Eolambia - Eolambia
| Eolambia | |
|---|---|
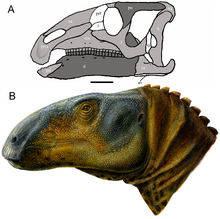 | |
| Kafatası rekonstrüksiyonu (A) ve yaşam onarımı (B) | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Sipariş: | †Ornithischia |
| Alttakım: | †Ornitopoda |
| Üst aile: | †Hadrosauroidea |
| Clade: | †Hadrosauromorpha |
| Cins: | †Eolambia Kirkland, 1998 |
| Türler | |
| †Eolambia caroljonesa Kirkland, 1998 | |
Eolambia ("şafak" anlamına gelir lambeosaurine ") bir cins otçul hadrosauroid Dinozor erkenden Geç Kretase of Amerika Birleşik Devletleri. Tek bir tür içerir, E. caroljonesa, tarafından adlandırıldı paleontolog James Kirkland 1998 yılında. tip numune nın-nin Eolambia 1993 yılında Carole ve Ramal Jones tarafından keşfedildi; tür adı Carole'u onurlandırır. O zamandan beri, hem yetişkinlerden hem de gençlerden, iskeletin neredeyse her unsurunu temsil eden yüzlerce kemik keşfedildi. Şimdiye kadar tüm numuneler bulundu Emery County, Utah Mussentuchit Üyesi olarak bilinen bir kaya katmanında Sedir Dağı Oluşumu.
6 metre (20 ft) uzunluğa kadar ölçüm, Eolambia grubunun büyük bir üyesidir. Asya hadrosauroidlerine yakından yaklaşırken Equijubus, Probactrosaurus, ve Choyrodon kafatasının özelliklerinde, omur ve uzuvlar, aslında Kuzey Amerika ile daha yakından ilişkili olabilir. Protohadros. Bu gruplama, dörtlü kemik ve kürek kemiği izole edilmiş bir endemik radyasyon hadrosauroidler. Benzemesine rağmen hadrosauridler - özellikle lambeosaurine hadrosauridler - çeşitli özelliklerinde, bunlardan biri olarak ilk tanımlanmasına yol açan bu benzerlikler, tamamen reddedildi. yakınsak veya yanlış yorumlanmış.
Eolambia göllerin kenarındaki ormanlık bir ortamda nemli bir ortamda yaşardı taşkın yatağı çevre, beslenmek jimnospermler, eğrelti otları, ve çiçekli bitkiler. Göllerdeki su seviyeleri zamanla, suların neden olduğu döngüsel ıslak ve kuru büyülerle değişti. devinim Mussentuchit Üyesinin çökeltilerinde değişen bantlar tarafından yansıtılan Dünya'nın. Bir çocuk olarak, Eolambia büyükler tarafından avlanırdı krokodilomorflar göl sularında ikamet ediyor. Yaş ilerledikçe, timsahlara karşı dayanıklı hale geldiler ve olgun bireyler (en az sekiz ila dokuz yaşları arasında) büyükler tarafından avlandılar. theropodlar benzeri neovenatorid Siats.
Keşif tarihi
İlk keşifler ve açıklama

1979'da Peter Galton ve James A. Jensen parçalı bir sağ femur tanımladı, BYU 2000, bir hadrosauriyen dinozorun ait olduğu çökeltilerde keşfedildi Sedir Dağı Oluşumu içinde Arches Ulusal Parkı, Utah. Zayıf materyal olmasına rağmen, onun için önemliydi (makalede anlatılan ikinci bir Kuzey Amerika uyluk kemiğinin yanı sıra), ilk hadrosaur örneğiydi. Alt Kretase dünyanın herhangi bir yerinde. Galton ve Jensen, gelecekte oluşumdan bir hadrosaurun daha eksiksiz kalıntılarının bulunabileceğini varsaydılar.[1] Batı bölgesindeki küçük omurgalıların ocaklarında da çeşitli hadrosauroid dişler bulunmuştur. San Rafael Swell, yakın Dale Kalesi içinde Emery County, Utah; 1991 yılında J. Michael Parrish tarafından tanımlanmışlardır. Ardından, 1993 yılında, Carole Jones ve eşi Ramal Jones, Swell'in kuzeybatı bölgesinde bulunan bir fosil alanında parçalanmış kemikler keşfettiler. Siteyi, kurumun yöneticisi Donald Burge'nin dikkatine sundular ve daha sonra Doğu Utah Prehistorik Müzesi Koleji (CEUM). Resmi olarak CEUM Locality 42em366v olarak bilinen site, daha sonra Carol'ın Sitesi (sic ) onuruna. CEUM 9758 numaralı numune altında saklanan fosiller, kafatası, omurlar, iskiyum ve bacak parçaları dahil olmak üzere yetişkin bir hadrosauroidin kısmi kalıntılarını temsil ediyor. Bir yetişkinin kısmi kafatası ve ön ayağı olan CEUM 5212, yakınlarda CEUM Locality 42em369v'de bulundu.[2] Bir yetişkinin sol uyluk kemiği olan CEUM 8786, Carol'un Sitesinde daha sonra keşfedildi ve 2012'ye kadar tanımlanmadı.[3]
Aynı zamanda Oklahoma Doğa Tarihi Müzesi (OMNH), Swell'in güneybatı bölgesindeki kazılarda aynı hadrosauroide ait kalıntıları bulmuştur.[2] OMNH tarafından keşfedilen örnekler başlangıçta beş bölgeden altı kişiden oluşuyordu: iki genç iskelet,[4] OMNH v237 lokalitesinden omurga, kürek kemiği, ulna, ilium ve iskiyum ve arka bacak elemanları dahil; OMNH v824'ten bir sırt omuru ile birlikte kafatasının parçaları, ön ayaklar ve arka bacakları içeren bir kısmi çocuk iskeleti; OMNH v696'dan bir sakrum ve iskiyum olan OMNH 27749; OMNH v214'ten bir iskiyum olan OMNH 24389; ve OMNH 32812, bir kürek kemiği, iki kaudal omur ve OMNH v866'dan kazılmamış diğer elemanları içeren kısmi bir iskelet. OMNH ayrıca çeşitli taş ocaklarında hadrosauroidin dişlerine ilişkin başka keşifler de yaptı. OMNH'den Richard Cifelli izin verdi James Kirkland toplanan örnekleri incelemek için.[2] Kirkland, 1994'teki örnekleri, yıllık toplantısında bir konferans özetinde ön olarak tanımlamaya devam etti. Omurgalı Paleontoloji Derneği.[5]

Bültende yayınlanan 1998 tarihli bir makalede New Mexico Doğa Tarihi ve Bilim Müzesi Kirkland, örnekleri resmen yeni bir cins, Eolambia, ile tip ve sadece türler Eolambia caroljonesa. Genel ad, Yunan önek eos/ ἠώς ("şafak", "sabah", "erken" anlamına gelen) son ek ile Lambia (elde edilen Lambeosaurus, daha sonra adını alan Kanadalı paleontolog Lawrence Lambe ). Genel olarak, cins adı "şafak (veya erken) anlamına gelir lambeosaurine ", en bazal lambeosaurin olduğu varsayılan konumuna referansla. Bu arada, belirli isim Carole Jones'u onurlandırıyor. İsim Eolambia tarafından önerildi paleoartist Michael Skrepnick; 1998 tarihli gazeteden önce Kirkland tarafından kullanılan gayri resmi adı "Eohadrosaurus caroljonesi" nin yerini aldı.[2]
Sonraki keşifler

1998'den beri yapılan ek kazılar, Eolambia. Şu anda en çok bilinen iguanodont. Senomaniyen ve Santoniyen aşamalar. Taksona ait hemen hemen her iskelet öğesi, mevcut koleksiyonlarda çok sayıda fosille temsil edilir. En az sekizi çocuk olmak üzere en az on iki kişi,[6] kasabasının güneyindeki bir taş ocağında keşfedildi. Zımpara hem kafatasından hem de vücudun geri kalanından parçalar dahil. Taş ocağı Cifelli # 2 olarak bilinir Eolambia caroljonesa Taş ocağı veya daha resmi olarak CEUM yerelliği 42em432v. Emery kasabasının güneyindeki bir başka taş ocağı - Willow Springs 8 ocağı veya CEUM mevkii 42Em576v - en az dört genci barındırmaktadır. Bu taş ocaklarından elde edilen malzemeler 2012 yılında Kirkland tarafından Andrew McDonald, John Bird ve Peter Dodson ile birlikte tanımlandı. Teşhis için Eolambia bu malzemeye göre.[3]
CEUM 34447 olarak kataloglanmış bir sağ diş hekimi olan Cifelli # 2 ocağında bulunan bir örnek, diğerlerine kıyasla sıra dışıdır. Eolambia hem yetişkinler hem de gençler dahil olmak üzere örnekler. Orta nokta derinliği ve diğer bireylerde diş hekimlerinin toplam uzunluğu arasındaki oran genellikle 0,19 ile 0,24 arasında değişirken, CEUM 34447'de aynı değer 0,31'dir ve bu da onu alışılmadık derecede derin yapar. Diş hekimi aynı zamanda ön uçta önemli ölçüde derinleşmez ve bu nedenle, diş hekimliği ayırt edici özelliklerinden birinden yoksundur. Eolambia diş hekimleri. Bununla birlikte, koronoid işlemin ön ucunun genişlemesi olan diğer ayırt edici özellik numunede mevcuttur. Bu kişinin alışılmadık bir şeyi temsil etmesi mümkün olsa da Eolambia McDonald ve meslektaşları, örnek olarak, farklı bir hadrosauroidi temsil edebileceği konusunda uyardı. morfoloji diş hekiminin Eolambia (taş ocağındaki diğer malzemelerin hiçbiri tarafından önerilmese de). Bu nedenle, onu belirsiz bir bazal hadrosauroid olarak kabul ettiler.[3]

Saha Doğa Tarihi Müzesi (FMNH) ayrıca 2009'dan 2010'a kadar Emery kasabasının güneyinde kazılar yaptı. Utah Jeolojik Etüt. Bu kazılar 2008 yılında Akiko Shinya tarafından keşfedilen ve onuruna Akiko'nun Yeri olarak adlandırılan FMNH mevkii UT080821-1'de gerçekleştirildi. Kazılar sırasında, 167 disartiküle edilmiş ancak yakından ilişkili eleman iki alçı ceket içinde toplanmıştır. Kalça ve omurlardan öğeler içeren bu alçı ceketlerden birine FMNH PR 3847 numune numarası verildi; numune daha sonra 2017'de Terry Gates, Lindsay Zanno ve Peter Makovicky ile birlikte McDonald tarafından tanımlandı. FMNH tarafından keşfedilen bir başka yer de, Üç Tepeli taş ocağı, FMNH mahalli UT130904-2, ait 400'den fazla genç kemik en az dört kişiye.[7]
Jeoloji ve tarihleme

Tüm yerellikler Eolambia Mussentuchit Üyesine ait olduğu tespit edildi. Sedir Dağı Oluşumu. Mussentuchit'teki baskın kayaçlar, aralıklı açık gri ila gri renkli katmanlardan oluşur. Bulanık kumtaşı ve çamurtaşı. İki ayrı temsil eden mevduatlar nehir kanalı sistemler de Mussentuchit'te mevcuttur. Bu sistemlerin alt kısmı sırasıyla çamurlu kumtaşı, ince taneli kumtaşı olmak üzere dört katmandan oluşur. çamurlu çamurtaşı ve ince taneli kumtaşı; son iki katmanda bitki artıkları bulunur. Cifelli # 2 Ocağı bu sistemle ilişkilidir. Üst sistem yeşilimsi ince taneli kumtaşından oluşur. Üç katman bentonit kil temsil eden katmanlar volkanik kül Mussentuchit'te sırasıyla beyazımsı gri, sarı ve ten rengi olan tortular da mevcuttur. Bu katmanların ortası Cifelli # 2 Ocağı'nı kesiyor.[6] Bentonit, Mussentuchit boyunca kumtaşı içinde de mevcuttur.[7] Cifelli # 2 taş ocağında, kemiklerin kendileri kararmış, kalsit, kuvars, pirit, ve karbonat -hidroksiapatit.[6]
Palinoloji (polen çalışması),[8][9][10] yanı sıra fosil çalışmaları megaflora ve omurgasızlar,[11] başlangıçta, Mussentuchit de dahil olmak üzere en üstteki Sedir Dağı Formasyonunun, Albiyen sahne.[2] Bir 1997 argon-argon yaş tayini nın-nin sanidin Cifelli ve meslektaşları tarafından üst Sedir Dağı Formasyonundaki bir volkanik kül tabakasından elde edilen kristaller, 97 ± 0.1 milyon yıllık bir yaşı gösterdi; bu, daha sonra Albiyen ve Senomaniyen aşamaları arasındaki sınırdan hemen sonra kabul edildi. Bu, üstte yatan için bildirilen erken Senomaniyen yaşına benzer. Dakota Formasyonu.[12] Daha sonra, 2007'de James Garrison ve meslektaşları, muhtemelen Cifelli ve meslektaşları tarafından daha önce tarihlendirilen aynı katman olan Cifelli # 2 Taş Ocağından geçen volkanik kül tabakasını 96.7 ± 0.5 milyon yıl öncesine tarihlendirdiler. Yaklaşık bir düzeyde, Cifelli ve Garrison tahminleri birbirleriyle aynı fikirde olduğundan, Eolambia. Garrison ve meslektaşları ayrıca, Mussentuchit'in her yerinden gelen polen topluluğunu yeniden analiz ederek, topluluğun Albian-Senomanian sınırını kapsadığı sonucuna vardılar.[6]
Açıklama

Eolambia büyük hadrosauroid. İlk tahminler kafatasının uzunluğunu 1 metre (3 ft 3 inç) uzunluğunda yerleştirdi,[2] bunun nedeni orantısız bir şekilde uzun bir burun olmasına rağmen, daha sonra ek materyallerin keşfi ile düzeltildi.[3] 2016 yılında Gregory S. Paul 6 metre (20 ft) vücut uzunluğu ve 1 metrik ton (0.98 uzun ton; 1.1 kısa ton) ağırlık tahmini Eolambia,[13] önceki vücut uzunluğu tahmini olan 6,1 metre (20 ft) ile uyumludur. Thomas R. Holtz Jr. 2012 yılında.[14] Daha önce, 2008'de, yetişkin bir örneğin 5,2 metre (17 ft) uzunluğunda ve 2 metre (6 ft 7 inç) yüksekliğinde olduğu tahmin ediliyordu.[6]
Kafatası

Tepesiz kafatası Eolambia genel olarak benzer bir şekle sahiptir Equijubus ve Probactrosaurus. Burnun ön kısmı oldukça pürüzlüdür ve birçok kişi tarafından delinmiştir. foramina (açıklıklar). Her birinin ucunda premaksilla En yakın akrabasında da görülen, diş etleri olarak bilinen iki diş benzeri yapı vardır. Protohadros. Daha geride, premaxilla'nın alt dalının arka kısmı aniden yukarı doğru çıkıntı yaparak, arka kısımdaki burun deliğini, Probactrosaurus, Protohadrosve diğer hadrosauroidler. Bu bölüm, iki parmak benzeri işlemle birleşir. üst çene benzer olan Protohadros. Üst çenenin gövdesi bir girinti veya herhangi bir belirti taşımaz. antorbital fenestra, sevmek Equijubus, Protohadrosve diğer hadrosauroidler. Ayırt etmek için kullanılan özelliklerden biri Eolambia yandan bakıldığında üst çenenin diş sırasının içbükey profilidir. Equijubus, Probactrosaurusve birkaç diğer Hadrosauriforms ama aksine Protohadros.[3]
Sevmek Probactrosaurus ve diğer hadrosauroidler, üst çenenin arkası cüce - göz yuvasının altını sınırlayan ve zamansız fenestra - bir girintiye uyan parmak benzeri bir çıkıntı aracılığıyla. Jugalin alt kenarı, infratemporal fenestranın seviyesinin altında güçlü bir çıkıntı taşır; bu aynı zamanda Equijubus, Probactrosaurus, Protohadrosve diğer birkaç hadrosauroid. Jugale yukarıdan bağlanmak, poztorbital göz çukurlarını çevreleyen pürüzlü bir yüzeye sahip olan (gibi Protohadros), ancak kemiğin yanı aksi takdirde pürüzsüzdür. Kafatasının arkasında uydurmak ile ifade eder skuamozal üstten bakıldığında D şeklinde bir eklem ile. Sol ve sağ skuamozaller birbirleriyle yoğun bir şekilde temas etmiş, sadece arkalarında küçük bir işlemle ayrılmış olacaklardır. parietal. supraoksipital kemik Kafatasının arka kısmının üst kısmını oluşturan, düz ve neredeyse dikeydir. Probactrosaurus ve diğer hadrosauroidler.[3]

Premaksillada olduğu gibi, öncül nın-nin Eolambia diş etlerini taşıdı. Belirgin bir dorsomedial süreç var, sekme benzeri bir yapı da Probactrosaurus ve diğer hadrosauriformlar. Dorsomedial işlemin her iki tarafında da aynı şekilde bulunan birkaç ek sekme benzeri diş eti mevcuttu. Probactrosaurus. Öncül, arkada diş hekimi alt çenenin çoğunu oluşturan. Kısa bir ara var veya diastema öncülün diş hekimi ile eklemlenmesi ile diş hekimi üzerinde gözlenen ilk diş pozisyonu arasında Equijubus, Probactrosaurusve diğer hadrosauroidler. Diş hekiminin ön kısmı karakteristik olarak derinleşir. Protohadros, Ouranosaurus, ve Baktrosaurus. Diş hekiminin dış yüzeyinde biri dişini temsil eden iki çıkıntı vardır. koronoid süreç de olduğu gibi Probactrosaurus ve diğer hadrosauroidler. Sevmek Protohadros ve diğer birkaç hadrosauriforms ProbactrosaurusBu sürecin sadece ön ucu kalınlaştırılır. De olduğu gibi Equijubus, Probactrosaurusve çeşitli diğer Iguanodonts küçük bir foramen yuvarlak Diş hekiminin arkasında bulunan.[3]
Diğer iguanodontlara benzer şekilde, dişlerin dişleri Eolambia sıkıca aralıklı ve birbirine geçen sıralar halinde düzenlenmiştir. Herhangi bir zamanda, 32 maksiller diş yuvasının her biri üç dişi tutarken, 30 diş diş yuvasının her biri dört dişi tutar. Bunlardan her yuvadaki iki diş, tıpkı dişlerinki gibi yedek dişlerdir. Probactrosaurus; diğerleri aktif dişlerdir. Her aktif dişin bir faset giymek. Sevmek Probactrosaurus, Gongpoquansaurus, Protohadrosve diğer hadrosauroidler için, her bir maksiller diş kronunun ağzın orta hattına doğru hafifçe kaymış olan sadece bir çıkıntısı vardır. Bu arada her dişçi diş tacı karakteristik olarak bir birincil çıkıntı ve ağzın orta hattına daha yakın bir aksesuar çıkıntısı taşır, bu da ağız içinde de mevcuttur. Protohadros ve diğer hadrosauroidler. Tüm kronlar ayrıca ön ve arka kenarlarda küçük, diş benzeri çentiklere sahiptir ve bu da Probactrosaurus.[3]
Omurga

Sevmek Equijubus, Probactrosaurus, Gongpoquansaurus, ve diğeri styracosternans, Eolambia vardır boyun omurları hangileri çok opisthocoelous yani ön uçları güçlü dışbükeyken arka uçları güçlü bir şekilde içbükeydir. Alışılmadık bir şekilde, üçüncü servikalin ön ucu, diğer herhangi bir servikal omurda görülmeyen arka uçtan biraz daha yükseğe yerleştirilmiştir. Boynun arkasından gelen omurlar, uzunlamasına arasında daha büyük bir açıya sahiptir. eklem süreçleri postzygapophyses olarak bilinir ve ayrıca daha uzun enine süreçler. Enine süreçlerde, başka eklem süreçleri de vardır - yuvarlak parapofizler ve çubuk benzeri diyapofizler - bulunur. En arkadaki servikallerde, postzigapofizleri ayıran derin bir depresyon da vardır ve sinir dikenleri Dik açılı ön kenar boşlukları vardır, bu da sivri uçlu çıkıntılar yaratma etkisine sahiptir.[3][7]

İlk birkaç sırt omurları servikal omurlara benzer, ancak daha uzun ve daha çatal benzeri sinir dikenlerine sahiptir. Gerisi amfiplatyan, yani her iki uçta da düz. Ayrıca daha uzun ve daha dikdörtgen sinir dikenlerine ek olarak daha az uzun ve daha kaide benzeri postzygapophyses vardır. Projeksiyonlar yerine çöküntüler olan parapofizler, enine süreçlerden uzaklaşarak sinir kemeri, enine süreçler ve omurların önündeki prezygapophyses olarak bilinen eklem süreçleri arasında. Bununla birlikte, son birkaç dorsalde enine işlemlerin temeline geri dönerler.[3] Üçüncü ya da dördüncü dorsalde parapofiz, dikiş sinirsel omurga ile merkez, bu diğer dorsallerden farklıdır ancak ilk birkaç dorsala benzerdir. Edmontosaurus. Ayrıca bunun gibi Edmontosaurusprezigapofizleri ayıran medyan çıkıntı, arka dorsallerde daha belirgin hale gelir.[7]
Arasında Eolambia bugüne kadar bulunan en iyi korunmuş örnekler sakrum yedi omur içerir. Bu bireyin olgunlaşmamış olduğu ve hadrosaurların yaşla birlikte sakral omur sayısını artırdığı göz önüne alındığında, yetişkinlerin daha fazla sakralı olmuş olabilir.[2] Kaudal (kuyruk) omurlar amfiko hem ön hem de arka uçları içbükeydir. Kaide benzeri prezygapophyses ve sekme benzeri postzygapophyses vardır, bunlardan ikincisi bir depresyonla ayrılır. Prezygapophyses, saplara dönüşmek üzere uzar ve postzygapophyses orta ve arka kuyruklarda küçülür. Dahası, sinir dikenleri dikdörtgenden güçlü kavisli hale (önde içbükey ve arkada dışbükey) geçiş yapar, merkez daha uzar ve enine süreçler de kaybolur.[3][7]
Uzuvlar ve uzuv kuşakları

Sevmek Probactrosaurus, skapular bıçak nın-nin Eolambia neredeyse dikdörtgendir, bıçağın kenarları düzdür ve bıçağın ucuyla neredeyse dik bir açıyla birleşir. humerus orta hatta doğru eğilir ve humerustaki deltopektoral tepe belirgin şekilde kalınlaşır. Eklemin dış yüzeyinde bir topuz bulunur. yarıçap humerus üzerinde. İkisi de ulna ve yarıçap nispeten düz kemiklerdir. Ulnanın tepesinde üç "çatal" bulunur, yani olekranon süreci ve iki ek üçgen flanş. Bu arada, yarıçap, üst uçta düzleştirilmiş bir iç yüzey ve alt uçta iç yüzeyde, her ikisi de ulna ile eklemlenmek için ayrı bir platform taşır. El beş hanelidir. Arasında metakarpal kemikler elin ikincisi eğimli, üçüncüsü düz ve dördüncüsü kuvvetli bir şekilde kavislidir. Sevmek Otçul kertenkele, Probactrosaurus, ve diğeri baz alınan iguanodonts, ilk basamaktaki pençe belirgin bir şekilde koni biçimindedir. İkinci ve üçüncü pençeler toynağa benzer, üçüncü pençenin yanlarında pürüzlü flanşlar bulunur - bu da diğer iguanodontlarla paylaşılan bir özelliktir.[3]

Preasetabular (ör. kalça soketi ) süreci ilium işlemin alt kısmı ile geniş bir açı oluşturan yatay bir "önyükleme" taşır. Ek olarak, kemiğin üst kenarı, iliumun üzerinde bulunan bir kenar oluşturmak için dışa doğru çıkıntı yapar.ischium kemiğin alt kenarında eklemlenme. Bu özellikler aynı zamanda Probactrosaurus ve diğer iguanodontlar.[3] Kalça soketinin kendisi, bir iguanodont için iliuma göre oldukça büyüktür.[4] İliumun üst yüzeyi boyunca, adsız bir hadrosauroid ile paylaşılan bir özellik olan ayırt edici bir flanş mevcuttur. Woodbine Oluşumu. İliumun postasetabular (yani kalça soketinin arkasında bulunan) süreci, üst kenarında herhangi bir kırılma olmadan arka tarafında daralır. Üzerinde kasık, ileriye dönük süreç öne doğru genişler, örneğin Probactrosaurus, Gongpoquansaurusve diğer iguanodontlar. Ischium'un şaftı, yetişkinlerde karakteristik olarak düzdür (gençlerin, iguanodontlar arasında daha yaygın olan kavisli bir şaftı vardır) ve şaftın dibinde, ileriye doğru genişleyen sıkıştırılmış bir "önyükleme" bulunur.[3][7]
Humerus gibi uyluk eğilir, ancak dışa doğru. Bununla birlikte, karakteristik olarak uyluğun alt yarısı düzdür. dördüncü trokanter uyluk kemiğinin yarısında yer alan, dikey olarak uzun ve üçgen şeklindedir. Probactrosaurus, Gongpoquansaurusve diğer iguanodontlar, ancak daha bazaldan farklı ornitopodlar. Ulna ve radius gibi, tibia ve fibula düz şaftlara sahipler ve birbirleriyle birleşiyorlar; Fibula'nın altındaki pürüzlü bir çıkıntı, iki ampul benzeri arasına sığar Kondiller hemen arkasında kemal arması tibianın alt ucunda. Tibianın tepesi pürüzlüdür ve başlığın varlığını gösterir. kıkırdak femur ve tibia arasında. Ayağın dört rakamı vardır. Metakarpaller gibi, ikinci metatarsal kavisli, üçüncüsü düz ve dördüncü kuvvetli kavisli. falankslar Her bir basamağın sonuna yakın olan terminal falankslar özellikle kalın olacak şekilde ayağın çok sağlamdır. Eldeki ikinci ve üçüncü pençelerde olduğu gibi, ayağın ikinci, üçüncü ve dördüncü pençeleri tek tip geniş, düz ve toynağa benzer.[3]
Sınıflandırma
İlk hadrosaurid tanımlama

Kirkland'ın ilk açıklamasında Eolambia, onun bir üyesi olduğunu düşündü Hadrosauridae tanımlandığı gibi David B. Weishampel, David B. Norman ve 1993 yılında Dan Grigorescu. Weishampel ve meslektaşları Hadrosauridae'yi tanımlamak için yedi birleştirici özellik kullandılar: maksilla'nın yükselen sürecinin yukarı doğru genişlemesi; kuadratı ayıran paraquadrate foramenin yokluğu ve dörtlü; yeri açısal alt çenenin iç yüzeyinde; dikdörtgen şeklindeki foramenlerin yokluğu; çenenin dar dişleri; her bir diş diş pozisyonunda üç veya daha fazla dişin varlığı; ve skapular bıçağın üst marjının azalması.[15] Bu özelliklerin birinci, beşinci, altıncı ve yedinci Eolambia, geri kalanı eksik malzeme nedeniyle bilinmemektedir. Kirkland ayrıca atandı Eolambia Euhadrosauria'ya, Weishampel ve meslektaşları tarafından ortak atayı içerecek şekilde tanımlanan Hadrosaurinae (şimdi Saurolophinae) ve Lambeosaurinae - hadrosauridlerin iki ana dalı - ve tüm soyundan gelenler.[15] Bu görev, tümü şu ülkelerde bulunan beş özelliğe dayanıyordu: Eolambia: premaksillada diş etlerinin varlığı; kuadratın alt çene ile dar eklemi; dar diş dişleri; her bir diş dişinde tek bir kenarın veya karinanın varlığı; ve humerus üzerindeki açılı deltopektoral tepe.[2]

Hadrosauridae içinde, Kirkland ayrıca Eolambia ya Lambeosaurinae'nin bazal üyesi olmak ya da kardeş grubu Lambeosaurinae. Çeşitli çalışmalarda tanımlandığı gibi, Lambeosaurinae ile paylaşılan beş özelliği belirledi:[15][16][17][18] premaksillada foramen olmaması; burun deliğinin premaksilla tarafından (en azından kısmen) kapatılması; üst çene üzerinde bir rafın geliştirilmesi; kuyruk omurlarının çok uzun sinir dikenleri; humerusun sağlamlığı; ve yetişkinlerde ischiumun büyük "botu". Kirkland, tepesizliği iki ek özelliğin ayırdığını buldu. Eolambia tepenin gelişimi ile ilgili olduğu düşünülen diğer tepeli lambeosaurinlerden: burun boşluğu göz yuvasının üstünde ve parietal kısalması. Eğer Eolambia bir lambeosaurine olarak tanınması için Lambeosaurinae'nin bu iki özelliği dışlamak için yeniden tanımlanması gerekecekti. Alternatif olarak, eğer Eolambia bunun yerine Lambeosaurinae'nin kardeş grubuydu, o zaman hadrosauridlerde hadrosaurinler ve lambeosaurinler arasındaki ayrışmaya yakın bir morfolojiyi temsil edecekti. Bu olasılık, sakrumun alt kısmında bir oluğun varlığı ile desteklenir. EolambiaKirkland tarafından hadrosaurinlerin tanımlayıcı bir özelliği olarak tanınan[17][18] - bir de olsa yakınsak mevcut Ankilosauri ve Ceratopsia.[2]
Lambeosaurine afinitelerini desteklemek için EolambiaKirkland bir Filogenetik analiz; ağaç buna göre yerleştirdiği analizi ile kurtarıldı Eolambia en bazal lambeosaurine olarak aşağıda yeniden üretilmiştir. Bu pozisyonu desteklemek için gösterdiği kanıtlara rağmen Kirkland, görünüşte tutarsız bir özelliğe de dikkat çekti. Çocuk iliumunda görüldüğü gibi, başlangıçta antitrokanter - ilium üzerinde, büyük trokanter femur - bir bazal hadrosaurid için beklenenden daha zayıf bir şekilde gelişmiştir. Michael Brett-Surman, yetişkinlerin daha gelişmiş bir antitrokantere sahip olabileceğini belirtti. Öte yandan, türetilmiş lambeosaurine Hipakrosaurus yumurtadan çıkma aşamasında bile iyi gelişmiş bir antitrokantere sahiptir. Kirkland, bazal hadrosauridlerin antitrokanterlerini, aşağıdaki gibi türetilmiş hadrosauridlerden daha sonra yaşamda geliştirdikleri olasılığına dikkat çekti. Hipakrosaurus.[19] Böylece o yetişkinin Eolambia örnekler bu sorunu çözecektir.[2]


| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Hadrosaurid olmayan olarak yeniden tanımlama

2001'de Jason Head, filogenetik konumunu yeniden değerlendirdi. Eolambia OMNH'de depolanan örneklere göre. Kirkland'ın sözde lambeosaurine'in ayırt edici özelliklerini tanımladığını fark etti. Eolambia hatalı veya şüpheli olarak. Head'in gösterdiği gibi, aslında premaksillada bir foramen vardır; premaksilla, burun deliğini yalnızca alt kenarda sınırlar; rafın yanı sıra üst çenenin önünde parmak benzeri iki işlem vardır; kuyruk omurlarının yüksek nöral dikenleri ayrıca lambeosaurine olmayanlarda da görülür. Ouranosaurus ve Baktrosaurus; ve ischiumun ucunun genişlemesi ayrıca Camptosaurus, Otçul kertenkele, Ouranosaurus, Gilmoreosaurus, ve Baktrosaurus, genişletmenin plesiomorfik iguanodontlar için ve ikincil olarak hadrosaurinlerde kayboldu.[18] M.L. 1999'da Casanovas ve meslektaşları, lambeosaurinleri "kulüp benzeri" bir durumun aksine, iskiyumun ucunda "genişletilmiş bir ayak" olarak nitelendirdiler.[20] Head şunu savundu Corythosaurus ve Lambeosaurus[21] ischial ipuçlarına sahip olmak, Mantellisaurus veya Ouranosaurus.[4][22]

Baş ayrıca dişlerin Eolambia Kirkland'ın sandığından daha geniş ve asimetrikti; ayrıca, hadrosauridlerden farklı olarak, alt yetişkinlerin diş pozisyonu başına yalnızca iki dişe sahip olduğunu kaydetti,[4] her ne kadar yetişkin örnekler aslında Kirkland tarafından gösterildiği gibi üç veya daha fazla diş pozisyonuna sahip olsa da.[3] Kafa daha da gösterdi ki Eolambia türetilmiş hadrosauridlerde sekiz veya daha fazlasının aksine yedi sakral omurlara sahipti ve antitrokanterinin aslında daha zayıf geliştiğini Ouranosaurus ve Altirhinus.[23] OMNH örneklerinin incelenmesi ayrıca iliumun postasetabular bölgesinin ayrı bir süreç oluşturmadığını ve femurun altındaki eklemlerin çok belirgin olmadığını gösterdi, her ikisi de ayrı Eolambia hadrosauridler gibi daha fazla türetilmiş iguanodontlardan. Böylece Head şu sonuca vardı: Eolambia bir baz alınan Hadrosauridae dışında, Hadrosauroidea üyesi, bir filogenetik analiz tarafından desteklenen bir sonuç, bunun hadrosaurid olmayan bir hadrosauroid olduğunu bulmuştur. Probactrosaurus.[4]
Sonraki araştırma

2004 yılında Norman dahil Eolambia ikinci baskısında iguanodontlara odaklanan bir bölümün parçası olarak, iguanodontların kapsamlı bir filogenetik analizinde ilk kez Dinosauria. Bunu buldu Eolambia kardeş grup muydu Altirhinusher ikisi de daha bazal olmak üzere Protohadros, Probactrosaurusveya Hadrosauridae.[24] Aynı cilt içinde Horner, Weishampel ve Catherine Forster Hadrosauridlerin ayrı bir filogenetik analizi yaptı, benzer şekilde Eolambia. Onlar buldular Eolambia yine hadrosaurid olmayan bir hadrosauroid olarak, kardeş grup olarak da olsa Protohadros. Head, Hadrosauridae'yi ortak özelliklere dayanarak özellikle yeniden tanımlamış olsa da, Protohadros,[4][25] Horner ve meslektaşları, takson temelli bir tanımı uyarladılar. Protohadros ve böylece Eolambia. Ayrıca farklılaştıran ek özellikler belirlediler Eolambia hadrosauridlerden: diş hekiminin dişlerinde kaba dişler vardır ve koronoid süreç zayıf bir şekilde genişler.[26]

İçin kurtarılmış filogenetik pozisyonlardaki varyans Eolambia sonraki yıllarda da devam etti. 2009 açıklamasında Levnesovia, Hans-Dieter Sues ve Alexander Averianov bunu buldu Protohadros göre ara bir pozisyon işgal etti Altirhinus ve Probactrosauruskardeş grubu olmak Fukuisaurus.[27] Bu arada, Horner ve meslektaşlarının sonuçlarına göre, Albert Prieto-Márquez ve Mark Norell 2010'da bulundu ki Eolambia onun yerine kardeş grubuydu Protohadros. Ayrıca, Eolambia hadrosauridlerle yakınsak olanlar: tek diş karinanın varlığı; önsözün neredeyse kareye yakın dış köşesi; ve (kuadratojugal ile eklemlenen) kuadratojugal çentiğin orta noktası, kuadratın yüzeyinin yaklaşık yarısına kadar yerleştirilmiştir (üstten kemiğin yüksekliğinin% 60'ından daha aşağıda yer alır).[28] 2012'de Holtz sınıflandırıldı Eolambia ilkel bir üyesi olarak Styracosterna ile birlikte Altirhinus.[14]
McDonald, 2012 yılında, iguanodontların filogenetik analizini yeni Eolambia örnekler. Önceki yazarların aksine, bunun yerine şunu buldu: Eolambia kardeş grubuydu Probactrosaurus. Bu yakın ilişki, iki ortak özelliğe dayanıyordu: dörtgen, geriye doğru eğimli olan üst uç dışında düz; ve skapular bıçağın üst ve alt kenarları neredeyse paraleldir.[3] Wenjie Zheng ve meslektaşları ekledi Jintasaurus 2014 yılında gruba,[29] José Gasca ve meslektaşları 2015 yılında orijinal grubu kurtardı.[30] Bununla birlikte, McDonald, FMNH örneklerinin 2017 açıklamasına hazırlık olarak filogenetik veri setini kapsamlı bir şekilde revize etti. Horner ve meslektaşlarının yanı sıra Prieto-Márquez ve Norell gibi, McDonald ve meslektaşları da şunu buldu: Eolambia kardeş grubuydu Protohadrosher ikisi de bazal olarak yerleştirilerek hadrosauromorphs (sonuçlar aşağıda gösterilmiştir).[7] Francisco Verdú ve meslektaşları, McDonald veri setinin 2012 versiyonunu kullanarak benzer bir sonucu bağımsız olarak kurtardılar.[31][32] ile Eolambia ile yakından ilgili olmak Protohadros ve Jeyawati; Bununla birlikte, McDonald ve meslektaşları ikincisinin 2017'de daha türetildiğini buldu.[7]

| Hadrosauriformes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Terry Gates ve meslektaşları 2018'de yeni iguanodot cinsini tanımladı Choyrodon, ile aynı bölgeden Altirhinusgeçmişte akraba olarak kabul edildi Eolambia. Norman'ın (2015) matrisine dayanan filogenetik analizleri şunu buldu: Choyrodon kardeş takson olmak Eolambia. Eolambia'sık görülen akraba Protohadros bu matriste mevcut değildi. Bu kardeş ilişkisini üç özellik destekledi: yamuk şekilli oksiput ve dişlerin ortak iki özelliği. Bu cinslerin diğer hadrosauriformlara göre yerleştirilmesi, antorbital fenestranın mevcut veya yok olarak kodlanmasına bağlıydı. Choyrodon; juvenil tip örnekte bir tane vardır, ancak yetişkinlerin kapalı bir örneğe sahip olma olasılığı göz ardı edilemez. Aşağıdaki iki ağaç (metinde Gates ve meslektaşları tarafından tanımlanan sınıflarla etiketlenmiştir), sırasıyla mevcut ve yok olarak kodlanmış sonuçları gösterir, ilki büyük bir politomi:[33][başarısız doğrulama ]
Topoloji A: Antorbital fenestra şu şekilde kodlanmıştır: Choyrodon[33]
| Topoloji B: Dıştan kapalı olarak kodlanan antorbital fenestra Choyrodon[33]
|
Paleobiyoloji
Muhtemelen ait olan yumurta kabukları Eolambia izole dişlerin yanında çeşitli yerlerde keşfedilmiştir. Retiküler (ağ benzeri) yüzey desenlerine sahiptirler ve 2 milimetre (0,079 inç) kalınlığa sahiptirler.[2]
Büyüme

Çeşitli genç ve yetişkin örneklerden tanınan değişiklikler Eolambia Büyüdükçe geçirilen iyi belgelenmiştir. Çenedeki diş sayısı, bir gençte 23'ten bir yetişkinde 33'e yükseldi. Benzer bir artış diş hekimi dişler, gençlerde 18-22, yetişkinlerde 25-30. Yavruların diş dişlerinin kronlarında ikincil çıkıntılar yokken, yetişkinlerde soluk ikincil çıkıntılar mevcuttur.[3] Kirkland tarafından belgelendiği gibi, iskiyumun "botunun" boyutu da yaşla birlikte arttı.[2] FMNH tarafından keşfedilen örneklerle belgelendiği gibi, iskiyumun şaftının düzlüğünde de değişiklikler meydana gelmiş olabilir.[7]

A'dan sonuçlar histolojik FMNH PR 3847 numunesinden alınan bir kaburga üzerinde gerçekleştirilen analiz, 2017 yılında McDonald ve arkadaşları tarafından rapor edilmiştir. kemik yeniden şekillenmesi kaburganın iç matrisinde meydana geldi. medüller boşluk, barındıran ilik, birçok boşluğun varlığı ile işaretlenmiştir veya lacunae. Kemiğin dış yüzeyinde, osteonlar kemiğin iç kısmını neredeyse tamamen değiştirmiş; kemiğin katmanlı büyümesi kemiğin iç kısmında daha belirgindir, burada beş tutuklanan büyüme hatları (LAG'ler) görülebilir. Medüller bölgede muhtemelen altıncı bir LAG mevcuttur, ancak kapsamı nispeten sınırlıdır, bu da bu tanımlamayı sorgulanabilir hale getirir.[7]
LAG'ler kemiğin yeniden şekillenmesiyle ortadan kaldırılabildiğinden, bu, gerçekte gelişen LAG'lerin gerçek sayısının eksik bir tahminini temsil eder; McDonald ve meslektaşları, görünür LAG'ler arasındaki aralığı inceleyerek üç veya dört eksik LAG'yi tahmin ettiler. Bu, hayvanın öldüğünde sekiz ila dokuz yaşında olduğunu gösterir. LAG'ler arasındaki boşluk, dış yüzeye doğru önemli ölçüde azalır, bu da büyümenin yavaşladığını gösterir, ancak dış temel sistemin (bir dizi sıkı aralıklı LAG) olmaması, büyümenin henüz durmadığını gösterir. Bu, servikal ve dorsal omurlardaki merkez ve nöral dikenler arasındaki kaynaşmamış sütürler ve sakral sentra ile karşılık gelen kaburgaları arasındaki kaynaşmamış sütürlerle tutarlıdır.[34] Genel olarak, büyüme oranı Eolambia ile karşılaştırılabilir görünüyor Hipakrosaurus ve Maiasaura yedi veya sekiz yıl sonra olgunluğa ulaşan.[7][35][36]
Paleoekoloji
Çevre ve tafonomi
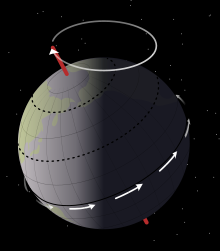
Sedimentolojik Mussentuchit Üyesinden alınan veriler şunu göstermektedir: Eolambia kötü süzülmüş bir yerde yaşadı taşkın yatağı, büyük, çok yıllık göller sistemi etrafında. Göller, yüksek ve düşük su seviyeleri arasında dalgalandı. Milankovitch döngüleri kabaca 21.000 yıllık, Dünya'nın eksenel devinim. Bu döngüler, günümüz kaya birikintilerinde değişen çamurlu kumtaşı ve çamurtaşı katmanları tarafından yansıtılır. During dry periods, the exposed lakebed formed a broad beach devoid of plants. cracked mud of the lakebed produced çamur tıkaçları, while organic debris and scavenged carcasses were scattered around the water's edge as the lake progressively retreated. Meanwhile, during periods in which the sea level rose, small rivers invaded the floodplain, which yeniden işlenmiş and displaced the debris that had been deposited during the dry season.[6]

Most of the fossils discovered in the Mussentuchit are scattered and disarticulated, with intact skeletons being relatively rare. Although the lakes did not actively destroy bones, it was not particularly conducive to good preservation either due to the long duration of the burial process. Before burial, scavengers could have disrupted the skeletal material, which is evident in the severed Eolambia tibiae bearing tooth marks discovered in the Cifelli #2 quarry. The regression of the lake could have occasionally created bataklıklar, but they would not have been permanent enough to entrap and preserve larger vertebrates. Action by water currents seems to have played a role in the deposition of Eolambia bones in the Cifelli #2 quarry, where the bones are largely deposited at 28°, 69°, 93°, 131° and 161° azimut (i.e. relative to north). These directions would have been parallel or sub-parallel to the edges of the lake, indicating orientation by lake currents, except for the 69° and 93° azimuths, which probably represent river currents.[6]
The abundant plant debris, combined with the absence of lavabo (playa) deposits, indicates that the local climate was relatively humid during the deposition of the Mussentuchit. This is in marked contrast to earlier deposits in the area, which are kireçli (chalky), indicating periods of aridity lasting up to 10,000 years. Paleoiklim reconstructions support these interpretations, with the encroachment of the Mowry Sea from the north (a process which would eventually form the Batı İç Denizyolu ) resulting in arid (Mid-latitude Continental Interior) climates being replaced by more humid (Mid-latitudeEast Coast) climates.[37] With a paleolatitude of 46° N, the Mussentuchit would have had a warm mean surface temperature of 40–45 °C (104–113 °F), about 6–8 °C (43–46 °F) warmer than the present day, according to genel dolaşım modelleri.[6][38]
Ekosistem

Krokodilomorflar are abundant in the Mussentuchit deposits, being represented largely by teeth. The largest known teeth have been referred to the Pholidosauridae;[a] they are broad with rounded bases, and some of them bear longitudinal ridges.[7] These large crocodilians would have been capable of preying upon at least juvenile Eolambia su kenarında. The abundance of juvenile Eolambia specimens in the lakeside fossil material supports this hypothesis; daha büyük Eolambia would have been invulnerable to crocodilians.[6] Smaller crocodilian teeth are also present, with two distinct morphotypes: an ovoid form bearing ridges, which has been referred to two different species of Bernissartia;[6][a] and a compressed, triangular form bearing heavy ornamentation, which has been attributed to the Atoposauridae.[12][40][a] However, given the uniform small size of these teeth, and the co-occurrence of these morphotypes in some crocodilians,[39] they may belong to the same species.[7]

Daha büyük Eolambia would have been vulnerable[6] to the large neovenatorid Theropod Siats,[41] which was discovered by expeditions from the FMNH.[7] A variety of smaller Maniraptoran theropods are also present in the Mussentuchit, which would have functioned as scavengers; their remains include teeth attributed to the Dromaeosauridae, Paronychodon, ve Richardoestesia.[40] As for herbivorous dinosaurs, the nodosaurid Animantarx was discovered in Carol's Site, near the type specimen of Eolambia.[42] Teeth belonging to more basal ornithopods have also been discovered, with similar teeth having been referred to the Pachycephalosauria or to toothed birds by Cifelli in 1999.[6][40] More complete remains referred to two different ornithopods – an orodrom and a basal iguanodont – remain unpublished.[43] Bir dev Oviraptorosaur has also been discovered,[44] alongside teeth from ceratopsians and Sauropodlar.[7][40]
Fish, which may have been preyed upon by smaller crocodilians, are represented by teeth and scales. İçerirler myliobatiform ray referred to the genus Baibisha;[40] an indeterminate member of the Neopterygii; a semionotid referred to the genus Lepidotlar; a gar ile ilgili Atraktosteus;[7] a pycnodontid referred to the genus Stephanodus; ve belirsiz bowfin. Belirsiz Kaudatanlar (salamanders) and mammals are also present. As for plants, pollen samples indicate a tiered forest surrounding the lakes of the Mussentuchit, containing a gölgelik nın-nin ağaç eğrelti otları, Araucarias, ve iğne yapraklılar, accompanying an undergrowth of eğrelti otları, küçük jimnospermler, ve çiçekli bitkiler. Algae would also have been present in the waters of the lakes.[9][45] These plants would have provided an abundant source of food for Eolambia.[6]
Paleobiyocoğrafya

Kirkland, Cifelli, and colleagues noted that the fauna of the Mussentuchit – iguanodonts, pachycephalosaurs, and ceratopsians – bears strong similarities to contemporary Asian faunas. They proposed that Eolambia was part of an influx of Asian dinosaurs into North America during the Cenomanian, which supplanted the earlier low-diversity native fauna.[2][12] This hypothesis is supported by the close relationship between Eolambia ya da Probactrosaurus veya Fukuisaurus, which have respectively been recovered by the phylogenies of Head and Sues & Averianov.[3][4][27]
However, the results of Horner and colleagues, Prieto-Márquez, and McDonald and colleagues, which consider the North American Protohadros to be the closest relative of Eolambia, contradict this hypothesis.[7][26][28][46] This alternative interpretation suggests that the common ancestor of Eolambia, Protohadros, and more derived hadrosauroids was not exclusively Asian, but instead distributed across Asia and North America.[3] McDonald and colleagues further suggested that Eolambia ve Protohadros represent a localized group of hadrosauroids endemik to North America, much like other groups of styracosternans throughout the Cretaceous. Such contemporary groups included a central Asian group of Shuangmiaosaurus, Zhanghenglong, ve Plesiohadros.[7]
Referanslar
- ^ Galton, Peter; Jensen, James A. (1979). "Galton, P. M. ve Jensen, J. A. (1979). Kuzey Amerika'nın Alt Kretase'sinden ornitopod dinozor kalıntıları" (PDF). Brigham Young Üniversitesi Jeoloji Çalışmaları. 25 (3): 1–10. ISSN 0068-1016.
- ^ a b c d e f g h ben j k l m Kirkland, J.I. (1998). "A new hadrosaurid from the upper Cedar Mountain Formation (Albian-Cenomanian: Cretaceous) of eastern Utah – the oldest known hadrosaurid (lambeosaurine?)". Lucas, S.G .; Kirkland, J.I .; Estep, J.W. (eds.). Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 14. s. 283–295.
- ^ a b c d e f g h ben j k l m n Ö p q r s t McDonald, A.T .; Bird, J .; Kirkland, J.I .; Dodson, P. (2012). "Osteology of the Basal Hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah". PLOS ONE. 7 (10): e45712. Bibcode:2012PLoSO...745712M. doi:10.1371/journal.pone.0045712. PMC 3471925. PMID 23077495.
- ^ a b c d e f g Baş, J.J. (2001). "A reanalysis of the phylogenetic position of Eolambia caroljonesa (Dinosauria, Iguanodontia)". Omurgalı Paleontoloji Dergisi. 21 (2): 392–396. doi:10.1671/0272-4634(2001)021[0392:AROTPP]2.0.CO;2.
- ^ Kirkland, J.I .; Burge, D. (1994). A large primitive hadrosaur from the Lower Cretaceous of Utah. Fifty-Fourth Annual Meeting, Society of Vertebrate Paleontology. Omurgalı Paleontoloji Dergisi. 14 (3). s. 32A. JSTOR i406457.
- ^ a b c d e f g h ben j k l m Garrison, J.R. Jr.; Brinkman, D.; Nichols, D.J.; Layer, P.; Burge, D .; Thayn, D. (2007). "A multidisciplinary study of the Lower Cretaceous Cedar Mountain Formation, Mussentuchit Wash, Utah: a determination of the paleoenvironment and paleoecology of the Eolambia caroljonesa dinosaur quarry". Kretase Araştırmaları. 28 (3): 461–494. doi:10.1016/j.cretres.2006.07.007.
- ^ a b c d e f g h ben j k l m n Ö p q r s McDonald, A.T .; Gates, T.A .; Zanno, L.E .; Makovicky, P.J. (2017). "Yeni bir örneğinin anatomisi, tafonomisi ve filogenetik çıkarımları Eolambia caroljonesa (Dinosauria: Ornithopoda) Cedar Mountain Formasyonundan, Utah, ABD ". PLOS ONE. 12 (5): e0176896. Bibcode:2017PLoSO..1276896M. doi:10.1371 / journal.pone.0176896. PMC 5425030. PMID 28489871.
- ^ Craig, L.C. (1981). "Lower Cretaceous rocks, southwestern Colorado and southeastern Utah". Rocky Mountain Association of Geologists 1981 Field Conference: 195–200.
- ^ a b Tschudy, R.H.; Tschudy, B.D.; Craig, L.C. (1984). Palynological evaluation of Cedar Mountain and Burro Canyon formations, Colorado Plateau (Bildiri). US Geological Survey, Professional Paper. 1821. Birleşik Devletler Jeoloji Araştırmaları. s. 1–24.
- ^ Nichols, D.J.; Sweet, A.R. (1993). "Biostratigraphy of Upper Cretaceous nonmarine palynofloras in a north-south transect of the Western Interior Basin". Caldwell, W.G.E .; Kauffman, E.G. (eds.). Evolution of the Western Interior Basin. Geological Association of Canada, Special Paper. 39. Kanada Jeoloji Derneği. pp. 539–584.
- ^ Young, R.G. (1960). "Dakota Group of Colorado Plateau". American Association of Petroleum Geologists Bulletin. 44 (2): 156–194. doi:10.1306/0bda5fd2-16bd-11d7-8645000102c1865d.
- ^ a b c Cifelli, R.L .; Kirkland, J.I .; Weil, A .; Deino, A.L.; Kowallis, B.J. (1997). "High-precision 40Ar/39Ar geochronology and the advent of North America's Late Cretaceous terrestrial fauna". Ulusal Bilimler Akademisi Bildiriler Kitabı. 94 (21): 11163–11167. Bibcode:1997PNAS...9411163C. doi:10.1073/pnas.94.21.11163. PMC 23402. PMID 9326579.
- ^ Paul, G.S. (2010). "Ornithischians". Princeton Dinozorlar Saha Rehberi. Princeton: Princeton Üniversitesi Yayınları. s. 327. ISBN 9780691167664.
- ^ a b Holtz, T.R. (2012). "Genus List for Holtz (2007) Dinozorlar" (PDF). Supplementary Information to Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages by Thomas R. Holtz, Jr., illustrations by Luis Rey. Department of Geology, University of Maryland. Alındı 30 Ağustos 2017.
- ^ a b c Weishampel, D.B .; Norman, D.B .; Grigorescu, D. (1993). "Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur". Paleontoloji. 36 (2): 361–385.
- ^ Sereno, P.C. (1986). "Phylogeny of the bird-hipped dinosaurs (Order Ornithischia)". National Geographic Araştırma. 2: 234–256.
- ^ a b Brett-Surman, M.K. (1989). Hadrosauridae'nin (Reptilia: Ornithischia) bir revizyonu ve Kampaniyen ve Maastrihtiyen sırasında evrimleri (PDF) (Doktora). Washington D.C .: George Washington Üniversitesi. pp. 1–373.
- ^ a b c Weishampel, D.B .; Horner, J.R. (1990). "Hadrosauridae". Weishampel, D.B .; Dodson, P .; Osmolska, H. (editörler). Dinosauria (1. baskı). Berkeley: California Üniversitesi Yayınları. s. 534–561.
- ^ Horner, J.R .; Currie, P.J. (1994). "Embryonic and neonatal morphology and ontogeny of a new species of Hipakrosaurus (Ornithischia, Lambeosauridae) from Montana and Alberta". Carpenter, K .; Hirsch, K.F.; Horner, J.R. (eds.). Dinosaur Eggs and Babies. Cambridge: Cambridge University Press. pp. 312–336. ISBN 9780521567237.
- ^ Casanovas, M.L.; Pereda Suberbiola, X.; Santafe, J.V.; Weishampel, D.B. (1999). "First lambeosaurine hadrosaurid from Europe: palaeobiogeographical implications". Jeoloji Dergisi. 136 (2): 205–211. Bibcode:1999GeoM..136..205C. doi:10.1017/s0016756899002319.
- ^ Lull, R.S .; Wright, N.E. (1942). "Kuzey Amerika'nın Hadrosaurian Dinozorları". Amerika Jeoloji Derneği Özel Belgeleri. 40: 1–242. doi:10.1130 / SPE40-p1.
- ^ Norman, D.B. (1986). "On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda)". Bulletin de l'Institut Royal des Sciences Naturelles de Belgique. 56: 281–372.
- ^ Norman, D.B. (1998). "Asya ornitopodlarında (Dinosauria: Ornithischia). 3. Yeni bir iguanodontid dinozor türü". Linnean Society'nin Zooloji Dergisi. 122 (1): 291–348. doi:10.1111 / j.1096-3642.1998.tb02533.x.
- ^ Norman, D.B. (2004). "Basal Iguanodontia". Weishampel, D.B .; Dodson, P .; Osmólska, H. (editörler). Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. sayfa 413–437.
- ^ Baş, J.J. (1998). "A new species of basal hadrosaurid (Dinosauria, Ornithischia) from the Cenomanian of Texas". Omurgalı Paleontoloji Dergisi. 18 (4): 718–738. doi:10.1080/02724634.1998.10011101.
- ^ a b Horner, J.R .; Weishampel, D.B .; Forster, C.A. (2004). "Basal Iguanodontia". Weishampel, D.B .; Dodson, P .; Osmólska, H. (editörler). Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. sayfa 438–463.
- ^ a b Sues, H.-D .; Averianov, A. (2009). "Özbekistan'ın Geç Kretase'sinden yeni bir bazal hadrosauroid dinozor ve ördek gagalı dinozorların erken radyasyonu". Royal Society B Tutanakları. 276 (1667): 2549–2555. doi:10.1098 / rspb.2009.0229. PMC 2686654. PMID 19386651.
- ^ a b Prieto-Márquez, A.; Norell, MA (2010). "Anatomi ve İlişkiler Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia". Amerikan Müzesi Novitates. 3694: 1–49. doi:10.1206/3694.2. S2CID 56372891.
- ^ Zheng, W .; Jin, X .; Shibata, M .; Azuma, Y. (2014). "Eski bir çocuk Bolong yixianensis (Ornithopoda: Iguanodontia) from the Lower Cretaceous of Ningcheng County, Nei Mongol, China". Tarihsel Biyoloji. 26 (2): 236–251. doi:10.1080/08912963.2013.809347. S2CID 129081459.
- ^ Gasca, J.M.; Moreno-Azanza, M .; Ruiz-Omeñaca, J.I.; Canudo, J.I. (2015). "New material and phylogenetic position of the basal iguanodont dinosaur Delapparentia turolensis from the Barremian (Early Cretaceous) of Spain". İber Jeolojisi Dergisi. 41 (1): 57–70. doi:10.5209/rev_jige.2015.v41.n1.48655.
- ^ Verdú, F.J .; Royo-Torres, R .; Cobos, A .; Alcalá, L. (2015). "Perinates of a new species of Otçul kertenkele (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain)". Kretase Araştırmaları. 56: 250–264. doi:10.1016/j.cretres.2015.05.010.
- ^ Verdú, F.J .; Royo-Torres, R .; Cobos, A .; Alcalá, L. (2017). "New systematic and phylogenetic data about the early Barremian İguanodon galvensis (Ornithopoda: Iguanodontoidea) from Spain". Tarihsel Biyoloji. 30 (4): 437–474. doi:10.1080/08912963.2017.1287179. S2CID 89715643.
- ^ a b c Gates, T.A .; Tsogtbaatar, K .; Zano, L.E.; Chinzorig, T .; Watabe, M. (2018). "A new iguanodontian (Dinosauria: Ornithopoda) from the Early Cretaceous of Mongolia". PeerJ. 6: e5300. doi:10.7717 / peerj.5300. PMC 6078070. PMID 30083450.
- ^ Brochu, C.A. (1994). "Timsah ontogenisi sırasında nörosentral sütürlerin kapatılması: Fosil archosaurlarda olgunluk değerlendirmesi için çıkarımlar". Omurgalı Paleontoloji Dergisi. 16 (1): 49–62. doi:10.1080/02724634.1996.10011283.
- ^ Horner, J.R .; de Ricqlès, A.; Padian, K. (1999). "Variation in dinosaur skeletochronology indicators: implications for age assessment and physiology". Paleobiyoloji. 25 (3): 295–304. doi:10.1017/S0094837300021308.
- ^ Woodward, H.N.; Freedman Fowler, E.A.; Farlow, J.O.; Horner, J.R. (2015). "Maiasaura, a model organism for extinct vertebrate population biology: a large sample statistical assessment of growth dynamics and survivorship". Paleobiyoloji. 41 (4): 503–527. doi:10.1017/pab.2015.19. S2CID 85902880.
- ^ Elder, W.P.; Kirkland, J.I. "Cretaceous Paleogeography of the Southern Western Interior Region". In Caputo, M.V.; Peterson, J.A.; Franczyk, K.J. (eds.). Mesozoic Systems of the Rocky Mountain Region, USA. Denver: Rocky Mountain Section SEPM. sayfa 415–440.
- ^ Glancy, T.J. Jr .; Arthur, M.A.; Barron, E.J.; Kaufman, E.G. (1993). "A paleoclimate model for the North American Cretaceous (Cenomanian-Turonian) epicontinental sea". Caldwell, W.G.E .; Kauffman, E.G. (eds.). Evolution of the Western Interior Basin. Geological Association of Canada, Special Papers. 39. Kanada Jeoloji Derneği. pp. 219–241.
- ^ a b Irmis, R.B .; Hutchison, J.H.; Sertich, J.J.W.; Titus, A.L. (2013). "Crocodyliforms from the Late Cretaceous of Grand Staircase-Escalante National Monument and Vicinity, Southern Utah, USA". In Titus, A.L.; Loewen, M.A. (eds.). At the Top of the Grand Staircase: the Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. pp. 424–444. ISBN 9780253008961.
- ^ a b c d e Cifelli, R.L .; Nydam, R.L .; Gardner, J.D.; Weil, A .; Eaton, J.G.; Kirkland, J.I .; Madsen, S.K. (1999). "Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County, Utah: the Mussentuchit local fauna". In Gillette, D. (ed.). Utah'da Omurgalı Paleontoloji. Utah Geological Survey Miscellaneous Publications. 99–1. s. 219–242.
- ^ Zanno, L.E .; Makovicky, P.J. (2013). "Neovenatorid theropodlar, Kuzey Amerika'nın Geç Kretase'sinde en uç yırtıcılardır". Doğa İletişimi. 4: 2827. Bibcode:2013NatCo ... 4.2827Z. doi:10.1038 / ncomms3827. PMID 24264527.
- ^ Carpenter, K .; Kirkland, J.I .; Burge, D.L .; Bird, J. (1999). "Ankylosaurs (Dinosauria: Ornithischia) of the Cedar Mountain Formation, Utah, and their stratigraphic distribution". In Gillette, D. (ed.). Utah'da Omurgalı Paleontoloji. Utah Geological Survey Miscellaneous Publications. 99–1. sayfa 243–251.
- ^ Zanno, L.E .; Makovicky, P.J. (2016). A New Species of Early Diverging Ornithopod Increases the Paleobiodiversity of Herbivorous Dinosaurs in Late Cretaceous Ecosystems in North America. Society of Vertebrate Paleontology 76th Annual Meeting. Salt Lake City: Society of Vertebrate Paleontology. s. 256.
- ^ Makovicky, P.J.; Zanno, L.E. (2014). Kirkland, J.I .; Hunt-Foster, R.; Liggett, G.A.; Trujillo, K. (eds.). New Discoveries Add to the Diversity of the Mussentuchit Member (Cedar Mountain Formation) Dinosaur Fauna. Mid-Mesozoic: The Age of Dinosaurs in Transition (PDF). Fruita: Utah Geological Survey. s. 70–71.
- ^ Ravn, R.L. (1995). "Miospores from the Muddy Sandstone (Upper Albian), Wind River Basin, Wyoming, USA". Palaeontographica Abteilung B. 234 (3): 41–91.
- ^ Prieto-Márquez, A. (2010). "Global historical biogeography of hadrosaurid dinosaurs". Linnean Society'nin Zooloji Dergisi. 159 (2): 503–525. doi:10.1111/j.1096-3642.2010.00642.x.
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||






