hidrojen bağı oluşturan çekirdek motif üzerindeki grup karboksilik asit bağlanma sahasındaki Asp-25 ve Asp-25´ tortuları üzerinde.[16][27] Ile50 ve Ile50'ye bağlı olan su molekülü arasındaki hidrojen bağları ve karbonil peptidomimetik inhibitör grupları, onları flep bölgeleri ile birleştiriyor gibi görünmektedir.[19] Öte yandan, peptidik olmayan inhibitörlerde, yerine geçen bir proton alıcısı vardır. dört koordineli su molekülü ve enzimin kanadı üzerindeki iki Ile50 kalıntısı ile doğrudan etkileşime girer.[28] Genellikle S1, S1 ', S2 ve S2' olarak anılan HIV proteazın bağlanma bölgesindeki spesifik cepler, doğal substratlar üzerindeki hidrofobik amino asitleri tanır. Bu alanları tamamlayan hidrofobik grupları taşıyan inhibitörlerin gücü bu nedenle artar.[29] Enzim bağlanma sahasındaki bazı tortular, inhibitör üzerindeki hidrofilik gruplarla, örneğin amprenavir ve darunavir üzerindeki THF kısımlarıyla hidrojen bağları oluşturabilir. Darunavir, amprenavir'deki gibi tek bir THF parçası yerine bir bis-THF parçasına sahip olduğundan, daha fazla hidrojen bağı oluşturabilir ve artabilir bağlanma enerjisi.[26]
Direnç
Konformasyonel şeklin değişikliklerini kodlayan mutasyonlar, HIV'in proteaz inhibitörlerine karşı direncini kolaylaştırır.[26] Bu mutasyonların konumları, Gag-Pol poliprotein öncülerindeki proteaz yarılma bölgelerinde bulunanlar dahil olmak üzere, öncelikle HIV proteaz enziminin aktif bölgesinde ve ayrıca aktif bölgenin dışındadır. Bölünme bölgeleri çok çeşitli sekanslara sahiptir, bu nedenle proteaz, substratlarını sekansa değil, aktif sahada bağlandıklarında substratların paylaştığı korunmuş 3B şekli tanır. Bu korunan şekil, alt tabaka zarfı.[30] Aktif bölge mutasyonlarının, inhibitörlerin etkileşimlerini doğrudan değiştirdiği ve çoğunlukla inhibitörlerin substrat zarfının ötesinde proteaz kalıntılarına temas ettiği pozisyonlarda meydana geldiği gösterilmiştir.[31] Aktif olmayan site mutasyonlarının, etkileme gibi diğer mekanizmalar tarafından etkilendiği kabul edilir. dimer kararlılık ve konformasyonel esneklik.[32][33]
100'den fazla tek gen nokta mutasyonları en az 26'sı proteaz inhibitörlerine özgü olan tarif edilmiştir. Bunlardan, ilaç aktivitesini değiştirecek kadar önemli olan yaklaşık 15 birincil veya büyük mutasyon vardır.[26]HIV-1 proteazında ilaç direncine neden olan birçok mutasyona uğramış kalıntı bulunmuştur, örneğin Leu33, Ile, Val veya Phe'ye değişir; Val82 - Ala, Phe, Leu veya Thr; Ile84 ila Val; ve Leu90'dan Met'e.[34] Farklı mutasyonlar, farklı proteaz inhibitörlerini etkiler. Örneğin, Leu90'daki mutasyonlar açıkça sakinavir ve nelfinavir'i etkilerken indinavir aktivitesi Met46, Val82 ve Ile84'teki mutasyonlardan etkilenir ve fosamprenavir Ile50, Val ve Ile84'te değiştiğinde etkilenir. Mutasyonların bir kombinasyonu, yüksek düzeyde ilaç direnci sağlayabilir ancak tekli mutasyonlar normalde proteaz inhibitörlerine karşı ilaç direnci ile eşit değildir.[26]Mutasyonlar, birincil mutasyonlara ve ikincil mutasyonlara ayrılabilir. Birincil mutasyonların genellikle direnç üzerinde sadece küçük bir etkisi vardır. Çoğu proteaz inhibitörünün kimyasal yapıları oldukça benzerdir, bu nedenle bazı birincil mutasyonların aynı anda birden fazla proteaz inhibitörüne dirence yol açması şaşırtıcı değildir. Çapraz direnç proteaz inhibitörü tedavisinin en önemli sorunlarından biridir.[35] Sürekli proteaz inhibitörü tedavisi sırasında proteazda ortaya çıkan ek mutasyonlar, genellikle ikincil mutasyonlar olarak adlandırılır. Bu, yüksek düzeyde proteaz inhibitörü direncine yol açabilir.[35]
Stanford HIV RT ve Proteaz Sekans Veritabanı ("HIV İlaç Direnci Veritabanı" olarak da adlandırılır) 1998 yılında iyi karakterize edilmiş antiretroviral tedavi geçmişlerine sahip kişilerden alınan HIV ters transkriptaz ve proteaz sekansları ile oluşturulmuştur ve direnç mutasyonlarını ve genotipini sorgulamak için halka açıktır. - tedavi, genotip-fenotip ve genotip-sonuç korelasyonları: http://hivdb.stanford.edu
Substrat zarfı, çoğu aktif saha mutasyonunun sağladığı direnci önlemek için substratı taklit eden ve zarf içinde kalan inhibitörler tasarlama genel stratejisini sağlasa da,[36][37] Özellikle aktif bölgeden uzakta olanlar nedeniyle, ilaç direnci sorununu çözmek için genel bir strateji yoktur. AIDS'i iyileştirmek için yeni tedavilerin geliştirilmesine yönelik araştırmalar, halihazırda piyasada bulunan ilaçlara karşı çapraz dirençten kaçınmaya odaklanmıştır.[12]
Şu anki durum
Ocak 2018'de darunavir, pazara ulaşan en son HIV proteaz inhibitörü oldu.[38]
2006 yılında GlaxoSmithKline faz II klinik gelişimini durdurdu Brecanavir, formülasyonla ilgili aşılmaz sorunlar nedeniyle HIV tedavisi için bir araştırma proteaz inhibitörü.[39]
2009 yazında, GlaxoSmithKline ve Concert Pharmaceuticals, geliştirmek ve ticarileştirmek için işbirliğini duyurdu. döteryum - içeren ilaçlar. Bunlardan biri, HIV tedavisi için bir proteaz inhibitörü olan CTP-518'dir ve 2009'un ikinci yarısında faz I klinik denemelere girmesi beklenmektedir. CTP-518, atazanavirin belirli anahtar hidrojen atomlarının yerine geçerek geliştirilen yeni bir HIV proteaz inhibitörüdür. döteryum. Klinik öncesi çalışmalar, bu modifikasyonun antiviral potensi tamamen koruduğunu ancak açıkça hepatik metabolizmayı yavaşlattığını ve böylece yarılanma ömrü ve plazmayı artırdığını göstermiştir çukur seviyeleri. Bu nedenle CTP-518, ritonavir gibi bir boost edici ajan ile birlikte dozlama ihtiyacını ortadan kaldıran ilk HIV proteaz inhibitörü olma potansiyeline sahiptir.[40]
Ayrıca bakınız
- Antiretroviral ilaç
- Ters transkriptaz inhibitörü
- Integrase inhibitörü
- Giriş engelleyici
- Nükleozid olmayan ters transkriptaz inhibitörlerinin keşfi ve geliştirilmesi
Referanslar
- ^ Cuccioloni, M; Mozzicafreddo, M; Bonfili, L; Cecarini, V; Eleuteri, A. M .; Angeletti, M (2009). "İlaç tasarımı için şablon olarak doğal oluşan polifenoller. Serin proteazlara odaklanın". Kimyasal Biyoloji ve İlaç Tasarımı. 74 (1): 1–15. doi:10.1111 / j.1747-0285.2009.00836.x. PMID 19519739.
- ^ Chen, X; Kempf, D. J .; Küçük; Sham, H.L .; Vasavanonda, S; Wideburg, N.E .; Saldivar, A; Marsh, K. C .; McDonald, E; Norbeck, D.W. (2003). "P2 ligandları olarak yeni dimetilfenoksil asetat içeren güçlü HIV proteaz inhibitörlerinin sentezi ve SAR çalışmaları". Biyorganik ve Tıbbi Kimya Mektupları. 13 (21): 3657–60. doi:10.1016 / j.bmcl.2003.08.043. PMID 14552751.
- ^ Adachi, M; Ohhara, T; Kurihara, K; Tamada, T; Honjo, E; Okazaki, N; Arai, S; Shoyama, Y; Kimura, K; Matsumura, H; Sugiyama, S; Adachi, H; Takano, K; Mori, Y; Hidaka, K; Kimura, T; Hayashi, Y; Kiso, Y; Kuroki, R (2009). "Güçlü inhibitör KNI-272 ile kompleks içindeki HIV-1 proteaz yapısı, yüksek çözünürlüklü X-ışını ve nötron kristalografisi ile belirlenir". Ulusal Bilimler Akademisi Bildiriler Kitabı. 106 (12): 4641–6. doi:10.1073 / pnas.0809400106. PMC 2660780. PMID 19273847.
- ^ a b Yanchunas Jr, J; Langley, D. R .; Tao, L; Rose, R. E .; Fribourg, J; Colonno, R. J .; Doyle, M.L. (2005). "Diğer proteaz inhibitörlerine I50L ikame sağlayan atazanavir direnci olan izolatların artan duyarlılığının moleküler temeli". Antimikrobiyal Ajanlar ve Kemoterapi. 49 (9): 3825–32. doi:10.1128 / AAC.49.9.3825-3832.2005. PMC 1195399. PMID 16127059.
- ^ a b Brower, E. T .; Bacha, U. M .; Kawasaki, Y; Freire, E (2008). "Klinik kullanımda HIV-1 proteaz inhibitörleri tarafından HIV-2 proteaz inhibisyonu". Kimyasal Biyoloji ve İlaç Tasarımı. 71 (4): 298–305. doi:10.1111 / j.1747-0285.2008.00647.x. PMID 18312292.
- ^ a b c d e f g h ben j k l m n Ö p Brunton, L.L .; Lazo, J.S .; Parker, K.L. (2006). Goodman ve Gilmans'ın Tedavinin Farmakolojik Temelleri (11. baskı). McGraw-Hill.[sayfa gerekli ]
- ^ a b HIV Hastalığı -de eTıp
- ^ Kurup, Alka; Mekapati, Suresh; Garg, Rajni; Hansch, Corwin (2003). "HIV-1 Proteaz İnhibitörleri: Karşılaştırmalı QSAR Analizi". Güncel Tıbbi Kimya. 10 (17): 1679–88. doi:10.2174/0929867033457070. PMID 12871116.
- ^ Shi, Haibin; Liu, Kai; Leong, Wendy W.Y .; Yao, Shao Q. (2009). "Aspartik proteazları hedefleyen hem simetrik hem de asimetrik diol kitaplıklarının uygun katı faz sentezi". Biyorganik ve Tıbbi Kimya Mektupları. 19 (14): 3945–8. doi:10.1016 / j.bmcl.2009.03.041. PMID 19328682.
- ^ Türk, Boris (2006). "Proteazları hedefleme: başarılar, başarısızlıklar ve gelecekteki beklentiler". Doğa İncelemeleri İlaç Keşfi. 5 (9): 785–99. doi:10.1038 / nrd2092. PMID 16955069.
- ^ a b c d e f g Graziani, Amy L (17 Haziran 2014). "HIV proteaz inhibitörleri". Güncel.
- ^ a b c d e f Brik, A. ve Wong, C.H. (2003) HIV-1 proteaz: mekanizma ve ilaç keşfi. Organik ve Biyomoleküler Kimya. 1(1); 5–14.
- ^ Warnke, D., Barreto, J. ve Temesgen, Z. (2007) Antiretroviral ilaçlar. Klinik Farmakoloji Dergisi. 47(12); 1570–1579.
- ^ Kim, R.J., Wilson, C.G., Wabitsch, M., Lazar, M.A. ve Steppan, C.M. (2006) İnsan adiposit farklılaşmasında ve metabolizmasında HIV proteaz inhibitörüne özgü değişiklikler. Obezite. 14; 994–1002.
- ^ a b De Clercq, E. (2009) Antiretrovirallerin tarihi: son 25 yıldaki temel keşifler. Tıbbi Viroloji İncelemeleri. 19; 287–299.
- ^ a b c Mimoto, T., Hattori, N., Takaku, H. vd. (2000) Hidroksimetilkarbonil izoster içeren oral olarak güçlü tripeptit bazlı HIV proteaz inhibitörlerinin yapı-aktivite ilişkisi. Kimya ve İlaç Bülteni. 48(9); 1310–1326.
- ^ a b c Perez, M.A.S., Fernandes, P.A. ve Ramos, M.J. (2007) İlaç tasarımı: Kurşun olarak Nelfinavire dayanan HIV-1 proteaz için yeni inhibitörler. Moleküler Grafik ve Modelleme Dergisi. 26; 634–642.
- ^ a b c d e f g h ben j k Flexner, C. (2007) HIV ilaç geliştirme: önümüzdeki 25 yıl. Doğa İncelemeleri İlaç Keşfi. 6; 959–966.
- ^ a b c d e f g h ben j k Wlodawer, A. (2002) Yapısal biyoloji aracılığıyla AIDS ilaç tasarımına akılcı yaklaşım. Yıllık Tıp İncelemesi. 53; 595–614.
- ^ Smith, H.J. ve Simons, C. (2005) Enzimler ve İnhibisyonu: İlaç Geliştirme (6. baskı). Amerika Birleşik Devletleri: CRC press
- ^ Chapman, T.M., Plosker, G.L. ve Perry, C.M. (2004) Fosamprenavir - HIV Enfeksiyonlu Antiretroviral Terapi Almamış Hastaların Yönetiminde Kullanımına İlişkin Bir İnceleme. İlaçlar. 64; 2101–2124.
- ^ a b c d e McCoy, C. (2007) Darunavir: Bir peptidik olmayan antiretroviral proteaz inhibitörü. Klinik Terapötikler. 29(8); 1559–1576.
- ^ Liu, F., Kovalevsky, A.Y., Tie, Y., Ghosh, A.K., Harrison, R.W. ve Weber, I.T. (2008) Flap Mutasyonlarının HIV-proteaz Yapısı ve Saquinavir ve Darunavir'in İnhibisyonu Üzerindeki Etkisi. Moleküler Biyoloji Dergisi. 381(1); 102–115
- ^ Lebon, F. ve Ledecq, M. (2000) Etkili HIV-1 Proteaz İnhibitörlerinin Tasarımına Yaklaşımlar. Güncel Tıbbi Kimya. 7; 455–477.
- ^ Blum, A. vd. (2008) Aspartik proteaz inhibitörlerinin geliştirilmesi için çok yönlü bir araç olarak aşiral oligoaminler. Biyorganik ve Tıbbi Kimya. 16; 8574–8586.
- ^ Prabu-Jeyabalan, Nalivaika E, Schiffer CA. (2002) Substrat şekli, HIV-1 proteaz için tanımanın özgüllüğünü belirler: altı substrat kompleksinin kristal yapılarının analizi. "Yapı" 10 (3): 369-81.
- ^ King NM, Prabu-Jeyabalan M, Nalivaika EA, Schiffer CA (2004) İlaç direncine duyarlılıkla mücadele: HIV-1 protease.Chem Biol'den dersler. Ekim; 11 (10): 1333-8.
- ^ Bihani, S. C., Das, A., Prashar, V., Ferrer, J.-L. ve Hosur; M.V. (2009) Bağımsız nelfinavire dirençli HIV-1 proteaz aktif olmayan site mutantları N88D ve N88S'nin kristal yapıları tarafından ortaya konan direnç mekanizması. Biyokimyasal ve Biyofiziksel Araştırma İletişimi. 389; 295–300.
- ^ de Vera IM, Smith AN, Dancel MC, Huang X, Dunn BM, Fanucci GE. (2013) Biyokimya. HIV-1 proteazda konformasyonel örnekleme ve ilaç direnci arasındaki ilişkiyi açıklamak. 14; 52 (19): 3278-88. doi: 10.1021 / bi400109d. Epub 2013 1 Mayıs.
- ^ Lemke, T.L., Williams, D.A., Roche, V.F. ve Zito, S.W. (2008) Fuaye'nin Tıbbi Kimya İlkeleri (6. baskı). Amerika Birleşik Devletleri: Lippincott williams & Wilkins, bir Wolters Kluwer işletmesi.
- ^ a b Maarseveen, N.V. ve Boucher, C. (2008) Klinik Uygulamada Antiretroviral Direnç. Londra: Mediscript Ltd.
- ^ Kairys V, Gilson MK, Lather V, Schiffer CA, Fernandes MX. (2009) Mutasyona dirençli enzim inhibitörlerinin tasarımına doğru: substrat zarfı hipotezinin daha fazla değerlendirilmesi. Chem Biol Drug Des. Eylül; 74 (3): 234-45. doi: 10.1111 / j.1747-0285.2009.00851.x
- ^ Nalam MN, Ali A, Altman MD, Reddy GS, Chellappan S, Kairys V, Ozen A, Cao H, Gilson MK, Tidor B, Rana TM, Schiffer CA. (2010) Substrat zarf hipotezinin değerlendirilmesi: ilaç direncine karşı sağlam olacak şekilde tasarlanmış yeni HIV-1 proteaz inhibitörlerinin yapısal analizi. J Virol. 2010 Mayıs; 84 (10): 5368-78. doi: 10.1128 / JVI.02531-09. Epub 2010 17 Mart.
- ^ De Clercq, E. (2009) Anti-HIV ilaçlar: HIV keşfinden sonraki 25 yıl içinde onaylanan 25 bileşik. International Journal of Antimicrobial Agents. 33; 307–320.
- ^ "Arşivlenmiş kopya". Arşivlenen orijinal 2008-12-03 tarihinde. Alındı 2008-06-11.CS1 Maint: başlık olarak arşivlenmiş kopya (bağlantı) GlaxoSmithKline, araştırma amaçlı proteaz inhibitörü brecanavirin klinik gelişimini durdurur (640385). 4 Kasım alındı. 2009.
- ^ "Arşivlenmiş kopya". Arşivlenen orijinal 2009-08-31 tarihinde. Alındı 2009-11-05.CS1 Maint: başlık olarak arşivlenmiş kopya (bağlantı)GSK ve Concert Pharmaceuticals, döteryumla modifiye edilmiş yeni ilaçlar geliştirmek için ittifak kuruyor. 4 Kasım alındı. 2009.