Dromaeosauridae - Dromaeosauridae
| Dromaeosauridler | |
|---|---|
 | |
| Dromaeosaurid fosil iskeletlerinin bir koleksiyonu. Sol üstten saat yönünde: Deinonychus antirrhopus (ağır yapılı bir dromaeosaurine), Buitreraptor gonzalezorum (uzun burunlu unenlagiine), Velociraptor mongoliensis (küçük bir velosiraptorin), Microraptor gui (kanatlı bir mikroraptor), Halszkaraptor escuilliei (yarı sulu bir halszkaraptorin), Zhenyuanlong suni (uzun kanatlı bir dromaeosaurid) | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Eumaniraptora |
| Aile: | †Dromaeosauridae Matthew & Kahverengi, 1922 |
| Türler | |
| †Dromaeosaurus albertensis Matthew ve Brown, 1922 | |
| Alt gruplar[1] | |
| Eş anlamlı | |
| |
Dromaeosauridae /ˌdrɒmbenəˈsɔːrɪdben/ bir aile tüylü Theropod dinozorlar. Genellikle küçük ila orta boy tüylüydüler etoburlar gelişti Kretase Periyot. Dromaeosauridae adı, 'koşan kertenkeleler' anlamına gelir. Yunan δρομεῦς (dromeus) 'koşucu' anlamına gelir ve σαῦρος (Sauros) 'kertenkele' anlamına gelir. Gayri resmi kullanımda genellikle denir Raptors[2] (sonra Velociraptor ), film tarafından popüler hale getirilen bir terim Jurassic Park; birkaç tür, doğrudan adlarına "yırtıcı hayvan" terimini dahil eder ve kuş benzeri görünümleri ve speküle edilmiş kuş benzeri davranışlarını vurgulamaya başlamıştır.
Dromaeosaurid fosilleri dünyanın dört bir yanında bulundu Kuzey Amerika, Avrupa, Afrika, Asya, Güney Amerika ve Antarktika,[3] fosilleşmiş dişler, yaşadıkları olasılığına inanır Avustralya yanı sıra.[4] İlk olarak orta Jura Döneminde (geç Batoiyen sahne, yaklaşık 167 milyon yıl önce) ve Kretase'nin sonuna kadar (Maastrihtiyen sahne, 66 ma), Kretase-Paleojen nesli tükenme olayı. Dromaeosauridlerin varlığı Orta Jura izole fosil dişlerin bulunmasıyla öne sürülmüştür, ancak bu döneme ait dromaeosaurid vücut fosillerine rastlanmamıştır.[5]
Açıklama

Ayırt edici dromaeosaurid vücut planı dinozorların aktif, hızlı ve kuşlarla yakından ilişkili olabileceği teorilerini yeniden canlandırmaya yardımcı oldu. Robert Bakker için resmi John Ostrom 1969 monografı,[6] dromaeosaurid gösteriliyor Deinonychus hızlı bir şekilde, tarihteki en etkili paleontolojik rekonstrüksiyonlardan biridir.[7] Dromaeosaurid vücut planı, nispeten büyük bir kafatası, tırtıklı dişler, dar burun içerir (bir istisna, Utahraptor ) ve bir dereceye kadar dürbün görüşünü gösteren öne bakan gözler.[8] Dromaeosauridler, diğer birçok theropod gibi, orta derecede uzun S-kıvrımlı bir boyuna sahipti ve gövdeleri nispeten kısa ve derindi. Diğerleri gibi Maniraptoranlar bazı türlerde vücuda katlanabilen uzun kolları ve büyük pençelerle biten üç uzun parmağı (orta parmak en uzun ve ilk parmak en kısadır) ile nispeten büyük elleri vardı.[9] Dromaeosaurid kalça yapısı, kuyruk tabanının altında çıkıntı yapan karakteristik olarak büyük bir kasık botuna sahipti. Dromaeosaurid ayakları, ikinci ayak parmağında büyük, kıvrık bir pençe taşıyordu. Kuyrukları ince, uzun, alçak, omur 14. kaudal omurdan sonra enine süreç ve sinir dikenleri yok.[9]
Dromaeosauridlerin en azından bazılarının ve muhtemelen hepsinin büyük, kanatlı, kanat ve kuyruk tüyleri dahil olmak üzere tüylerle kaplı olduğu artık biliniyor. İlk olarak 1980'lerin ortalarından sonlarına kadar varsayımı yapılan ve 1999'daki fosil keşifleriyle doğrulanan bu gelişme, dromaeosauridlerin sanat ve filmde tarihsel olarak tasvir edilme biçiminde önemli bir değişikliği temsil ediyor.[10]
Ayak

Diğer theropodlar gibi, dromaeosauridler de iki ayaklıydı; yani arka ayakları üzerinde yürüdüler. Bununla birlikte, theropodların çoğu yere temas eden üç ayak parmağıyla yürürken, fosilleşmiş ayak izi izleri paravyan dromaeosauridler de dahil olmak üzere gruplar, hayvanın ağırlığını yalnızca üçüncü ve dördüncü ayak parmakları ile aşırı genişletilmiş bir pozisyonda yerden tuttu. Buna işlevsel öğretici olarak denir.[11] Büyütülmüş ikinci ayak parmağı alışılmadık derecede büyük, kavisli, falciform (orak biçimli, alt. drepanoid) Avı yakalamada ve ağaçlara tırmanmada kullanıldığı düşünülen pençe (yerden tutulmuş veya "geri çekilmiş") (aşağıdaki "Pençe işlevi" bölümüne bakın). Bu pençe özellikle geniş gövdeli yırtıcı hayvanlarda bıçak gibiydi. eudromaeosaurs.[12] Olası bir dromaeosaurid türü, Balaur bondoc ayrıca ikinciye paralel olarak oldukça modifiye edilmiş bir ilk parmağa sahipti. Her ayağın hem birinci hem de ikinci ayak parmakları B. bondoc ayrıca geri çekilmiş ve büyütülmüş, orak şeklindeki pençelere sahipti.[13]
Kuyruk
Dromaeosauridlerin uzun kuyrukları vardı. Kuyruk omurlarının çoğu, bazı türlerde kemikli, çubuk benzeri uzantıların yanı sıra kemikli tendonlar taşıyordu. Onun çalışmasında Deinonychus Ostrom, bu özelliklerin kuyruğu sertleştirdiğini, böylece sadece tabanda esneyebileceğini ve daha sonra tüm kuyruğun tek, sert bir kaldıraç olarak hareket edeceğini öne sürdü.[6] Bununla birlikte, iyi korunmuş bir örnek Velociraptor mongoliensis (IGM 100/986), uzun S şeklinde yatay olarak kavisli mafsallı bir kuyruk iskeletine sahiptir. Bu, yaşamda kuyruğun önemli derecede bir esneklikle bir yandan diğer yana bükülebileceğini göstermektedir.[14] Bu kuyruğun koşarken veya havada dengeleyici veya karşı ağırlık olarak kullanılması önerilmiştir;[14] içinde Microraptor Kuyruğun ucunda elmas şeklindeki uzun bir tüy yelpazesi korunmuştur. Bu, süzülme veya motorlu uçuş sırasında aerodinamik dengeleyici ve dümen olarak kullanılmış olabilir (aşağıdaki "Uçuş ve süzülme" bölümüne bakın).[15]
Boyut


Dromaeosauridler, uzunlukları yaklaşık 0,7 metre (2,3 ft) arasında değişen küçük ila orta büyüklükteki dinozorlardı. Mahakala ) yaklaşan veya 6 m (20 ft) (inç Utahraptor, Dakotaraptor ve Achillobator ).[16][17] Bazıları daha büyük olabilir; tarifsiz örnekler Utahraptor içinde Brigham Young Üniversitesi Koleksiyonlar, 11 m (36 ft) uzunluğa ulaşmış olabilecek kişilere aitti, ancak bunlar daha ayrıntılı bir çalışmayı bekliyor.[18] Büyük boyut, dromaeosauridler arasında en az iki kez evrimleşmiş görünmektedir; dromaeosaurines arasında bir kez Utahraptor ve Achillobatorve yine laktikler arasında (Austroraptor, 5 m [16 ft] uzunluğunda ölçülen). Muhtemel üçüncü bir dev dromaeosaurid soyu, üzerinde bulunan izole dişlerle temsil edilir. Wight Adası, İngiltere. Dişler, dromaeosaurine büyüklüğünde bir hayvana aittir. Utahraptorancak dişlerin şekline bakılırsa velosiraptorinlere ait gibi görünüyorlar.[19][20]
Mahakala hem şimdiye kadar tanımlanan en ilkel dromaeosaurid hem de en küçüğüdür. Bu kanıt, diğer ilkel akrabaların küçük boyutu ile birleştiğinde Microraptor ve Troodontid Anchiornis, dromaeosauridler, troodontidler ve kuşların ortak atasının (atadan kalma paravyan olarak adlandırılır) çok küçük olabileceğini, yaklaşık 65 cm uzunluğunda ve 600 ila 700 gram kütleye sahip olabileceğini gösterir.[21]
Tüyler

Dromaeosauridlerin kaplandığını gösteren çok sayıda kanıt var. tüyler. Bazı dromaeosaurid fosilleri, ellerde ve kollarda uzun, yapraklı tüyleri korur (Remiges) ve kuyruk (dikdörtgenler) ve vücudu kaplayan daha kısa, tüy gibi tüyler.[10][22] Gerçek tüy izlenimlerini korumayan diğer fosiller, yaşamda uzun kanat tüylerinin yapışmış olacağı önkol kemiklerindeki ilgili çıkıntıları hala korumaktadır.[23] Genel olarak, bu tüy deseni çok benziyor Archæopteryx.[10]
Kesin tüy kanıtı olan ilk bilinen dromaeosaurid, Sinornithosaurus, bildirildi Çin Xu tarafından et al. 1999'da.[22] Diğer birçok dromaeosaurid fosili, vücutlarını örten tüylerle, bazıları tamamen gelişmiş tüylü kanatlarla bulundu. Microraptor hatta arka ayaklarda ikinci bir çift kanat olduğuna dair kanıtlar var.[10] Doğrudan tüy izlenimi yalnızca ince taneli tortularda mümkün olsa da, daha iri kayalarda bulunan bazı fosiller, bazı kuşların sahip olduğu kanat tüyleri için bağlantı noktaları olan tüy yumrularının varlığıyla tüylerin kanıtını göstermektedir. Dromaeosauridler Rahonavis ve Velociraptor Her ikisi de tüy topuzlarıyla birlikte bulunmuş, bu da hiçbir izlenim bulunmamasına rağmen bu formların tüylere sahip olduğunu göstermektedir. Bunun ışığında, büyük olasılıkla yerde yaşayan daha büyük dromaeosauridler bile tüy taşıyorlar, çünkü bugün uçamayan kuşlar bile tüylerinin çoğunu ve nispeten büyük dromaeosauridleri koruyorlar. Velociraptor, yapraklı tüyleri muhafaza ettiği bilinmektedir.[23][24] Bazı bilim adamları, daha büyük dromaeosauridlerin yalıtım örtülerinin bir kısmını veya tamamını kaybettiklerini öne sürdüler. Velociraptor Örnekler, ailenin tüm üyelerinin tüyleri tuttuğunun kanıtı olarak gösterildi.[23][25]
Daha yakın zamanlarda, keşfi Zhenyuanlong nispeten büyük dromaeosauridlerde tam tüylü bir kaplamanın varlığını kurdu. Ek olarak, hayvan orantılı olarak büyük, aerodinamik kanat tüyleri ve kuyruk boyunca uzanan bir yelpaze sergiliyor; bunların her ikisi de büyük dromaeosauridlerin bütünlüğünü anlamayı sağlayabilecek beklenmedik özelliklerdir.[26] Dakotaraptor tüy yumruları şeklinde dolaylı olsa da tüylerin kanıtı olan daha da büyük bir dromaeosaurid türüdür.[27]
Teknik teşhis

Dromaeosauridler aşağıdaki özelliklerle teşhis edilir; kısa T-şekilli cephe bu form rostral sınırı supratemporal fenestra; kaudolateral sarkan raf skuamozal; yanal bir süreç uydurmak o temas dörtlü; üzerinde yükseltilmiş, takip edilmiş, parapofizler sırt omurları değiştirilmiş bir pedal rakamı II; köşeli çift ayraçlar ve prezygapophysis of kaudal omur uzun ve birkaç omuru kapsayan; üzerinde subglenoid fossa varlığı korakoid.[9]
Sınıflandırma
Kuşlarla ilişki

Dromaeosaurids, birçok özelliği erken kuşlar (clade Avialae veya Aves ). Kuşlarla olan ilişkilerinin kesin doğası çok fazla araştırıldı ve bu ilişki hakkındaki hipotezler, büyük miktarlarda yeni kanıtlar ortaya çıktıkça değişti. 2001 gibi geç bir tarihte, Mark Norell ve meslektaşları büyük bir anket analiz etti Coelurosaur fosiller oluşturmuş ve dromaeosauridlerin kuşlarla en yakın akraba olduklarına dair kesin bir sonuç vermiştir. Troodontidler daha uzak bir grup olarak. Hatta Dromaeosauridae'nin parafiletik Avialae'ye göre.[28] 2002'de Hwang ve meslektaşları Norell'in çalışmalarını kullandı. et al.kuşların (avialanlar) dromaeosauridlerin kuzeni olarak daha iyi düşünüldüğünü belirlemek için yeni karakterler ve daha iyi fosil kanıtları da dahil olmak üzere Troodontidler.[16] Paleontologların bir fikir birliği, herhangi bir dromaeosauridin uçup uçamayacağını veya süzülüp uçamayacağını veya yapabilecek atalardan evrimleşip evrimleşmediklerini belirlemek için henüz yeterli kanıt olmadığı sonucuna varmıştır.[29]
Alternatif teoriler ve uçamama
Dromaeosauridler o kadar kuşa benzerler ki, bazı araştırmacıları kuş olarak daha iyi sınıflandırılabileceklerini iddia etmeye yönlendirdiler. Birincisi, tüylere sahip oldukları için, dromaeosauridler (diğer birçok coelurosaurian theropod dinozorları ile birlikte), "kuş" veya "Aves" kelimesinin geleneksel tanımlarına göre tüylerin bulundurulmasına dayanan "kuşlar" dır. Bununla birlikte, Lawrence Witmer gibi diğer bilim adamları, bir theropodu şöyle çağırmanın Caudipteryx Kuş tüyleri olduğu için kelimeyi herhangi bir yararlı anlamın ötesine uzatabilir.[30]

En az iki araştırmacı okulu, dromaeosauridlerin aslında uçan atalardan gelebileceğini öne sürdü. Dromaeosaurids için uçan bir atayı içeren hipotezlere bazen "Önce Kuşlar" (BCF) adı verilir. George Olshevsky, genellikle BCF'nin ilk yazarı olarak anılır.[31] Kendi işinde Gregory S. Paul tüm grubun uçan, dinozor atalarından, belki de benzeri bir hayvandan evrimleştiğinin kanıtı olarak yorumladığı dromaeosaurid iskeletinin sayısız özelliğine işaret etti. Archæopteryx. Bu durumda, daha büyük dromaeosauridler, modern gibi ikincil olarak uçamazdı. devekuşu.[24] 1988'de Paul, dromaeosauridlerin aslında modern kuşlarla daha yakından ilişkili olabileceğini öne sürdü. Archæopteryx. Ancak 2002 yılına kadar Paul, dromaeosauridleri yerleştirdi ve Archæopteryx birbirlerine en yakın akraba olarak.[32]
2002 yılında, Hwang et al. bulundu Microraptor en ilkel dromaeosaurid idi.[16] Xu ve arkadaşları 2003 yılında, Microraptor, tüy ve kanat özellikleriyle birlikte, atalardan kalma dromaeosaurid'in süzüldüğünün kanıtı olarak. Bu durumda, daha büyük dromaeosauridler ikincil olarak karasal olacaktır - evrimsel tarihlerinde daha sonra süzülme yeteneklerini kaybettiler.[10]
Ayrıca 2002'de Steven Czerkas, Kriptovanlar Muhtemelen küçük eşanlamlı olmasına rağmen Microraptor. Fosili yanlış bir şekilde sadece iki kanatla yeniden inşa etti ve böylece dromaeosauridlerin pasif planörlerden ziyade güçlendirilmiş uçucular olduğunu savundu. Daha sonra yeniden yapılanma ile uyumlu olarak revize edilmiş bir yeniden yapılanma yayınladı. Microraptor[33]
Gibi diğer araştırmacılar Larry Martin, dromaeosauridlerin tüm maniraptoranlarla birlikte dinozor olmadığını öne sürdüler. Martin onlarca yıldır kuşların maniraptoranlarla alakası olmadığını iddia etti, ancak 2004'te ikisinin yakın akraba olduğunu kabul ederek pozisyonunu değiştirdi. Ancak Martin, maniraptoranların ikincil olarak uçamayan kuşlar olduğuna ve kuşların dinozorlardan değil, dinozor olmayan arkozorlardan evrimleştiğine inanıyordu.[34]

2005 yılında Mayr ve Peters, çok iyi korunmuş bir örneğin anatomisini tanımladılar. Archæopteryxve anatomisinin daha önce anlaşılandan daha çok kuş olmayan theropodlara benzediğini belirledi. Özellikle şunu buldular: Archæopteryx ilkel bir palatin, geri alınmamış halluks ve hiper-uzayabilir ikinci ayak parmağı. Filogenetik analizleri tartışmalı sonucu üretti: Konfüçyüs daha yakındı Microraptor daha Archæopteryx, Avialae'yi parafiletik bir takson yapıyor. Ayrıca atalardan kalma paravianın uçabildiğini veya süzüldüğünü ve dromaeosauridlerin ve troodontidlerin ikincil olarak uçamadıklarını (veya süzülme yeteneklerini kaybettiklerini) öne sürdüler.[36][37] Corfe ve Butler, bu çalışmayı metodolojik gerekçelerle eleştirdi.[38]
Turner ve meslektaşları 2007'de yeni bir dromaeosaurid tanımladığında, tüm bu alternatif senaryolara bir meydan okuma geldi, Mahakala Dromaeosauridae'nin en temel ve en ilkel üyesi olduğunu buldukları, Microraptor. Mahakala kısa kolları vardı ve kayma yeteneği yoktu. Turner et al. ayrıca, uçuşun yalnızca Avialae'de evrimleştiği sonucuna vardı ve bu iki nokta, ata dromaeosaurid'in süzülüp uçamayacağını öne sürdü. Bu kladistik analize dayanarak, Mahakala dromaeosaurids için atadan kalma koşulunuçucu.[39] Bununla birlikte, 2012 yılında, en son dromaeosaurid bulgularını içeren genişletilmiş ve revize edilmiş bir çalışma, Archæopteryx-sevmek Xiaotingia Dromaeosauridae kuşağının en ilkel üyesi olarak, kladın en eski üyelerinin uçma yeteneğine sahip olabileceğini düşündürüyor gibi görünüyor.[40]
Taksonomi
Dromaeosauridae ailesinin yazarlığı, William Diller Matthew ve Barnum Brown, onu ailenin bir alt ailesi (Dromaeosaurinae) olarak kuran Deinodontidae 1922'de sadece yeni cinsi içeren Dromaeosaurus.[41]
Dromaeosauridae'nin alt aileleri, yeni analize dayalı olarak içerikte sıklıkla değişir, ancak tipik olarak aşağıdaki gruplardan oluşur. Bazı dromaeosauridler, belirli bir alt aileye atanmamıştır, çünkü çoğu zaman, güvenle yerleştirilemeyecek kadar zayıf bir şekilde korunmuşlardır. filogenetik analiz (bkz. bölüm Filogeni aşağıda) veya belirsizdir, farklı makalelerde kullanılan metodolojiye bağlı olarak farklı gruplara atanır. Dromaeosauridlerin bilinen en bazal alt familyası, uzun parmakları ve boyunları olan, çok sayıda küçük dişleri ve olası yarı karasal alışkanlıkları olan bir grup tuhaf yaratık olan Halszkaraptorinae'dir.[42] Diğer bir esrarengiz grup olan Unenlagiinae, dromaeosauridlerin en zayıf şekilde desteklenen alt familyasıdır ve üyelerinin bir kısmının veya tamamının Dromaeosauridae'nin dışında olması mümkündür.[21][43] Daha büyük, yerde yaşayan üyeler gibi Buitreraptor ve Unenlagia Muhtemelen "havalanmak" için çok büyük olmalarına rağmen, güçlü uçuş uyarlamaları gösterirler. Bu grubun olası bir üyesi, Rahonavis, çok küçüktür, tüy topuzlarının (uçuş tüyleri için bağlantı noktaları) kanıtlarını gösteren iyi gelişmiş kanatlara sahiptir ve uçma olasılığı çok yüksektir. Bir sonraki en ilkel dromaeosaurid türü Microraptoria'dır. Bu grup, ağaçlarda yaşamak için uyarlamalar gösteren en küçük dromaeosauridlerin çoğunu içerir. Bilinen tüm dromaeosaurid deri izlenimleri bu gruptan geliyor ve tümü geniş bir tüy örtüsünü ve iyi gelişmiş kanatları gösteriyor. Unenlagiines gibi, bazı türler aktif olarak uçma yeteneğine sahip olabilir. Dromaeosauridlerin en gelişmiş alt grubu olan Eudromaeosauria, muhtemelen pusu avcıları olan tıknaz ve kısa bacaklı cinsleri içerir. Bu grup Velociraptorinae, Dromaeosaurinae'yi ve bazı çalışmalarda üçüncü bir grubu içerir: Saurornitholestinae. Velociraptorinae alt ailesi geleneksel olarak şunları içerir: Velociraptor, Deinonychus, ve Saurornitholestes ve keşfedilirken Tsaagan bu gruplamaya destek verdi, dahil Deinonychus, Saurornitholestes, ve diğer birkaç cins hala belirsizdir. Dromaeosaurinae'nin genellikle orta ila dev boyutlu türlerden oluştuğu ve genellikle kutu şeklindeki kafatasları olduğu bulunmuştur (diğer alt ailelerin genellikle daha dar burunları vardır).[1]





Çeşitli dromaeosaurid cinslerinin aşağıdaki sınıflandırması, aksi belirtilmedikçe Holtz, 2011'de verilen tabloyu takip eder.[1]
- Aile Dromaeosauridae
Cinslerin tarihi
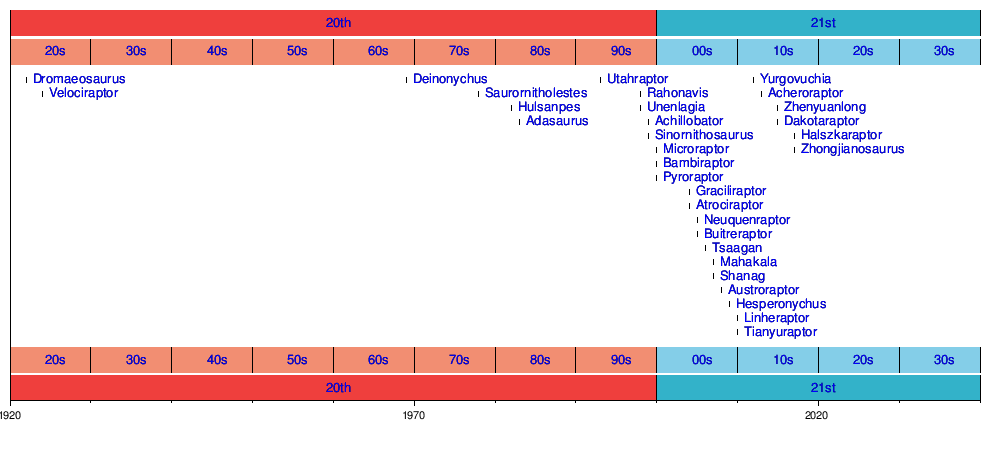
Filogeni
Dromaeosauridae ilk olarak bir clade tarafından Paul Sereno 1998 yılında en kapsayıcı doğal grup olarak Dromaeosaurus Ama değil Troodon, Ornitomimus veya Pasör. Çeşitli "alt aileler" de sınıflar olarak yeniden tanımlanmış, genellikle grup adaşına daha yakın olan tüm türler olarak tanımlanmıştır. Dromaeosaurus veya diğer alt sınıflardan herhangi bir adaş (örneğin, Makovicky, Unenlagiinae sınıfını, yakınlardaki tüm dromaeosauridler olarak tanımlamıştır) Unenlagia daha Velociraptor ). Microraptoria, bir alt aileden dönüştürülmemiş tek dromaeosaurid alt sınıftır. Senter ve meslektaşları, adı alt aile soneki olmadan açıkça uydurdular. -inae bir montajla ilgili algılanan sorunları önlemek için geleneksel aile grubu taksonu, grubun uygun dromaeosauridae dışında bulunduğu tespit edilirse.[45] Sereno, aşağıdakileri içeren alt grubun gözden geçirilmiş bir tanımını sundu Microraptor Dromaeosauridae'ye girmesini sağlamak ve Microraptorinae alt ailesini kurarak onu Senter'e atfedmek et al.ancak bu kullanım yalnızca çevrimiçi TaxonSearch veritabanında görünmüş ve resmi olarak yayınlanmamıştır.[46] Turner tarafından yürütülen kapsamlı kladistik analiz et al. (2012) ayrıca Dromaeosauridae'nin monofiliğini destekledi.[47]
Aşağıdaki kladogram, DePalma ve diğerleri tarafından yapılan 2015 analizini takip eder. Theropod Çalışma Grubundan güncellenmiş verileri kullanarak.[27]
| Dromaeosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Aşağıda inşa edilen başka bir cladogram, 2017 yılında Cau tarafından yapılan filogenetik analizi takip eder. et al. Theropod Çalışma Grubundan güncellenmiş verileri kullanarak Halszkaraptor.[42]
| Dromaeosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Paleobiyoloji
Pençe işlevi
Şu anda ikinci ayak parmağındaki genişlemiş "orak pençe" nin işlevi konusunda bir anlaşmazlık var. John Ostrom bunu şöyle tarif ettiğinde Deinonychus 1969'da pençeyi bıçak benzeri bir kesen silah olarak yorumladı, bazılarının köpek dişleri gibi kılıç dişli kediler, güçlü tekmelerle avı kesmek için kullanılır. Adams (1987), talonun geniş çaplı iç organları sökmek için kullanıldığını öne sürdü. Ceratopsian dinozorlar.[48] Orak pençenin bir öldürme silahı olarak yorumlanması tüm dromaeosauridlere uygulandı. Ancak, Manning et al. pençenin bunun yerine bir kanca görevi gördüğünü, keratinli kılıfı daha önce elde edilen ters gözyaşı damlası şekli yerine eliptik bir enine kesitle yeniden yapılandırdığını savundu.[49] Manning'in yorumuna göre, ikinci ayak parmağı pençesi, daha büyük bir avı bastırırken tırmanma yardımı olarak ve ayrıca bir bıçaklama silahı olarak kullanılacaktır.

Ostrom karşılaştırıldığında Deinonychus için devekuşu ve kasırga. Kuş türlerinin ikinci ayak parmağındaki büyük pençe ile ciddi yaralanmalara neden olabileceğini kaydetti.[6] Cassowary, 125 milimetre (4,9 inç) uzunluğa kadar pençelere sahiptir.[50] Ostrom, Gilliard'ı (1958) bir kolunu kesebileceklerini veya bir erkeğin karnını sökebileceklerini söylerken aktardı.[51] Kofron (1999 ve 2003), 241 belgelenmiş kasırga saldırısını inceledi ve bir insan ve iki köpeğin öldürüldüğünü buldu, ancak kasırgaların diğer hayvanların içlerini parçalayabileceğine veya parçalayabileceğine dair bir kanıt yok.[52][53] Kasavarlar pençelerini kendilerini savunmak, tehdit eden hayvanlara saldırmak için ve Bowed Threat Display gibi agonist gösterilerde kullanırlar.[50] Seriema ayrıca genişletilmiş bir ikinci parmak pençesine sahiptir ve onu yutmak için küçük av öğelerini parçalamak için kullanır.[54]
Phillip Manning ve arkadaşları (2009), orak pençe ve benzer şekilli pençelerin ön ayaklardaki işlevini test etmeye çalıştılar. Baskıların ve gerilmelerin pençeler boyunca ve uzuvlara nasıl dağıtılacağına dair biyo-mekaniği analiz ettiler. Röntgen ön ayak pençesinin üç boyutlu bir kontur haritasını oluşturmak için görüntüleme Velociraptor. Karşılaştırma için, modern bir yırtıcı kuşun pençesinin yapısını analiz ettiler. kartal baykuş. Pençe boyunca uygulanan strese dayanarak, tırmanmak için ideal olduklarını buldular. Bilim adamları, pençenin keskin ucunun bir delme ve kavrama aracı olduğunu, kavisli ve genişletilmiş pençe tabanının stres yüklerinin eşit olarak aktarılmasına yardımcı olduğunu buldu.[55]
Manning ekibi ayrıca, dromaeosaurid "orak pençesinin" ayaktaki eğriliğini modern kuşlar ve memelilerdeki eğrilik ile karşılaştırdı. Önceki çalışmalar, bir pençedeki eğrilik miktarının, hayvanın sahip olduğu yaşam tarzına karşılık geldiğini göstermişti: belirli bir şekle sahip güçlü kavisli pençelere sahip hayvanlar tırmanma eğilimindeyken, daha düz pençeler yerde yaşayan yaşam tarzlarını gösterir. Dromaeosaurid'in orak pençeleri Deinonychus 160 derecelik bir eğriliği var ve bu da tırmanan hayvanların menzilinde. İnceledikleri ön ayak pençeleri de eğriliğin tırmanma aralığına düştü.[55]
Paleontolog Peter Mackovicky, Manning ekibinin çalışmasına yorum yaparak, küçük, ilkel dromaeosauridlerin (örneğin Microraptor ) muhtemelen ağaç tırmanıcıydılar, ancak bu tırmanma neden daha sonra nedenini açıklamadı, örneğin devasa dromaeosauridler Achillobator ağaçlara tırmanmak için çok büyük olduklarında oldukça kıvrımlı pençeleri korudular. Mackovicky, dev dromaeosauridlerin pençeyi yalnızca avı mandallamak için kullanacak şekilde uyarlamış olabileceğini tahmin etti.[56]
2009 yılında Phil Senter, dromaeosaurid ayak parmakları üzerine bir çalışma yayınladı ve hareket açıklıklarının sert böcek yuvalarının kazılmasıyla uyumlu olduğunu gösterdi. Senter, küçük dromaeosauridlerin Rahonavis ve Buitreraptor kısmi olacak kadar küçüktü böcekçiller gibi daha büyük cinsler ise Deinonychus ve Neuquenraptor bu yeteneği yakalamak için kullanabilirdi omurgalı böcek yuvalarında yaşayan av. Bununla birlikte, Senter, dromaeosaurid pençelerinin güçlü eğriliğinin bu tür faaliyetler için de uygun olup olmadığını test etmedi.[57]
2011'de Denver Fowler ve meslektaşları, dromaeosauridlerin daha küçük avlar alabilecekleri yeni bir yöntem önerdiler. Yırtıcı hayvanın "yırtıcı avını kısıtlama" (RPR) modeli olarak bilinen bu model, dromaeosauridlerin avlarını günümüze çok benzer bir şekilde öldürdüğünü öne sürüyor. Accipitrid yırtıcı kuşlar: taş ocağına atlayarak, onu vücut ağırlıklarının altına sıkıştırarak ve büyük, orak biçimli pençelerle sıkıca kavrayarak. Accipitrids gibi, dromaeosaurid de daha sonra kan kaybından ve organ yetmezliğinden ölünceye kadar canlıyken hayvanla beslenmeye başlayacaktı. Bu öneri, öncelikle, dromaeosauridlerin morfolojisi ve oranları ile bilinen yırtıcı davranışları olan birkaç avcı kuş grubu arasındaki karşılaştırmalara dayanmaktadır. Fowler, dromaeosauridlerin ayaklarının ve bacaklarının, en çok kartallar ve şahinler özellikle genişletilmiş bir ikinci pençe ve benzer bir kavrama hareketi aralığı açısından. Kısa metatars ve ayak kuvveti, bununla birlikte, baykuşlar. RPR avlanma yöntemi, alışılmadık diş yapısı ve kol morfolojisi gibi dromaeosaurid anatomisinin diğer yönleriyle tutarlı olacaktır. Çok fazla güç uygulayabilen ancak muhtemelen uzun tüylerle kaplı olan kollar, sert dengeleme kuyruğunun yanı sıra mücadele eden bir av hayvanının tepesinde denge için kanat çırpma dengeleyicileri olarak kullanılmış olabilir. Fowler ve meslektaşları tarafından nispeten zayıf olduğu düşünülen dromaeosaurid çeneleri, avı canlı canlı yemek için yararlı olabilirdi, ancak avın hızlı ve güçlü bir şekilde gönderilmesi için o kadar yararlı değildi. Birlikte çalışan bu yıkıcı uyarlamaların aynı zamanda kanat çırpmanın kökeni içinde paravanlar.[58][59]
Gianechini tarafından yapılan bir 2020 çalışması ve diğerleri., ayrıca Laurasia'daki velociraptorinlerin, dromaeosaurinlerin ve diğer eudromaeosaurların lokomotif ve öldürme tekniklerinde Gondwana'daki unenlagiine dromaeosauridlerden büyük ölçüde farklı olduğunu gösterir. Ayağın ikinci basamağındaki daha kısa ikinci falanks, daha kısa ve daha geniş bir metatarsus ve muhtemelen metatarsalların ve falanksların eklem yüzeylerinin belirgin bir menteşe benzeri morfolojisi ile birleşen bu basamak tarafından daha fazla kuvvet üretilmesine izin verdi. eudromaeosaurların unenlagiine'lerden daha büyük bir kavrama gücü uygulamasına izin vererek, büyük avların daha etkili bir şekilde bastırılmasına ve öldürülmesine izin verdi. Buna karşılık, unenlagiine dromaeosauridler daha uzun ve ince bir subarktometatarsusa ve daha az iyi işaretlenmiş menteşe eklemlerine sahipti, bu da onlara muhtemelen daha büyük kürsör kapasiteleri veren ve daha fazla hıza izin veren bir özellikti. Ek olarak, ikinci basamağın daha uzun olan ikinci falanksı, unenlagiines'in daha küçük ve daha zor av türlerini avlamak için ayaklarının ikinci basamaklarının hızlı hareketlerine izin verdi. Lokomotor ve yırtıcı uzmanlaşmalardaki bu farklılıklar, kuzey ve güney yarım kürelerde her iki dromaeosaur grubunu da şekillendiren evrimsel yolları etkileyen önemli bir özellik olabilir.[60]
Besleme
Dromaeosaurid beslemesinin, karakteristik bir "delin ve çekerek" besleme yöntemiyle, coelurosaurian theropodlara özgü olduğu keşfedildi. Angelica Torices tarafından dromaeosauridlerin dişlerindeki aşınma modellerinin incelenmesi et al. dromaeosaurid dişlerin Tyrannosauridae ve Troodontidae'de görülenlere benzer aşınma modellerini paylaştığını belirtmektedir. Bununla birlikte, dişlerdeki mikro dalga, dromaeosauridlerin muhtemelen çevrelerini paylaştıkları troodontidlerden daha büyük av ürünlerini tercih ettiklerini gösterdi. Bu tür beslenme farklılıkları muhtemelen aynı ortamda yaşamalarına izin verdi. Aynı çalışma ayrıca dromaeosauridlerin Dromaeosaurus ve Saurornitholestes (çalışmada analiz edilen iki dromaeosaurid) muhtemelen diyetlerine kemiği dahil etti ve daha zayıf çenelerle donatılmış troodontidler daha yumuşak hayvanları ve omurgasızlar ve leş gibi avları avlarken, mücadele eden avları idare etmek için daha iyi adapte oldular.[61]
Grup davranışı

Deinonychus otobur kalıntılarının yakınında küçük gruplar halinde fosiller ortaya çıkarılmıştır. Tenontosaurus, daha geniş ornitorik Dinozor. Bu, bu dromaeosauridlerin bazı modern türler gibi eşgüdümlü paketler halinde avlandıklarının kanıtı olarak yorumlanmıştı. memeliler.[63] Ancak hepsi değil paleontologlar Kanıtı kesin buldu ve 2007'de Roach ve Brinkman tarafından yayınlanan sonraki bir çalışma, Deinonychus aslında düzensiz bir mobbing davranışı sergilemiş olabilir. Modern diyapsitler, dahil olmak üzere kuşlar ve timsahlar (dromaeosauridlerin en yakın akrabaları), minimum uzun vadeli ortak avlanma sergiler ( aplomado şahin ve Harris'in şahin ); bunun yerine, genellikle ya avlanma başarısını artırmak için zaman zaman güçlerini birleştiren (timsahların bazen yaptığı gibi) ya da aynı türden bireyler arasında sıklıkla çatışmanın meydana geldiği daha önce öldürülmüş leşleri çeken yalnız avcılardır. Örneğin, grupların bulunduğu durumlarda Komodo ejderhaları birlikte yiyorlarsa, en büyük bireyler önce yer ve beslenmeye çalışan daha küçük Komodo ejderhalarına saldırabilir; daha küçük hayvan ölürse, genellikle yamyam. Bu bilgi, dromaeosauridlerde varsayılan paket avlanma davranışını içeren sitelere uygulandığında, Komodo ejderhası benzeri bir besleme stratejisi ile bir şekilde tutarlı görünmektedir. Deinonychus Bu yerlerde bulunan iskelet kalıntıları, alt yetişkinlere aittir ve başkaları tarafından yenmiş olabilecek eksik parçalar Deinonychus Roach tarafından yapılan bir çalışma et al. hayvanların avda işbirliği yaptığı fikrine karşı delil olarak sunuldu.[64] Yavru ve yetişkin arasında farklı beslenme tercihleri Deinonychus 2020'de yayımlanan yayın, hayvanın paket avlanan hayvanlarda görülen karmaşık, işbirlikçi davranışlar sergilemediğini göstermektedir. Bunun diğer dromaeosaurlara yayılıp yayılmadığı şu anda bilinmemektedir.[65] Üçüncü bir olası seçenek, dromaeosauridlerin uzun vadeli işbirliği davranışı sergilememesi, ancak av için hem gerçek işbirliği hem de rekabet gösteren timsahlarda görüldüğü gibi kısa vadeli işbirliği davranışı göstermesidir.
2001 yılında, bazıları tarafından bataklık ve avcı tuzağı olarak kabul edilen bir bölgede, yaşları tamamen yetişkin bir yetişkinden üç ayak uzunluğundaki minik bebeğe kadar değişen çok sayıda Utahraptor örneği bulundu. Bazıları bunu aile avlanma davranışının kanıtı olarak görüyor; ancak, kumtaşı bloğunun tamamı henüz açılmadı ve araştırmacılar, hayvanların aynı anda ölüp ölmediğinden emin değiller.[66]
2007'de bilim adamları bilinen ilk geniş dromaeosauridi tanımladılar yol, Shandong, Çin'de. Orak pençesinin yerden geri çekilmiş olduğu hipotezini doğrulamaya ek olarak, yol (büyük bir Achillobator boyutundaki türler), kıyı şeridi boyunca yaklaşık eşit büyüklükte altı bireyin birlikte hareket ettiğine dair kanıtlar gösterdi. Bireyler birbirlerinden yaklaşık bir metre uzakta, aynı yönde seyahat ediyor ve oldukça yavaş bir hızda yürüyorlardı. Bu ayak izlerini tanımlayan makalenin yazarları, iz yollarını bazı dromaeosaurid türlerinin gruplar halinde yaşadığının kanıtı olarak yorumladılar. Yollar açıkça avlanma davranışını temsil etmese de, yazarlara göre dromaeosaurid gruplarının birlikte avlanmış olabileceği fikri göz ardı edilemez.[11]
Uçmak ve süzülmek
Yeteneği uç ya da süzül en az beş dromaeosaurid türü için önerilmiştir. İlk, Rahonavis ostromi (başlangıçta kuş kuşu olarak sınıflandırıldı, ancak daha sonraki çalışmalarda bir dromaeosaurid olduğu bulundu[8][67]) güçlendirilmiş olabilir uçuş, uzun, sağlam uçuş tüyleri için tüy topuzu eklerinin kanıtı olan uzun ön ayaklarıyla gösterildiği gibi.[68] Ön ayakları Rahonavis daha güçlü bir şekilde inşa edildi Archæopteryxve kanat çırpma uçuşu için gerekli olan güçlü bağ bağlantılarını taşıdıklarına dair kanıt gösterin. Luis Chiappe, bu uyarlamalar göz önüne alındığında, Rahonavis muhtemelen uçabilirdi ama havada modern kuşlardan daha sakar olurdu.[69]

Başka bir dromaeosaurid türü, Microraptor gui iyi gelişmiş kanatlarını hem ön hem de arka bacaklarda kullanarak süzülme yeteneğine sahip olabilir. 2005 tarihli bir çalışma Sankar Chatterjee kanatlarını önerdi Microraptor iki seviyeli gibi çalıştı "çift kanatlı uçak "ve muhtemelen bir fugoid bir levrekden fırlayıp U şeklindeki bir eğri üzerinde aşağıya doğru süzüldüğü, daha sonra başka bir ağaca inmek için tekrar yükseldiği, kuyruğu ve arka kanatları konumunu ve hızını kontrol etmeye yardımcı olacak şekilde süzülme tarzı. Chatterjee ayrıca şunu da buldu Microraptor Süzülmeye ek olarak düz uçuşu sürdürmek için temel gereksinimlere sahipti.[15]
Changyuraptor yangi yakın akrabası Microraptor gui Ayrıca, dört kanat ve benzer uzuv oranlarının varlığına dayanarak bir planör veya el ilanı olduğu düşünülüyordu. Bununla birlikte, oldukça büyük bir hayvandır, yaklaşık bir vahşi Türkiye, bilinen en büyük uçan Mesozoyik paravianlar arasındadır.
Başka bir dromaeosaurid türü, Deinonychus antirrhopus, kısmi uçuş kapasiteleri gösterebilir. Bu türün gençleri, yetişkinlere göre daha uzun kollara ve daha sağlam göğüs kuşağına sahipti ve diğer çırpınan theropodlarda görülenlere benziyordu, bu da gençken uçabildikleri ve büyüdükçe yeteneklerini yitirdikleri anlamına geliyordu.[70]
Olasılığı Sinornithosaurus millenii Süzülme kabiliyetine sahipti ve hatta motorlu uçuş da birkaç kez gündeme getirildi,[71][72] daha fazla çalışma yapılmamasına rağmen.
Zhenyuanlong Aerodinamik olarak şekillendirilmiş kanat tüylerini, özellikle kuş benzeri örtülerle, daha uzun, daha geniş kapsayan örtülerin aksine korur. Archæopteryx ve Anchiornis yanı sıra kaynaşmış sternal plakalar. Büyüklüğü ve kısa kolları nedeniyle Zhenyuanlong motorlu uçuş yeteneğine sahipti (bu bağlamda biyomekanik modellemenin önemi vurgulanmış olsa da)[26]), ancak uçan atalardan nispeten yakın bir alçalmayı, hatta biraz kayma veya kanat destekli eğimli koşu.
Duyular
Arasındaki karşılaştırmalar skleral halkalar birkaç dromaeosauridsin (Microraptor, Sinornithosaurus, ve Velociraptor ) ve modern kuşlar ve sürüngenler, bazı dromaeosauridlerin ( Microraptor ve Velociraptor ) olabilir Gece gündüz yırtıcılar Sinornithosaurus olduğu anlaşılıyor katemeral (gün boyunca kısa aralıklarla aktiftir).[73] Ancak, yanardöner tüylerin keşfi Microraptor yanardöner tüylere sahip hiçbir modern kuşun gece olduğu bilinmediğinden, bu cinste gecelilik çıkarımı konusunda şüphe uyandırmıştır.[74]
Çalışmaları koku soğanları Dromaeosauridlerin% 'si, boyutlarına göre diğer kuş olmayanlara benzer koku alma oranlarına sahip olduklarını ortaya koymaktadır. theropodlar ve keskin bir koku alma duyusuna sahip modern kuşlar, örneğin tyrannosaurids ve hindi akbabası muhtemelen önemini yansıtan koku alma dromaeosauridlerin yiyecek bulma gibi günlük aktivitelerini hissedin.[75][76]
Paleopatoloji
2001 yılında, Bruce Rothschild ve diğerleri, aşağıdakiler için kanıtları inceleyen bir çalışma yayınladılar: Gerilme kırıkları ve tendon avülsiyonları içinde Theropod dinozorlar ve davranışlarının etkileri. Since stress fractures are caused by repeated trauma rather than singular events they are more likely to be caused by regular behavior than other types of injuries. The researchers found lesions like those caused by stress fractures on a dromaeosaurid hand claw, one of only two such claw lesions discovered in the course of the study. Stress fractures in the hands have special behavioral significance compared to those found in the feet, since stress fractures in the feet can be obtained while running or during migration. Hand injuries, by contrast, are more likely to be obtained while in contact with struggling prey.[77]
Yüzme
En az bir tür, Halszkaraptor escuilliei, appears to have been specialised for aquatic habits, having developed limb proportions akin to those of diving birds.[42] Fishing habits have been proposed for unenlagiines, including comparisons to the unambiguously aquatic spinosaurs,[78] but any aquatic propulsion mechanisms have not been discussed so far.
Üreme
In 2006, Grellet-Tinner and Makovicky reported an egg associated with a specimen of Deinonychus. The egg shares similarities with oviraptorid eggs, and the authors interpreted the association as potentially indicative of brooding.[79] A study published in November 2018 by Norell, Yang and Wiemann et al., indicates that Deinonychus laid blue eggs, likely to camouflage them as well as creating open nests. Other dromaeosaurids may have done the same, and it is theorized that they and other maniraptoran dinosaurs may have been an origin point for laying colored eggs and creating open nests as many birds do today.[80][81][82]
popüler kültürde
Velociraptor, a dromaeosaurid, gained much attention after it was featured prominently in the 1993 Steven Spielberg film Jurassic Park. However, the dimensions of the Velociraptor in the film are much larger than the largest members of that genus. Robert Bakker recalled that Spielberg had been disappointed with the dimensions of Velociraptor and so upsized it.[83] Gregory S. Paul, in his 1988 book Dünyanın Yırtıcı Dinozorları, also considered Deinonychus antirrhopus bir tür Velociraptor, and so rechristened the species Velociraptor antirrhopus.[32] This taxonomic opinion has not been widely followed.[9][84][85]
Ayrıca bakınız
Referanslar
- ^ a b c Holtz, Thomas R. Jr. (2012) Dinozorlar: Her Yaştan Dinozor Severler için En Eksiksiz, En Güncel Ansiklopedi, Kış 2011 Ek.
- ^ Acorn, J. (2007). Derin Alberta: Fosil Gerçekleri ve Dinozor Kazıları. Alberta Üniversitesi Yayınları. pp.13. ISBN 978-0-88864-481-7.
- ^ Case, Judd A.; Martin, James E.; Reguero, Marcelo (2007). A dromaeosaur from the Maastrichtian of James Ross Island and the Late Cretaceous Antarctic dinosaur fauna. 10th International Symposium on Antarctic Earth Sciences. CiteSeerX 10.1.1.546.3890. doi:10.3133/ofr20071047SRP083.
- ^ "Oz dromaeosaurs(Re: Megaraptor)". dml.cmnh.org. Alındı 28 Mart 2018.[güvenilmez kaynak? ]
- ^ Metcalf, S.J.; Vaughan, R.F.; Benton, M.J .; Cole, J.; Simms, M.J.; Dartnall, D.L. (Ocak 1992). "A new Bathonian (Middle Jurassic) microvertebrate site, within the Chipping Norton Limestone Formation at Hornsleasow Quarry, Gloucestershire". Jeologlar Derneği Bildirileri. 103 (4): 321–342. doi:10.1016/S0016-7878(08)80129-0.
- ^ a b c Ostrom, J. H. (1969). "Osteoloji Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana" (PDF). Peabody Doğa Tarihi Müzesi Bülteni. 30: 1–165.
- ^ Paul, Greg (January 2006). "Drawing Dinosaurs". Coğrafi zamanlar.
- ^ a b Makovicky, Peter J.; Apesteguía, Sebastián; Agnolín, Federico L. (October 2005). "The earliest dromaeosaurid theropod from South America". Doğa. 437 (7061): 1007–1011. Bibcode:2005Natur.437.1007M. doi:10.1038/nature03996. PMID 16222297. S2CID 27078534.
- ^ a b c d Norell, M.A .; Makovicky, P.J. (2004). "Dromaeosauridae". Weishampel, D.B .; Dodson, P .; Osmólska, H. (editörler). Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. pp. 196–210. ISBN 978-0-520-24209-8.
- ^ a b c d e Xu, Xing; Zhou, Zhonghe; Wang, Xiaolin; Kuang, Xuewen; Zhang, Fucheng; Du, Xiangke (January 2003). "Four-winged dinosaurs from China". Doğa. 421 (6921): 335–340. Bibcode:2003Natur.421..335X. doi:10.1038/nature01342. PMID 12540892. S2CID 1160118.
- ^ a b Li, Rihui; Lockley, Martin G .; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A .; Harris, Jerald D.; Liu, Mingwei (19 October 2007). "Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China". Naturwissenschaften. 95 (3): 185–191. Bibcode:2008NW.....95..185L. doi:10.1007/s00114-007-0310-7. PMID 17952398. S2CID 16380823.
- ^ a b Longrich, N.R .; Currie, P.J. (2009). "A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America". PNAS. 106 (13): 5002–7. Bibcode:2009PNAS..106.5002L. doi:10.1073/pnas.0811664106. PMC 2664043. PMID 19289829.
- ^ Csiki, Zoltán; Vremir, Mátyás; Brusatte, Stephen L .; Norell, Mark A. (31 August 2010). "An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 107 (35): 15357–15361. Bibcode:2010PNAS..10715357C. doi:10.1073/pnas.1006970107. PMC 2932599. PMID 20805514.
- ^ a b Norell, Mark A .; Makovicky, Peter J. (1999). "Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis". Amerikan Müzesi Novitates. 3282: 1–45. hdl:2246/3025.
- ^ a b Chatterjee, Sankar; Templin, R. Jack (30 January 2007). "Tüylü dinozor Microraptor gui'nin çift kanatlı plan formu ve uçuş performansı". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 104 (5): 1576–1580. Bibcode:2007PNAS..104.1576C. doi:10.1073 / pnas.0609975104. PMC 1780066. PMID 17242354.
- ^ a b c Hwang, S.H .; Norell, M.A .; Ji, Q .; Gao, K. (2002). "New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) Kuzeydoğu Çin'den ". Amerikan Müzesi Novitates (3381): 44pp. hdl:2246/2870.
- ^ Perle, A.; Norell, M.A .; Clark, J. (1999). "A new maniraptoran theropod – Achillobator giganticus (Dromaeosauridae) – from the Upper Cretaceous of Burkhant, Mongolia". Contributions of the Mongolian-American Paleontological Project. 101: 1–105.
- ^ Britt; Chure; Stadtman; Madsen; Scheetz; Burge (2001). "New osteological data and the affinities of Utahraptor from the Cedar Mountain Fm. (Early Cretaceous) of Utah". Omurgalı Paleontoloji Dergisi. 21 (3): 36A. doi:10.1080/02724634.2001.10010852. S2CID 220414868.
- ^ Naish, D. Hutt, and Martill, D.M. (2001). "Saurischian dinosaurs: theropods." in Martill, D.M. and Naish, D. (eds). Wight Adası Dinozorları. The Palaeontological Association, Field Guides to Fossils. 10, 242–309.
- ^ "Dino hunts net rare raptor teeth". BBC haberleri. 26 April 2004.
- ^ a b Turner, Alan H .; Hwang, Sunny H.; Norell, Mark A. (2007). "A Small Derived Theropod from Öösh, Early Cretaceous, Baykhangor Mongolia". Amerikan Müzesi Novitates. 3557 (1): 1. doi:10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2. hdl:2246/5845.
- ^ a b Xu, Xing; Wang, Xiao-Lin; Wu, Xiao-Chun (September 1999). "A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China". Doğa. 401 (6750): 262–266. Bibcode:1999Natur.401..262X. doi:10.1038/45769. S2CID 4430574.
- ^ a b c Turner, A. H .; Makovicky, P. J .; Norell, M. A. (21 September 2007). "Feather Quill Knobs in the Dinosaur Velociraptor". Bilim. 317 (5845): 1721. Bibcode:2007Sci...317.1721T. doi:10.1126/science.1145076. PMID 17885130.
- ^ a b Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins Üniversitesi Yayınları. 472 s.
- ^ Prum, Richard O .; Brush, Alan H. (September 2002). "The Evolutionary Origin And Diversification Of Feathers". Biyolojinin Üç Aylık İncelemesi. 77 (3): 261–295. doi:10.1086/341993. PMID 12365352.
- ^ a b Lü, Junchang; Brusatte, Stephen L. (16 July 2015). "Çin'in Erken Kretase Dönemi'nden büyük, kısa kollu, kanatlı bir dromaeosaurid (Dinosauria: Theropoda) ve bunun tüy evrimi üzerindeki etkileri". Bilimsel Raporlar. 5 (1): 11775. Bibcode:2015NatSR ... 511775L. doi:10.1038 / srep11775. PMC 4504142. PMID 26181289.
- ^ a b "The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation". Paleontolojik Katkılar. 30 Ekim 2015. doi:10.17161 / paleo.1808.18764.
- ^ Norell, M. Clark, J.M., Makovicky, P.J. (2001). "Phylogenetic relationships among coelurosaurian theropods. " New Perspectives on the Origin and Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom", Yale Peabody Museum: 49–67
- ^ Padian, Kevin; Dial, Kenneth P. (2005). "Origin of flight: Could 'four-winged' dinosaurs fly?". Doğa. 438 (7066): E3, discussion E3–4. Bibcode:2005Natur.438E...3P. doi:10.1038/nature04354. PMID 16292258. S2CID 4314459.
- ^ Witmer, L.M. (2005) "The Debate on Avian Ancestry; Phylogeny, Function and Fossils.” In "Mesozoic Birds: Above the Heads of Dinosaurs”, pp. 3–30. ISBN 0-520-20094-2
- ^ Olshevsky, George. (1994). "The birds first? A theory to fit the facts — evolution of reptiles into birds". Omni, June 1994. Volume 16 No. 9
- ^ a b Paul Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon ve Schuster. 464 s.
- ^ Czerkas, S.A., Zhang, D., Li, J., and Li, Y. (2002). "Flying Dromaeosaurs", in Czerkas, S.J. (ed.): Feathered Dinosaurs and the Origin of Flight: The Dinosaur Museum Journal 1. Blanding: The Dinosaur Museum, 16–26.
- ^ Martin, L.D. (2004). "A basal archosaurian origin for birds" (PDF). Acta Geologica Sinica. 50 (6): 978–990.
- ^ National Geographic News- Earliest Bird Had Feet Like Dinosaur, Fossil Shows – Nicholas Bakalar, December 1, 2005, Page 2. Retrieved 2006-10-18.
- ^ Mayr, G .; Pohl, B; Peters, DS (2 December 2005). "A Well-Preserved Archaeopteryx Specimen with Theropod Features". Bilim. 310 (5753): 1483–1486. Bibcode:2005Sci...310.1483M. doi:10.1126/science.1120331. PMID 16322455. S2CID 28611454.
- ^ Mayr, G. (1 September 2006). "Response to Comment on 'A Well-Preserved Archaeopteryx Specimen with Theropod Features'". Bilim. 313 (5791): 1238c. Bibcode:2006Sci...313.1238M. doi:10.1126/science.1130964.
- ^ Corfe, I. J .; Butler, RJ (1 September 2006). "Comment on 'A Well-Preserved Archaeopteryx Specimen with Theropod Features'". Bilim. 313 (5791): 1238b. Bibcode:2006Sci...313.1238C. doi:10.1126/science.1130800. PMID 16946054. S2CID 19859320.
- ^ Turner, Alan H .; Pol, Diego; Clarke, Julia A .; Erickson, Gregory M .; Norell, Mark (2007). "Bir bazal dromaeosaurid ve kuş uçuşundan önce boyut evrimi". Bilim. 317 (5843): 1378–1381. Bibcode:2007Sci ... 317.1378T. doi:10.1126 / science.1144066. PMID 17823350.
- ^ a b Senter, P .; Kirkland, J. I .; Deblieux, D. D .; Madsen, S .; Toth, N. (2012). Dodson, Peter (ed.). "Utah'ın Alt Kretase Dönemi'nden Yeni Dromaeosaurids (Dinosauria: Theropoda) ve Dromaeosaurid Tail'in Evrimi". PLOS ONE. 7 (5): e36790. Bibcode:2012PLoSO...736790S. doi:10.1371 / journal.pone.0036790. PMC 3352940. PMID 22615813.
- ^ Matthew, W. D.; Brown, B. (1922). "The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta". Amerikan Doğa Tarihi Müzesi Bülteni. 46: 367–385. hdl:2246/1300.
- ^ a b c d Cau, Andrea; Beyrand, Vincent; Voeten, Dennis F. A. E.; Fernandez, Vincent; Tafforeau, Paul; Stein, Koen; Barsbold, Rinchen; Tsogtbaatar, Khishigjav; Currie, Philip J .; Godefroit, Pascal (6 December 2017). "Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs". Doğa. 552 (7685): 395–399. Bibcode:2017Natur.552..395C. doi:10.1038/nature24679. PMID 29211712. S2CID 4471941.
- ^ Agnolin, Federico L .; Novas, Fernando E. (March 2011). "Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)?". Anais da Academia Brasileira de Ciências. 83 (1): 117–162. doi:10.1590/s0001-37652011000100008. PMID 21437379.
- ^ a b Longrich, Nicholas R .; Currie, Philip J. (31 March 2009). "A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 106 (13): 5002–5007. Bibcode:2009PNAS..106.5002L. doi:10.1073/pnas.0811664106. PMC 2664043. PMID 19289829.
- ^ Senter, Phil; Barsbold, R .; Britt, Brooks B.; Burnham, David B. (2004). "Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda)". Gunma Doğa Tarihi Müzesi Bülteni. 8: 1–20.
- ^ Sereno, P. C. 2005. Stem Archosauria—TaxonSearch Arşivlendi 2009-01-15 Wayback Makinesi, version 1.0, November 7, 2005
- ^ Turner, Alan H .; Makovicky, Peter J.; Norell, Mark A. (17 August 2012). "A Review of Dromaeosaurid Systematics and Paravian Phylogeny". Amerikan Doğa Tarihi Müzesi Bülteni. 371: 1–206. doi:10.1206/748.1. hdl:2246/6352. S2CID 83572446.
- ^ Adams, Dawn (1987). "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs". Currie'de, Philip J .; Koster, E. (eds.). Fourth symposium on mesozoic terrestrial ecosystems. Drumheller, Canada: Tyrrell Museum. s. 1–6.
- ^ Manning, Phillip L; Payne, David; Pennicott, John; Barrett, Paul M; Ennos, Roland A (11 October 2005). "Dinosaur killer claws or climbing crampons?". Biyoloji Mektupları. 2 (1): 110–112. doi:10.1098/rsbl.2005.0395. PMC 1617199. PMID 17148340.
- ^ a b Davies, S.J.J.F. (2002) "Ratites and Tinamous" Oxford University Press. New York, ABD[sayfa gerekli ]
- ^ Gilliard, Ernest Thomas (1958). Living birds of the world. Doubleday. OCLC 681791956.[sayfa gerekli ]
- ^ Kofron, Christopher P. (1999). "Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia". Zooloji Dergisi. 249 (4): 375–381. doi:10.1111 / j.1469-7998.1999.tb01206.x.
- ^ Kofron, Christopher P. (2003). "Queensland'deki güney kedinin saldırılarının vaka öyküleri". Queensland Müzesi Anıları. 49 (1): 335–338.
- ^ Redford, Kent H.; Peters, Gustav (1986). "Notes on the Biology and Song of the Red-Legged Seriema (Cariama cristata)". Alan Ornitoloji Dergisi. 57 (4): 261–269. JSTOR 4513154.
- ^ a b Manning, Phillip L.; Margetts, Lee; Johnson, Mark R.; Withers, Philip J.; Sellers, William I.; Falkingham, Peter L.; Mummery, Paul M.; Barrett, Paul M .; Raymont, David R. (September 2009). "Biomechanics of Dromaeosaurid Dinosaur Claws: Application of X-Ray Microtomography, Nanoindentation, and Finite Element Analysis". Anatomik Kayıt: Bütünleştirici Anatomi ve Evrimsel Biyolojideki Gelişmeler. 292 (9): 1397–1405. doi:10.1002 / ar.20986. PMID 19711472.
- ^ "Velociraptor's 'killing' claws were for climbing". Yeni Bilim Adamı. 203 (2725): 10. September 2009. doi:10.1016/s0262-4079(09)62381-2. Alındı 2009-09-15.
- ^ Senter, Phil (2009). "Pedal function in deinonychosaurs (Dinosauria: Theropoda): a comparative study" (PDF). Gunma Doğa Tarihi Müzesi Bülteni. 13: 1–14.
- ^ Fowler, Denver W .; Freedman, Elizabeth A .; Scannella, John B .; Kambic, Robert E. (14 December 2011). "The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds". PLOS ONE. 6 (12): e28964. Bibcode:2011PLoSO...628964F. doi:10.1371 / journal.pone.0028964. PMC 3237572. PMID 22194962.
- ^ Choi, Charles (14 December 2011). "Velociraptors' Killer Claws Helped Them Eat Prey Alive". LiveScience.
- ^ Gianechini, Federico A.; Ercoli, Marcos D.; Díaz‐Martínez, Ignacio (5 February 2020). "Differential locomotor and predatory strategies of Gondwanan and derived Laurasian dromaeosaurids (Dinosauria, Theropoda, Paraves): Inferences from morphometric and comparative anatomical studies". Anatomi Dergisi. 236 (5): 772–797. doi:10.1111/joa.13153. PMC 7163733. PMID 32023660.
- ^ Torices, A.; Wilkinson, R.; Arbor, V.M .; Ruiz-Omeñaca, J.I.; Currie, P.J. (2018). "Puncture-and-Pull Biomechanics in the Teeth of Predatory Coelurosaurian Dinosaurs". Güncel Biyoloji. 28 (9): 1467–1474. doi:10.1016/j.cub.2018.03.042. PMID 29706515.
- ^ Mudroch, A.; Richter, U.; Joger, U.; Kosma, R.; Idé, O.; Maga, A. (2011). "Didactyl tracks of paravian theropods (Maniraptora) from the ?Middle Jurassic of Africa". PLOS ONE. 6 (2): e1462. Bibcode:2011PLoSO...614642M. doi:10.1371/journal.pone.0014642. PMC 3038851. PMID 21339816. Şekil 4.
- ^ Maxwell, W. Desmond; Ostrom, John H. (27 December 1995). "Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations". Omurgalı Paleontoloji Dergisi. 15 (4): 707–712. doi:10.1080/02724634.1995.10011256.
- ^ Roach, Brian T.; Brinkman, Daniel L. (April 2007). "A Reevaluation of Cooperative Pack Hunting and Gregariousness in Deinonychus antirrhopus and Other Nonavian Theropod Dinosaurs". Peabody Doğa Tarihi Müzesi Bülteni. 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ^ Frederickson, J. A.; Engel, M. H.; Cifelli, R. L. (15 August 2020). "Ontogenetic dietary shifts in Deinonychus antirrhopus (Theropoda; Dromaeosauridae): Insights into the ecology and social behavior of raptorial dinosaurs through stable isotope analysis". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 552: 109780. Bibcode:2020PPP...552j9780F. doi:10.1016/j.palaeo.2020.109780.
- ^ Switek, Brian (7 January 2015). "Utah's Dinosaur 'Death Trap' Reveals Trove of Giant Predators". National Geographic Haberleri.
- ^ Norell, Mark A .; Clark, James M .; Turner, Alan H .; Makovicky, Peter J.; Barsbold, Rinchen; Rowe, Timothy (2006). "A New Dromaeosaurid Theropod from Ukhaa Tolgod (Ömnögov, Mongolia)". Amerikan Müzesi Novitates. 3545 (1): 1. doi:10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2. hdl:2246/5823.
- ^ Forster, C. A .; Sampson, SD; Chiappe, LM; Krause, DW (20 March 1998). "The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar". Bilim. 279 (5358): 1915–1919. Bibcode:1998Sci...279.1915F. doi:10.1126/science.279.5358.1915. PMID 9506938.
- ^ Chiappe, L.M. (2007-02-02). Glorified Dinosaurs: The Origin and Early Evolution of Birds. Sydney: UNSW Basın. ISBN 978-0-471-24723-4.[sayfa gerekli ]
- ^ Parsons, William L.; Parsons, Kristen M.; Dodson, Peter (15 April 2015). "Morphological Variations within the Ontogeny of Deinonychus antirrhopus (Theropoda, Dromaeosauridae)". PLOS ONE. 10 (4): e0121476. Bibcode:2015PLoSO..1021476P. doi:10.1371/journal.pone.0121476. PMC 4398413. PMID 25875499.
- ^ Chatterjee, Sankar; Templin, R. J. (2004). "Feathered Coelurosaurs from China: New Light on the Arboreal Origin of Avian Flight". Currie'de, Philip J .; Koppelhus, Eva B .; Shugar, Martin A.; Wright, Joanna L. (eds.). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Indiana University Press. pp. 251–281. ISBN 978-0-253-34373-4.
- ^ Naish, Darren (2012). Planet Dinosaur : The Next Generation of Killer Giants. Ateşböceği Kitapları. s.192. ISBN 978-1-77085-049-1.
- ^ Schmitz, L.; Motani, R. (14 April 2011). "Dinozorlarda Gece, Skleral Halka ve Yörünge Morfolojisinden Çıkarım". Bilim. 332 (6030): 705–708. Bibcode:2011Sci ... 332..705S. doi:10.1126 / science.1200043. PMID 21493820. S2CID 33253407.
- ^ Li, Quanguo; Gao, K.-Q.; Meng, Q .; Clarke, J. A.; Shawkey, M. D.; d'Alba, L.; Pei, R.; Ellison, M.; Norell, M. A .; Vinther, J. (9 March 2012). "Yeniden yapılanma Microraptor ve Yanardöner Tüylerin Evrimi ". Bilim. 335 (6073): 1215–1219. Bibcode:2012Sci ... 335.1215L. doi:10.1126 / science.1213780. PMID 22403389. S2CID 206537426.
- ^ Zelenitsky, Darla K; Therrien, François; Kobayashi, Yoshitsugu (28 October 2008). "Olfactory acuity in theropods: palaeobiological and evolutionary implications". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 276 (1657): 667–673. doi:10.1098/rspb.2008.1075. PMC 2660930. PMID 18957367.
- ^ Zelenitsky, Darla K.; Therrien, François; Ridgely, Ryan C.; McGee, Amanda R.; Witmer, Lawrence M. (13 April 2011). "Evolution of olfaction in non-avian theropod dinosaurs and birds". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 278 (1725): 3625–3634. doi:10.1098/rspb.2011.0238. PMC 3203493. PMID 21490022.
- ^ Rothschild, B.; Tanke, D. H .; Ford, T. L. (2001). "Theropod stress fractures and tendon avulsions as a clue to activity". Tanke, D. H .; Carpenter, K. (editörler). Mesozoik Omurgalı Yaşamı. Indiana University Press. s. 331–336.
- ^ Currie, Philip J .; Carabajal, Ariana Paulina (December 2012). "A New Specimen of Austroraptor cabazai Novas, Pol, Canale, Porfiri and Calvo, 2008 (Dinosauria, Theropoda, Unenlagiidae) from the Latest Cretaceous (Maastrichtian) of Río Negro, Argentina". Ameghiniana. 49 (4): 662–667. doi:10.5710/AMGH.30.8.2012.574. hdl:11336/9090. S2CID 129058582.
- ^ Grellet-Tinner, Gerald; Makovicky, Peter (1 June 2006). "A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications". Kanada Yer Bilimleri Dergisi. 43 (6): 705–719. Bibcode:2006CaJES..43..705G. doi:10.1139/E06-033.
- ^ Wiemann, Jasmina; Yang, Tzu-Ruei; Norell, Mark A. (31 October 2018). "Dinosaur egg colour had a single evolutionary origin". Doğa. 563 (7732): 555–558. Bibcode:2018Natur.563..555W. doi:10.1038/s41586-018-0646-5. PMID 30464264. S2CID 53188171.
- ^ "Dinosaur Egg Color Had a Single Evolutionary Origin". Kasım 2018.
- ^ "Dinosaurs put all colored bird eggs in one basket, evolutionarily speaking". 2018-10-31.
- ^ Bakker, Robert T. (1995). Raptor Red. New York: Bantam Books. s. 4. ISBN 978-0-553-57561-3.
- ^ Pérez-Moreno, B.P.; Sanz, J. L.; Sudre, J .; Sigé, B. (1994). "A theropod dinosaur from the Lower Cretaceous of southern France". Dinosaurs and Other Fossil Reptiles of Europe, Second Georges Cuvier Symposium, Montbéliard; Revue de Paléobiologie, Volume Spécial. 7: 173–188.
- ^ Currie, P. J. (1995). "New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda)". Omurgalı Paleontoloji Dergisi. 15 (3): 576–591. doi:10.1080/02724634.1995.10011250. (Öz Arşivlendi 2007-09-27 de Wayback Makinesi )
Dış bağlantılar
- The Dromaeosauridae: The Raptors!, from the University of California Berkeley Museum of Paleontology.
- Dinozorlar – Complete and free online edition of the book "Dinosaurs" as written by W. D. Matthew (cited in this article with authorship of the family Dromaeosauridae), and former Curator of Vertebrate Paleontology at the American Museum of Natural History in New York; Originally published in 1915
- Dromaeosauridae, Dinosaur-world reference with in-depth description and pictures of many dromaeosauridae dinosaurs
















