Archosaur - Archosaur - Wikipedia
Bu makale için ek alıntılara ihtiyaç var doğrulama. (Aralık 2020) (Bu şablon mesajını nasıl ve ne zaman kaldıracağınızı öğrenin) |
| Archosaurlar | |
|---|---|
 | |
| Kuşlar ve timsahlar (bu durumda sarı gagalı leylek ve bir Nil timsahı ) bilinen tek yaşayan archosaur gruplarıdır. | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Sınıf: | Reptilia |
| Clade: | Eucrocopoda |
| Clade: | Archosauria Başa çıkmak, 1869 |
| Alt gruplar | |
| |
| Eş anlamlı | |
| |
Archosauria ("yönetici sürüngenler") bir clade nın-nin diyapsitler, ile kuşlar ve timsahlar yaşayan tek temsilci olarak. Archosaurlar genel olarak şöyle sınıflandırılır: sürüngenler, kuşları içeren kladistik anlamda. Soyu tükenmiş arkozorlar kuş olmayanları içerir dinozorlar, pterozorlar ve soyu tükenmiş timsah akrabaları. Modern paleontologlar Archosauria'yı bir taç grubu içerir en son ortak ata yaşayan kuşların, timsahların ve tüm soyundan gelenlerin. Archosauria'nın temeli iki sınıfa ayrılır: Pseudosuchia timsahları ve soyu tükenmiş akrabalarını içeren ve Avemetatarsalia kuşları ve soyu tükenmiş akrabalarını (kuş olmayan dinozorlar ve pterozorlar ).[1]
Archosauria grubunun daha eski tanımları, paylaşılan morfolojik gibi özellikler antorbital fenestra kafatasında tırtıklı dişler ve dik bir duruş. Bazı soyu tükenmiş sürüngenler, örneğin proterosuchidler ve euparkeriidler, bu özelliklere sahipti, ancak timsah ve kuş soyları arasındaki bölünmeden önce ortaya çıktı. Bugünlerde Archosauria'nın eski morfolojik tanımı kabaca Archosauriformes, taç grubu archosaurs ve yakın akrabalarını kapsayacak şekilde adlandırılan bir grup.[1] En eski gerçek archosaur fosilleri, Erken Triyas ilk archosauriforms ve Archosauromorphs (sürüngenler kertenkelelere veya diğerlerine göre archosaurlara daha yakın lepidosaurlar ) göründü Permiyen. Archosaurlar, Permiyen-Triyas kitlesel yok oluş (~252 Anne ), en büyük ve ekolojik olarak en baskın kara omurgalıları haline geldi. Orta Triyas kadar süre K-Pg kitlesel yok oluş (~ 66 Ma).[2] Kuşlar ve birkaç krokodilform soylar, K-Pg neslinin tükenmesinden sağ kurtulan ve sonraki yıllarda yeniden çeşitlenen tek archozorlardı. Senozoik çağ. Özellikle kuşlar, günümüzde tür bakımından en zengin kara omurgalıları grupları arasında yer almıştır.
Ayırt edici özellikleri
Archosaurlar geleneksel olarak diğer dörtayaklılardan birkaç temelde ayırt edilebilir. sinapomorfiler ya da ortak özellikler son ortak ata. Bu özelliklerin çoğu, Archosauria klanının başlangıcından önce ortaya çıktı, çünkü bunlar Archosauriforms gibi Proterosuchus ve Euparkeria dışında olan taç grubu.[1]
En belirgin özellikleri derin yuvalara yerleştirilmiş dişlerdir, antorbital ve mandibular fenestra (sırasıyla gözlerin önünde ve çenede açıklıklar),[3] ve belirgin dördüncü trokanter (üzerinde belirgin bir sırt uyluk ).[4] Yuvalara yerleştirilen dişlerin beslenme sırasında gevşeme olasılığı daha düşüktü. Bu özellik adından sorumludur "Thecodont "(" yuva dişleri "anlamına gelir),[5] Erken paleontologların birçok Triyas arozora uyguladığı.[4] Kuşlar gibi bazı archosaurlar ikincil olarak dişsizdir. Antorbital fenestrae, erken dönem arkozorlarda nispeten büyük olan kafatasının ağırlığını, modern timsahlar. Mandibular fenestrae bazı şekillerde çene ağırlığını da azaltmış olabilir. Dördüncü trokanter, femurdaki kasların bağlanması için geniş bir alan sağlar. Daha güçlü kaslar, erken dönem archosaurlarda dik yürüyüşlere izin verdi ve aynı zamanda arkozorların veya onların yakın atalarının felaketten sağ çıkma yetenekleriyle de bağlantılı olabilir. Permiyen-Triyas yok oluş olayı.[kaynak belirtilmeli ]
Kökenleri
Archosaurs bir alt gruptur Archosauriforms, kendilerinin bir alt grubu olan Archosauromorphs. Hem en eski archosauromorph (Protorosaurus speneri ) ve en eski archosauriform (Archosaurus rossicus ) Geç Permiyen'de yaşadı. En eski gerçek arkozorlar, Olenekiyen evresi (247-251 Ma) Erken Triyas. Büyük etobur timsah sıralı archosaurların birkaç parçalı fosili (gayri resmi olarak "rauisuchians ") bu aşamadan bilinmektedir. Bunlar arasında Scythosuchus ve Tsylmosuchus (her ikisi de bulundu Rusya ),[6] yanı sıra Xilousuchus, bir ctenosauriscid itibaren Çin.[1] Kuş sıralı arkozorların bilinen en eski fosilleri, Anisiyen evre (247-242 Ma) Tanzanya ve şunları içerir Asilisaurus (erken Silesaurid ), Teleocrater (bir afanozor ), ve Nyasasaurus (olası bir erken dinozor).[kaynak belirtilmeli ]
Triyas'ta Archosaur'un ele geçirilmesi
Sinapsitler içeren bir sınıftır memeliler ve soyu tükenmiş ataları. Genellikle "memeli benzeri sürüngenler" olarak adlandırılan ancak "olarak adlandırılması gereken ikinci grup"protomamallar," "gövde memelileri, "veya"bazal sinapsitler "çünkü modern tarafından gerçek sürüngenler değiller kladistik sınıflandırma, egemen kara omurgalılarıydı. Permiyen ama en çok telef oldu Permiyen-Triyas yok oluş olayı. Olaydan çok az sayıda büyük sinapsid hayatta kaldı ve bir form, Lystrosaurus (bir otçul Dicynodont ), neslinin tükenmesinden kısa süre sonra yaygın bir dağılıma ulaştı.[kaynak belirtilmeli ] Bunun yerine, arkozorlar ve diğer arkosauriformlar, erken dönemlerde hızla baskın kara omurgalıları haline geldi. Triyas. Kitlesel yok oluştan önceki fosiller yalnızca Ekvator civarında bulundu, ancak olaydan sonra fosiller dünyanın her yerinde bulunabilir.[7] Bunun için en sık önerilen üç açıklama şunlardır:[kaynak belirtilmeli ]
- Archosaurlar, sinapsidlerden daha dik uzuvlara doğru daha hızlı ilerleme kaydetti ve bu onlara kaçınarak daha fazla dayanıklılık verdi. Operatörün kısıtlaması. Bu açıklamaya bir itiraz, archosaurların hala yayılmış veya yarı dik uzuvları varken baskın hale gelmeleridir. Lystrosaurus ve diğer sinapsidler.[kaynak belirtilmeli ]
- Archosaurlar, tek yönlü hava akışına sahip daha verimli solunum sistemlerine sahiptir. Peter Ward, bunun Permiyen sonunda oksijen seviyelerinde şüpheli bir düşüşte avantajlı olduğunu kanıtlamış olabileceğini öne sürüyor.[kaynak belirtilmeli ]
- Erken Triyas çoğunlukla kuraktı, çünkü çoğu Dünya arazisi süper kıta Pangea. Archosaurlar muhtemelen su tasarrufu konusunda erken sinapsidlerden daha iyiydi çünkü:
- Modern diyapsitler (kertenkeleler, yılanlar, timsahlar, kuşlar) salgılar ürik asit Bu, bir macun olarak atılabilir ve daha seyreltik bir idrar yerine düşük su kaybına neden olur. Timsahların, dinozorların ve pterozorların ataları olan archosaurların da ürik asit salgıladıklarını ve bu nedenle suyu korumada iyi olduklarını varsaymak mantıklıdır. Diapsidlerin aglandüler (bezsiz) derileri de suyun korunmasına yardımcı olabilirdi.[kaynak belirtilmeli ]
- Modern memeliler salgılar üre Böbrek tübüllerinde difüzyon yoluyla idrarı terk etmesini önlemek için nispeten yüksek bir idrar hızı gerektirir. Derilerinde ayrıca su kaybeden birçok bez bulunur. Erken sinapsidlerin benzer özelliklere sahip olduğunu varsayarsak, örneğin, yazarların iddia ettiği gibi Palaeos, çoğunlukla kurak bir dünyada dezavantajlı durumdaydılar. Aynı saygın site, "Avustralya'nın çoğu için Pliyo-Pleistosen Koşulların muhtemelen benzer olduğu tarih, en büyük karasal yırtıcılar memeliler değil, devasa Varanid kertenkeleler (Megalanya ) ve toprak timsahları. "[5]
Bununla birlikte, bu teori, sinapsidlerin su tutma konusunda zorunlu olarak daha az avantajlı olduklarını, sinapsid düşüşünün iklim değişiklikleri veya archosaur çeşitliliği ile çakıştığını (ikisi de test edilmedi) ve çölde yaşayan memelilerin bu bölümde de iyi adapte edildiği gerçeğini ima ettiği için sorgulanmıştır. archosaurs olarak[8] ve bazı sinodontlar gibi Trukidosinodon büyük boyutlu yırtıcılardı.[9]
Ana formlar


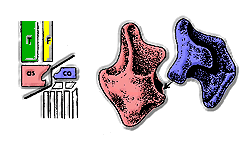
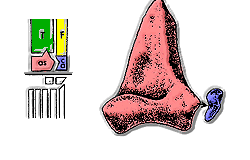
1970'lerden bu yana, bilim adamları, arkozorları esas olarak ayak bileklerine göre sınıflandırdılar.[10] En eski archosaurların "ilkel mezotarsal" ayak bilekleri vardı: astragalus ve kalkaneum sabitlendi tibia ve fibula tarafından dikişler ve eklem bu kemikler ile ayak arasındaki temas etrafında eğildi.
Pseudosuchia erken ortaya çıktı Triyas. Ayak bileklerinde, astragalus tibiaya bir dikiş ve eklem, kalkaneumdaki bir yuvaya takılan astragalus üzerindeki bir çivi etrafında dönüyordu. İlk "crurotarsans" hala genişleyen uzuvlarla yürüyorlardı, ancak daha sonra bazı krurotarsanlar tamamen dik uzuvlar geliştirdiler. Modern timsahlar, hareket hızına bağlı olarak uzuvları genişleyerek veya dik olarak yürüyebilen krurotarsanlardır.[kaynak belirtilmeli ]
Euparkeria ve Ornithosuchidae kalkaneumda bir çivi ve astragalusta bir yuva ile "ters krurotarsal" ayak bilekleri vardı.
En eski fosiller Avemetatarsalia ("kuş bilekleri"), Anisiyen geç yaş Triyas. Ornithodiran'ların çoğunun "ileri mezotarsal" ayak bilekleri vardı. Bu ayak bileği formu, çok büyük bir astragalus ve çok küçük kalkaneum içeriyordu ve basit bir menteşe gibi yalnızca bir düzlemde hareket edebiliyordu. Sadece uzuvları dik olan hayvanlar için uygun olan bu düzenleme, hayvanlar koşarken daha fazla denge sağlıyordu. Teleocrater ve Asilisaurus gibi ilk avemetatarsalılar, "ilkel mezotarsal" ayak bileklerini korudular. Ornitoviranlar, diğer arsozorlardan başka yönlerden farklıydılar: Hafifçe inşa edilmişlerdi ve genellikle küçüklerdi, boyunları uzundu ve S şeklinde bir eğriye sahiptiler, kafatasları çok daha hafif yapılıydı ve birçok ornitovir tamamen iki ayaklı. Femur üzerindeki archosaurian dördüncü trokanter, ornitoviranların iki ayaklı olmalarını kolaylaştırmış olabilir çünkü uyluk kasları için daha fazla kaldıraç sağlıyordu. Triyas'ın sonlarında, ornitodiranlar üretmek için çeşitlendi dinozorlar ve pterozorlar.
Sınıflandırma
Modern sınıflandırma
Archosauria normalde bir taç grubu Bu, yalnızca yaşayan temsilcilerinin son ortak atalarının torunlarını içerdiği anlamına gelir. Archosaurlar söz konusu olduğunda bunlar kuşlar ve timsahlardır. Archosauria daha büyük sınıfta Archosauriformes gibi bazı yakın akrabalarını içeren Proterochampsids ve euparkeriidler. Bu akrabalar, Archosauria taç grubunun dışında yer almalarına rağmen, genellikle archosaurs olarak anılır. baz alınan Archosauriformes içinde konum.[11] Tarihsel olarak, birçok archosauriform, aşağıdakiler de dahil olmak üzere, arkozorlar olarak tanımlandı. proterosuchidler ve eritrosuchidler, bir antorbital fenestranın varlığına göre. Birçok araştırmacı, Archosauria'yı sıralanmamış olarak görmeyi tercih ederken clade bazıları buna geleneksel bir biyolojik sıralama atamaya devam ediyor. Archosauria geleneksel olarak bir Üst Sipariş olarak görülse de, birkaç 21. yüzyıl araştırmacısı onu Bölüm dahil olmak üzere farklı kademelere atadı.[12] ve Sınıf.[13]
Sınıflandırma tarihi
Archosauria bir terim olarak ilk olarak Amerikalı paleontolog tarafından icat edildi Edward Drinker Cope 1869'da ve dahil olmak üzere geniş bir takson yelpazesini içeriyordu dinozorlar, timsahlar, kodonlar, Sauropterygians (kaplumbağalarla ilgili olabilir), Rhynchocephalians (Cope'a göre bir grup dahil) gergedanlar, günümüzde daha bazal olarak kabul edilen Archosauromorphs, ve Tuataras, hangileri lepidosaurlar ), ve Anomodontlar şimdi sinapsid olarak kabul edilenler.[14] 1986 yılına kadar, Archosauria bir taç kuşağı olarak tanımlanmadı ve kullanımını daha fazlasıyla sınırladı. türetilmiş takson.[15]


Cope'un terimi Yunanca-Latince idi melez kafatası kemerlerini kastetmesi amaçlanmıştır, ancak daha sonra Yunanca ile birlikte "önde gelen sürüngenler" veya "hüküm süren sürüngenler" olarak da anlaşılmıştır. ἀρχός "lider, hükümdar".[16]
Artık eski bir terim olarak kabul edilen "thecodont" terimi ilk olarak İngiliz paleontolog tarafından kullanılmıştır. Richard Owen 1859'da Trias dönemine ait arozorları tanımlamak için kullanıldı ve 20. yüzyılda yaygın olarak kullanıldı. Kodonlar, daha gelişmiş arkozorların indiği "temel stok" olarak kabul edildi. Daha sonraki kuş ve timsah soylarında görülen özelliklere sahip değillerdi ve bu nedenle iki grup için daha ilkel ve ataları olarak kabul edildi. İle kladistik devrim 1980'lerin ve 90'ların kladistik organizmaları sınıflandırmada en yaygın kullanılan yöntem haline geldi, kodonlar artık geçerli bir gruplama olarak kabul edilmiyordu. "Bazal stok" olarak kabul edildikleri için kodonlar parafiletik Bu, son ortak atasının tüm soyundan gelenleri içermeyen bir grup oluşturdukları anlamına gelir: bu durumda, daha fazla türetilmiş timsahlar ve kuşlar, daha önce anlaşıldığı gibi "Thecodontia" dan hariç tutulur. Bazalin açıklaması ornitolar Lagerpeton ve Lagosuchus 1970'lerde kodonları dinozorlarla ilişkilendiren kanıtlar sağladı ve birçok kladistin yapay bir gruplama olarak gördüğü "Thecodontia" teriminin kullanılmamasına katkıda bulundu.[17]
"Crocodilian normal" ve "crocodilian reversed" ayak bileklerinin tanımlanmasıyla Sankar Chatterjee 1978'de Archosauria'da bir bazal bölünme tespit edildi. Chatterjee bu iki grubu "normal" ayak bileği ile Pseudosuchia ve "ters" ayak bileği ile Ornithosuchidae olarak kabul etti. Ornitosuchidlerin bu dönemde dinozorların atası olduğu düşünülüyordu. 1979'da, A.R.I. Cruickshank bazal bölünmeyi tespit etti ve krurotarsan ayak bileğinin bu iki grupta bağımsız olarak, ancak zıt yönlerde geliştiğini düşündü. Cruickshank ayrıca, bu ayak bileği tiplerinin gelişiminin, ileri düzey üyelerin yarı dik (timsahlar durumunda) veya dik (dinozorlar durumunda) yürüyüşlere sahip olmalarına izin vermek için her grupta ilerlediğini düşünüyordu.[17]
Filogeni
Çoğunda filogenetik analizler, arkozorların bir monofiletik gruplama, böylece gerçek bir sınıf oluşturur. Archosaur soyoluşunun ilk çalışmalardan biri Fransız paleontolog tarafından yazılmıştır. Jacques Gauthier 1986'da. Gauthier, Archosauria'yı ikiye böldü. Pseudosuchia timsah hattı ve Ornitosuchia, dinozor ve pterosaur hattı. Pseudosuchia, timsahlarla daha yakın akraba olan tüm arkozorlar olarak tanımlanırken, Ornithosuchia, kuşlarla daha yakın akraba olan tüm archosaurlar olarak tanımlandı. Proterochampsids, eritrosuchidler ve proterosuchidler, ortaya çıkan ağaçta Archosauria'nın dışına art arda düştü. Aşağıda kladogram Gauthier'den (1986):[18]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
1988'de paleontologlar Michael Benton ve J.M. Clark, bazal arkozorların filogenetik çalışmasında yeni bir ağaç üretti. Gauthier'in ağacında olduğu gibi, Benton ve Clark'ınki de Archosauria'da bir bazal bölünmeyi ortaya çıkardı. İki gruba Crocodylotarsi ve Ornithosuchia olarak atıfta bulundular. Crocodylotarsi bir apomorphy "timsah-normal" ayak bileği ekleminin varlığına dayalı olan takson (kladın tanımlayıcı apomorfisi olarak kabul edilir). Gauthier'in Pseudosuchia'sı, aksine, bir sap temelli takson. Gauthier'in ağacının aksine, Benton ve Clark'ın yerleri Euparkeria Ornithosuchia dışında ve Archosauria taç grubunun tamamen dışında.[19]
Crurotarsi ve Ornithodira klanları ilk olarak 1990 yılında paleontolog tarafından birlikte kullanılmıştır. Paul Sereno ve A.B. Arcucci, archosaurs filogenetik çalışmalarında. Ornithodira, 1986'da Gauthier tarafından seçildi. Crurotarsi ve Ornithodira, sırasıyla, Pseudosuchia ve Ornithosuchia'nın yerini alırken, Crurotarsi sınıfını ilk kuranlar oldular.[17][20] Sereno ve Arcucci, analizlerinde ayak bileği tipleri dışındaki archosaur özelliklerini de dahil ederek önceki analizlerden farklı bir ağaç ortaya çıkardı. Aşağıda, Sereno ve Arcucci tarafından üretilene benzeyen Sereno (1991) 'e dayanan bir kladogram bulunmaktadır:[17]
| Archosauriformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ornithodira ve Crurotarsi'nin ikisi de düğüm tabanlı sınıflar, yani bunları içerecek şekilde tanımlandıkları anlamına gelir son ortak ata iki veya daha fazla takson ve soyundan gelenlerin tümü. Ornithodira, pterozorların ve dinozorların (kuşlar da dahil) son ortak atasını içerirken, Crurotarsi, yaşayan timsahların son ortak atasını ve Trias dönemine ait üç arkozor grubunu içerir: ornitoşidler, Aetosaurlar, ve Fitosaurlar. Bu sınıflar, "kuş çizgisi" ve "timsah çizgisi" arkozorlarına eşdeğer değildir. şube bazlı sınıflar, bir canlı gruba (kuşlar veya timsahlar) diğerinden daha yakından ilişkili tüm taksonlar olarak tanımlanır.
Benton, 1999 yılında Avemetatarsalia adını tüm kuş sıralı arkozorları içerecek şekilde önerdi (onun tanımına göre, tüm arkozorlar timsahlardan çok dinozorlarla daha yakından ilgilidir). Küçük Triyas arkozoru analizi Scleromochlus kuş-sıralı archosaurlar içine ama Ornithodira'nın dışına yerleştirdi, yani Ornithodira artık kuş-çizgi archosaurlara eşdeğer değildi. Aşağıda, bu filogeniyi gösteren Benton'dan (2004) değiştirilmiş bir kladogram bulunmaktadır:[15]
| Archosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

İçinde Sterling Nesbitt 2011'in erken dönem arozorlar üzerine monografisi olan filogenetik bir analiz, Archosauria'nın dışına düşen fitosaurlar için güçlü bir destek buldu. Sonraki birçok çalışma bu soyoluşu destekledi. Crurotarsi, fitosaurların dahil edilmesiyle tanımlandığından, fitozorların Archosauria dışına yerleştirilmesi, Crurotarsi'nin tüm Archosauria'yı içermesi gerektiği anlamına gelir. Nesbitt, Pseudosuchia'yı kök temelli bir takson olarak kullanarak timsah sıralı archosaurlar için bir soy adı olarak eski haline getirdi. Aşağıda Nesbitt'ten (2011) değiştirilmiş bir kladogram var:[1]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Yok olma ve hayatta kalma
Crocodylomorphs, pterosaurs ve dinozorlar hayatta kaldı Triyas-Jura neslinin tükenmesi olayı yaklaşık 200 milyon yıl önce, ancak diğer arkozorların nesli Triyas-Jura sınırında veya öncesinde tükenmişti.
Kuş olmayan dinozorlar ve pterozorlar telef oldu Kretase-Paleojen nesli tükenme olayı yaklaşık 66 milyon yıl önce meydana gelen, ancak taç grubu kuşlar (kalan tek dinozor grubu) ve birçok krokodilform hayatta kaldı. Her ikisi de arkozorların torunlarıdır ve bu nedenle kendi başlarına arkozorlardır. filogenetik taksonomi.
Timsahlar (tüm modern timsahlar, timsahlar, ve gharials ) ve kuşlar bugün Holosen. Genel olarak, kuşların tüm kara omurgalılarının en çok türüne sahip olduğu kabul edilir.[kaynak belirtilmeli ]
Archosaur yaşam tarzı
Kalça eklemleri ve hareket

Erken gibi dört ayaklılar erken dönem archosaurlar geniş bir yürüyüşe sahipti çünkü kalça yuvaları yana doğru bakıyordu ve tepelerindeki topuzlar femurlar uyluk kemiği ile aynı hizadaydı. erken ve orta Triyas Bazı archosaur grupları, daha dik bir yürüyüşe izin veren (veya gerekli olan) kalça eklemleri geliştirdi. Bu onlara daha fazla dayanıklılık verdi çünkü Operatörün kısıtlaması yani aynı anda rahatlıkla koşup nefes alabilirler. Bacakların dikleşmesine izin veren iki ana eklem türü vardı:
- Kalça yuvaları yanlara dönüktü, ancak uyluk kemiği üzerindeki topuzlar uyluk kemiğinin geri kalanına dik açıdaydı ve bu nedenle aşağıya dönüktü. Dinozorlar, bu kalça düzenlemesi ile arkozorlardan evrimleşti.
- Kalça yuvaları aşağı dönüktü ve uyluk kemiği üzerindeki topuzlar uyluk kemiği ile aynı hizadaydı. Bu "sütun dik" düzenleme, çeşitli archosaur soylarında bağımsız olarak evrimleşmiş gibi görünmektedir, örneğin "Rauisuchia" (non-crocodylomorph parakrokodilomorflar ) ve ayrıca bazılarında Aetosaurlar.
Dik bir duruşun daha fazla enerji gerektirdiği, bu nedenle daha yüksek bir metabolizma ve daha yüksek bir vücut sıcaklığına işaret edebileceği belirtilmiştir.[21]
Diyet
Çoğu büyük yırtıcılardı, ancak çeşitli soyların üyeleri diğer nişlere doğru çeşitlendi. Aetosaurlar otçullardı ve bazıları geniş zırhlar geliştirdi. Birkaç crocodyliform otoburdu, ör. Simosuchus, Phyllodontosuchus. Büyük crocodyliform Stomatosuchus olabilirdi filtre besleyici. Sauropodomorflar ve ornitorik dinozorlar, beslenme için çeşitli uyarlamalara sahip otoburlardı biyomekanik.
Kara, su ve hava
Archosaurlar esas olarak şu şekilde tasvir edilir: arazi hayvanlar, ancak:
- Birçok fitosaur ve crocodyliform nehirlere ve bataklıklara hükmetti ve hatta denizleri istila etti (örn. teleozorlar, Metriorhynchidae ve Dyrosauridae ). Metriorhynchidae, daha çok yunus gibiydi, kürek benzeri ön ayakları, kuyruk paraziti ve pürüzsüz, zırhsız derileri vardı.
- İki kuşak ornitolar, pterozorlar ve kuşlar, uçucu bir yaşam tarzına adapte olduktan sonra havaya hakim oldular.
Metabolizma
Archosaurların metabolizması hala tartışmalı bir konudur. Kesinlikle soğukkanlı atalardan evrimleştiler ve hayatta kalan dinozor olmayan arozorlar, timsahlar soğukkanlı. Ancak timsahlar, normalde sıcak kanlı bir metabolizma ile ilişkilendirilen bazı özelliklere sahiptir, çünkü hayvanın oksijen kaynağını iyileştirirler:
- 4 odacıklı kalpler. Hem kuşlar hem de memeliler, oksijenli ve oksijensiz akışları tamamen ayıran 4 odacıklı kalplere sahiptir. kan. Timsah olmayan sürüngenlerin 3 odacıklı kalpler Bu daha az etkilidir çünkü iki akışın karışmasına izin verirler ve böylece oksijenli kanı akciğerler yerine vücuda gönderirler. Modern timsahların kalpleri 4 odacıklıdır, ancak vücut boyutuna göre daha küçüktür ve modern kuşlara göre daha düşük basınçta çalışır ve memeliler. Ayrıca bir pulmoner baypas, su altındayken işlevsel olarak 3 odacıklı hale getiren oksijen.
- a ikincil damak Bu, hayvanın aynı anda yemek yemesine ve nefes almasına izin verir.
- a hepatik piston pompalama mekanizması akciğerler. Bu, memelilerin ve kuşların akciğer pompalama mekanizmalarından farklıdır, ancak bazı araştırmacıların bazı dinozorlarda bulduklarını iddia ettiklerine benzer.[22][23]
Tarihsel olarak neden olduğuna dair belirsizlik olmuştur. Doğal seçilim aktif sıcakkanlı canlılar için çok önemli olan, ancak soğukkanlı su tuzağına pek az faydası olan bu özelliklerin geliştirilmesini destekledi. avcılar Zamanlarının büyük çoğunluğunu suda yüzerek veya nehir kıyısında yatarak geçirenler?


Paleontolojik kanıt[açıklama gerekli ] yaşayan timsahların atalarının aktif ve endotermik (sıcak kanlı) olduğunu gösterir. Bazı uzmanlar[DSÖ? ] Archosaur atalarının da sıcakkanlı olduğuna inanırlar. Bunun nedeni muhtemelen tüy benzeri filamentlerin tüm vücudu kaplayacak şekilde gelişmesi ve ısı yalıtımı sağlayabilmesidir.[24] Timsah kalbinin fizyolojik, anatomik ve gelişimsel özellikleri, paleontolojik kanıtları destekliyor ve bu soyun sucul, pusu avcı nişini istila ettiğinde ektotermiye geri döndüğünü gösteriyor. Timsah embriyoları erken bir aşamada tamamen 4 odacıklı kalpler geliştirir. Büyüyen kalpte yapılan modifikasyonlar, sol kanadı içeren bir pulmoner baypas şantı oluşturur. aort kemeri sağdan kaynaklanan ventrikül, foramen of Panizza sol ve sağ aort arkları ve tabanındaki dişli kapakçık arasında pulmoner arter. Şant, dalış sırasında kalbin 3 odacıklı kalp olarak işlev görmesini sağlamak için kullanılır ve timsahın ektotermler tarafından kullanılan nöral kontrollü şantı sağlar. Araştırmacılar, yaşayan timsahların atalarının tamamen 4 odacıklı kalplere sahip oldukları ve bu nedenle soğuk kanlı veya ektotermik metabolizmaya dönmeden önce sıcak kanlı oldukları sonucuna vardılar. Yazarlar ayrıca kök arkozorlarda endotermi için başka kanıtlar da sağlıyor.[25][26] Daha sonra timsahların soğukkanlı, suda yaşayan ve daha az aktif hale geldiklerinde pulmoner baypas şantı geliştirdiklerini öne sürmek mantıklıdır.
Timsah ataları ve diğerleri Triyas Archosaurlar sıcakkanlıydı, bu bazı evrimsel bulmacaları çözmeye yardımcı olacaktı:
- İlk krokodilomorflar, ör. Terrestrisuchus, yapıları oldukça aktif bir yaşam tarzı öneren, oldukça hızlı bir metabolizma gerektiren ince, uzun bacaklı karasal yırtıcılardı. Ve diğer bazı crurotarsan archosaurs uzuvları dikmiş gibi görünürken, rauisuchians diğer herhangi bir duruş için çok zayıf bir şekilde uyarlanmıştır. Dik uzuvlar aktif hayvanlar için avantajlıdır çünkü Operatör kısıtlaması, ancak daha halsiz hayvanlar için dezavantajlıdır çünkü ayağa kalkmanın ve yatmanın enerji maliyetlerini arttırırlar.
- Eğer erken dönem arzorlar tamamen soğukkanlıysa ve (büyük ihtimalle göründüğü gibi) dinozorlar en azından oldukça sıcakkanlıydı dinozorlar, geçen sürenin yarısından daha kısa bir sürede sıcak kanlı metabolizmaları evrimleştirmek zorunda kalacaklardı. sinapsitler aynısını yapmak.
Solunum sistemi
Akciğerler üzerine yeni bir çalışma Timsah mississippiensis ( Amerikan timsahı ) içlerindeki hava akışının tek yönlü olduğunu, inhalasyon ve ekshalasyon sırasında aynı yönde hareket ettiğini göstermiştir.[27] Bu aynı zamanda kuşlarda ve kuş olmayan birçok dinozorda da görülür. hava keseciklerinin solunuma daha fazla yardımcı olmak için. Hem kuşlar hem de timsahlar, varlığıyla tek yönlü hava akışı sağlar. parabronchi sorumlu olan gaz takası. Çalışma, timsahlarda havanın ikinciden girdiğini buldu. bronş dalı, parabronşilerden geçer ve ilk bronşiyal daldan çıkar. Hem kuşlarda hem de timsahlarda tek yönlü hava akımı, bu tür solunumun Archosauria'nın tabanında mevcut olduğunu ve hem dinozorlar hem de aetosaurlar, "rauisuchians" (timsah olmayan parakrokodilomorflar), timsahlar ve pardilomorflar gibi dinozor olmayan arkozorlar tarafından korunduğunu göstermektedir.[27] Archosaurların akciğerlerinde tek yönlü hava akışının kullanılması, gruba, havanın sonlandırılan bir bronş ağı yoluyla gelgit olarak içeri ve dışarı hareket ettiği akciğerlere sahip olan sinapsidlere göre bir avantaj vermiş olabilir. alveoller çıkmaz sokaklardı. Archosaur akciğerlerinde görülen gaz transferindeki daha iyi verimlilik, Mesozoik sırasında var olduğu düşünülen düşük atmosferik oksijen zamanlarında avantajlı olabilir.[28]
Üreme
Çoğu archosaurs yumurtlayan. Kuşlar ve timsahlar, soyu tükenmiş dinozorlar ve krokodilomorflar gibi sert kabuklu yumurtalar bırakırlar. Sert kabuklu yumurtalar, hem dinozorlarda hem de timsahlarda bulunur ve bu, yokluğunun bir açıklaması olarak kullanılmıştır. canlılık veya ovoviviparite Archosaurlarda.[29] Ancak, her iki pterosaur da[30] ve Baurusuchids[31] yumuşak kabuklu yumurtalara sahip olması, sert kabuğun plesiomorfik bir durum olmadığını gösterir. Pelvik anatomisi Cricosaurus ve diğeri Metriorhynchids[32] ve archosaur olmayan archosauromorph'a ait fosilleşmiş embriyolar Dinocephalosaurus,[33] birlikte, arkozorlar arasındaki canlılık eksikliğinin soya özgü kısıtlamaların bir sonucu olabileceğini düşündürmektedir.[açıklama gerekli ]
Archosaurlar atadan kalma süper precocial çeşitli dinozorlarda, pterozorlarda ve timsahlarda görüldüğü gibi.[34] Bununla birlikte, ebeveyn bakımı, timsahlarda, dinozorlarda ve dinozorlarda bağımsız olarak birçok kez gelişti. Aetosaurlar.[35] Bu tür türlerin çoğunda hayvanlar yumurtalarını gömer ve sıcaklığa bağlı cinsiyet belirleme. Dikkate değer istisna Neornithes Yumurtalarını kuluçkaya yatıran ve genetik cinsiyet belirlemeye dayanan bu özellik, onlara diğer dinozorlara göre hayatta kalma avantajı sağlayabilecek bir özelliktir.[36]
Referanslar
- ^ a b c d e Nesbitt, S.J. (2011). "Archosaurların erken evrimi: ilişkiler ve ana soyların kökeni" (PDF). Amerikan Doğa Tarihi Müzesi Bülteni. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112. S2CID 83493714.
- ^ Ezcurra, M.N.D .; Scheyer, T. M .; Butler, R.J. (2014). "Sauria'nın Kökeni ve Erken Evrimi: Permiyen Saurian Fosil Kayıtlarının Yeniden Değerlendirilmesi ve Timsah-Kertenkele Ayrışmasının Zamanlaması". PLOS ONE. 9 (2): e89165. doi:10.1371 / journal.pone.0089165. PMC 3937355. PMID 24586565.
- ^ Gareth Dyke; Gary Kaiser, editörler. (2011). Yaşayan Dinozorlar: Modern Kuşların Evrimsel Tarihi. John Wiley & Sons. pp.10. ISBN 978-0470656662.
- ^ a b Khanna, D.R. (2004). Sürüngenlerin Biyolojisi. Discovery Yayınevi. sayfa 78ff. ISBN 978-8171419074.
- ^ a b White, T .; Kazlev, M.A. "Archosauromorpha: Genel Bakış". Palaeos.com. Arşivlenen orijinal 20 Aralık 2010. Alındı 6 Eylül 2012.
- ^ Gower, D. J .; Sennikov, A. G. (2003). "Rusya'dan ilk arkeozlar". Benton, M.J .; Shishkin, M.A .; Unwin, D.M. (eds.). Rusya ve Moğolistan'da Dinozorlar Çağı. Cambridge: Cambridge University Press. s. 140–159.
- ^ "Antarktika'da İguana büyüklüğünde dinozor kuzeni keşfedildi - ScienceDaily".
- ^ Darren Naish, 38.Bölüm: A Not Too Shabby Podcarts Arşivlendi 2016-01-27 de Wayback Makinesi
- ^ Oliveira, T.V .; Soares, M.B .; Schultz, C.L. (2010). "Trucidocynodon riograndensis gen. Nov. Et sp. Nov. (Eucynodontia), Brezilya Üst Triyasından (Santa Maria Formasyonu) yeni bir cynodont". Zootaxa. 2382: 1–71. doi:10.11646 / zootaxa.2382.1.1.
- ^ Archosauromorpha: Archosauria - Palaeos Arşivlendi 2005-04-05 Wayback Makinesi
- ^ Gower, D. J .; Wilkinson, M. (1996). "Bazal archosaur filogenisi konusunda herhangi bir fikir birliği var mı?" (PDF). Royal Society B Tutanakları. 263 (1375): 1399–1406. doi:10.1098 / rspb.1996.0205. S2CID 86610229.
- ^ Benton, M.J. (2005). Omurgalı Paleontoloji, 3. baskı. Blackwell Science Ltd
- ^ Göhlich, U.B .; Chiappe, L.M .; Clark, J.M .; Sues, H.-D. (2005). "Geç Jura dönemine ait olduğu iddia edilen dinozorun sistematik konumu Macelognathus (Crocodylomorpha: Sphenosuchia) ". Kanada Yer Bilimleri Dergisi. 42 (3): 307–321. doi:10.1139 / e05-005.
- ^ Cope, Edward Drinker (1869). "Kuzey Amerika'nın nesli tükenmiş Batrachia, Reptilia ve Aves'lerinin özeti". Amerikan Felsefe Derneği'nin İşlemleri. 14 (1): 1–252. doi:10.2307/1005355. hdl:2027 / nyp.33433090912423. JSTOR 1005355.
- ^ a b Benton, M.J. (2004). "Dinosauria'nın kökeni ve ilişkileri". Weishampel, D.B .; Dodson, P.r; Osmólska, H. (editörler). Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. pp.7 –19. ISBN 978-0-520-24209-8.
- ^ Biyoloji Broşürleri: Kofoid koleksiyonu, cilt. 2900 (1878), s. 731
- ^ a b c d Sereno, P.C. (1991). "Bazal archosaurs: filogenetik ilişkiler ve işlevsel çıkarımlar". Anı (Omurgalı Paleontoloji Derneği). 2: 1–53. doi:10.2307/3889336. JSTOR 3889336.
- ^ Gauthier, J.A. (1986). "Saurischian monophyly ve kuşların kökeni". Padian, K. (ed.). Kuşların Kökeni ve Uçuşun Evrimi. California Bilimler Akademisi'nin Anıları. 8. San Francisco: Kaliforniya Bilimler Akademisi. s. 1–55.
- ^ Benton, M. J .; Clark, J.M. (1985). "Archosaur filogeni ve Crocodylia'nın ilişkileri". Benton, M.J. (ed.). Tetrapodların Filogeni ve Sınıflandırılması. 1. Oxford: Clarendon Press. s. 295–338. ISBN 978-0-19-857712-6.
- ^ Sereno, P. C .; Arcucci, A.B. (1990). "Crurotarsal archosaurs monophyly ve kuş ve timsah ayak bileği eklemlerinin kökeni". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 180: 21–52.
- ^ Desmond, Adrián J., Sıcakkanlı dinozorlar: paleontolojide bir devrim. 1976, Dial Press, sayfa 87.
- ^ Ruben, J .; et al. (1996). "Bazı Geç Kretase dinozorlarının metabolik durumu". Bilim. 273 (5279): 120–147. Bibcode:1996Sci ... 273.1204R. doi:10.1126 / science.273.5279.1204. S2CID 84693210.
- ^ Ruben, J .; et al. (1997). "Theropod dinozorlarında ve erken kuşlarda akciğer yapısı ve havalandırma". Bilim. 278 (5341): 1267–1270. Bibcode:1997Sci ... 278.1267R. doi:10.1126 / science.278.5341.1267.
- ^ Kişiler, Walter S .; Currie, Philip J. (2015). "Tüyler aşağıdan önce: Tüylerin işlevsel kökenine yeni bir bakış açısı". Evrim. 69 (4): 857–862. doi:10.1111 / evo.12634. ISSN 1558-5646. PMID 25756292. S2CID 24319963.
- ^ Seymour, R. S .; Bennett-Stamper, C. L .; Johnston, S. D .; Carrier, D.R. ve Grigg, G.C. (2004). "Archosaur evriminin kökünde timsahların endotermik atalarının kanıtı" (PDF). Physiol. Biochem. Zool. 77 (6): 1051–1067. doi:10.1086/422766. hdl:2440/1933. PMID 15674775. S2CID 10111065.[kalıcı ölü bağlantı ]
- ^ Summers, A.P. (2005). "Evrim: Sıcak kalpli timsahlar". Doğa. 434 (7035): 833–834. Bibcode:2005Natur.434..833S. doi:10.1038 / 434833a. PMID 15829945. S2CID 4399224.
- ^ a b Farmer, C. G .; Sanders, K. (2010). "Timsahların akciğerlerinde tek yönlü hava akışı". Bilim. 327 (5963): 338–340. Bibcode:2010Sci ... 327..338F. doi:10.1126 / science.1180219. PMID 20075253. S2CID 206522844.
- ^ Lisa Grossman (14 Ocak 2010). "Timsahlar kuşlar gibi nefes alır". Bilim Haberleri. Alındı 14 Ocak 2010.
- ^ Robin M. Andrews; Tom Matematik (2000). "Sürüngen Gelişiminin Doğal Tarihi: Canlılığın Evrimi Üzerindeki Kısıtlamalar". BioScience. 50 (3): 227–238. doi:10.1641 / 0006-3568 (2000) 050 [0227: NHORDC] 2.3.CO; 2.
- ^ Ji, Q; Ji, SA; Cheng, YN; et al. (Aralık 2004). ""(Aralık 2004). "Paleontoloji: kösele kabuklu pterozor yumurtası". Doğa. 432 (7017): 572. doi:10.1038 / 432572a. PMID 15577900. S2CID 4416203.
- ^ Oliveira, C.E.M .; Santucci, R.M .; Andrade, M.B .; Fulfaro, V.J .; Basílo, J.A.F .; Benton, M.J. (2011). "Adamantina Formasyonundan (Bauru Grubu), Brezilya Üst Kretase'sinden timsahlar ve yumurta kabukları". Paleontoloji. 54 (2): 309–321. doi:10.1111 / j.1475-4983.2010.01028.x.
- ^ Herrera, Yanina; Fernández, Marta S .; Lamas, Susana G .; Campos, Lisandro; Talevi, Marianella; Gasparini, Zulma (2017/02/01). "Sakral bölgenin morfolojisi ve Metriorhynchidae'nin üreme stratejileri: karşı indüktif bir yaklaşım". Edinburgh Kraliyet Topluluğu'nun Dünya ve Çevre Bilimleri İşlemleri. 106 (4): 247–255. doi:10.1017 / S1755691016000165. ISSN 1755-6910.
- ^ Liu, Haz; Organ, Chris L .; Benton, Michael J .; Brandley, Matthew C .; Aitchison Jonathan C. (2017/02/14). "Archosauromorph sürüngeninde canlı doğum". Doğa İletişimi. 8: 14445. Bibcode:2017NatCo ... 814445L. doi:10.1038 / ncomms14445. ISSN 2041-1723. PMC 5316873. PMID 28195584.
- ^ Mark P. Witton (2013), Pterosaurs: Natural History, Evolution, Anatomy, Princeton University Press, ISBN 978-0-691-15061-1
- ^ Avanzini, M .; Dalla; Mietto, P; Piubelli, D; Preto, N; Rigo, M; Roghi, G (2007). "Kuzeydoğu İtalya'daki bir omurgalı yuvalama alanı, geç Karnaval sürüngenleri için beklenmedik şekilde karmaşık davranışlar ortaya koyuyor". PALAIOS. 22 (5): 465–475. doi:10.2110 / palo.2005.p05-137r. S2CID 131332250.
- ^ Tanaka, Kohei (2015). "Yumurta Kabuğu Gözenekliliği Dinozorlarda Yuvalanmanın Evrimi Hakkında Bilgi Sağlıyor". PLOS ONE. 10 (11): e0142829. Bibcode:2015PLoSO..1042829T. doi:10.1371 / journal.pone.0142829. PMC 4659668. PMID 26605799.
Kaynaklar
- Benton, M.J. (2004). Omurgalı Paleontoloji (3. baskı). Blackwell Science.
- Carroll, R.L. (1988). Omurgalı Paleontoloji ve Evrim '. New York: W. H. Freeman.
Dış bağlantılar
- UCMP
- Paleos Archosaur filogenisinin (soy ağacı) karmaşık tarihini inceler ve çeşitli archosaur ayak bileği tiplerinin mükemmel bir görüntüsüne sahiptir.
- Mikko'nun Filogeni Arşivi Archosauria
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||







{kind=link}