Dinozorların fizyolojisi - Physiology of dinosaurs
- Genel olarak dinozorlar sürüngenlerdir. Ancak ön not: Bu makalede "dinozor", "kuş olmayan dinozor" anlamına gelmektedir. monofiletik içindeki takson clade Dinosauria ve çoğu uzman, kuşları dinozorlar.
dinozorların fizyolojisi tarihsel olarak tartışmalı bir konu olmuştur, özellikle termoregülasyon. Son zamanlarda, dinozor hakkında birçok yeni kanıt getirildi. fizyoloji genel olarak, sadece metabolik sistemler ve termoregülasyon değil, aynı zamanda solunum ve kardiyovasküler sistemler de dahil.
Dinozor paleontolojisinin ilk yıllarında, yaygın olarak halsiz, hantal ve yayılan oldukları düşünülüyordu. Soğuk kanlı kertenkele. Bununla birlikte, 1870'lerden başlayarak batı Amerika Birleşik Devletleri'nde çok daha eksiksiz iskeletlerin keşfedilmesiyle, bilim adamları dinozor biyolojisi ve fizyolojisi hakkında daha bilinçli yorumlar yapabildiler. Edward Drinker Cope, rakibi Othniel Charles Marsh içinde Kemik Savaşları, en azından bazı dinozorları, iki dövüşün resminde görüldüğü gibi aktif ve çevik olarak öne sürdü. Laelaps onun yönetimi altında üretti Charles R. Şövalye.[1]
Paralel olarak, gelişimi Darwinci evrim ve keşifleri Archæopteryx ve Compsognathus, Led Thomas Henry Huxley dinozorların kuşlarla yakından ilişkili olduğunu öne sürmek.[2] Bu değerlendirmelere rağmen, dinozorların büyük sürüngenler imajı çoktan kök salmıştı.[1] ve onların çoğu yönü paleobiyoloji yirminci yüzyılın ilk yarısı için tipik olarak sürüngen olarak yorumlandı.[3] 1960'larda başlayarak ve Dinozor Rönesansı, dinozorların görüşleri ve fizyolojileri dramatik bir şekilde değişti. tüylü dinozorlar içinde Erken Kretase yaş Çin'deki mevduatlar, kuşlar gelişti son derece çevik Maniraptoran dinozorlar.
Çalışma tarihi
Dinozorların erken yorumları: 1820'lerden 1900'lerin başına


Dinozorların incelenmesi 1820'lerde İngiltere'de başladı. Alandaki öncüler, örneğin William Buckland, Gideon Mantell, ve Richard Owen, ilk, çok parçalı kalıntıları büyüklere ait olarak yorumladı dört ayaklı canavarlar.[4] Erken çalışmaları bugün Kristal Saray Dinozorları 1850'lerde inşa edilen ve bilinen dinozorları fil olarak sunan kertenkele sürüngenler gibi.[5] Bu sürüngen görünümlerine rağmen, Owen bu dinozorun kalp ve solunum sistemleri sürüngenden çok memeliye benziyordu.[4]
Değişen görüş ve dinozor rönesansı
1960'ların sonlarında, benzer fikirler yeniden ortaya çıktı. John Ostrom üzerinde çalışmak Deinonychus ve kuş evrimi.[6] Onun öğrencisi, Bob Bakker, değişen düşünceyi bir dizi makalede popüler hale getirdi. Dinozorların üstünlüğü 1968'de.[7] Bu yayınlarda, dinozorların sıcakkanlı ve aktif hayvanlar olduğunu, sürekli yüksek aktivite dönemlerini sürdürebileceğini şiddetle savundu. Yazılarının çoğunda Bakker, argümanlarını 19. yüzyılın sonlarında popüler olan fikirlerin yeniden canlanmasına yol açan yeni kanıtlar olarak çerçeveledi ve sıklıkla devam eden bir dinozor rönesansı. Davasını savunmak için çeşitli anatomik ve istatistiksel argümanlar kullandı.[8][9] metodolojisi bilim adamları arasında şiddetle tartışılan bir konu.[10]
Bu tartışmalar, nesli tükenmiş hayvanların paleobiyolojisini tespit etmek için yeni yöntemlere olan ilgiyi uyandırdı. kemik histolojisi, birçok dinozorun büyüme oranlarını belirlemek için başarıyla uygulanmıştır.
Bugün, genellikle dinozorların çoğunun veya belki de tümünün yaşayan sürüngenlerden daha yüksek metabolik hızlara sahip olduğu, ancak aynı zamanda durumun Bakker'in ilk önerdiğinden daha karmaşık ve çeşitli olduğu düşünülmektedir. Örneğin, daha küçük dinozorlar doğru olabilirken endotermler daha büyük formlar olabilirdi atalet homeotermleri,[11][12] ya da birçok dinozorun ara metabolik hızlara sahip olabileceği.[13]
Beslenme ve sindirim
En eski dinozorlar neredeyse kesinlikle yırtıcılardı ve dinozor olmayan en yakın akrabalarıyla birkaç yırtıcı özelliği paylaştılar. Lagosuchus dahil: makas gibi kapanan büyük, geniş açık çenelerde nispeten büyük, kavisli, bıçak benzeri dişler; nispeten küçük karınlar etoburlar için büyük sindirim sistemleri. Daha sonra avcı olarak kabul edilen dinozorlar bazen çok daha fazla büyüdüler, ancak aynı özellikleri korudular. Bu yırtıcılar yiyeceklerini çiğnemek yerine bütünüyle yuttu.[14]
Beslenme alışkanlıkları ornitomimozorlar ve Oviraptorozorlar bir muamma: bir yırtıcıdan evrimleşmelerine rağmen Theropod soyları, küçük çeneleri vardır ve tipik yırtıcı hayvanların bıçak benzeri dişlerinden yoksundurlar, ancak diyetlerine veya nasıl yediklerine ve sindirdiklerine dair hiçbir kanıt yoktur.[14]
Diğer dinozor gruplarının özellikleri, onların otçul olduklarını gösterir. Bu özellikler şunları içerir:
- Tüm dişlerin aynı anda buluşması için sadece hafifçe açılıp kapanan çeneler
- Büyük miktarda bitki örtüsünü barındırabilen ve onu bitki örtüsünü sindirmek için uzun süre depolayan büyük karınlar
- Muhtemelen içerdiği bağırsaklar Endosimbiyotik sindiren mikro organizmalar selüloz Bilinen hiçbir hayvan bu sert materyali doğrudan sindiremeyeceğinden[14]
Sauropodlar Otçul olan, dişleri ve çeneleri sadece bitkilerdeki yaprakları soymaya uygun göründüğünden yiyeceklerini çiğnemiyorlardı. Ornithischians ve otçullar da çeşitli yaklaşımlar sergilerler. Zırhlı Ankylosaurlar ve stegosaurlar küçük kafaları ve zayıf çeneleri ve dişleri vardı ve sauropodlarla aynı şekilde beslendikleri düşünülüyordu. pachycephalosaurs küçük kafaları, zayıf çeneleri ve dişleri vardı, ancak büyük sindirim sistemlerinden yoksun olmaları, yapraklardan daha besleyici olabilecek meyveler, tohumlar veya genç sürgünler gibi farklı bir diyete işaret ediyordu.[14]
Diğer taraftan, ornitopodlar gibi Hipsilofodon, Otçul kertenkele ve çeşitli hadrosaurlar bitki örtüsünü kesmek için azgın gagaları ve çiğneme için iyi uyarlanmış çeneleri ve dişleri vardı. Boynuzlu Ceratopsia'cılar benzer mekanizmalara sahipti.[14]
Çoğu zaman, en azından bazı dinozorların, şu adla bilinen yutulmuş taşlar kullandığı öne sürülmüştür. mide taşı, yiyeceklerini kas içinde öğüterek sindirime yardımcı olmak için taşlık ve bunun kuşlarla paylaştıkları bir özellik olduğunu. 2007 yılında Oliver Wings, bilimsel literatürdeki gastrolitlere yapılan atıfları gözden geçirdi ve "mide taşı" nın üzerinde mutabık kalınan ve objektif bir tanımın bulunmamasından başlayarak önemli bir kafa karışıklığı buldu. Yutulan sert taşların veya kumun, esas olarak tahılla beslenen ancak gerekli olmayabilen kuşlarda sindirime yardımcı olabileceğini ve yazın böcekleri ve kışın tahılları yiyen kuşların genellikle yazın taşlardan ve kumdan kurtulduklarını keşfetti. Gastrolitler genellikle Sauropod Bitki örtüsüyle beslenmeleri çok kapsamlı bir sindirim gerektiren dinozorlar, ancak Wings bu fikrin yanlış olduğu sonucuna vardı: Sauropod fosillerinin sadece küçük bir yüzdesinde mide taşları bulunur; bulundukları yerlerde, miktarlar çok azdır ve çoğu durumda taşlar, yiyecek öğütmede etkili olamayacak kadar yumuşaktır; bu gastrolitlerin çoğu son derece cilalıdır, ancak modern hayvanların yiyecekleri öğütmek için kullandıkları mide taşları aşınmayla pürüzlenir ve mide asitleri tarafından aşındırılır; bu nedenle sauropod gastrolitleri muhtemelen kazara yutulmuştur. Öte yandan, gastrolitlerin gelişmiş fosillerle bulunduğu sonucuna varmıştır. Theropod gibi dinozorlar Sinornithomimus ve Caudipteryx kuşlarınkine benziyor ve yiyecek öğütmek için gastrolitlerin kullanımı, hem bu dinozorların hem de kuşların evrimleştiği dinozorlar grubunun başlarında ortaya çıkmış olabilir.[15][16]
Üreme biyolojisi
Dişi kuşlar, yumurtalarken, sert dış kemik ile bacakları arasında uzuvlarında özel bir kemik türü çıkarırlar. ilik.[17] Zengin olan bu medüller kemik kalsiyum, yumurta kabuğu yapmak için kullanılır ve onu üreten kuşlar yumurtlamayı bitirdiklerinde onu emerler.[18] Theropod fosillerinde medüller kemik bulundu Tyrannosaurus ve Allosaurus ve ornitopodun Tenontosaurus.[18][19]
Çünkü içeren dinozorlar dizisi Allosaurus ve Tyrannosaurus yol açan çizgiden saptı Tenontosaurus dinozorların evriminin çok erken dönemlerinde, her iki grupta da medüller kemiğin varlığı, dinozorların genel olarak medüller doku ürettiğini göstermektedir. Diğer taraftan, timsahlar dinozorlara en yakın ikinci olan kaybolmamış akrabalar kuşlardan sonra, medüller kemik üretmezler. Bu doku ilk olarak ornitolar, Triyas Archosaur dinozorların evrimleştiği düşünülen grup.[18]
Medüller kemik, yetişkin altı boyuttaki örneklerde bulunmuştur, bu da dinozorların ulaştığını düşündürmektedir. cinsel olgunluk tam büyümeden önce. Yetişkinlik altı boyutta cinsel olgunluk, sürüngenlerde ve orta ila büyük boyutlu memelilerde de bulunur, ancak kuşlar ve küçük memeliler ancak tam büyüdükten sonra cinsel olgunluğa ulaşırlar - ki bu, ilk yıllarında gerçekleşir. Erken cinsel olgunluk, hayvanların yaşam döngülerinin belirli özellikleriyle de ilişkilidir: gençler çaresiz değil, nispeten iyi gelişmiş doğarlar; ve yetişkinler arasında ölüm oranı yüksektir.[18]
Solunum sistemi
Hava keseciklerinin

Yaklaşık 1870'den itibaren bilim adamları, birçok dinozorun kafa sonrası iskeletlerinin birçok hava dolu boşluk içerdiği konusunda genel olarak hemfikir oldular (postkranial iskelet pnömatikliği özellikle omurlarda. Kafatasının pnömatizasyonu (örneğin paranazal sinüsler ) ikisinde de bulunur sinapsitler ve Archosaurs ancak postkraniyal pnömatizasyon yalnızca kuşlarda bulunur, kuş olmayan Saurischian dinozorlar ve pterozorlar.
Uzun bir süre bu boşluklar sadece ağırlık tasarrufu sağlayan cihazlar olarak görülüyordu, ancak Bakker bağlantılı olduklarını önerdiler hava keseciklerinin kuş yapanlar gibi solunum sistemleri tüm hayvanlar arasında en verimli olanı.[9]
John Ruben et al. (1997, 1999, 2003, 2004) buna itiraz etti ve dinozorların timsah benzeri bir sistemden güç alan "gelgit" solunum sistemine (içeri ve dışarı) sahip olduğunu öne sürdü. hepatik piston mekanizma - esas olarak kaslara bağlı kaslar kasık çek karaciğer geriye doğru, akciğerlerin nefes almasını sağlar; Bu kaslar gevşediğinde, akciğerler önceki boyutlarına ve şekline geri döner ve hayvan nefes verir. Ayrıca bunu şüphe etmek için bir neden olarak sundular. dinozorlardan gelen kuşlar.[20][21][22][23][24]
Eleştirmenler, kuş hava keseleri olmadan modern bir sürüngenin birkaç yönündeki mütevazı iyileştirmelerin olduğunu iddia ettiler. dolaşım ve solunum sistemleri, sürüngenin benzer büyüklükteki bir memelinin oksijen akışının% 50 ila% 70'ini elde etmesini sağlar,[25] ve kuşlarda hava kesesi olmaması endoterminin gelişmesini engellemeyecektir.[26] Ruben'in bilimsel dergilerinde çok az resmi çürütme yayınlanmıştır. ve diğerleri. 'Dinozorların kuş tipi hava keselerine sahip olamayacağı iddiası; ama biri şu noktaya işaret ediyor: Sinosauropteryx İddialarının çoğunu dayandırdıkları fosil ciddi şekilde düzleşmişti ve bu nedenle karaciğerin hepatik bir piston mekanizmasının parçası olarak hareket etmek için doğru şeklin olup olmadığını söylemek imkansızdı.[27] Son zamanlarda yayınlanan bazı makaleler, Ruben'in et al. dinozorlarda hava keselerinin varlığına karşı çıktı.[28]
Araştırmacılar, hava keseleri için kanıt ve argümanlar sundular. Sauropodlar, "prosauropodlar ", Coelurosaurs, Ceratosaurlar ve theropodlar Aerosteon ve Kölofiz.
Gelişmiş sauropodlarda ("neosauropodlar") bel ve kalça bölgelerinin omurları hava kesesi belirtileri gösterir. Erken dönem sauropodlarda sadece servikal (boyun) omurlar bu özellikleri gösterir. Kuş embriyolarında gelişim sırası bulunursa bir rehber Hava keseleri aslında iskeletteki kanallardan önce gelişti ve onları daha sonraki şekillerde barındırdı.[29][30]

Theropodlarda hava kesesi kanıtı da bulunmuştur. Araştırmalar gösteriyor ki fosiller Coelurosaurs,[31] Ceratosaurlar,[28] ve theropodlar Kölofiz ve Aerosteon hava kesesi kanıtı sergileyin. Kölofiz, geç Triyas, fosillerinde hava kesesi kanallarının kanıtlarını gösteren en eski dinozorlardan biridir.[30] Aerosteon, Geç Kretase allosaur Şimdiye kadar en kuş benzeri hava kesesi bulundu.[32]
erken Sauropodomorflar geleneksel olarak "prosauropodlar" olarak adlandırılan grup da dahil olmak üzere, hava keseleri de olabilir. Olası pnömatik girintiler bulunmasına rağmen Plateosaurus ve Thecodontosaurus girintiler çok küçük. 2007'de yapılan bir çalışma, kardeş taksonlardaki (theropodlar ve sauropodlar) kanıtlara dayanarak, prosauropodların muhtemelen abdominal ve servikal hava keselerine sahip oldukları sonucuna varmıştır. Çalışma, prosauropodların kuş benzeri bir akciğere sahip olup olmadığını belirlemenin imkansız olduğu, ancak hava keselerinin neredeyse kesin olarak mevcut olduğu sonucuna vardı.[33] Hava keselerinin varlığının ve bunların akciğer ventilasyonunda kullanımının bir başka göstergesi, hava değişim hacminin (her nefeste değiş tokuş edilen hava hacmi) yeniden yapılandırılmasından gelir. Plateosaurus, 29 ml / kg vücut ağırlığı başına hava hacmi oranı olarak ifade edildiğinde kaz ve diğer kuşların değerlerine benzer ve tipik memeli değerlerinden çok daha yüksektir.[34]
Şu ana kadar hava kesesine dair hiçbir kanıt bulunamadı ornitorik dinozorlar. Ancak bu, ornithischians'ın memelilerinkine benzer metabolik hızlara sahip olamayacağı anlamına gelmez, çünkü memelilerde hava kesesi de yoktur.[35]
Dinozorlarda hava kesesi gelişimi için üç açıklama öne sürüldü:[32]
- Solunum kapasitesinde artış. Bu muhtemelen en yaygın hipotezdir ve birçok dinozorun oldukça yüksek olduğu fikrine uymaktadır. metabolik hızlar.
- Denge ve manevra kabiliyetini düşürerek iyileştirme ağırlık merkezi ve azaltma dönme ataleti. Ancak bu, dört ayaklı sauropodlarda hava keselerinin genişlemesini açıklamaz.
- Soğutma mekanizması olarak. Görünüşe göre hava keseleri ve tüyler yaklaşık aynı zamanda Coelurosaurs. Tüyler ısıyı muhafaza ederse, sahipleri aşırı ısıyı dağıtmak için bir araca ihtiyaç duyacaktı. Bu fikir makuldür, ancak daha fazla ampirik desteğe ihtiyaç duyar.
Sauropodun çeşitli bölümlerinin hacimlerinin hesaplanması Apatosaurus ' solunum sistemi, sauropodlarda kuş benzeri hava keselerinin kanıtlarını destekler:
- Varsayalım ki Apatozorlardinozorların hayatta kalan en yakın akrabaları gibi timsahlar ve kuşlar yoktu diyafram, ölü boşluk hacmi 30 tonluk bir örneğin yaklaşık 184 litre olacaktır. Bu, ağız, soluk borusu ve hava tüplerinin toplam hacmidir. Hayvan bundan daha az nefes verirse, bayat hava dışarı atılmaz ve sonraki inhalasyonda akciğerlere geri çekilir.
- Tahminleri gelgit hacmi - tek bir nefeste akciğerlere giren veya çıkan hava miktarı - türüne bağlıdır. solunum sistemi hayvanın vardı: 904 litre kuş ise; Memeliyse 225 litre; Sürüngen ise 19 litre.
Bu temelde, Apatozorlar bir sürüngen solunum sistemine sahip olamazdı, çünkü gelgit hacmi ölü alan hacminden daha az olacaktı, böylece bayat hava dışarı atılmayacaktı, ancak akciğerlere geri çekilecekti. Benzer şekilde, bir memeli sistemi akciğerlere her nefeste sadece 225 - 184 = 41 litre temiz, oksijenli hava sağlayacaktır. Apatozorlar bu nedenle ya modern dünyada bilinmeyen bir sisteme sahip olmalı ya da kuşlar ', birden fazla hava kesesi ve içinden akan bir akciğer ile. Dahası, bir kuş sistemi yalnızca yaklaşık 600 litrelik bir akciğer hacmine ihtiyaç duyarken, bir memeli için yaklaşık 2.950 litre gerekirdi ki bu, 30 tonluk bir alandaki tahmini 1.700 litrelik alanı aşacaktır. Apatozorlar′ Göğüs.[36]
Kuş benzeri hava keselerine sahip dinozor solunum sistemleri, benzer büyüklükteki ve yapıdaki memelilerin sürdürebileceğinden daha yüksek aktivite seviyelerini sürdürebilirdi. Çok verimli bir oksijen kaynağı sağlamanın yanı sıra, hızlı hava akışı, aktif ancak derilerindeki aşırı ısıdan kurtulamayacak kadar büyük hayvanlar için gerekli olan etkili bir soğutma mekanizması olurdu.[35]
Paleontolog Peter Ward ilk olarak en eski dinozorlarda ortaya çıkan hava kesesi sisteminin evriminin, çok düşük (% 11) atmosferik oksijene yanıt olabileceğini savundu. Karniyen ve Norian Triyas Dönemi çağları.[37]

Kaburgalardaki uncinate işlemleri
Kuşların "belirsiz süreçler "kaburgalarının arka kenarlarında ve bunlar, oksijen beslemesini iyileştirmek için göğsü pompalarken göğüs kaslarına daha fazla baskı sağlar. Uninat süreçlerin boyutu, kuşun yaşam tarzı ve oksijen gereksinimleri ile ilgilidir: yürüyen kuşlarda en kısadır ve en uzundur. Yüzeye çıktıklarında oksijen rezervlerini hızla yenilemeleri gereken dalış kuşlarında. Maniraptoran Dinozorlar da bu belirsiz süreçlere sahipti ve orantılı olarak modern dalış kuşlarındaki kadar uzundu, bu da maniraptoranların yüksek kapasiteli bir oksijen kaynağına ihtiyaç duyduklarını gösteriyor.[38][39]
Uninate süreçlerle aynı şekilde işlev görmüş olabilecek plakalar, Güney Afrika'daki fosillerde gözlemlenmiştir. ornitorik Dinozor Thescelosaurus ve yüksek oksijen tüketiminin ve dolayısıyla yüksek metabolik hızın kanıtı olarak yorumlanmıştır.[40]
Burun türbinleri
Burun türbinleri kıvrımlı ince kemik yapılarıdır. burun boşluğu. Çoğu memelide ve kuşta bunlar bulunur ve mukoza zarları iki işlevi yerine getiren. Havadaki kimyasalları absorbe etmek için mevcut alanı artırarak koku alma duyusunu geliştirir ve solunan havayı ısıtır ve nemlendirir ve solunan havadan ısıyı ve nemi çeker. kuruma akciğerlerin.
John Ruben ve diğerleri, dinozorlarda burun türbinlerine dair hiçbir kanıt bulunmadığını iddia ettiler. İnceledikleri tüm dinozorlar, burun türbinlerine uyum sağlayamayacak kadar dar ve kısa burun pasajlarına sahipti, bu nedenle dinozorlar, akciğerleri kurumuş olduğu için, dinozorlar memeli benzeri veya kuş benzeri bir metabolizma hızı için gereken solunum hızını sürdüremezdi. dışarı.[20][21][41][42] Ancak bu iddiaya karşı itirazlar var. Bazı kuşlarda burun konkaları yoktur veya çok küçüktür (örn. Ratites, Procellariiformes ve Falconiformes ) ve memeliler (örneğin balinalar, karıncayiyenler, yarasalar, filler ve çoğu primatlar ), bu hayvanlar tamamen endotermik olmasına ve bazı durumlarda çok aktif olmasına rağmen.[43][44][45][46] Diğer çalışmalar, burun türbinlerinin kırılgan olduğu ve nadiren fosillerde bulunduğu sonucuna varıyor. Özellikle fosil kuşlarda hiçbiri bulunamamıştır.[47]
2014'te Jason Bourke ve diğerleri Anatomik Kayıt pachycephalosaurlarda nazal türbinler bulduklarını bildirdi.[48]
Kardiyovasküler sistem

Prensipte, dinozorların dört odacıklı kalpler tarafından yönlendirilen iki parçalı dolaşıma sahip olması beklenirdi, çünkü birçoğu yerden yüksekte olan kafalarına kan vermek için yüksek tansiyona ihtiyaç duyacaktı, ancak omurgalı akciğerleri ancak oldukça düşük bir seviyeye dayanabilir. tansiyon.[35] 2000 yılında bir iskelet Thescelosaurus, şimdi sergileniyor Kuzey Carolina Doğa Bilimleri Müzesi, dört odacıklı bir evin kalıntılarını içerecek şekilde tanımlandı kalp ve bir aort. Yazarlar, kalbin yapısını yükselmiş bir metabolizma hızı için Thescelosaurus, değil sürüngen soğukkanlılık.[49] Onların vardığı sonuçlar tartışmalı; diğer araştırmacılar, kalbin gerçekten bir kalp olduğunu iddia ettikleri bir makale yayınladılar. somutlaştırma tamamen mineral "çimento". Dikkat ettikleri gibi: nesne için verilen anatomi yanlıştır, örneğin iddia edilen "aort", "kalp" ile buluştuğu yerde en dardır ve eksiktir. arterler ondan dallanma; "kalp", nervürlerden birini kısmen içine alır ve bazı yerlerde eşmerkezli tabakalardan oluşan bir iç yapıya sahiptir; ve sağ bacağın arkasında başka bir beton korunmuştur.[50] Orijinal yazarlar kendi konumlarını savundu; göğsün bir tür konkresyon içerdiğini, ancak kalbin ve aortun daha kaslı kısımlarını kısmen koruyan ve çevresinde oluştuğunu kabul ettiler.[51]
Nesnenin kimliğine bakılmaksızın, dinozorların iç anatomisi ve metabolik hızıyla çok az ilgisi olabilir. Hem modern timsahlar ve dinozorların en yakın yaşayan akrabaları olan kuşlar, timsahlarda modifiye edilmiş olmasına rağmen dört odacıklı kalplere sahiptir ve bu nedenle dinozorlar da muhtemelen onlara sahipti. Ancak bu tür kalplerin metabolik hıza bağlı olması gerekmez.[52]
Büyüme ve yaşam döngüsü
Bir basket topundan daha büyük hiçbir dinozor yumurtası bulunamamıştır ve nispeten küçük yumurtalarda büyük dinozorların embriyoları bulunmuştur. Maiasaura.[53] Memeliler gibi, dinozorlar da türlerinin tipik yetişkin boyutuna ulaştıklarında büyümeyi durdururken, olgun sürüngenler yeterli besinleri varsa yavaş büyümeye devam ettiler. Her boyuttaki dinozorlar, benzer boyuttaki modern sürüngenlerden daha hızlı büyüdüler; ancak benzer büyüklükteki "sıcakkanlı" modern hayvanlarla yapılan karşılaştırmaların sonuçları boyutlarına bağlıdır:[54][55]
| Ağırlık (kg) | Dinozorların karşılaştırmalı büyüme oranı | Bu boyut aralığındaki modern hayvanlar |
|---|---|---|
| 0.22 | Daha yavaş keseli hayvanlar | Sıçan |
| 1 – 20 | Keseli hayvanlara benzer, daha yavaş erken dönem kuşlar (koşabilen doğanlar) | Nereden Gine domuzu -e And kondoru |
| 100 – 1000 | Keseli hayvanlardan daha hızlı, erken dönem kuşlara benzer, daha yavaş plasental memeliler | Nereden kırmızı kanguru -e kutup ayısı |
| 1500 – 3500 | Çoğu plasentalı memeliye benzer | Nereden Amerikan bizonu -e gergedan |
| 25000 ve üzeri | Moderne benzer çok hızlı balinalar; ama büyütülmüş olanın yaklaşık yarısı altricial kuş (çaresiz doğan biri) - Bir kuşu 25.000 kilograma (25 uzun ton; 28 kısa ton) çıkarabilirse | Balinalar |

Tyrannosaurus rex bir "genç büyüme hamlesi" gösterdi:[56][57]
- 10 yaşında ½ ton
- onlu yaşların ortalarında yaklaşık 2 tona kadar çok hızlı büyüme (yılda yaklaşık ½ ton).
- ikinci on yıldan sonra ihmal edilebilir büyüme.
Bir iskeletin 2008 yılı çalışması Hadrosaur Hipakrosaurus bu dinozorun daha da hızlı büyüdüğü ve yaklaşık 15 yaşında tam boyutuna ulaştığı sonucuna vardı; ana kanıt kemiklerindeki büyüme halkalarının sayısı ve aralıkıydı. Yazarlar, bunu, av türlerinin avcılara çok sayıda yavru kaybetmeleri ve yerel çevrenin hızlı büyüme için yeterli kaynak sağlaması durumunda av türlerinin avcılarından daha hızlı büyümesi gerektiği şeklindeki bir yaşam döngüsü teorisiyle tutarlı buldular.[58]
Görünüşe göre bireysel dinozorlar oldukça kısa ömürlüdür, örn. en yaşlı (ölümde) Tyrannosaurus şimdiye kadar bulunan 28 ve en eskisi Sauropod 38 yaşındaydı.[56] Predasyon muhtemelen çok genç dinozorların yüksek ölüm oranından ve cinsel olarak olgun dinozorların yüksek ölüm oranlarından kaynaklanan cinsel rekabetten sorumluydu.[59]
Metabolizma
Yaşam tarzı hakkında bilimsel görüş, metabolizma ve sıcaklık düzenlemesi 19. yüzyılın ortalarında dinozorların keşfinden bu yana zamanla çeşitlilik göstermiştir. Metabolik enzimlerin aktivitesi sıcaklığa göre değişir, bu nedenle sıcaklık kontrolü, ister endotermik ister ektotermik olsun, herhangi bir organizma için hayati önem taşır. Organizmalar şu şekilde kategorize edilebilir: poikilotermler (poikilo - değişen), iç sıcaklık dalgalanmalarına toleranslı ve Homeotherms (homeo - aynı), sabit bir çekirdek sıcaklığı sürdürmesi gerekir. Hayvanlar ayrıca şu şekilde kategorize edilebilir: endotermler, sıcaklıklarını dahili olarak düzenleyen ve ektotermler, harici ısı kaynaklarının kullanımıyla sıcaklığı düzenleyen.
Tartışma ne hakkında
"Sıcakkanlılık "karmaşık ve belirsiz bir terimdir, çünkü aşağıdakilerin bir kısmını veya tamamını içerir:
- Ana Sayfayani oldukça sabit bir vücut ısısını korumak. Modern endotermler çeşitli sıcaklıkları korur: 28 ° C (82 ° F) ila 30 ° C (86 ° F) tekdelikliler ve tembel hayvanlar; 33 ° C (91 ° F) ila 36 ° C (97 ° F) keseli hayvanlar; 36 ° C (97 ° F) ila 38 ° C (100 ° F) çoğu durumda plasentaller; ve kuşlarda yaklaşık 41 ° C (106 ° F).[35]
- Taşimetabolizma, yani yüksek tutmak metabolizma hızı özellikle dinlenirken. Biyokimyasal işlemler, bir hayvanın sıcaklığı 10C ° düşerse yaklaşık yarısı kadar hızlı çalıştığından, bu oldukça yüksek ve sabit bir vücut ısısı gerektirir; çoğu enzimler optimum olmak Çalışma sıcaklığı ve verimlilikleri tercih edilen aralığın dışında hızla düşer.[60]
- Endotermiyani, güneşlenme veya kas aktivitesi gibi davranışlar yerine örneğin yağı "yakarak" dahili olarak ısı üretme yeteneği. Endotermi prensipte oldukça sabit bir sıcaklığı korumanın en güvenilir yolu olmasına rağmen, pahalıdır; örneğin modern memeliler, modern sürüngenlerden 10 ila 13 kat daha fazla yiyeceğe ihtiyaç duyar.[35]
Büyük dinozorlar, aynı zamanda "toplu homeotermi" veya "kitlesel homeotermi" olarak da bilinen ataletli homeotermi ile sıcaklıklarını korumuş olabilirler. Başka bir deyişle, termal kapasite Bu kadar büyük hayvanların oranı o kadar yüksekti ki, sıcaklıklarının önemli ölçüde değişmesi iki gün veya daha uzun sürebilirdi ve bu, günlük sıcaklık döngülerinin neden olduğu değişiklikleri düzeltirdi. Bu yumuşatma etkisi büyük kaplumbağalarda gözlenmiştir ve timsahlar, fakat Plateosaurus Yaklaşık 700 kilogram (1.500 lb) ağırlığında olan, içinde etkili olabileceği en küçük dinozor olabilir. Ataletsel homeotermi küçük türler için veya daha büyük türlerin gençleri için mümkün olamazdı.[35] Büyük otoburların bağırsaklarında bitki fermantasyonu da önemli ölçüde ısı üretebilir, ancak yüksek ve sabit bir sıcaklığı muhafaza etmenin bu yöntemi etoburlar, küçük otoburlar veya daha büyük otoburların yavruları için mümkün olamazdı.[61]
Soyu tükenmiş canlıların iç mekanizmaları bilinemediğinden, çoğu tartışma homeotermi ve taşimetabolizma odaklıdır.
Metabolik hızların değerlendirilmesi, dinlenirken ve aktifken hızlar arasındaki ayrım nedeniyle karmaşıktır. Tüm modern sürüngenlerde ve çoğu memeli ve kuşta, topyekun aktivite sırasındaki maksimum oranlar, dinlenirken minimum oranlardan 10 ila 20 kat daha yüksektir. Bununla birlikte, birkaç memelide bu oranlar 70 kat farklıdır. Teorik olarak, kara omurgalılarının dinlenme sırasında bir sürüngen metabolizma hızına ve düz bir şekilde çalışırken kuş benzeri bir hızına sahip olması mümkün olacaktır. Bununla birlikte, bu kadar düşük bir dinlenme oranına sahip bir hayvan hızlı bir şekilde büyüyemez. Dev otçul Sauropodlar Yiyecek aramak için o kadar sürekli hareket halindeydiler ki, enerji harcamaları, istirahat halindeki metabolik hızlarının yüksek veya düşük olmasına bakılmaksızın hemen hemen aynı olacaktı.[62]
Metabolik seçenekler
Ana olasılıklar şu şekildedir:[35]
- Dinozorlar, modern sürüngenler gibi soğukkanlıydı, tek fark, pek çoğunun büyük boyutlarının vücut sıcaklıklarını dengelemiş olması dışında.
- Sıcakkanlıydılar, modern sürüngenlerden çok modern memeliler veya kuşlar gibi.
- Modern terimlerle ne soğukkanlı ne de sıcakkanlıydılar, ancak modern soğukkanlı ve sıcakkanlı hayvanlardan farklı ve bazı açılardan bunlar arasında orta düzeydeki metabolizmaları vardı.
- Bu tür metabolizma türlerinden iki veya üçüne sahip hayvanları dahil ettiler.
Dinozorlar yaklaşık 150 milyon yıldır ortalıktaydı, bu nedenle farklı grupların farklı metabolizmalar ve ısı düzenleyici rejimler geliştirmesi ve bazılarının ilk dinozorlardan farklı fizyolojiler geliştirmesi çok muhtemeldir.
Dinozorların tümü veya bazılarının ara metabolizmaları varsa, aşağıdaki özelliklere sahip olabilirler:[35]
- Düşük dinlenme metabolik hızları - ihtiyaç duydukları yiyecek miktarını azaltacak ve büyümeleri için bu yiyeceklerden daha fazla dinlenme metabolik hızına sahip hayvanlardan daha fazla yararlanmalarına izin verecektir.
- Ataletsel homeotermi
- Birçok modern sürüngen gibi, derinin hemen altındaki kan damarlarını genişleterek ve daraltarak ısı kaybını kontrol etme yeteneği.
- Dört odacıklı kalpler tarafından yönlendirilen iki parçalı dolaşımlar.
- Yüksek aerobik kapasite, sürekli aktiviteye izin verir.
Robert Reid, bu tür hayvanların "başarısız endotermler" olarak kabul edilebileceğini öne sürdü. Bu özelliklerle hem dinozorların hem de memelilerin Triyas dönemindeki atalarının bir aşamadan geçtiklerini tasavvur etti. Memeliler küçülmeye zorlandı Archosaurs hakim olmaya geldi Ekolojik nişler orta ila büyük hayvanlar için. Boyutlarının küçülmesi onları ısı kaybına karşı daha savunmasız hale getirdi çünkü yüzey alanı kütleye oranlarını artırdı ve böylece onları dahili ısı üretimini artırmaya ve böylece tam endoterm haline gelmeye zorladı. Öte yandan, dinozorlar orta ila çok büyük hayvanlar haline geldi ve bu nedenle "orta" tipte metabolizmayı koruyabildiler.[35]
Kemik yapısı
Armand de Ricqlès keşfedildi Haversian kanalları dinozor kemiklerinde ve dinozorlarda endotermi kanıtı olduğunu savundu. Bu kanallar, "sıcakkanlı" hayvanlarda yaygındır ve hızlı büyüme ve aktif bir yaşam tarzı ile ilişkilidir çünkü hızlı büyümeyi kolaylaştırmak ve stres veya yaralanmaların neden olduğu hasarı onarmak için kemiği geri dönüştürmeye yardımcı olurlar.[63] Yeniden yapılanma sırasında oluşan yoğun ikincil Haversian kemiği, birçok canlı endotermin yanı sıra dinozorlar, pterosaurlar ve therapsidlerde bulunur. İkincil Haversian kanalları boyut ve yaş, mekanik stres ve besin devri ile ilişkilidir. İkincil Haversian kanallarının varlığı, memelilerde ve dinozorlarda benzer kemik büyümesi ve yaşam süreleri olduğunu göstermektedir.[64] Bakker, dinozor fosillerinde fibrolameller kemiğin (hızlı üretilen ve lifli, dokuma bir görünüme sahip) varlığının endoterminin kanıtı olduğunu savundu.[9]
Bununla birlikte, daha sonra yapılan diğer araştırmaların bir sonucu olarak, kemik yapısı dinozorlarda, memelilerde veya sürüngenlerde güvenilir bir metabolizma göstergesi olarak kabul edilmez:
- Dinozor kemikleri genellikle yavaş ve hızlı büyümenin değişen dönemlerinden oluşan tutuklanmış büyüme hatları (LAG'ler) içerir; Aslında birçok çalışma, dinozorların yaşlarını tahmin etmek için büyüme halkalarını sayar.[55][56] Büyüme halkalarının oluşumu genellikle sıcaklıktaki mevsimsel değişikliklerden kaynaklanır ve bu mevsimsel etki bazen yavaş metabolizmanın ve ektoterminin bir işareti olarak görülmüştür. Ancak büyüme halkaları, kutup ayılarında ve kış uykusuna yatan memelilerde bulunur.[65][66] YEG'ler ve mevsimsel büyüme bağımlılığı arasındaki ilişki çözülmeden kalır.[67]
- Fibrolamellar kemik, genç timsahlarda oldukça yaygındır ve bazen yetişkinlerde bulunur.[68][69]
- Haversian kemiği kaplumbağalarda, timsahlarda ve kaplumbağalarda bulundu.[70] ancak genellikle küçük kuşlarda, yarasalarda, farelerde ve kemirgenlerde yoktur.[69]
Bununla birlikte, de Ricqlès, dinozorların kemik yapısı üzerine yapılan çalışmalarla ısrar etti ve Archosaurs. 2008 ortalarında, erken dinozorlar da dahil olmak üzere çok çeşitli arkozorlardan kemik örneklerini inceleyen bir makalenin ortak yazarlığını yaptı ve şu sonuca vardı:[71]
- En erken bile Archosauriforms çok hızlı büyüme kapasitesine sahip olabilirler, bu da oldukça yüksek metabolik hızlara sahip olduklarını gösterir. Daha sonraki formlardan en eski arkosauriformlar hakkında sonuçlar çıkarmak zor olsa da, kemik yapısı ve büyüme oranındaki türe özgü varyasyonlar çok muhtemel olduğundan, bu tür faktörlerin analizde hatalara neden olma riskini en aza indirebilecek araştırma stratejileri vardır.
- Archosaurs, üç ana gruba ayrılır. Triyas: ornitozanlar dinozorların evrimleştiği, hızlı büyümeye bağlı kaldı; timsahların ataları daha tipik "sürüngen" yavaş büyüme oranları benimsedi; ve diğer Triyas dönemine ait arozorların çoğunun orta büyüme oranları vardı.
Osteositlerin vasküler yoğunluğu ve yoğunluğu, şekli ve alanının osteohistolojik analizi, kuş olmayan dinozorları ve arkosauriformların çoğunu ( Proterosuchus, timsahlar ve Fitosaurlar ) ısıyı korudu ve mevcut memeliler ve kuşlarınkine benzer dinlenme metabolik hızlarına sahipti.[72]
Metabolizma hızı, kan basıncı ve akış
Endotermler aerobik metabolizmaya oldukça güvenir ve aktivite ve dinlenme sırasında yüksek oksijen tüketim oranlarına sahiptir. Dokuların ihtiyaç duyduğu oksijen kan tarafından taşınır ve dolayısıyla sıcak kanlı endotermlerin kalbindeki kan akış hızları ve kan basınçları, soğuk kanlı ektotermlere göre oldukça yüksektir.[73] Dinozorların minimum kan basıncını, kalp ile başın üst kısmı arasındaki dikey mesafeyi tahmin ederek ölçmek mümkündür, çünkü bu kan sütununun alt kısmında, kan basıncına eşit bir basınç olması gerekir. hidrostatik basınç kan ve yerçekiminin yoğunluğundan elde edilir. Bu basınca eklenen, kanın dolaşım sisteminden geçmesi için gerekli olan basınçtır. 1976 yılında, yükseklikleri nedeniyle, birçok dinozorun endotermik aralık içinde minimum kan basıncına sahip olduğu ve yüksek basınç devresini düşük basınç devresinden vücuda ayırmak için dört odacıklı kalpleri olması gerektiği belirtildi. akciğerler.[74] Bu dinozorların basitçe kan sütununu desteklemek için mi yoksa endoterminin veya her ikisinin gerektirdiği yüksek kan akış oranlarını desteklemek için mi yüksek tansiyona sahip olduğu açık değildi.
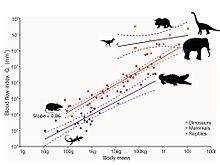
Bununla birlikte, dinozorların fosil bacak kemiklerindeki küçük deliklerin son analizi, kan akış hızı ve dolayısıyla metabolik hız için bir gösterge sağlıyor.[75] Deliklere besin foramina adı verilir ve besleyici arter, kemiğin küçük damarlarına dallandığı kemiğin iç kısmından geçen ana kan damarıdır. Haversian kanalı sistemi. Bu sistem, eski kemiğin yeni kemikle değiştirilmesinden ve böylece hareket sırasında doğal olarak meydana gelen mikro kırıkların onarılmasından sorumludur. Bu onarım olmadan, mikro kırıklar oluşacak ve Gerilme kırıkları ve nihayetinde feci kemik yetmezliği. Besin forameninin boyutu, kan akışına göre bir kan akışı indeksi sağlar. Hagen-Poiseuille denklemi. Elbette, boyut aynı zamanda hayvanın vücut boyutuyla da ilgilidir, bu nedenle bu etki, allometri. Canlı memelilerde femurların besleyici foramenlerinin kan akış indeksi, maksimum sürekli hareket sırasında ölçülen, hayvanların maksimum metabolik hızlarıyla doğru orantılı olarak artar. Memeli kan akış indeksi, ektotermik sürüngenler. Beş taksonomik gruptan on fosil dinozor türü, vücut büyüklükleri hesaba katıldığında memelilerdekinden bile daha yüksek indisler ortaya koyuyor, bu da onların oldukça aktif, aerobik hayvanlar olduklarını gösteriyor. Dolayısıyla yüksek kan akış hızı, yüksek tansiyon, dört odacıklı bir kalp ve sürekli aerobik metabolizma, endotermi.
Büyüme oranları
Dinozorlar, nispeten hızlı bir şekilde küçük yumurtalardan birkaç tona kadar büyüdüler. A natural interpretation of this is that dinosaurs converted food into body weight very quickly, which requires a fairly fast metabolism both to forage actively and to assimilate the food quickly.[76] Developing bone found in juveniles is distinctly porous, which has been linked to vascularization and bone deposition rate, all suggesting growth rates close to those observed in modern birds.
But a preliminary study of the relationship between adult size, growth rate, and body temperature concluded that larger dinosaurs had higher body temperatures than smaller ones had; Apatozorlar, the largest dinosaur in the sample, was estimated to have a body temperature exceeding 41 °C (106 °F), whereas smaller dinosaurs were estimated to have body temperatures around 25 °C (77 °F)[77] – for comparison, normal human body temperature is about 37 °C (99 °F).[78][79] Based on these estimations, the study concluded that large dinosaurs were inertial homeotherms (their temperatures were stabilized by their sheer bulk) and that dinosaurs were ectothermic (in colloquial terms, "cold-blooded", because they did not generate as much heat as mammals when not moving or digesting food).[77] These results are consistent with the relationship between dinosaurs' sizes and growth rates (described above).[80][81] Studies of the sauropodomorph Massospondylus and early theropod Syntarsus (Megapnosaurus ) reveal growth rates of 3 kg/year and 17 kg/year, respectively, much slower than those estimated of Maiasaura and observed in modern birds.[82]
Oxygen isotope ratios in bone
Oranı izotoplar 16O ve 18O in bone depends on the temperature the bone formed at: the higher the temperature, the more 16O. Barrick and Showers (1999) analyzed the isotope ratios in two theropodlar that lived in temperate regions with seasonal variation in temperature, Tyrannosaurus (ABD) ve Giganotosaurus (Arjantin):[83]
- dorsal vertebrae from both dinosaurs showed no sign of seasonal variation, indicating that both maintained a constant core temperature despite seasonal variations in air temperature.
- ribs and leg bones from both dinosaurs showed greater variability in temperature and a lower average temperature as the distance from the vertebrae increased.
Barrick and Showers concluded that both dinosaurs were endothermic but at lower metabolic levels than modern mammals, and that inertial homeothermy was an important part of their temperature regulation as adults. Their similar analysis of some Late Cretaceous ornithischians in 1996 concluded that these animals showed a similar pattern.[84]
However this view has been challenged. The evidence indicates homeothermy, but by itself cannot prove endothermy. Secondly, the production of bone may not have been continuous in areas near the extremities of limbs – in allosaur skeketons lines of arrested growth ("LAGs"; rather like büyüme halkaları ) are sparse or absent in large limb bones but common in the fingers and toes. While there is no absolute proof that LAGs are temperature-related, they could mark times when the extremities were so cool that the bones ceased to grow. If so, the data about oxygen isotope ratios would be incomplete, especially for times when the extremities were coolest. Oxygen isotope ratios may be an unreliable method of estimating temperatures if it cannot be shown that bone growth was equally continuous in all parts of the animal.[35]
Predator–prey ratios
- cold-blooded predators need much less food than warm-blooded ones, so a given mass of prey can support far more cold-blooded predators than warm-blooded ones.
- the ratio of the total mass of predators to prey in dinosaur communities was much more like that of modern and recent warm-blooded communities than that of recent or fossil cold-blooded communities.
- hence predatory dinosaurs were warm-blooded. And since the earliest dinosaurs (e.g. Staurikosaurus, Herrerasaurus ) were predators, all dinosaurs must have been warm-blooded.
This argument was criticized on several grounds and is no longer taken seriously (the following list of criticisms is far from exhaustive):[86][87]
- Estimates of dinosaur weights vary widely, and even a small variation can make a large difference to the calculated predator–prey ratio.
- Onun örneklem may not have been representative. Bakker obtained his numbers by counting museum specimens, but these have a bias towards rare or especially well-preserved specimens, and do not represent what exists in fossil beds. Even fossil beds may not accurately represent the actual populations, for example smaller and younger animals have less robust bones and are therefore less likely to be preserved.
- There are no published predator–prey ratios for large ectothermic predators, because such predators are very rare and mostly occur only on fairly small islands. Large ectothermic herbivores are equally rare. So Bakker was forced to compare mammalian predator–prey ratios with those of fish and invertebrate communities, where life expectancies are much shorter and other differences also distort the comparison.
- The concept assumes that predator populations are limited only by the availability of prey. However other factors such as shortage of nesting sites, cannibalism or predation of one predator on another can hold predator populations below the limit imposed by prey biomass, and this would misleadingly reduce the predator–prey ratio.
- Ecological factors can misleadingly reduce the predator–prey ratio, for example: a predator might prey on only some of the "prey" species present; disease, parasites and starvation might kill some of the prey animals before the predators get a chance to hunt them.
- It is very difficult to state precisely what preys on what. For example, the young of herbivores may be preyed upon by lizards and snakes while the adults are preyed on by mammals. Conversely the young of many predators live largely on invertebrates and switch to vertebrates as they grow.
Duruş ve yürüyüş
Bu bölüm için ek alıntılara ihtiyaç var doğrulama. (Eylül 2008) (Bu şablon mesajını nasıl ve ne zaman kaldıracağınızı öğrenin) |

Dinosaurs' limbs were erect and held under their bodies, rather than sprawling out to the sides like those of lizards and newts. The evidence for this is the angles of the joint surfaces and the locations of muscle and tendon attachments on the bones. Attempts to represent dinosaurs with sprawling limbs result in creatures with dislocated hips, knees, shoulders and elbows.[88]
Carrier's constraint states that air-breathing vertebrates with two lungs that flex their bodies sideways during locomotion find it difficult to move and breathe at the same time. This severely limits stamina, and forces them to spend more time resting than moving.[89]
Sprawling limbs require sideways flexing during locomotion (except for tortoises and turtles, which are very slow and whose armor keeps their bodies fairly rigid). However, despite Carrier's constraint, sprawling limbs are efficient for creatures that spend most of their time resting on their bellies and only move for a few seconds at a time—because this arrangement minimizes the energy costs of getting up and lying down.
Erect limbs increase the costs of getting up and lying down, but avoid Carrier's constraint. This indicates that dinosaurs were active animals because Doğal seçilim would have favored the retention of sprawling limbs if dinosaurs had been sluggish and spent most of their waking time resting. An active lifestyle requires a metabolism that quickly regenerates energy supplies and breaks down waste products which cause fatigue, i.e., it requires a fairly fast metabolism and a considerable degree of homeothermy.
Additionally, an erect posture demands precise balance, the result of a rapidly functioning neuromuscular system. This suggests endothermic metabolism, because an ectothermic animal would be unable to walk or run, and thus to evade predators, when its core temperature was lowered. Other evidence for endothermy includes limb length (many dinosaurs possessed comparatively long limbs) and bipedalism, both found today only in endotherms.[90] Many bipedal dinosaurs possessed gracile leg bones with a short thigh relative to calf length. This is generally an adaptation to frequent sustained running, characteristic of endotherms which, unlike ectotherms, are capable of producing sufficient energy to stave off the onset of anaerobic metabolism in the muscle.[91]
Bakker and Ostrom both pointed out that all dinosaurs had erect hindlimbs and that all quadrupedal dinosaurs had erect forelimbs; and that among living animals only the endothermic ("warm-blooded") mammals and birds have erect limbs (Ostrom acknowledged that timsahlar ' occasional "high walk" was a partial exception). Bakker claimed this was clear evidence of endothermy in dinosaurs, while Ostrom regarded it as persuasive but not conclusive.[9][92]
A 2009 study supported the hypothesis that endothermy was widespread in at least larger non-avian dinosaurs, and that It was plausibly ancestral for all dinosauriforms, based on the biomechanics of running,[93] though it has also been suggested that endothermy appeared much earlier in Archosauromorph evolution, perhaps even preceding the origin of Archosauriformes.[72]

There is now no doubt that many Theropod dinosaur species had feathers, including Shuvuuia, Sinosauropteryx ve Dilong (an early tyrannosaur ).[94][27][95] These have been interpreted as insulation and therefore evidence of warm-bloodedness.
But direct, unambiguous impressions of feathers have only been found in Coelurosaurs (which include the birds and tyrannosaurs, among others), so at present feathers give us no information about the metabolisms of the other major dinosaur groups, e.g. Kölofizitler, Ceratosaurlar, karnozorlar veya Sauropodlar. Filamentous integument was also present in at least some ornithischians, such as Tianyulong, Kulindadromeus ve Psittacosaurus, not only indicating endothermy in this group, but also that feathers were already present in the first ornithodiran (the last common ancestor of dinosaurs and pterosaurs). Their absence in certain groups like Ankilosauri could be the result of suppression of feather genes.[96]
The fossilised skin of Carnotaurus (bir Abelisaurid and therefore not a coelurosaur) shows an unfeathered, reptile-like skin with rows of bumps,[97] but the conclusion that Carnotaurus was necessarily featherless has been criticized as the impressions do not cover the whole body, being found only in the lateral region but not the dorsum.[98] Bir yetişkin Carnotaurus weighed about 2 tonnes,[99] and mammals of this size and larger have either very short, sparse hair or naked skins, so perhaps the skin of Carnotaurus tells us nothing about whether smaller non-coelurosaurid theropods had feathers. The tyrannosauroid Yutyrannus is known to have possessed feathers and weighed 1.1 tonne.[99]
Skin-impressions of Pelorosaurus ve diğeri Sauropodlar (dinosaurs with elephantine bodies and long necks) reveal large hexagonal scales, and some sauropods, such as Saltasaurus, had bony plates in their skin.[100] The skin of Ceratopsia'cılar consisted of large polygonal scales, sometimes with scattered circular plates.[101] "Mummified" remains and skin impressions of hadrosaurids reveal pebbly scales. It is unlikely that the ankylosaurids, such as Euoplocephalus, had insulation, as most of their surface area was covered in bony knobs and plates.[102] Likewise there is no evidence of insulation in the stegosaurlar. Thus insulation, and the elevated metabolic rate behind evolving them, may have been limited to the theropods, or even just a subset of theropods.[kaynak belirtilmeli ] Lack of feathers or other sort of insulation does not indicate ectothermy or low metabolisms, as observed in the relative hairlessness of mammalian megafauna, pigs, human children and the hairless bat being compatible with endothermy.[99]
Polar dinosaurs
Dinosaur fossils have been found in regions that were close to the poles at the relevant times, notably in southeastern Avustralya, Antarktika ve Kuzey Yamacı nın-nin Alaska. There is no evidence of major changes in the angle of the Earth's eksen, so polar dinosaurs and the rest of these ecosystems would have had to cope with the same extreme variation of day length through the year that occurs at similar enlemler today (up to a full day with no darkness in summer, and a full day with no sunlight in winter).[103]
Studies of fossilized vegetation suggest that the Alaska North Slope had a maximum temperature of 13 °C (55 °F) and a minimum temperature of 2 °C (36 °F) to 8 °C (46 °F) in the last 35 million years of the Kretase (slightly cooler than Portland, Oregon but slightly warmer than Calgary, Alberta). Even so, the Alaska North Slope has no fossils of large cold-blooded animals such as lizards and crocodilians, which were common at the same time in Alberta, Montana, ve Wyoming. This suggests that at least some non-avian dinosaurs were warm-blooded.[103] It has been proposed that North American polar dinosaurs may have migrated to warmer regions as winter approached, which would allow them to inhabit Alaska during the summers even if they were cold-blooded.[104] But a round trip between there and Montana would probably have used more energy than a cold-blooded land vertebrate produces in a year; in other words the Alaskan dinosaurs would have to be warm-blooded, irrespective of whether they migrated or stayed for the winter.[105] A 2008 paper on dinosaur migration by Phil R. Bell and Eric Snively proposed that most polar dinosaurs, including theropodlar, Sauropodlar, Ankylosaurians, ve Hipsilofodontlar, muhtemelen overwintered, olmasına rağmen hadrosaurids sevmek Edmontosaurus were probably capable of annual 2,600 km (1,600 mi) round trips.[106][107]

It is more difficult to determine the climate of southeastern Australia when the dinosaur fossil beds were laid down 115 to 105 milyon yıl önce, towards the end of the Early Cretaceous: these deposits contain evidence of permafrost, buz dilimleri, and hummocky ground formed by the movement of subterranean ice, which suggests mean annual temperatures ranged between −6 °C (21 °F) and 5 °C (41 °F); oksijen izotopu studies of these deposits give a mean annual temperature of 1.5 °C (34.7 °F) to 2.5 °C (36.5 °F). However the diversity of fossil vegetation and the large size of some of fossil trees exceed what is found in such cold environments today, and no-one has explained how such vegetation could have survived in the cold temperatures suggested by the physical indicators – for comparison Fairbanks, Alaska presently has a mean annual temperature of 2.9 °C (37.2 °F).[103] An annual migration from and to southeastern Australia would have been very difficult for fairly small dinosaurs in such as Leaellynasaura, a herbivore about 60 centimetres (2.0 ft) to 90 centimetres (3.0 ft) long, because seaways to the north blocked the passage to warmer latitudes.[103] Bone samples from Leaellynasaura ve Timimus, bir ornitomimid about 3.5 metres (11 ft) long and 1.5 metres (4.9 ft) high at the hip, suggested these two dinosaurs had different ways of surviving the cold, dark winters: the Timimus sample had lines of arrested growth (LAGs for short; similar to büyüme halkaları ), and it may have hibernated; ama Leaellynasaura sample showed no signs of LAGs, so it may have remained active throughout the winter.[108] A 2011 study focusing on hypsilophodont and theropod bones also concluded that these dinosaurs did not hibernate through the winter, but stayed active.[109]
Evidence for behavioral thermoregulation
Some dinosaurs, e.g. Spinosaurus ve Ouranosaurus, had on their backs "sails" supported by spines growing up from the omur. (This was also true, incidentally, for the sinapsit Dimetrodon.) Such dinosaurs could have used these sails to:
- take in heat by basking with the "sails" at right angles to the sun's rays.
- to lose heat by using the "sails" as radiators while standing in the shade or while facing directly towards or away from the sun.
But these were a very small minority of known dinosaur species.One common interpretation of the plates on stegosaurs' backs is as ısı eşanjörleri for thermoregulation, as the plates are filled with blood vessels, which, theoretically, could absorb and dissipate heat.[110]
This might have worked for a stegosaur with large plates, such as Stegosaurus, but other stegosaurs, such as Wuerhosaurus, Tuojiangosaurus ve Kentrosaurus possessed much smaller plates with a surface area of doubtful value for thermo-regulation. However, the idea of stegosaurian plates as heat exchangers has recently been questioned.[111]
Other evidence
Endothermy demands frequent respiration, which can result in water loss. In living birds and mammals, water loss is limited by pulling moisture out of exhaled air with mucous-covered respiratory turbinates, tissue-covered bony sheets in the nasal cavity. Several dinosaurs have olfactory turbinates, used for smell, but none have yet been identified with respiratory turbinates.[112]
Because endothermy allows refined neuromuscular control, and because brain matter requires large amounts of energy to sustain, some speculate that increased brain size indicates increased activity and, thus, endothermy. The encephalization quotient (EQ) of dinosaurs, a measure of brain size calculated using brain endocasts, varies on a spectrum from bird-like to reptile-like. Using EQ alone, coelosaurs appear to have been as active as living mammals, while theropods and ornithopods fall somewhere between mammals and reptiles, and other dinosaurs resemble reptiles.[112]
A study published by Roger Seymour in 2013 added more support to the idea that dinosaurs were endothermic. After studying saltwater crocodiles, Seymour found that even if their large sizes could provide stable and high body temperatures, during activity the crocodile's ectothermic metabolism provided less aerobic abilities and generate only 14% of the total muscle power of a similar sized endothermic mammal before full fatigue. Seymour reasoned that dinosaurs would have needed to be endothermic since they would have needed better aerobic abilities and higher power generation to compete with and dominate over mammals as active land animals throughout the Mesozoic era.[113]
The crocodilian puzzle and early archosaur metabolism
Bu bölüm için ek alıntılara ihtiyaç var doğrulama. (Eylül 2008) (Bu şablon mesajını nasıl ve ne zaman kaldıracağınızı öğrenin) |
It appears that the earliest dinosaurs had the features that form the basis for arguments for warm-blooded dinosaurs—especially erect limbs. This raises the question "How did dinosaurs become warm-blooded?" The most obvious possible answers are:
- "Their immediate ancestors (Archosaurs ) were cold-blooded, and dinosaurs began developing warm-bloodedness very early in their evolution." This implies that dinosaurs developed a significant degree of warm-bloodedness in a very short time, possibly less than 20M years. But in mammals' ancestors the evolution of warm-bloodedness seems to have taken much longer, starting with the beginnings of a ikincil damak around the beginning of the mid-Permiyen[114] and going on possibly until the appearance of hair about 164M years ago in the mid Jurassic[115]).
- "Dinosaurs' immediate ancestors (Archosaurs ) were at least fairly warm-blooded, and dinosaurs evolved further in that direction." This answer raises 2 problems: (Bir) The early evolution of archosaurs is still very poorly understood – large numbers of individuals and species are found from the start of the Triyas but only 2 species are known from the very late Permiyen (Archosaurus rossicus ve Protorosaurus speneri); (B) Crocodilians evolved shortly before dinosaurs and are closely related to them, but are cold-blooded (see below).
Crocodilians present some puzzles if one regards dinosaurs as active animals with fairly constant body temperatures. Crocodilians evolved shortly before dinosaurs and, second to birds, are dinosaurs' closest living relatives – but modern crocodilians are cold-blooded. This raises some questions:
- If dinosaurs were to a large extent "warm-blooded", when and how fast did warm-bloodedness evolve in their lineage?
- Modern crocodilians are cold-blooded but have several features associated with warm-bloodedness. How did they acquire these features?
Modern crocodilians are cold-blooded but can move with their limbs erect, and have several features normally associated with warm-bloodedness because they improve the animal's oxygen supply:[116]
- 4-chambered hearts. Mammals and birds have four-chambered hearts. Non-crocodilian reptiles have three-chambered hearts, which are less efficient because they allow oxygenated and de-oxygenated blood to mix and therefore send some de-oxygenated blood out to the body instead of to the lungs. Modern crocodilians' hearts are four-chambered, but are smaller relative to body size and run at lower pressure than those of modern mammals and birds. They also have a bypass that makes them functionally three-chambered when under water, conserving oxygen.[117]
- a diyafram, which aids breathing.
- a ikincil damak, which allows the animal to eat and breathe at the same time.
- a hepatic piston mechanism for pumping the lungs. This is different from the lung-pumping mechanisms of mammals and birds but similar to what some researchers claim to have found in some dinosaurs.[20][22]
Öyleyse neden yaptın Doğal seçilim favor these features, which are important for active warm-blooded creatures but of little apparent use to cold-blooded aquatic ambush predators that spend most of their time floating in water or lying on river banks?
It was suggested in the late 1980s that crocodilians were originally active, warm-blooded predators and that their archosaur ancestors were warm-blooded.[89][116] More recently, developmental studies indicate that crocodilian embryos develop fully four-chambered hearts first—then develop the modifications that make their hearts function as three-chambered under water. Using the principle that ontogeny recapitulates phylogeny, the researchers concluded that the original crocodilians had fully 4-chambered hearts and were therefore warm-blooded and that later crocodilians developed the bypass as they reverted to being cold-blooded aquatic ambush predators.[118][119]
More recent research on archosaur bone structures and their implications for growth rates also suggests that early archosaurs had fairly high metabolic rates and that the Triyas ancestors of crocodilians dropped back to more typically "reptilian" metabolic rates.[71]
If this view is correct, the development of warm-bloodedness in archosaurs (reaching its peak in dinosaurs) and in mammals would have taken more similar amounts of time. It would also be consistent with the fossil evidence:
- The earliest crocodylomorphs, e.g. Terrestrisuchus, were slim, leggy terrestrial predators.
- Erect limbs appeared quite early in Archosaurs ' evolution, and those of rauisuchians are very poorly adapted for any other posture.[120]
Ayrıca bakınız
- Dinosaur classification
- Dinosaur renaissance
- Dinozorların evrimi
- Evrimsel fizyoloji
- Dinozorların listesi
- Kuşların kökeni
Referanslar
- ^ a b Lucas, Spencer G. (1999). Dinosaurs: The Textbook (3. baskı). McGraw-Hill Companies, Inc. pp.3–9. ISBN 978-0-07-303642-7.
- ^ Fastovsky DE, Weishampel DB (2005). "Theropoda I: Diş ve pençede doğa kırmızısı". In Fastovsky DE, Weishampel DB (eds.). The Evolution and Extinction of the Dinosaurs (2. baskı). Cambridge University Press. pp. 265–299. ISBN 978-0-521-81172-9.
- ^ E.H., Cowles, R.B. and Bogert, C.M. (Haziran 1947). "Temperature Tolerances in the American Alligator and Their Bearing on the Habits, Evolution, and Extinction of the Dinosaurs". Biyolojinin Üç Aylık İncelemesi. 22 (2): 145–146. doi:10.1086/395712.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ a b Lucas, Spencer G. (2000). Dinosaurs: The Textbook (3. baskı). McGraw-Hill Companies, Inc. pp.1–3. ISBN 978-0-07-303642-7.
- ^ Torrens, Hugh (1997). "Politics and Paleontology". In Farlow, James O.; Brett-Surman, Michael K. (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. pp.175–190. ISBN 978-0-253-33349-0.
- ^ Benton, Michael J. (2000). "A brief history of dinosaur paleontology". Paul, Gregory S. (ed.). Dinozorların Bilimsel Amerikan Kitabı. New York: St. Martin's Press. pp. 10–44. ISBN 978-0-312-26226-6.
- ^ Bakker, R.T. (1968). "Dinozorların üstünlüğü". Keşif. 3 (2): 11–22.
- ^ Bakker, R.T. (1987). "The Return of the Dancing Dinosaurs". Czerkas, S. J .; Olson, E. C. (editörler). Geçmiş ve Günümüz Dinozorları, cilt. ben. Washington Üniversitesi Yayınları. ISBN 978-0-295-96541-3.
- ^ a b c d Bakker, Robert T. (1972). "Dinozorlarda endoterminin anatomik ve ekolojik kanıtı". Doğa. 238 (5359): 81–85. Bibcode:1972Natur.238 ... 81B. doi:10.1038 / 238081a0. S2CID 4176132.
- ^ Thomas, R.D.K .; Olson, E.C. (1980). Sıcakkanlı Dinozorlara Soğuk Bir Bakış. Westview Press. ISBN 978-0-89158-464-3.
- ^ Benton, M.J. (2005). Omurgalı Paleontoloji (3. baskı). Wiley-Blackwell. sayfa 221–223. ISBN 978-0-632-05637-8.
- ^ Paladino, F.V.; O'Connor, M.P.; Spotila, J.R. (1990). "Deri sırtlı kaplumbağaların metabolizması, gigantotermi ve dinozorların termoregülasyonu". Doğa. 344 (6269): 858–860. Bibcode:1990Natur.344..858P. doi:10.1038 / 344858a0. S2CID 4321764.
- ^ Barrick, R.E.; Showers, W.J.; Fischer, A.G. (1996). "Dört Ornithischian Dinozorunun ve Kretase İki Tıp Formasyonundan Bir Varanid Kertenkelesinin Termoregülasyonunun Karşılaştırılması: Oksijen İzotoplarından Kanıt". PALAIOS. 11 (4): 295–305. Bibcode:1996Palai. 11..295B. doi:10.2307/3515240. JSTOR 3515240.
- ^ a b c d e Norman, D.B. (Nisan 2001). "Dinosaur Feeding". Yaşam Bilimleri Ansiklopedisi. John Wiley & Sons. doi:10.1038/npg.els.0003321. ISBN 978-0470016176. Alındı 10 Eylül 2009.
- ^ Wings, O. (2007). "A review of gastrolith function with implications for fossil vertebrates and a revised classification" (PDF). Acta Palaeontologica Polonica. 52 (1): 1–16. Alındı 10 Eylül 2008.
- ^ Wings, O. & Sander. P.M. (Mart 2007). "No gastric mill in sauropod dinosaurs: new evidence from analysis of gastrolith mass and function in ostriches". Royal Society B Tutanakları. 274 (1610): 635–640. doi:10.1098/rspb.2006.3763. PMC 2197205. PMID 17254987.
- ^ http://jeb.biologists.org/content/184/1/63.full.pdf
- ^ a b c d Lee, Andrew H.; Werning, Sarah (2008). "Sexual maturity in growing dinosaurs does not fit reptilian growth models". Ulusal Bilimler Akademisi Bildiriler Kitabı. 105 (2): 582–587. Bibcode:2008PNAS..105..582L. doi:10.1073/pnas.0708903105. PMC 2206579. PMID 18195356.
- ^ Schweitzer, M.H .; Wittmeyer, J.L .; Horner, J.R. (2005). "Gender-specific reproductive tissue in ratites and Tyrannosaurus rex". Bilim. 308 (5727): 1456–1460. Bibcode:2005Sci...308.1456S. doi:10.1126/science.1112158. PMID 15933198. S2CID 30264554.
- ^ a b c Ruben, J.A., Jones, T.D., Geist, N.R. ve Hillenius, W. J. (Kasım 1997). "Theropod dinozorlarında ve erken kuşlarda akciğer yapısı ve havalandırma". Bilim. 278 (5341): 1267–1270. Bibcode:1997Sci ... 278.1267R. doi:10.1126/science.278.5341.1267.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ a b Ruben, J.A., Jones, T.D., Geist, N.R., Leitch, A., and Hillenius, W.J. (1997). "Lung ventilation and gas exchange in theropod dinosaurs". Bilim. 278 (5341): 1267–1270. Bibcode:1997Sci ... 278.1267R. doi:10.1126/science.278.5341.1267.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ a b Ruben, J.A., Dal Sasso, C., Geist, N.R., Hillenius, W. J., Jones, T.D., and Signore, M. (January 1999). "Pulmonary function and metabolic physiology of theropod dinosaurs". Bilim. 283 (5401): 514–516. Bibcode:1999Sci...283..514R. doi:10.1126/science.283.5401.514. PMID 9915693.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Ruben, J. A., Jones, T. D. and Geist, N. R. (2003). "Respiration and reproductive paleophysiology of dinosaurs and early birds" (PDF). Physiol. Biochem. Zool. 76 (2): 141–164. doi:10.1086/375425. hdl:10211.1/1472. PMID 12794669.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Hillenius, W. J. & Ruben, J.A. (November–December 2004). "The Evolution of Endothermy in Terrestrial Vertebrates: Who? When? Why?". Fizyolojik ve Biyokimyasal Zooloji. 77 (6): 1019–1042. doi:10.1086/425185. PMID 15674773.
- ^ Hicks, J.W. & Farmer, C.G. (Kasım 1997). "Lung Ventilation and Gas Exchange in Theropod Dinosaurs". Bilim. 278 (5341): 1267–1270. Bibcode:1997Sci ... 278.1267R. doi:10.1126/science.278.5341.1267.
- ^ Hicks, J.W. & Farmer, C.G. (Eylül 1999). "Gas exchange potential in reptilian lungs: implications for the dinosaur–avian connection". Solunum Fizyolojisi. 117 (2–3): 73–83. doi:10.1016/S0034-5687(99)00060-2. PMID 10563436.
- ^ a b Currie, P.J. & Chen, P-j. (Aralık 2001). "Anatomisi Sinosauropteryx prima from Liaoning, northeastern China". Kanada Yer Bilimleri Dergisi. 38 (12): 1705–1727. Bibcode:2001CaJES..38.1705C. doi:10.1139 / cjes-38-12-1705.
- ^ a b O'Connor, P. & Claessens, L. (July 2005). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Doğa. 436 (7048): 253–256. Bibcode:2005Natur.436..253O. doi:10.1038/nature03716. PMID 16015329. S2CID 4390587.
- ^ Wedel, M.J. (2003). "Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs". Paleobiyoloji. 29 (2): 243–255. doi:10.1666/0094-8373(2003)029<0243:VPASAT>2.0.CO;2. Full text currently online at "Findarticles.com: Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs". Paleobiyoloji. 2003. ve "Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs" (PDF). Arşivlenen orijinal (PDF) 17 Aralık 2008. Detailed anatomical analyses can be found at Wedel, M.J. (2003). "The Evolution of Vertebral Pneumaticity in Sauropod Dinosaurs". Omurgalı Paleontoloji Dergisi. 23 (2): 344–357. doi:10.1671/0272-4634(2003)023[0344:TEOVPI]2.0.CO;2.
- ^ a b Wedel, M.J. (June 2006). "Origin of postcranial skeletal pneumaticity in dinosaurs". Bütünleştirici Zooloji. 1 (2): 80–85. doi:10.1111/j.1749-4877.2006.00019.x. PMID 21395998.
- ^ Naish, D., Martill, D. M. and Frey, E. (June 2004). "Ecology, systematics and biogeographical relationships of dinosaurs, including a new theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil". Tarihsel Biyoloji. 16 (2–4): 57–70. CiteSeerX 10.1.1.394.9219. doi:10.1080/08912960410001674200. S2CID 18592288.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) This is also one of several topics featured in a post on Naish's blog, "Basal tyrant dinosaurs and my pet Mirischia". – note Mirischia bir Coelurosaur, which Naish believes was closely related to Compsognathus.
- ^ a b Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA, et al. (Eylül 2008). Kemp T (ed.). "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina". PLOS ONE. 3 (9): e3303. Bibcode:2008PLoSO...3.3303S. doi:10.1371/journal.pone.0003303. PMC 2553519. PMID 18825273.
- ^ Wedel, Mathew (2007). "What pneumaticity tells us about 'prosauropods', and vice versa" (PDF). Paleontolojide Özel Makaleler. 77: 207–222. Arşivlenen orijinal (PDF) 5 Temmuz 2008'de. Alındı 31 Ekim 2007.
- ^ Mallison, H. (2010). "The digital Plateosaurus II: an assessment of the range of motion of the limbs and vertebral column and of previous reconstructions using a digital skeletal mount". Acta Palaeontologica Polonica. 55 (3): 433–458. doi:10.4202/app.2009.0075.
- ^ a b c d e f g h ben j Reid, R.E.H. (1997). "Dinosaurian Physiology: the Case for "Intermediate" Dinosaurs". In Farlow, J.O.; Brett-Surman, M.K. (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. pp. 449–473. ISBN 978-0-253-33349-0. Alındı 12 Eylül 2008.
- ^ Paladino, F.V., Spotila, J.R., and Dodson, P. (1997). "A Blueprint for Giants: Modeling the Physiology of Large Dinosaurs". In Farlow, J.O.; Brett-Surman, M.K. (eds.). The Complete Dinosaur. Bloomington, Ind .: Indiana University Press. pp. 491–504. ISBN 978-0-253-21313-6.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Ward, Peter (2006) Out of thin air: Dinosaurs, birds, and earth's ancient atmosphere Pages 159–198, National Academies Press. ISBN 9780309141239.
- ^ Codd, J.R., Manning, P.L., Norell, M.A., and Perry, S.F. (Ocak 2008). "Avian-like breathing mechanics in maniraptoran dinosaurs". Royal Society B Tutanakları. 275 (1631): 157–161. doi:10.1098/rspb.2007.1233. PMC 2596187. PMID 17986432.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) News summary at "Why Dinosaurs Had 'Fowl' Breath". 7 Kasım 2007.
- ^ Tickle, P.G., Ennos, A.R., Lennox, L.E., Perry, S.F. and Codd, J.R. (November 2007). "Functional significance of the uncinate processes in birds". Deneysel Biyoloji Dergisi. 210 (Pt 22): 3955–3961. doi:10.1242/jeb.008953. PMID 17981863.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Fisher, P.E., Russell, D.A., Stoskopf, M.K., Barrick, R.E., Hammer, M., and Kuzmitz A.A. (Nisan 2000). "Cardiovascular Evidence for an Intermediate or Higher Metabolic Rate in an Ornithischian Dinosaur". Bilim. 288 (5465): 503–505. Bibcode:2000Sci...288..503F. doi:10.1126/science.288.5465.503. PMID 10775107.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) But note that this paper's main subject is that the fossil provided strong evidence of a 4-chambered heart, which is not widely accepted.
- ^ Ruben, J.A., Hillenius, W.J., Geist, N.R., Leitch, A., Jones, T.D., Currie, P.J., Horner, J.R. ve Espe, G. (Ağustos 1996). "Bazı Geç Kretase dinozorlarının metabolik durumu". Bilim. 273 (5279): 1204–1207. Bibcode:1996Sci ... 273.1204R. doi:10.1126 / science.273.5279.1204. S2CID 84693210.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Ruben, J.A. & Jones, T.D. (2000). "Selective Factors Associated with the Origin of Fur and Feathers". Amerikalı Zoolog. 40 (4): 585–596. doi:10.1093/icb/40.4.585.
- ^ Bang, B.G. (1966). "Procellariiformes'in koku alma cihazı". Acta Anatomica. 65 (1): 391–415. doi:10.1159/000142884. PMID 5965973.
- ^ Bang, B.G. (1971). "23 kuş sırasındaki koku alma sisteminin fonksiyonel anatomisi". Acta Anatomica. 79. 79: 1–76. doi:10.1159 / isbn.978-3-318-01866-0. ISBN 978-3-8055-1193-3. PMID 5133493.
- ^ Scott, J.H. (1954). "Nazal mukoza zarının ısı düzenleyici işlevi". Gırtlak ve Otoloji Dergisi. 68 (5): 308–317. doi:10.1017 / S0022215100049707. PMID 13163588.
- ^ Coulombe, H.N., Sam H. Ridgway, S.H. ve Evans, W.E. (1965). "İki domuzbalığı türünde solunum suyu değişimi". Bilim. 149 (3679): 86–88. Bibcode:1965Sci ... 149 ... 86C. doi:10.1126 / science.149.3679.86. PMID 17737801. S2CID 38947951.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Witmer, L.M. (Ağustos 2001). "Dinozorlarda ve Diğer Omurgalılarda Burun Deliği Konumu ve Burun İşlevi Açısından Önemi". Bilim. 293 (5531): 850–853. CiteSeerX 10.1.1.629.1744. doi:10.1126 / science.1062681. PMID 11486085. S2CID 7328047.
- ^ "Dinosaur breathing study shows that noses enhanced smelling and cooled brain".
- ^ Fisher, Paul E.; Russell, Dale A.; Stoskopf, Michael K.; Barrick, Reese E.; Hammer, Michael; Kuzmitz, Andrew A. (April 2000). "Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur". Bilim. 288 (5465): 503–505. Bibcode:2000Sci...288..503F. doi:10.1126/science.288.5465.503. PMID 10775107.
- ^ Rowe, Timothy; McBride, Earle F .; Sereno, Paul C. (February 2001). "Technical comment: dinosaur with a heart of stone". Bilim. 291 (5505): 783a. doi:10.1126/science.291.5505.783a. PMID 11157158.
- ^ Russell, Dale A.; Fisher, Paul E.; Barrick, Reese E.; Stoskopf, Michael K. (February 2001). "Reply: dinosaur with a heart of stone". Bilim. 291 (5505): 783a. doi:10.1126/science.291.5505.783a. PMID 11157158.
- ^ Chinsamy, Anusuya; ve Hillenius, Willem J. (2004). "Nonavian dinozorların fizyolojisi". Dinosauria, 2. 643–659.
- ^ Carpenter, K., Hirsch, K.F., and Horner, J.R. (1994). "Giriş". In Carpenter, K.; Hirsch, K.F.; Horner, J.R. (eds.). Dinozor Yumurtaları ve Bebekler. Cambridge University Press. ISBN 978-0-521-56723-7.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Erickson, G.M., Curry Rogers, K., Yerby, S.A. (July 2001). "Dinosaurian growth patterns and rapid avian growth rates". Doğa. 412 (6845): 429–433. Bibcode:2001Natur.412..429E. doi:10.1038/35086558. PMID 11473315. S2CID 4319534.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) Note Kristina Rogers also published papers under her maiden name, Kristina Curry.
- ^ a b Curry, K.A. (1999). "Ontogenetic Histology of Apatozorlar (Dinosauria: Sauropoda): New Insights on Growth Rates and Longevity". Omurgalı Paleontoloji Dergisi. 19 (4): 654–665. doi:10.1080/02724634.1999.10011179. Arşivlenen orijinal on 15 July 2010.
- ^ a b c Erickson, G.M., Makovicky, P.J., Currie, P.J., Norell, M.A., Yerby, S.A. & Brochu, C.A. (Ağustos 2004). "Gigantizm ve tyrannosaurid dinozorların karşılaştırmalı yaşam öyküsü parametreleri". Doğa. 430 (7001): 772–775. Bibcode:2004Natur.430..772E. doi:10.1038 / nature02699. PMID 15306807. S2CID 4404887.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Horner, J. R., and Padian, K. (September 2004). "Age and growth dynamics of Tyrannosaurus rex". Royal Society B Tutanakları. 271 (1551): 1875–1880. doi:10.1098/rspb.2004.2829. PMC 1691809. PMID 15347508.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Cooper,, L.N., Lee, A.H., Taper, M.L., and Horner, J.R. (August 2008). "Yırtıcı ve av dinozorlarının göreceli büyüme oranları, avlanmanın etkilerini yansıtır". Royal Society B Tutanakları. 275 (1651): 2609–2615. doi:10.1098 / rspb.2008.0912. PMC 2605812. PMID 18682367. Alındı 26 Ağustos 2008.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Erickson, G.M., Currie, P.J., Inouye, B.D. and Winn, A.A. (Temmuz 2006). "Tyrannosaur Life Tables: An Example of Nonavian Dinosaur Population Biology". Bilim. 313 (5784): 213–217. Bibcode:2006Sci...313..213E. doi:10.1126/science.1125721. PMID 16840697. S2CID 34191607.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ McGowan, C. (1991). Dinosaurs, Spitfires and Sea Dragons. Harvard Üniversitesi Yayınları. pp.133–135. ISBN 978-0-674-20769-1.
- ^ Mackie, R.I. (2002). "Mutualistic Fermentative Digestion in the Gastrointestinal Tract: Diversity and Evolution". Bütünleştirici ve Karşılaştırmalı Biyoloji. 42 (2): 319–326. doi:10.1093/icb/42.2.319. PMID 21708724. Alındı 12 Eylül 2008.
- ^ Paul, G.S. (1998). "Cope's rule". Carpenter, K .; Fraser, N .; Chure, D .; Kirkland, J.I. (eds.). Morrison Sempozyumu Bildiriler: Modern Jeoloji Dergisi'nin Özel Sayısı. Taylor ve Francis. s. 211. ISBN 978-90-5699-183-8. Alındı 12 Eylül 2008.
- ^ Ricqles, A. J. de. (1974). Endoterminin evrimi: histolojik kanıt. Evrim Teorisi 1: 51–80
- ^ Fastovsky & Weishampel 2009, s. 258.
- ^ Chinsamy, A., Rich, T. ve Vickers-Rich, P. (1998). "Kutupsal dinozor kemik histolojisi". Omurgalı Paleontoloji Dergisi. 18 (2): 385–390. doi:10.1080/02724634.1998.10011066. Arşivlenen orijinal 15 Temmuz 2010'da.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Klevezal, G.A., Mina, M.V. ve Oreshkin, A.V. (1996). Memelilerin yapılarının kaydedilmesi. Yaşın belirlenmesi ve yaşam öyküsünün yeniden yapılandırılması. CRC Basın. ISBN 978-90-5410-621-0.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Fastovsky & Weishampel 2009, s. 260.
- ^ Enlow, D.H. (1963). Kemik Tadilatının Prensipleri. Uzun kemiklerde ve mandibulada doğum sonrası büyüme ve yeniden şekillenme süreçlerinin bir hesabı. Springfield, IL: C.C. Thomas.
- ^ a b Reid, R.E.H. (1984). "Birincil kemik ve dinozor fizyolojisi". Jeoloji Dergisi. 121 (6): 589–598. Bibcode:1984GeoM..121..589R. doi:10.1017 / S0016756800030739.
- ^ Reid, R.E.H (1997). "Dinozorlar nasıl büyüdü". Farlow, J.O .; Brett-Surman, M.K. (eds.). Komple Dinozor. Bloomington: Indiana University Press. pp.403–413. ISBN 978-0-253-33349-0.
- ^ a b de Ricqlès, A., Padian, K., Knoll, F. ve Horner, J.R. (Nisan – Haziran 2008). "Archosaurlar ve onların eski akrabalarındaki yüksek büyüme oranlarının kökeni hakkında: Triyas arkosauriformları üzerine tamamlayıcı histolojik çalışmalar ve kemik histolojisinde" filogenetik sinyal "problemi". Annales de Paléontologie. 94 (2): 57–76. doi:10.1016 / j.annpal.2008.03.002.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) Özet ayrıca çevrimiçi olarak "Archosaurlarda Yüksek Büyüme Oranlarının Kökeni". 31 Mayıs 2008. Alındı 3 Haziran 2008.
- ^ a b Legendre, Lucas J .; Guénard, Guillaume; Botha-Brink, Jennifer; Cubo, Jorge (1 Kasım 2016). "Archosaurlarda Ataların Yüksek Metabolizma Hızı İçin Paleohistolojik Kanıtlar". Sistematik Biyoloji. 65 (6): 989–996. doi:10.1093 / sysbio / syw033. ISSN 1063-5157. PMID 27073251.
- ^ Seymour, Roger S .; Bennett-Stamper, Christina L .; Johnston, Sonya D .; Taşıyıcı, David R .; Grigg Gordon C. (2004). "Archosaur evriminin kökünde timsahların endotermik atalarının kanıtı" (PDF). Fizyolojik ve Biyokimyasal Zooloji. 77 (6): 1051–1067. doi:10.1086/422766. PMID 15674775.
- ^ Seymour Roger S. (1976). "Dinozorlar, endotermi ve kan basıncı". Doğa. 262 (5565): 207–208. Bibcode:1976Natur.262..207S. doi:10.1038 / 262207a0. S2CID 4157084.
- ^ Seymour, Roger S .; Smith, Sarah L; White, Craig R .; Henderson, Donald M .; Schwarz-Wings, Daniela (2012). "Uzun kemiklere kan akışı, memelilerde, sürüngenlerde ve dinozorlarda aktivite metabolizmasını gösterir". Royal Society B Tutanakları. 279 (1728): 451–456. doi:10.1098 / rspb.2011.0968. PMC 3234558. PMID 21733896.
- ^ Paul, G.S. (1988). Dünyanın Yırtıcı Dinozorları. New York: Simon ve Schuster. s.155. ISBN 978-0-671-61946-6.
- ^ a b Gillooly, J.F., Allen, A.P. ve Charnov, E.L. (Ağustos 2006). "Dinozor Fosilleri Vücut Sıcaklığını Tahmin Ediyor". PLOS Biyoloji. 4 (8): e248. doi:10.1371 / journal.pbio.0040248. PMC 1489189. PMID 16817695.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) Adresinde daha az teknik bir özet var Gross, L. (Ağustos 2006). "Matematik ve Fosiller Dinozor Metabolizması Üzerine Bir Tartışmayı Çözüyor". PLOS Biyoloji. 4 (8): e255. doi:10.1371 / journal.pbio.0040255. PMC 1489188. PMID 20076616.
- ^ Mackowiak, P. A .; S. S. Wasserman; M. M. Levine (23 Eylül 1992). "98.6 ° F (37.0 ° C) kritik değerlendirme, normal vücut ısısının üst sınırı ve Carl Reinhold August Wunderlich'in diğer mirası". JAMA. 268 (12): 1578–1580. doi:10.1001 / jama.1992.03490120092034. PMID 1302471.
- ^ Sund-Levander, Märtha; Forsberg, Christina; Wahren, Lis Karin (2002). "Yetişkin erkeklerde ve kadınlarda normal oral, rektal, timpanik ve aksiller vücut ısısı: sistematik bir literatür taraması". Scandinavian Journal of Caring Sciences. 16 (2): 122–128. doi:10.1046 / j.1471-6712.2002.00069.x. PMID 12000664.
- ^ Erickson, Gregory M .; Rogers, Kristina Curry; Yerby, Scott A. (2001). "Dinozorların büyüme modelleri ve hızlı kuş büyüme oranları". Doğa. 412 (6845): 429–433. Bibcode:2001Natur.412..429E. doi:10.1038/35086558. PMID 11473315. S2CID 4319534.
- ^ Curry, Kristina A. (1999). "Apatosaurus'un (Dinosauria: Sauropoda) Ontogenetik histolojisi: Büyüme oranları ve uzun ömür hakkında yeni bilgiler". Omurgalı Paleontoloji Dergisi. 19 (4): 654–665. doi:10.1080/02724634.1999.10011179.
- ^ Fastovsky, David E .; Weishampel, Dvid B. (2009). Dinozorlar: Kısa Bir Doğa Tarihi (PDF). s. 258.
- ^ Barrick, R.E. & Showers, W.J. (1999). "Termofizyoloji ve biyolojisi Giganotosaurus: ile karşılaştırıldığında Tyrannosaurus". Paleontoloji Electronica. 2 (2). Alındı 10 Eylül 2008.
- ^ Barrick, R.E, Showers, W.J. ve Fischer, A.G. (Ağustos 1996). "Dört Ornithischian Dinozorunun ve Kretase İki Tıp Formasyonundan Bir Varanid Kertenkelesinin Termoregülasyonunun Karşılaştırılması: Oksijen İzotoplarından Kanıt". PALAIOS. 11 (4): 295–305. Bibcode:1996Palai. 11..295B. doi:10.2307/3515240. JSTOR 3515240.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Bakker, R.T. (Eylül 1974). "Dinozor Biyoenerjetiği - Bennett ve Dalzell ve Feduccia'ya Bir Cevap". Evrim. 28 (3): 497–503. doi:10.2307/2407178. JSTOR 2407178. PMID 28564852.
- ^ Fastovsky, D.E., Weishampel, D.B. ve Sibbick, J. (2005). Dinozorların Evrimi ve Yok Olması. Cambridge University Press. ISBN 978-0-521-81172-9.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Farlow, J.O. (1980). "Yırtıcı / av biyokütle oranları, topluluk besin ağları ve dinozor fizyolojisi" (PDF). Thomas, R.D.K .; Olson, E.C. (editörler). Sıcakkanlı dinozorlara soğuk bir bakış. Boulder, CO: American Association for the Advancement of Science. sayfa 55–83. Arşivlenen orijinal (PDF) 17 Aralık 2008.
- ^ Bu, en geç 1909'da tanındı: "Dr. Holland ve Yayılan Sauropodlar". Arşivlenen orijinal 12 Haziran 2011. Argümanlar ve görüntülerin çoğu da Desmond, A. (1976). Sıcak Kanlı Dinozorlar. DoubleDay. ISBN 978-0-385-27063-2.
- ^ a b Taşıyıcı, D.R. (1987). "Dört ayaklılarda lokomotor dayanıklılığın evrimi: mekanik bir kısıtlamanın üstesinden gelmek". Paleobiyoloji. 13 (3): 326–341. doi:10.1017 / S0094837300008903. JSTOR 2400734.
- ^ Fastovsky & Weishampel 2009, s. 251.
- ^ Fastovsky & Weishampel 2009, s. 252.
- ^ Ostrom, J.H. (1980). "Dinozorlarda endoterminin kanıtı" (PDF). Thomas, R.D.K .; Olson, E.C. (editörler). Sıcakkanlı dinozorlara soğuk bir bakış. Boulder, CO: American Association for the Advancement of Science. s. 82–105. Arşivlenen orijinal (PDF) 27 Mayıs 2008.
- ^ Pontzer, H. .; Allen, V. .; Hutchinson, J. .; Farke, A. A. (2009). Farke, Andrew Allen (ed.). "Koşmanın biyomekaniği iki ayaklı dinozorlarda endotermiyi gösterir". PLOS ONE. 4 (11): e7783. Bibcode:2009PLoSO ... 4.7783P. doi:10.1371 / journal.pone.0007783. PMC 2772121. PMID 19911059.
- ^ Schweitzer, M.H., J.A. Watt, R. Avci, L. Knapp, L. Chiappe, M. Norell ve M. Marshall. (Temmuz 1999). "Kretase alvarezsaurid'in tüy benzeri yapılarında beta-keratine özgü immünolojik reaktivite, Shuvuuia deserti". Deneysel Zooloji Dergisi (Mol Dev Evol). 285 (2): 146–157. doi:10.1002 / (SICI) 1097-010X (19990815) 285: 2 <146 :: AID-JEZ7> 3.0.CO; 2-A. PMID 10440726. Arşivlenen orijinal 10 Aralık 2012.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Xu X .; Norell, M.A .; Kuang X .; Wang X .; Zhao Q. ve Jia C. (2004). "Çin'den gelen bazal tyrannosauroidler ve tyrannosauroidlerdeki protofeathers için kanıtlar". Doğa. 431 (7009): 680–684. Bibcode:2004Natur.431..680X. doi:10.1038 / nature02855. PMID 15470426. S2CID 4381777.
- ^ Benton, Michael J .; Dhouailly, Danielle; Jiang, Baoyu; McNamara, Maria (1 Eylül 2019). "Tüylerin Erken Kökeni". Ekoloji ve Evrimdeki Eğilimler. 34 (9): 856–869. doi:10.1016 / j.tree.2019.04.018. ISSN 0169-5347. PMID 31164250.
- ^ Bonaparte, J.F., Novas, E.E. ve Coria, R.A. (1990). "Carnotaurus sastrei Bonaparte, Patagonya'nın Orta Kretase'sinden boynuzlu, hafif yapılı karnozor ". Los Angeles County Doğa Tarihi Müzesi Bilime Katkılar. 416: 1–42. Arşivlenen orijinal 10 Mayıs 2008.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ Doktora, Andrea Cau (1 Mart 2016). "Theropoda: Miti e leggende post-moderne sui dinosauri mesozoici: Le squame di Carnotaurus". Theropoda. Alındı 25 Nisan 2020.
- ^ a b c Paul Gregory S. (2016). The Princeton Field Guide to Dinosaurs - 2nd Edition. Princeton University Press. ISBN 978-0-691-16766-4.
- ^ Czerkas, S.A. (1994). Lockley, M.G .; dos Santos, V.F .; Meyer, C.A .; Hunt, A.P. (editörler). "Sauropod Paleobiyolojisinin Yönleri". Gaia. Lizbon, Portekiz. 10.
| katkı =yok sayıldı (Yardım) - ^ Dodson, P. & Barlowe, W.D. (1996). Boynuzlu Dinozorlar: Doğal Bir Tarih. Princeton University Press. ISBN 978-0-691-05900-6. Ayrıca şuradaki resme bakın "Ceratopsian cilt". Arşivlenen orijinal 3 Şubat 2014.
- ^ Gosline, A. (16 Kasım 2004). "Dinozorların" kurşun geçirmez "zırhı ortaya çıktı". New Scientist.com haber servisi.
- ^ a b c d Rich, T.H., Vickers-Rich, P. ve Gangloff, R.A. (Şubat 2002). "Kutup Dinozorları". Bilim. 295 (5557): 979–980. doi:10.1126 / science.1068920. PMID 11834803. S2CID 28065814.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) Ayrıca bakınız Vickers-Rich, P. & Rich, T.H. (2004). "Antarktika Dinozorları" (PDF). Bilimsel amerikalı. Arşivlenen orijinal (PDF) 17 Mayıs 2008. ve Fiorillo, A.R. (2004). "Arktik Alaska Dinozorları" (PDF). Bilimsel amerikalı. 291 (6): 84–91. Bibcode:2004SciAm.291f..84F. doi:10.1038 / bilimselamerican1204-84. PMID 15597984. Arşivlenen orijinal (PDF) 17 Mayıs 2008.
- ^ Hotton, N. (1980). "Dinozor endotermisine bir alternatif: Mutlu gezginler". Thomas, R.D.K .; Olson, E.C. (editörler). Sıcakkanlı dinozorlara soğuk bir bakış. Boulder, CO: American Association for the Advancement of Science. sayfa 311–350. ISBN 978-0-89158-464-3.
- ^ Paul, G.S. (1988). Dünyanın Yırtıcı Dinozorları. New York: Simon ve Schuster. pp.160–161. ISBN 978-0-671-61946-6.
- ^ Bell, Phil R .; Snively, E. (2008). "Geçit töreninde kutup dinozorları: dinozor göçünün gözden geçirilmesi". Alcheringa. 32 (3): 271–284. doi:10.1080/03115510802096101.
- ^ Lloyd, Robin (4 Aralık 2008). "Kutup Dinozorları Soğuk Karanlık Kışlara Dayandı". LiveScience.com. Imaginova. Alındı 11 Aralık 2008.
- ^ Chinsamy, A., Rich, T.H. ve Vickers-Rich, P. (1998). "Kutupsal dinozor kemik histolojisi". Omurgalı Paleontoloji Dergisi. 18 (2): 385–390. doi:10.1080/02724634.1998.10011066. JSTOR 4523908.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) Ayrıca bakınız Leslie, M. (Aralık 2007). "Bir Dinozorun Ateşini Ölçmek". Smithsonian dergisi.[kalıcı ölü bağlantı ]
- ^ Woodward, Holly N .; Rich, Thomas H .; Chinsamy, Anusuya; Vickers-Rich, Patricia (2011). "Avustralya Kutup Dinozorlarının Büyüme Dinamikleri". PLOS ONE. 6 (8): e23339. Bibcode:2011PLoSO ... 623339W. doi:10.1371 / journal.pone.0023339. PMC 3149654. PMID 21826250.
- ^ de Bufrenil, V., Farlow, J.O. ve de Riqles, A. (1986). "Büyüme ve işlev Stegosaurus plakalar: kemik histolojisinden kanıt ". Paleobiyoloji. 12 (4): 459–473. doi:10.1017 / S0094837300003171. JSTOR 2400518.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı)
- ^ "Stegosaur plakaları ve sivri uçları sadece görünüş için". 18 Mayıs 2005. Arşivlenen orijinal 27 Eylül 2011.
- ^ a b Fastovsky & Weishampel 2009, s. 255.
- ^ Seymour Roger S. (5 Temmuz 2013). "Memelilere Karşı Büyük Timsahlarda Maksimal Aerobik ve Anaerobik Güç Üretimi: Dinozor Gigantothermy için Çıkarımlar". PLOS ONE. 8 (7): e69361. Bibcode:2013PLoSO ... 869361S. doi:10.1371 / journal.pone.0069361. PMC 3702618. PMID 23861968.
- ^ Kermack, D.M. & Kermack, K.A. (1984). Memeli karakterlerin evrimi. Londra: Croom Helm Kapitan Szabo Yayıncılar. s. 149. ISBN 978-0-7099-1534-8.
- ^ Saç veya kürkün en eski açık kanıtı, fosillerdedir. Castorocauda, 164 milyon yıl öncesinden Jurassic, Ji, Q .; Luo, Z-X, Yuan, C-X ve Tabrum, A.R. (Şubat 2006). "Orta Jura ve Erken Memelilerin Ekomorfolojik Çeşitlendirilmesinden Yüzen Bir Memeli Biçimi". Bilim. 311 (5764): 1123–7. Bibcode:2006Sci ... 311.1123J. doi:10.1126 / science.1123026. PMID 16497926. S2CID 46067702.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) Ayrıca şu adresteki haberlere bakın: "Jurassic" Kunduz "Bulundu; Memelilerin Tarihini Yeniden Yazıyor". En geç 1950'lerden beri saçın erken dönemlerde kanıt olabileceği tartışıldı.Triyas Sinodontlar gibi Thrinaxodon: Brink, A.S. (1955). "İskelet üzerine bir çalışma Diademodon". Paleontoloji Africana. 3: 3–39. ve Kemp, T.S. (1982). Memeli benzeri sürüngenler ve memelilerin kökeni. Londra: Akademik Basın. s.363. ISBN 978-0-12-404120-2. ancak cynodont burun kemiklerindeki foramina (küçük tüneller), en iyi ihtimalle belirsiz kanıtlardır, çünkü benzer foramina birkaç yaşayan sürüngende bulunur: Estes, R. (1961). "Cynodont sürüngeninin kafatası anatomisi Thrinaxodon liorhinus". Karşılaştırmalı Zooloji Müzesi Bülteni (1253): 165–180. ve Ruben, J.A. & Jones, T.D. (2000). "Kürk ve Tüylerin Kökeni ile İlişkili Seçici Faktörler". Amerikalı Zoolog. 40 (4): 585–596. doi:10.1093 / icb / 40.4.585.
- ^ a b McGowan, C. (1991). Dinozorlar, Ateşler ve Deniz Ejderhaları. Harvard Üniversitesi Yayınları. pp.151–152. ISBN 978-0-674-20769-1.
- ^ "Timsah kalbinin sırrı".
- ^ Summers, A.P. (2005). "Evrim: Sıcak kalpli timsahlar". Doğa. 434 (7035): 833–834. Bibcode:2005Natur.434..833S. doi:10.1038 / 434833a. PMID 15829945. S2CID 4399224.
- ^ Seymour, R. S., Bennett-Stamper, C.L., Johnston, S. D., Carrier, D.R. ve Grigg, G.C. (2004). "Archosaur evriminin kökünde timsahların endotermik atalarının kanıtı" (PDF). Fizyolojik ve Biyokimyasal Zooloji. 77 (6): 1051–1067. doi:10.1086/422766. PMID 15674775.CS1 bakım: birden çok isim: yazarlar listesi (bağlantı) Ayrıca, yararlı resimlerle bunun açıklamasına da bakınız. Myers, P.Z. (19 Nisan 2005). "Sıcak kanlı timsahlar?". Arşivlenen orijinal 16 Şubat 2008.
- ^ Hutchinson, J.R. (Mart – Nisan 2006). "Archosaurlarda hareketin evrimi". Rendus Palevol Comptes. 5 (3–4): 519–530. doi:10.1016 / j.crpv.2005.09.002.
Dış bağlantılar
- Giganotosaurus'un Termofizyolojisi ve Biyolojisi: Tyrannosaurus ile Karşılaştırma RE Barrick ve WJ Showers (1999) tarafından
- Bir Dinozorun Kalbi Bulundu Bildirildi
- Timsahın evrimi kalp ısıtıcı değil
Fizyoloji türleri | ||
|---|---|---|
| Hayvanlar |  | |
| Bitkiler | ||
| Hücreler | ||
| İlgili konular | ||