Endojen retrovirüs - Endogenous retrovirus - Wikipedia
Endojen retrovirüsler (ERV'ler) endojen viral elementler içinde genetik şifre yakından benzeyen ve türetilebilen retrovirüsler. Genomlarında bol miktarda bulunurlar çeneli omurgalılar ve insan genomunun% 5-8'ini oluştururlar (daha düşük tahminler ~% 1).[1][2] ERV'ler, bir tür gen deniliyor transpozon içinde yaşamsal bir rol oynaması için genom içinde paketlenip taşınabilen gen ifadesi ve düzenleme.[3][4] Olarak ayırt edilirler retrotranspozonlar Sınıf I öğeleridir.[5] Araştırmacılar, retrovirüslerin, ERV'leri içeren, retrotranspozon adı verilen bir tür transpoze edilebilir genden evrimleştiğini öne sürmüşlerdir; bu genler mutasyona uğrayabilir ve genomda başka bir yere taşınmak yerine eksojen veya patojenik hale gelebilir. Bu, tüm ERV'lerin bir retrovirüs tarafından bir ekleme olarak ortaya çıkmamış olabileceği, ancak bazılarının benzedikleri retrovirüslerdeki genetik bilginin kaynağı olabileceği anlamına gelir.[6] Viral DNA'nın germ hattında entegrasyonu gerçekleştiğinde, daha sonra konakçı popülasyonun gen havuzunda sabitlenebilen bir ERV'ye yol açabilir.[1][7]
Oluşumu
Bir çoğaltma döngüsü retrovirüs viral DNA kopyasının eklenmesini ("entegrasyon") gerektirir genetik şifre içine nükleer genomu ev sahibi hücre. Retrovirüslerin çoğu enfekte somatik hücreler, ancak ara sıra enfeksiyon germ hattı hücreler (yumurta ve sperm üreten hücreler) de oluşabilir. Nadiren, retroviral entegrasyon, yaşayabilir bir organizmaya dönüşmeye devam eden bir germ hattı hücresinde meydana gelebilir. Bu organizma, eklenen retroviral genomu kendi genomunun ayrılmaz bir parçası olarak taşıyacaktır - "endojen" retrovirüs (ERV) miras onun tarafından yavru roman olarak alel. Birçok ERV, konakçılarının genomunda milyonlarca yıldır varlığını sürdürmüştür. Bununla birlikte, bunların çoğu inaktive edici hale geldi mutasyonlar ev sahibi sırasında DNA kopyalama ve artık virüsü üretemez. ERV'ler ayrıca rekombinasyonel delesyon olarak bilinen bir işlemle genomdan kısmen çıkarılabilir; rekombinasyon yeni entegre retrovirüsleri çevreleyen özdeş diziler arasında, iç kısımların silinmesine neden olur, protein - viral genomun kodlama bölgeleri.
Genel retrovirüs genomu, viral genomunun istilası, replikasyonu, kaçışı ve yayılması için hayati önem taşıyan üç genden oluşur. Bu üç gen gag'dir (viral çekirdek için yapısal proteinleri kodlar), pol (için kodlar) ters transkriptaz, integral, ve proteaz ) ve env (kodlar kaplama proteinleri virüsün dışı için). Bu viral proteinler şu şekilde kodlanır: poliproteinler. Retrovirüs, yaşam döngülerini gerçekleştirmek için büyük ölçüde konakçı hücrenin mekanizmasına dayanır. Proteaz, viral poliproteinlerin peptit bağlarını bozarak ayrı proteinleri işlevsel hale getirir. Ters transkriptaz, nükleusa girmeden önce konakçı hücrenin sitoplazmasındaki viral RNA'dan viral DNA sentezleme işlevi görür. Integrase, viral DNA'nın konak genomuna entegrasyonuna rehberlik eder.[7][8]
Zamanla, ERV'lerin genomu yalnızca nokta mutasyonları elde etmekle kalmaz, aynı zamanda diğer ERV'lerle karıştırılır ve yeniden birleşir.[9] İle ERV'ler env çürüyen kürkün yayılma olasılığı daha yüksektir.[10]
Genom evrimindeki rol

Endojen retrovirüsler, genomların şekillendirilmesinde aktif bir rol oynayabilir. Bu alandaki çoğu çalışma, insanların ve daha yüksek primatların genomlarına odaklanmıştır, ancak fareler ve koyunlar gibi diğer omurgalılar da derinlemesine incelenmiştir.[11][12][13][14] Uzun terminal tekrarı (LTR ) ERV genomlarını çevreleyen diziler sıklıkla alternatif olarak hareket eder destekçiler ve geliştiriciler, genellikle katkıda bulunur transkriptom dokuya özgü varyantlar üreterek. Ek olarak, retroviral proteinler kendileri, özellikle üreme ve geliştirmede yeni konakçı işlevlerine hizmet etmek üzere seçilmiştir. Homolog retroviral diziler arasındaki rekombinasyon, gen karıştırılmasına ve genetik varyasyonun oluşmasına da katkıda bulunmuştur. Dahası, retroviral sekansların potansiyel olarak antagonistik etkileri durumunda, baskılayıcı genler onlarla savaşmak için birlikte gelişti.
Endojen retrovirüslerin yaklaşık% 90'ı tek başına LTR'lerdir, açık okuma çerçeveleri (ORF'ler).[15] Tam retroviral sekanslarla ilişkili solo LTR'lerin ve LTR'lerin konakçı genler üzerinde transkripsiyonel elementler olarak hareket ettiği gösterilmiştir. Eylem aralıkları esas olarak 5 ' UTR'ler protein kodlayan genlerin; ancak, 70-100'e kadar genler üzerinde etkili oldukları bilinmektedir.kb uzakta.[11][16][17][18] Bu elementlerin çoğu, karşılık gelen genlerine duyu yönünde eklenir, ancak LTR'lerin antisens yönde ve komşu genler için çift yönlü bir destekleyici olarak hareket ettiğine dair kanıtlar vardır.[19][20] Birkaç durumda, LTR gen için ana destekleyici olarak işlev görür. Örneğin insanlarda AMY1C, promoter bölgesinde tam bir ERV sekansına sahiptir; ilişkili LTR, sindirimin tükürüğe özgü ekspresyonunu sağlar enzim amilaz.[21] Ayrıca, birincil destekleyici safra asidi-CoA: amino asit N-asiltransferaz Safra metabolizmasının ayrılmaz bir parçası olan bir enzimi kodlayan (BAAT), LTR kökenlidir.[17][22]
Yalnız bir ERV-9 LTR'nin takılması, işlevsel bir açık okuma çerçevesi oluşturmuş olabilir ve bu da insanın yeniden doğmasına neden olmuş olabilir. bağışıklık ile ilgili GTPaz geni (IRGM).[23] ERV eklemelerinin, ya insan leptin hormonu reseptöründe olduğu gibi gene doğrudan entegrasyon yoluyla ya da fosfolipaz A-2 benzeri proteinde olduğu gibi bir akış yukarı LTR'nin ekspresyonu tarafından tahrik edilerek alternatif ek yerleri oluşturduğu da gösterilmiştir.[24]
Bununla birlikte çoğu zaman, LTR birçok alternatif promotörden biri olarak işlev görür ve sıklıkla üreme ve gelişmeyle ilgili dokuya özgü ekspresyon verir. Aslında, bilinen LTR destekli transkripsiyon varyantlarının% 64'ü ifade üreme dokularında.[25] Örneğin, CYP19 geni, aromataz Östrojen sentezi için önemli bir enzim olan P450, normalde çoğu memelinin beyninde ve üreme organlarında eksprese edilir.[17] Bununla birlikte, primatlarda, bir LTR destekli transkripsiyonel varyant, plasentaya ekspresyon sağlar ve gebelik sırasında östrojen seviyelerini kontrol etmekten sorumludur.[17] Ayrıca, nöronal apoptoz inhibe edici protein Normalde yaygın olan (NAIP), testis ve prostata ekspresyon sağlayan bir promoter olarak hareket eden HERV-P ailesinden bir LTR'ye sahiptir.[26] Nitrik oksit sentaz 3 (NOS3), interlökin-2 reseptörü B (IL2RB) ve östrojen sentezinin başka bir aracı olan HSD17B1 gibi diğer proteinler de alternatif olarak plasental ekspresyon sağlayan LTR'ler tarafından düzenlenir, ancak bunların spesifik işlevleri henüz bilinmemektedir. .[22][27] Yüksek dereceli üreme ifadesinin, endojenize edildikleri yöntemin bir sonraki etkisi olduğu düşünülmektedir; bununla birlikte, bu aynı zamanda germ hattı dokularında DNA metilasyon eksikliğinden de kaynaklanıyor olabilir.[22]
Plasental protein ekspresyonunun en iyi karakterize edilen örneği, alternatif olarak teşvik edilen bir konakçı genden değil, bir retroviral proteinin tam bir ortak seçeneğinden gelir. Virionun konakçı hücreye girişinde rol oynayan retroviral füzojenik env proteinleri, memelinin gelişimi üzerinde önemli bir etkiye sahip olmuştur. plasenta. Memelilerde, bozulmamış env proteinleri sinsitinler oluşumundan ve işlevinden sorumludur sinsitiyotrofoblastlar.[13] Bu çok çekirdekli hücreler temel olarak besin alışverişini sürdürmekten ve fetüsü annenin bağışıklık sisteminden ayırmaktan sorumludur.[13] Bu proteinlerin bu işlev için seçilmesinin ve sabitlenmesinin, gelişiminde kritik bir rol oynadığı öne sürülmüştür. canlılık.[28]
Ek olarak, ERV'lerin ve ilgili LTR'lerin sokulması, kromozomlar arası lokuslarda viral sekanslar arasındaki rekombinasyona bağlı olarak kromozomal yeniden düzenlemeyi indükleme potansiyeline sahiptir. Bu yeniden düzenlemelerin, genom plastisitesine büyük ölçüde katkıda bulunan ve gen fonksiyonunun dinamiğini dramatik bir şekilde değiştiren gen duplikasyonlarını ve delesyonlarını indüklediği gösterilmiştir.[29] Dahası, genel olarak retroelementler, işlevi büyük ölçüde strese ve dış uyaranlara yanıtla ilgili olan, hızla gelişen, memeliye özgü gen ailelerinde büyük ölçüde yaygındır.[17] Özellikle hem insan sınıf I ve sınıf II MHC genler, diğer çoklu lokus gen ailelerine kıyasla yüksek yoğunlukta HERV elementine sahiptir.[24] HERV'lerin, HLA sınıf 1 gen ailesini oluşturan geniş ölçüde kopyalanmış duplikon blokların oluşumuna katkıda bulunduğu gösterilmiştir.[30] Daha spesifik olarak, HERV'ler esas olarak bu bloklar arasındaki kırılma noktalarının içinde ve arasındaki bölgeleri işgal eder, bu da tipik olarak eşit olmayan çapraz geçiş ile ilişkili olan önemli çoğaltma ve silme olaylarının oluşumlarını kolaylaştırdığını gösterir.[31] İmmünohaplotipler olarak miras alınan bu blokların üretimi, insanlara diğer primatlara göre bir avantaj aşılamış olabilecek çok çeşitli antijenlere karşı koruyucu bir polimorfizm görevi görür.[30]
Özelliği Plasentalar farklı türler arasında çok evrimsel olarak farklı organlar olmasının, yardımcı seçenek ERV geliştiricilerinin. Kodlayan genlerdeki mutasyonlar yerine düzenleyici mutasyonlar hormonlar ve büyüme faktörleri, özellikle hormon ve büyüme faktörü genlerinin çoğu plasenta gelişimi sırasında değil, gebeliğe yanıt olarak ifade edildiğinden, plasental morfolojinin bilinen evrimini destekler. Araştırmacılar, yakından ilişkili iki tür olan sıçan ve fare arasındaki plasental gelişimin düzenleyici manzarasını inceledi. Bu, farenin tüm düzenleyici unsurlarını haritalayarak yapıldı. trofoblast kök hücreler (TSC'ler) ve bunları kendi ortologlar fare TSC'lerinde. TSC'ler fetal plasentada gelişen ilk hücreleri yansıttıkları için gözlendi. Somut benzerliklerinden bağımsız olarak, güçlendirici ve bastırılmış bölgeler çoğunlukla türe özgüdür. Bununla birlikte, çoğu promoter sekansı, fare ve sıçan arasında korunmuştur. Araştırmacılar çalışmalarının sonucunda, ERV'lerin plasenta büyümesi aracılığıyla türe özgü plasental evrimi etkilediğini öne sürdüler. immünosupresyon, ve hücre füzyonu.[32]
Hücresel mekanizmalardan yararlanan başka bir ERV örneği s53, bir tümör baskılayıcı gen (TSG). DNA hasarı ve hücresel stres, hücreye neden olan p53 yolunu indükler. apoptoz. Sekanslama ile birlikte kromatin immüno-çökeltme kullanılarak, tüm p53 bağlama bölgelerinin yüzde otuzu, birkaç primat spesifik ERV ailesinin kopyaları içinde konumlandırıldı. Bir çalışma, bunun retrovirüslere fayda sağladığını ileri sürdü çünkü p53'ün mekanizması, viral RNA'nın konakçı hücreden çıkışına yol açan hızlı bir transkripsiyon indüksiyonu sağlar.[5]
Son olarak, ERV'lerin veya ERV elemanlarının konakçı DNA'nın genik bölgelerine sokulması veya bunların transkripsiyonel varyantlarının aşırı ekspresyonu, pozitif olanlardan çok daha fazla zararlı etkiler üretme potansiyeline sahiptir. Genomdaki görünümleri bir konak-parazit birlikte evrimsel dinamik baskılayıcı genlerin çoğalmasını ve genişlemesini çoğaltan. Bunun en net örneği, tandemin hızlı çoğaltılması ve çoğalmasıdır. çinko parmak memeli genomlarındaki genler. Çinko parmak genleri, özellikle bir KRAB alanı içerenler, omurgalı genomlarında yüksek kopya sayısında bulunur ve bunların işlevleri, transkripsiyonel rollerle sınırlıdır.[33] Bununla birlikte, memelilerde, bu genlerin çeşitlendirilmesinin, yeni retroviral dizilere veya bunların transkripsiyonlarını bastırmak için endojen kopyalarına yanıt olarak çoklu çoğaltma ve sabitleme olaylarına bağlı olduğu gösterilmiştir.[18]
Hastalıktaki rolü
Omurgalı genomlarında meydana gelen ERV'lerin çoğu eskidir, mutasyonla etkisiz hale getirilmiştir ve genetik sabitleme ev sahibi türlerinde. Bu nedenlerden ötürü, olağandışı durumlar haricinde, ev sahiplerini olumsuz etkileme olasılıkları son derece düşüktür. Bununla birlikte, kuşlar ve fareler, kediler ve hayvanlar da dahil olmak üzere insan dışı memeli türlerinde yapılan çalışmalardan açıkça anlaşılmaktadır. Koalas, daha genç (yani daha yakın zamanda entegre olmuş) ERV'ler hastalıkla ilişkilendirilebilir. Memelilerin genomundaki aktif ERV'lerin sayısı, vücut büyüklükleriyle negatif olarak ilişkilidir ve bu da Peto'nun paradoksu kanser patogenezi yoluyla.[34] Bu, araştırmacıların, insan kanserinin çeşitli biçimlerinde ERV'ler için bir rol önermesine ve Otoimmün rahatsızlığı kesin kanıtlar olmamasına rağmen.[35][36][37][38]
Nörolojik bozukluklar
İnsanlarda, ERV'lerin dahil olduğu ileri sürülmüştür. multipl Skleroz (HANIM). MS ve MS arasında belirli bir ilişki ERVWE1 veya bir ERV eklemesinden türetilen "sinsitin" geni, hastalığı olan hastalarda bir "MS ile ilişkili retrovirüs" (MSRV) mevcudiyeti ile birlikte rapor edilmiştir.[39][40] İnsan ERV'leri (HERV'ler) de dahil edilmiştir ALS[41] ve bağımlılık.[42][43][44]
2004 yılında, HERV'lere karşı antikorların, sera olanların şizofreni. Ek olarak, Beyin omurilik sıvısı Yakın zamanda başlayan şizofreni hastalarının içinde retroviral belirteç seviyeleri vardı, ters transkriptaz, kontrol deneklerinden dört kat daha fazla.[45] Araştırmacılar, HERV'ler ve şizofreni arasındaki olası bir bağlantıya, ek bir tetikleme olasılığı ile bakmaya devam ediyor. şizofreniye neden olan enfeksiyon.[46]
Bağışıklık
ERV'lerin sadece hastalığa neden olan ilişkiler yoluyla değil, aynı zamanda bağışıklık yoluyla da hastalıkla ilişkili olduğu bulunmuştur. Uzun terminal tekrarlarında (LTR'ler) ERV'lerin frekansı, viral transkripsiyonu ve replikasyonu teşvik eden bağışıklık sinyal yollarından yararlanmak için muhtemelen viral adaptasyonlarla ilişkilidir. 2016 yılında yapılan bir çalışma, bir konakçıya entegre edilmiş antik viral DNA'nın, aşağıdakilerin neden olduğu gen düzenleme ağları aracılığıyla faydasını araştırdı. interferonlar, doğuştan gelen bir bağışıklık dalı.[47] Bunlar sitokinler Viral enfeksiyona ilk yanıt verenlerdir ve aynı zamanda kötü huylu hücreler için immüno-gözetimde önemlidir.[48] ERV'lerin cis-düzenleyici unsurlar olarak hareket ettiği tahmin edilmektedir, ancak bunun belirli fizyolojik işlevler için uyarlanabilir sonuçlarının çoğu hala bilinmemektedir. ERV'lerin, özellikle insan interferon yanıtının düzenlenmesindeki genel rolünü destekleyen veriler vardır. interferon-gama (IFNG). Örneğin, interferon ile uyarılan genlerin, sinyal dönüştürücü ve transkripsiyon aktivatörü (STAT1) ve / veya tarafından bağlanan ERV'ler ile büyük ölçüde zenginleştirildiği bulunmuştur. İnterferon düzenleyici faktör (IRF1) içinde CD14 + makrofajlar.[1]
HERV'ler ayrıca insanı şekillendiren çeşitli roller oynar. doğuştan gelen bağışıklık yanıt, bazı diziler sistemi etkinleştirirken diğerleri bastırıyor. Ayrıca eksojen retroviral enfeksiyonlardan da koruyabilirler: virüs benzeri transkriptler Model tanıma reseptörleri ve proteinler aktif retrovirüslere müdahale edebilir. HERV-K'den (HML2) bir gag proteininin HIV Gag ile karıştığı ve bunun sonucunda HIV kapsid oluşumunu bozduğu gösterilmiştir.[49]
Gen düzenlemesi
Önerilen başka bir fikir, aynı aileden ERV'lerin birden fazla genin aynı düzenleme ağına dahil edilmesinde rol oynadığı yönündeydi. MER41 elemanlarının STAT1 bağlanma bölgelerinin yakınında bulunan genlere fazladan düzenleyici geliştirme sağladığı bulundu.[1]
Tıpta rolü
Domuz endojen retrovirüsü
İnsanlar için domuz endojen retrovirüsler (PERV'ler), ksenotransplantasyonda domuz dokuları ve organları kullanıldığında, canlı hücrelerin, dokuların ve organların bir tür organizmadan farklı türlerden bir organizmaya nakledilmesinde endişe yaratır. Domuzlar genellikle pratik, mali, güvenlik ve etik nedenlerle insan organ hastalıklarını tedavi etmek için en uygun donörler olsa da,[47] Daha önce PERV'ler, konakçı genomuna entegre olma ve yavrulara geçme gibi viral doğası nedeniyle Dr. George Church'ün laboratuvarı domuz genomundan 62 retrovirüsün tamamını çıkardığı 2017 yılına kadar domuzlardan çıkarılamıyordu.[50] Türler arası bulaşmanın sonuçları keşfedilmemiş olarak kalır ve çok tehlikeli potansiyele sahiptir.[51]
Araştırmacılar, özellikle bağışıklığı baskılanmış kişilerde PERV'ler tarafından insan dokularına enfeksiyonun çok olası olduğunu belirtti. Bağışıklık sistemi baskılanmış bir durum potansiyel olarak viral DNA'nın daha hızlı ve inatçı bir replikasyonuna izin verebilir ve daha sonra insandan insana geçişe adapte olma konusunda daha az zorluk çekebilir. Donör organ / dokusunda bulunan bilinen enfeksiyöz patojenler, patojen içermeyen sürülerin yetiştirilmesiyle ortadan kaldırılabilmesine rağmen, donörde bilinmeyen retrovirüsler bulunabilir. Bu retrovirüsler genellikle donörde gizli ve asemptomatiktir, ancak alıcıda aktif hale gelebilir. İnsan hücrelerinde enfekte olabilen ve çoğalabilen bazı endojen virüs örnekleri, babunlardan (BaEV), kedilerden (RD114) ve farelerden gelmektedir.[47]
Üç farklı PERV sınıfı vardır: PERV-A, PERV-B ve PERV-C. PERV-A ve PERV-B politropik ve insan hücrelerini in vitro enfekte edebilirken, PERV-C ekotropik ve insan hücrelerinde çoğalmaz. Sınıflar arasındaki temel farklar, env protein ve her sınıfın replikasyonunu etkileyen uzun terminal tekrarları (LTR'ler). PERV-A ve PERV-B, içinde tekrarları olan LTR'leri görüntüler. U3 bölge. Bununla birlikte, PERV-A ve PERV-C, tekrarlanmayan LTR'leri gösterir. Araştırmacılar, kültürdeki PERV'lerin, bir konakçı hücrenin gerçekleştirebileceği en iyi replikasyon performansıyla eşleşmek için LTR'lerinin tekrar yapısına aktif olarak adapte olduklarını buldular. Çalışmalarının sonunda araştırmacılar, tekrarlanmayan PERV LTR'nin tekrar barındıran LTR'den evrimleştiği sonucuna vardılar. Bu büyük olasılıkla insersiyonel mutasyondan kaynaklanmıştı ve LTR ve LTR'deki verilerin kullanımı ile kanıtlandı. env/ Env. Tekrarlanmayan LTR'lerin oluşumunun, virüsün eksojen bir yaşam tarzından içsel bir yaşam tarzına değişen bir adaptasyon sürecini yansıtabileceği düşünülmektedir.[52]
1999'da gerçekleştirilen bir klinik çalışma çalışması, farklı canlı domuz dokuları ile tedavi edilen ve PERV dizilerinin amplifikasyonu için PCR için yeterli miktarda DNA bulunan hastaların% 97'sinde kalıcı bir PERV enfeksiyonu kanıtı gözlemlemeyen 160 hastayı örnekledi. Bu çalışma, geriye dönük çalışmaların gerçek enfeksiyon insidansını veya ilişkili klinik semptomları bulmak için sınırlı olduğunu belirtti. Muhtemel çapraz türler PERV aktarımının daha eksiksiz ve ayrıntılı bir değerlendirmesini ve PERV'nin karşılaştırılmasını sağlayacak, yakından izlenen ileriye dönük denemelerin kullanılmasını önerdi.[53]
İnsan endojen retrovirüsleri
İnsan endojen retrovirüsleri (HERV), insan genomu yaklaşık 98.000 ERV öğesi ve% 5-8'i oluşturan fragmanlarla.[1] 2005 yılında yayınlanan bir araştırmaya göre, replikasyon yapabilen HERV tanımlanmamıştır; hepsi kusurlu görünüyordu, büyük delesyonlar veya anlamsız mutasyonlar içeriyordu. Bunun nedeni, çoğu HERV'nin, ilk olarak milyonlarca yıl önce entegre olmuş orijinal virüslerin izleri olmasıdır. HERV entegrasyonlarının bir analizi, 100.000 Genom Projesi.[54]
İnsan endojen retrovirüsleri, birkaç farklı deney kullanılarak tesadüfen keşfedildi. İnsan genomik kitaplıkları, hayvan retrovirüslerinden alınan sondalar kullanılarak düşük sertlik koşulları altında tarandı, bu da çeşitli aileleri temsil eden, kusurlu olsa da birçok provirüsün izolasyonuna ve karakterizasyonuna izin verdi. Başka bir deney, viral primer bağlanma bölgelerine homolojiye sahip oligonükleotitlere bağlıydı.[1]
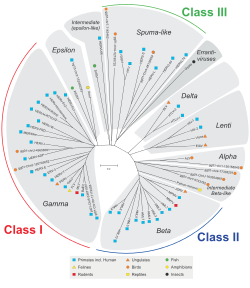
HERV'ler, hayvan retrovirüslerine olan homolojilerine göre sınıflandırılır. Sınıf I'e ait olan aileler, memelilere sırayla benzerdir. Gammaretrovirüsler (C tipi) ve Epsilonretrovirüsler (E Tipi). Sınıf II'ye ait olan aileler, memelilere homoloji gösterir. Betaretrovirüsler (B Tipi) ve Deltaretrovirüsler (D Tipi). Sınıf III'e ait olan aileler, köpüklü virüsler. Tüm sınıflar için, homolojiler, şaka, pol, ve env gen, bir üst aile. Var olduğu bilinen daha fazla Sınıf I ailesi var.[1][9] Ailelerin kendileri, eksojen bir retrovirüse, priming tRNA'ya (HERV-W, K) veya bazı komşu genlere (HERV-ADP), küme sayısına (HERV-S71) dayalı bir adlandırma karışımı ile daha az tekdüze bir şekilde adlandırılır. veya bazı amino asit motifleri (HERV-FRD). Önerilen bir isimlendirme, bazen parafiletik standartları temizlemeyi amaçlamaktadır.[55]
HERV'lerin insan genomunda nasıl sabitlendiğine dair iki öneri var. Birincisi, insan evrimi sırasında bir zaman, HERV'nin eksojen öncülerinin kendilerini germ hattı hücrelerine soktuklarını ve daha sonra konakçının hücresel mekanizmalarını kullanarak ve istismar ederek konağın genleriyle birlikte kopyaladıklarını varsayar. Farklı genomik yapıları nedeniyle HERV'ler, retroviral DNA'nın yaygın bir dağılımına yol açan birçok amplifikasyon ve transpozisyon turuna tabi tutuldu. İkinci hipotez, retro unsurların daha basit yapılandırılmış atalardan sürekli evrim geçirdiğini iddia ediyor.[1]
Bununla birlikte, bir virüs ailesi, insanlar ve şempanzeler. Bu aile, HERV-K (HML2), HERV elementlerinin% 1'inden daha azını oluşturur ancak en çok çalışılanlardan biridir. Son birkaç yüz bin yıldır aktif olduğuna dair göstergeler var, örneğin bazı insan bireyler diğerlerinden daha fazla HML2 kopyası taşıyor.[56] Geleneksel olarak, HERV'lerin yaş tahminleri 5 've 3' karşılaştırılarak gerçekleştirilir. LTR HERV'nin; ancak bu yöntem yalnızca tam uzunluktaki HERV'ler için geçerlidir. Kesitsel tarihleme adı verilen yeni bir yöntem,[57] HERV eklemelerinin yaşlarını tahmin etmek için tek bir LTR içindeki varyasyonları kullanır. Bu yöntem, HERV yaşlarının tahmin edilmesinde daha kesindir ve herhangi bir HERV eklemesi için kullanılabilir. Kesitsel tarihleme, HERV-K (HML2), HERV-K106 ve HERV-K116'nın iki üyesinin son 800.000 yılda aktif olduğunu ve HERV-K106'nın 150.000 yıl önce modern insanları enfekte etmiş olabileceğini öne sürmek için kullanılmıştır.[58] Bununla birlikte, HERV-K (HML2) ailesinin bilinen bulaşıcı üyelerinin yokluğu ve yayınlanan insan genom dizisinde tam bir kodlama potansiyeline sahip elemanların olmaması, bazılarına, ailenin şu anda daha az aktif olma olasılığının düşük olduğunu göstermektedir. 2006 ve 2007'de, Fransa ve ABD'de bağımsız olarak çalışan araştırmacılar, HERV-K'nin (HML2) işlevsel versiyonlarını yeniden oluşturdu.[59][60]
MER41.AIM2, yabancı sitozolik DNA sensörünü kodlayan AIM2'nin (Melanom 2'de Yoktur) transkripsiyonunu düzenleyen bir HERV'dir. Bu, AIM2 için bir bağlayıcı site görevi görür, yani AIM2'nin transkripsiyonu için gerekli olduğu anlamına gelir. Araştırmacılar, bunu, CRISPR / Cas9 kullanarak HeLa hücrelerinde MER41.AIM2'yi silerek gösterdiler ve bu, modifiye edilmiş HeLa hücrelerinde saptanamayan bir AIM2 transkript seviyesine yol açtı. Halen MER41.AIM2 ERV'yi içeren kontrol hücreleri, normal miktarlarda AIM2 transkripti ile gözlendi. Bağışıklık açısından araştırmacılar, MER41.AIM2'nin enfeksiyona enflamatuar bir yanıt için gerekli olduğu sonucuna vardı.[61]
İmmünolojik çalışmalar, bazı kanıtlar göstermiştir. T hücresi HIV ile enfekte kişilerde HERV'lere karşı bağışıklık tepkileri.[62] HIV'in HIV ile enfekte olmuş hücrelerde HERV ekspresyonunu indüklediği hipotezi, HERV antijenlerini hedefleyen bir aşının HIV ile enfekte olmuş hücreleri spesifik olarak ortadan kaldırabileceği önerisine yol açtı. Bu yeni yaklaşımın potansiyel avantajı, HERV antijenlerini HIV ile enfekte olmuş hücrelerin vekil markörleri olarak kullanarak, kötü şöhretli çeşitli ve hızlı mutasyona uğramış HIV antijenlerini doğrudan hedeflemenin doğasında bulunan güçlüğü aşabilmesidir.[62]
Hala bozulmamış açık okuma çerçevelerine sahip birkaç insan endojen retrovirüs sınıfı vardır. Örneğin biyolojik olarak aktif bir HERV ailesi olan HERV-K'nin ekspresyonu, plasentada bulunan proteinleri üretir. Ayrıca, zarf genlerinin ifadesi HERV-W (ERVW-1 )ve HERV-FRD (ERVFRD-1 ), hücre-hücre füzyonunu indükleyerek plasentogenez sırasında sinsitiyotrofoblast hücre tabakasının üretimi için önemli olan sinsitinleri üretir.[63] HUGO Gen İsimlendirme Komitesi (HGNC), kopyalanmış insan ERV'leri için gen sembollerini onaylar.[64]
ERV'leri karakterize etmek için teknikler
Tüm genom dizileme
Örnek: Çin doğumlu domuz ERV (PERV) minipig izolatı, PERV-A-BM, genetik varyasyonunu ve evrimini anlamak için tamamen ve farklı ırklar ve hücre soyları ile birlikte dizildi. Gözlemlenen nükleotid ikamelerinin sayısı ve farklı genom dizileri arasında, araştırmacıların, PERV-A-BM'nin konakçı genomuna entegre edildiğine dair tahmini bir yaş belirlemesine yardımcı oldu; bu, Avrupa doğumlu domuz izolatlarından daha önce evrimsel bir yaşta olduğu bulundu.[51]
Sıralama ile kromatin immüno-çökeltme (ChIP-seq)
Bu teknik, DNA proteinleri, bastırılmış bölgeler ve trimetilasyon için bağlanma yerleri olan promoterlerin ve geliştiricilerin göstergesi olan histon işaretlerini bulmak için kullanılır.[32] DNA metilasyonunun fare somatik hücrelerinde ERV'lerin susturulmasını sürdürmek için hayati olduğu gösterilmiştir, histon işaretleri ise aynı amaç için hayati önem taşımaktadır. embriyonik kök hücreleri (ESC'ler) ve erken embriyogenez.[5]
Başvurular
Filogeniler inşa etmek
Çoğu HERV'nin hiçbir işlevi olmadığından, seçici olarak nötr olduklarından ve primat genomlarında çok bol olduklarından, bağlantı analizi için kolayca filogenetik belirteçler olarak hizmet ederler. Entegrasyon bölgesi polimorfizmlerini veya ortologların gelişen, proviral, nükleotid dizilerini karşılaştırarak bunlardan yararlanılabilir. Entegrasyonun ne zaman gerçekleştiğini tahmin etmek için araştırmacılar, her bir belirli lokustaki moleküler evrim oranını bulmak için her filogenetik ağaçtan mesafeleri kullandılar. ERV'lerin birçok tür genomu (yani bitkiler, böcekler, yumuşakçalar, balıklar, kemirgenler, evcil hayvanlar ve çiftlik hayvanları) açısından zengin olması da faydalıdır çünkü uygulaması çeşitli filogenetik soruları yanıtlamak için kullanılabilir.[7]
Provirüs yaşının ve tür ayrımı olaylarının zaman noktalarının belirlenmesi
Bu, farklı evrim dönemlerinden farklı HERV'leri karşılaştırarak gerçekleştirilir. Örneğin, bu çalışma, insanlardan maymunlara ve maymunlara kadar değişen farklı hominoidler için yapıldı. Mevcut büyük çeşitlilik nedeniyle bunu PERV ile yapmak zordur.[52]
Daha fazla araştırma
Epigenetik değişkenlik
Araştırmacılar bireysel epigenomları analiz edebilir ve transkriptomlar epigenetik salım yoluyla ve bunların insan hastalıkları ile potansiyel ilişkilerini ve gen düzenleyici ağların özelliklerini keşfetme yoluyla uykuda yer değiştirebilir öğelerin yeniden aktivasyonunu incelemek.[5]
Ksenotransplantasyonun immünolojik sorunları
Üstesinden gelmenin etkili bir yolu hakkında çok az şey biliniyor hiperakut reddi (HAR), donör epitelinde galaktosil-alfa1-3galatosil (alfa-Gal) antijenlerini tanıyan ksenoreaktif antikorlar tarafından başlatılan tamamlayıcı aktivasyonunu takip eder.[47]
Gen terapisinde HERV'lerin risk faktörleri
Retrovirüsler birbirleriyle ve diğer endojen DNA dizileriyle yeniden birleşebildikleri için, varsa HERV'lerin neden olabileceği potansiyel riskleri keşfetmek gen terapisi için faydalı olacaktır. Ayrıca HERV'lerin bu yeniden birleşme yeteneği, retroviral vektörlere HERV sekansları dahil edilerek sahaya yönelik entegrasyon için manipüle edilebilir.[1]
HERV gen ifadesi
Araştırmacılar, HERV genleri tarafından kodlanan RNA ve proteinlerin, hücre fizyolojisinde ve patolojik koşullarda varsayılan işlev için araştırılmaya devam edilmesi gerektiğine inanıyor. Sentezlenen proteinlerin biyolojik önemini daha derinlemesine tanımlamak için bunu incelemek mantıklı olacaktır.[1]
Ayrıca bakınız
- Kuş sarkomu lökoz virüsü (ASLV)
- Endojen viral eleman
- ERV3
- HERV-FRD
- Yatay gen transferi
- Jaagsiekte koyun retrovirüsü (JSRV)
- Koala retrovirüsü (KoRV)
- Fare meme tümörü virüsü (MMTV)
- Murin lösemi virüsü (MLV) ve ksenotropik murin lösemi virüsü ile ilgili virüs (XMRV)
- Paleoviroloji
Referanslar
- ^ a b c d e f g h ben j Belshaw R, Pereira V, Katzourakis A, Talbot G, Paces J, Burt A, Tristem M (Nisan 2004). "İnsan genomunun endojen retrovirüsler tarafından uzun vadeli yeniden enfeksiyonu". Proc Natl Acad Sci ABD. 101 (14): 4894–99. Bibcode:2004PNAS..101.4894B. doi:10.1073 / pnas.0307800101. PMC 387345. PMID 15044706.
- ^ Nelson PN, Hooley P, Roden D, Davari Ejtehadi H, Rylance P, Warren P, vd. (Ekim 2004). "İnsan endojen retrovirüsleri: potansiyeli olan yer değiştirebilir öğeler mi?". Klinik ve Deneysel İmmünoloji. 138 (1): 1–9. doi:10.1111 / j.1365-2249.2004.02592.x. PMC 1809191. PMID 15373898.
- ^ Khodosevich K, Lebedev Y, Sverdlov E (Ekim 2002). "Endojen retrovirüsler ve insan evrimi". Karşılaştırmalı ve Fonksiyonel Genomik. 3 (6): 494–98. doi:10.1002 / cfg.216. PMC 2448423. PMID 18629260.
- ^ Kim FJ, Battini JL, Manel N, Sitbon M (Ocak 2004). "Omurgalı retrovirüslerinin ortaya çıkışı ve zarf yakalama". Viroloji. 318 (1): 183–91. doi:10.1016 / j.virol.2003.09.026. PMID 14972546.
- ^ a b c d Rebollo R, Romanca MT, Mager DL (2012-01-01). "Değiştirilebilir elemanlar: konakçı genler için bol ve doğal bir düzenleyici sekans kaynağı". Genetik Yıllık İnceleme. 46 (1): 21–42. doi:10.1146 / annurev-genet-110711-155621. PMID 22905872.
- ^ Cotton, J. (2001). "Retotranspozonlardan retrovirüsler". Genom Biyolojisi. 2 (2): 6. doi:10.1186 / gb-2001-2-2-raporlar0006.
Görünüşe göre, nonviral retrotranspozondan retrovirüse geçiş bağımsız olarak en az sekiz kez meydana gelmiştir ve bulaşıcı yetenekten sorumlu zarf geninin kaynağı artık bu örneklerin en az dördünde bir virüse kadar izlenebilmektedir. Bu şunu önerir potansiyel olarak, herhangi bir LTR retrotranspozonu bir virüs olabilir mevcut viral genlerin edinilmesi yoluyla.
- ^ a b c Johnson WE, Coffin JM (Ağustos 1999). "Antik retrovirüs dizilerinden primat filogenileri oluşturmak". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 96 (18): 10254–60. Bibcode:1999PNAS ... 9610254J. doi:10.1073 / pnas.96.18.10254. PMC 17875. PMID 10468595.
- ^ Fujiwara T, Mizuuchi K (Ağustos 1988). "Retroviral DNA entegrasyonu: bir entegrasyon ara ürününün yapısı". Hücre. 54 (4): 497–504. doi:10.1016/0092-8674(88)90071-2. PMID 3401925.
- ^ a b Vargiu, L; Rodriguez-Tomé, P; Sperber, GİT; Cadeddu, M; Grandi, N; Blikstad, V; Tramontano, E; Blomberg, J (22 Ocak 2016). "İnsan endojen retrovirüslerinin sınıflandırılması ve karakterizasyonu; mozaik formlar yaygındır". Retroviroloji. 13: 7. doi:10.1186 / s12977-015-0232-y. PMC 4724089. PMID 26800882.
- ^ Magiorkinis, G; Gifford, RJ; Katzourakis, A; De Ranter, J; Belshaw, R (8 Mayıs 2012). "Zarfsız endojen retrovirüsler, genomik süper yayıcılardır". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 109 (19): 7385–90. doi:10.1073 / pnas.1200913109. PMC 3358877. PMID 22529376.
- ^ a b Li J, Akagi K, Hu Y, Trivett AL, Hlynialuk CJ, Swing DA, Volfovsky N, Morgan TC, Golubeva Y, Stephens RM, Smith DE, Symer DE (Mart 2012). "Fare endojen retrovirüsleri, belli bir mesafeden erken transkripsiyonel sonlandırmayı tetikleyebilir". Genom Res. 22 (5): 870–84. doi:10.1101 / gr.130740.111. PMC 3337433. PMID 22367191.
- ^ Spencer TE, Palmarini M (2012). "Koyunların endojen retrovirüsleri: gelişen bir geviş getiren genomuna fizyolojik adaptasyonu anlamak için bir model sistem". J Reprod Dev. 58 (1): 33–37. doi:10.1262 / jrd.2011-026. PMID 22450282.
- ^ a b c Siyah SG, Arnaud F, Palmarini M, Spencer TE (2010). "Trofoblast Farklılaşmasında ve Plasental Gelişimde Endojen Retrovirüsler". American Journal of Reproductive Immunology. 64 (4): 255–64. doi:10.1111 / j.1600-0897.2010.00860.x. PMC 4198168. PMID 20528833.
- ^ Ryan FP (Aralık 2004). "Sağlık ve hastalıkta insan endojen retrovirüsleri: simbiyotik bir bakış açısı". Kraliyet Tıp Derneği Dergisi. 97 (12): 560–65. doi:10.1258 / jrsm.97.12.560. PMC 1079666. PMID 15574851.
- ^ Ishak, Charles A .; De Carvalho, Daniel D. (2020). "Kanser Gelişiminde ve Tedavisinde Endojen Retroelementlerin Reaktivasyonu". Kanser Biyolojisinin Yıllık İncelemesi. 4: 159–176. doi:10.1146 / annurev-kanserbio-030419-033525.
- ^ Pi W, Zhu X, Wu M, Wang Y, Fulzele S, Eroglu A, Ling J, Tuan D (Temmuz 2010). "Bir intergenik retrotranspozonun uzun menzilli işlevi". PNAS. 107 (29): 12992–97. Bibcode:2010PNAS..10712992P. doi:10.1073 / pnas.1004139107. PMC 2919959. PMID 20615953.
- ^ a b c d e van de Lagemaat LN, Landry JR, Mager DL, Medstrand P (Ekim 2003). "Memelilerde yer değiştirebilir öğeler, özel işlevlere sahip genlerin düzenleyici çeşitliliğini ve çeşitlendirmesini teşvik eder". Trendler Genet. 19 (10): 530–36. doi:10.1016 / j.tig.2003.08.004. PMID 14550626.
- ^ a b Kovalskaya E, Buzdin A, Gogvadze E, Vinogradova T, Sverdlov E (2006). "Fonksiyonel insan endojen retroviral LTR transkripsiyon başlangıç siteleri, R ve U5 bölgeleri arasında yer alır". Viroloji. 346 (2): 373–78. doi:10.1016 / j.virol.2005.11.007. PMID 16337666.
- ^ Dunn CA, Romanish MT, Gutierrez LE, van de Lagemaat LN, Mager DL (2006). "İki yönlü bir endojen retrovirüs promotöründen iki insan geninin transkripsiyonu". Gen. 366 (2): 335–42. doi:10.1016 / j.gene.2005.09.003. PMID 16288839.
- ^ Gogvadze E, Stukacheva E, Buzdin A, Sverdlov E (2009). "Endojen retroviral eklemeler tarafından sağlanan transkripsiyonel aktivitenin insana özgü modülasyonu". J Virol. 83 (12): 6098–105. doi:10.1128 / JVI.00123-09. PMC 2687385. PMID 19339349.
- ^ Ting CN, Rosenberg MP, Snow CM, Samuelson LC, Meisler MH (1992). "Endojen retroviral sekanslar, bir insan tükürük amilaz geninin dokuya özgü ekspresyonu için gereklidir". Genes Dev. 6 (8): 1457–65. doi:10.1101 / gad.6.8.1457. PMID 1379564.
- ^ a b c Cohen CJ, Lock WM, Mager DL (2009). "İnsan genleri için promoter olarak endojen retroviral LTR'ler: kritik bir değerlendirme". Gen. 448 (2): 105–14. doi:10.1016 / j.gene.2009.06.020. PMID 19577618.
- ^ Bekpen C, Marques-Bonet T, Alkan C, Antonacci F, Leogrande MB, Ventura M, Kidd JM, Siswara P, Howard JC, Eichler EE (2009). "İnsan IRGM geninin ölümü ve dirilişi". PLOS Genet. 5 (3): e1000403. doi:10.1371 / journal.pgen.1000403. PMC 2644816. PMID 19266026.
- ^ a b Jern P, Coffin JM (2008). "Retrovirüslerin Konakçı Genom Fonksiyonu Üzerindeki Etkileri". Annu Rev Genet. 42: 709–32. doi:10.1146 / annurev.genet.42.110807.091501. PMID 18694346.
- ^ Oliver KR, Greene WK (2011). "Mobil DNA ve TE-Thrust hipotezi: primatlardan destekleyici kanıtlar". Mob DNA. 2 (1): 8. doi:10.1186/1759-8753-2-8. PMC 3123540. PMID 21627776.
- ^ Romanca MT, Lock WM, van de Lagemaat LN, Dunn CA, Mager DL (2007). "Memeli evrimi sırasında antiapoptotik lokus NAIP tarafından destekleyici olarak LTR retrotranspozonlarının tekrar tekrar görevlendirilmesi". PLOS Genet. 3 (1): e10. doi:10.1371 / dergi.pgen.0030010. PMC 1781489. PMID 17222062.
- ^ Huh JW, Ha HS, Kim DS, Kim HS (2008). "LTR'den türetilen NOS3'ün plasenta sınırlı ifadesi". Plasenta. 29 (7): 602–08. doi:10.1016 / j.placenta.2008.04.002. PMID 18474398.
- ^ Villarreal LP, Villareal LP (Şubat 1997). "On viruses, sex, and motherhood". J. Virol. 71 (2): 859–65. doi:10.1128/JVI.71.2.859-865.1997. PMC 191132. PMID 8995601.
- ^ Hughes JF, Coffin JM (December 2001). "Evidence for genomic rearrangements mediated by human endogenous retroviruses during primate evolution". Doğa Genetiği. 29 (4): 487–89. doi:10.1038/ng775. PMID 11704760. S2CID 20262212.
- ^ a b Dawkins R, Leelayuwat C, Gaudieri S, Tay G, Hui J, Cattley S, et al. (Şubat 1999). "Genomics of the major histocompatibility complex: haplotypes, duplication, retroviruses and disease". İmmünolojik İncelemeler. 167: 275–304. doi:10.1111/j.1600-065X.1999.tb01399.x. PMID 10319268.
- ^ Doxiadis GG, de Groot N, Bontrop RE (July 2008). "Impact of endogenous intronic retroviruses on major histocompatibility complex class II diversity and stability". Journal of Virology. 82 (13): 6667–77. doi:10.1128/JVI.00097-08. PMC 2447082. PMID 18448532.
- ^ a b Chuong EB, Rumi MA, Soares MJ, Baker JC (March 2013). "Endogenous retroviruses function as species-specific enhancer elements in the placenta". Doğa Genetiği. 45 (3): 325–29. doi:10.1038/ng.2553. PMC 3789077. PMID 23396136.
- ^ Thomas JH, Schneider S (2011). "Coevolution of retroelements and tandem zinc finger genes". Genom Res. 21 (11): 1800–12. doi:10.1101/gr.121749.111. PMC 3205565. PMID 21784874.
- ^ Katzourakis A, Magiorkinis G, Lim AG, Gupta S, Belshaw R, Gifford R (July 2014). "Larger mammalian body size leads to lower retroviral activity". PLOS Patojenleri. 10 (7): e1004214. doi:10.1371/journal.ppat.1004214. PMC 4102558. PMID 25033295.
- ^ Bannert N, Kurth R (October 2004). "Retroelements and the human genome: new perspectives on an old relation". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 101 Suppl 2 (Suppl 2): 14572–79. Bibcode:2004PNAS..10114572B. doi:10.1073/pnas.0404838101. PMC 521986. PMID 15310846.
- ^ Nelson PN, Carnegie PR, Martin J, Davari Ejtehadi H, Hooley P, Roden D, et al. (Şubat 2003). "Demystified. Human endogenous retroviruses". Moleküler Patoloji. 56 (1): 11–18. doi:10.1136/mp.56.1.11. PMC 1187282. PMID 12560456.
- ^ Singh SK (June 2007). "Endogenous retroviruses: suspects in the disease world". Geleceğin Mikrobiyolojisi. 2 (3): 269–75. doi:10.2217/17460913.2.3.269. PMID 17661701.
- ^ Magiorkinis G, Belshaw R, Katzourakis A (September 2013). "'There and back again': revisiting the pathophysiological roles of human endogenous retroviruses in the post-genomic era". Londra Kraliyet Cemiyeti'nin Felsefi İşlemleri. Seri B, Biyolojik Bilimler. 368 (1626): 20120504. doi:10.1098/rstb.2012.0504. PMC 3758188. PMID 23938753.
- ^ Mameli G, Astone V, Arru G, Marconi S, Lovato L, Serra C, et al. (Ocak 2007). "Brains and peripheral blood mononuclear cells of multiple sclerosis (MS) patients hyperexpress MS-associated retrovirus/HERV-W endogenous retrovirus, but not Human herpesvirus 6". Genel Viroloji Dergisi. 88 (Pt 1): 264–74. doi:10.1099/vir.0.81890-0. PMID 17170460.
- ^ Serra C, Mameli G, Arru G, Sotgiu S, Rosati G, Dolei A (December 2003). "In vitro modulation of the multiple sclerosis (MS)-associated retrovirus by cytokines: implications for MS pathogenesis". Nöroviroloji Dergisi. 9 (6): 637–43. doi:10.1080/714044485. PMID 14602576.
- ^ "Reactivated Virus May Contribute to ALS". 2016-01-23.
- ^ Karamitros T, Hurst T, Marchi E, Karamichali E, Georgopoulou U, Mentis A, et al. (Ekim 2018). "RASGRF2 is associated with intravenous drug abuse and modulates transcription in a cell-line model". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 115 (41): 10434–39. doi:10.1073/pnas.1811940115. PMC 6187174. PMID 30249655.
- ^ Rob Picheta. "Addiction may stem from ancient retrovirus, study says". CNN. Alındı 2019-10-13.
- ^ "An ancient virus may promote addiction in modern people". Ekonomist. 2018-10-02. ISSN 0013-0613. Alındı 2019-10-13.
- ^ Yolken R (June 2004). "Viruses and schizophrenia: a focus on herpes simplex virus". Uçuk. 11 (Suppl 2): 83A–88A. PMID 15319094.
- ^ Fox D (2010). "The Insanity Virus". Keşfedin. Alındı 2011-02-17.
- ^ a b c d Patience C, Takeuchi Y, Weiss RA (March 1997). "Infection of human cells by an endogenous retrovirus of pigs". Doğa Tıbbı. 3 (3): 282–86. doi:10.1038/nm0397-282. PMID 9055854.
- ^ Platanias LC (Mayıs 2005). "Tip-I- ve tip-II-interferon aracılı sinyalleşme mekanizmaları". Doğa Yorumları. İmmünoloji. 5 (5): 375–86. doi:10.1038 / nri1604. PMID 15864272.
- ^ Grandi, N; Tramontano, E (2018). "Human Endogenous Retroviruses Are Ancient Acquired Elements Still Shaping Innate Immune Responses". İmmünolojide Sınırlar. 9: 2039. doi:10.3389/fimmu.2018.02039. PMC 6139349. PMID 30250470.
- ^ Niu D, Wei HJ, Lin L, George H, Wang T, Lee IH, Zhao HY, Wang Y, Kan Y, Shrock E, Lesha E, Wang G, Luo Y, Qing Y, Jiao D, Zhao H, Zhou X, Wang S, Wei H, Güell M, Church GM, Yang L (September 2017). "Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9". Bilim. 357 (6357): 1303–07. Bibcode:2017Sci...357.1303N. doi:10.1126/science.aan4187. PMC 5813284. PMID 28798043.
- ^ a b Tang HB, Ouyang K, Rao GB, Ma L, Zhong H, Bai A, et al. (2016-01-01). "Characterization of Complete Genome Sequences of a Porcine Endogenous Retrovirus Isolated From China Bama Minipig Reveals an Evolutionary Time Earlier Than That of Isolates From European Minipigs". Nakil İşlemleri. 48 (1): 222–28. doi:10.1016/j.transproceed.2015.12.005. PMID 26915872.
- ^ a b Tönjes RR, Niebert M (November 2003). "Relative age of proviral porcine endogenous retrovirus sequences in Sus scrofa based on the molecular clock hypothesis". Journal of Virology. 77 (22): 12363–68. doi:10.1128/JVI.77.22.12363-12368.2003. PMC 254287. PMID 14581574.
- ^ Paradis K, Langford G, Long Z, Heneine W, Sandstrom P, Switzer WM, et al. (Ağustos 1999). "Search for cross-species transmission of porcine endogenous retrovirus in patients treated with living pig tissue. The XEN 111 Study Group". Bilim. 285 (5431): 1236–41. doi:10.1126/science.285.5431.1236. PMID 10455044.
- ^ "Genomics England › Integrated Pathogens and Mobile Elements GeCIP Domain". Alındı 2019-10-13.
- ^ Gifford, RJ; Blomberg, J; Coffin, JM; Fan, H; Heidmann, T; Mayer, J; Stoye, J; Tristem, M; Johnson, WE (28 August 2018). "Nomenclature for endogenous retrovirus (ERV) loci". Retroviroloji. 15 (1): 59. doi:10.1186/s12977-018-0442-1. PMC 6114882. PMID 30153831.
- ^ Belshaw R, Dawson AL, Woolven-Allen J, Redding J, Burt A, Tristem M (October 2005). "Genomewide Screening Reveals High Levels of Insertional Polymorphism in the Human Endogenous Retrovirus Family HERV-K(HML2): Implications for Present-Day Activity". J. Virol. 79 (19): 12507–14. doi:10.1128/JVI.79.19.12507-12514.2005. PMC 1211540. PMID 16160178.
- ^ Jha AR, Pillai SK, York VA, Sharp ER, Storm EC, Wachter DJ, Martin JN, Deeks SG, Rosenberg MG, Nixon DF, Garrison KE (August 2009). "Cross-Sectional Dating of Novel Haplotypes of HERV-K 113 and HERV-K 115 Indicate These Proviruses Originated in Africa before Homo sapiens". Mol Biol Evol. 26 (11): 2617–26. doi:10.1093/molbev/msp180. PMC 2760466. PMID 19666991.
- ^ Jha AR, Nixon DF, Rosenberg MG, Martin JN, Deeks SG, Hudson RR, Garrison KE, Pillai SK (May 2011). "Human Endogenous Retrovirus K106 (HERV-K106) Was Infectious after the Emergence of Anatomically Modern Humans". PLOS ONE. 6 (5): e20234. Bibcode:2011PLoSO...620234J. doi:10.1371/journal.pone.0020234. PMC 3102101. PMID 21633511.
- ^ Bieniasz, Paul; Lee, Young Nam (January 2007). "Reconstitution of an infectious human endogenous retrovirus". PLOS Pathog. 3 (1): e10. doi:10.1371/journal.ppat.0030010. PMC 1781480. PMID 17257061.
- ^ Dewannieux M, Harper F, Richaud A, Letzelter C, Ribet D, Pierron G, Heidmann T (December 2006). "Identification of an infectious progenitor for the multiple-copy HERV-K human endogenous retroelements". Genom Res. 16 (12): 1548–56. doi:10.1101/gr.5565706. PMC 1665638. PMID 17077319. Lay özeti – ScienceNOW Günlük Haberler.
- ^ Chuong EB, Elde NC, Feschotte C (March 2016). "Regulatory evolution of innate immunity through co-option of endogenous retroviruses". Bilim. 351 (6277): 1083–87. Bibcode:2016Sci...351.1083C. doi:10.1126/science.aad5497. PMC 4887275. PMID 26941318.
- ^ a b Garrison KE, Jones RB, Meiklejohn DA, Anwar N, Ndhlovu LC, Chapman JM, Erickson AL, Agrawal A, Spotts G, Hecht FM, Rakoff-Nahoum S, Lenz J, Ostrowski MA, Nixon DF (November 2007). "T cell responses to human endogenous retroviruses in HIV-1 infection". PLOS Pathog. 3 (11): e165. doi:10.1371/journal.ppat.0030165. PMC 2065876. PMID 17997601.
- ^ Morozov VA, Dao Thi VL, Denner J (2013). "The transmembrane protein of the human endogenous retrovirus – K (HERV-K) modulates cytokine release and gene expression". PLOS ONE. 8 (8): e70399. Bibcode:2013PLoSO...870399M. doi:10.1371/journal.pone.0070399. PMC 3737193. PMID 23950929.
- ^ Mayer J, Blomberg J, Seal RL (May 2011). "A revised nomenclature for transcribed human endogenous retroviral loci". Mobil DNA. 2 (1): 7. doi:10.1186/1759-8753-2-7. PMC 3113919. PMID 21542922.
daha fazla okuma
- Löwer R, Löwer J, Kurth R (May 1996). "The viruses in all of us: characteristics and biological significance of human endogenous retrovirus sequences". Proc Natl Acad Sci ABD. 93 (11): 5177–84. Bibcode:1996PNAS...93.5177L. doi:10.1073/pnas.93.11.5177. PMC 39218. PMID 8643549.
- Molès JP, Tesniere A, Guilhou JJ (Jul 2005). "A new endogenous retroviral sequence is expressed in skin of patients with psoriasis". Br J Dermatol. 153 (1): 83–89. doi:10.1111/j.1365-2133.2005.06555.x. PMID 16029331.
- Seifarth W, Frank O, Zeilfelder U, Spiess B, Greenwood AD, Hehlmann R, Leib-Mösch C (Jan 2005). "Comprehensive analysis of human endogenous retrovirus transcriptional activity in human tissues with a retrovirus-specific microarray". J. Virol. 79 (1): 341–52. doi:10.1128/JVI.79.1.341-352.2005. PMC 538696. PMID 15596828.
- Knerr I, Beinder E, Rascher W (Feb 2002). "Syncytin, a novel human endogenous retroviral gene in human placenta: evidence for its dysregulation in preeclampsia and HELLP syndrome". Am J Obstet Gynecol. 186 (2): 210–13. doi:10.1067/mob.2002.119636. PMID 11854637.
- Gifford R, Tristem M (May 2003). "The evolution, distribution and diversity of endogenous retroviruses" (PDF). Virüs Genleri. 26 (3): 291–315. doi:10.1023/A:1024455415443. PMID 12876457. Arşivlenen orijinal (PDF) 2013-02-15 tarihinde. Alındı 2008-01-16.
Dış bağlantılar
- Endogenous+Retroviruses ABD Ulusal Tıp Kütüphanesinde Tıbbi Konu Başlıkları (MeSH)
- HERVd – human endogenous retrovirus database