Uykunun sinirbilimi - Neuroscience of sleep

uyku nörobilim çalışmasıdır nörobilimsel ve fizyolojik doğasının temeli uyku ve işlevleri. Geleneksel olarak uyku, Psikoloji ve ilaç.[1] Nörobilim perspektifinden uyku çalışması, yirminci yüzyılın ikinci yarısından itibaren teknolojideki ilerlemeler ve nörobilim araştırmalarının yaygınlaşmasıyla öne çıktı.
Uykunun önemi, organizmaların her gün saatlerini uykuda geçirmeleriyle ve uyku eksikliği sonuçta ölüme yol açan feci etkilere neden olabilir.[2][3] Bu kadar önemli bir fenomen için, uykunun amaçları ve mekanizmaları yalnızca kısmen anlaşılmıştır, öyle ki 1990'ların sonlarında olduğu gibi.[4] şöyleydi: "Uykunun bilinen tek işlevi uykuyu iyileştirmektir".[5] Ancak, aşağıdaki gibi gelişmiş görüntüleme tekniklerinin geliştirilmesi EEG, EVCİL HAYVAN ve fMRI yüksek hesaplama gücüyle birlikte, uykunun altında yatan mekanizmaların giderek daha iyi anlaşılmasına yol açtı.
Nörobilimsel uyku çalışmasındaki temel sorular şunlardır:
- Uykunun bağıntıları nelerdir, yani organizmanın uyuduğunu doğrulayabilecek en küçük olaylar dizisi nelerdir?
- Uyku nasıl tetiklenir ve düzenlenir? beyin ve gergin sistem ?
- Uyku sırasında beyinde ne olur?
- Beyindeki fizyolojik değişikliklere dayanarak uyku işlevini nasıl anlayabiliriz?
- Çeşitli nedenler uyku bozuklukları ve nasıl tedavi edilebilirler?[6]
Modern sinirbilim uyku araştırmalarının diğer alanları şunları içerir: evrim uyku sırasında gelişme ve yaşlanma, hayvan uykusu uyuşturucunun uyku, rüyalar ve kabuslar üzerindeki etki mekanizması ve uyarılma uyku ve uyanıklık arasında.[7]
Giriş
Hızlı göz hareketi uykusu (REM), hızlı olmayan göz hareketi uykusu (NREM veya REM olmayan) ve uyanma, üç ana bilinç, sinirsel aktivite ve fizyolojik düzenlemeyi temsil eder.[8] NREM uykusunun kendisi birden çok aşamaya ayrılmıştır - N1, N2 ve N3. Uyku, 90 dakikalık REM ve NREM döngülerinde ilerler, sıra normalde N1 → N2 → N3 → N2 → REM'dir. İnsanlar uykuya daldıkça vücut aktivitesi yavaşlar. Vücut ısısı, kalp atış hızı, nefes alma hızı ve enerji kullanımı azalır. Beyin dalgaları yavaşlar ve büyür. Uyarıcı nörotransmiter asetilkolin beyinde daha az bulunur hale gelir.[9] İnsanlar genellikle termal olarak dost bir ortam yaratmak için manevra yaparlar - örneğin, soğuksa bir top şeklinde kıvrılırlar. Refleksler oldukça aktif kalır.
REM uykusu uyanıklığa daha yakın kabul edilir ve hızlı göz hareketi ve kas atonisi ile karakterizedir. NREM, derin uyku olarak kabul edilir (NREM'in en derin kısmına yavaş dalga uykusu ) ve belirgin göz hareketi eksikliği veya kas felci ile karakterizedir. Özellikle REM olmayan uyku sırasında beyin, uyku sırasında uyanmaya kıyasla önemli ölçüde daha az enerji kullanır. Aktivitenin azaldığı bölgelerde beyin, adenozin trifosfat (ATP), enerjinin kısa süreli depolanması ve taşınması için kullanılan molekül.[10] (Sessizce uyanırken beyin vücudun enerji kullanımının% 20'sinden sorumlu olduğu için, bu azalmanın genel enerji tüketimi üzerinde bağımsız olarak fark edilebilir bir etkisi vardır.)[11] Yavaş dalga uykusu sırasında, insanlar patlamaları salgılarlar. büyüme hormonu. Gün içinde bile tüm uyku, salgı ile ilişkilidir. prolaktin.[12]
Hobson & McCarley'e göre aktivasyon-sentez hipotezi, 1975–1977'de önerilen, REM ve non-REM arasındaki değişim, döngüsel, karşılıklı etkili nörotransmiter sistemleri açısından açıklanabilir.[13] Uyku zamanlaması, Sirkadiyen saat ve insanlarda, bir dereceye kadar iradeli davranışla. Sirkadiyen terimi Latince'den geliyor yaklaşık, "etrafında" (veya "yaklaşık olarak") anlamına gelir ve günlük veya ölür, "gün" anlamına gelir. Sirkadiyen saat, çoklu biyolojik süreçleri yöneten ve bunların yaklaşık 24 saatlik içsel, sürüklenebilir bir salınım sergilemelerine neden olan biyolojik bir mekanizmayı ifade eder. Bu ritimler bitkilerde, hayvanlarda, mantarlarda ve siyanobakterilerde yaygın olarak gözlemlenmiştir.
Uykunun bağıntıları
Uyku araştırmalarında önemli sorulardan biri, uyku durumunu açıkça tanımlamaktır. Bu sorun, uyku geleneksel olarak bir durum olarak tanımlandığı için ortaya çıkmaktadır. bilinç ve fizyolojik bir durum olarak değil,[14][15] bu nedenle, hangi minimum olaylar dizisinin uykuyu oluşturduğuna ve onu diğer kısmi ya da hiç bilinç durumlarından ayıran net bir tanım yoktu. Böyle bir tanım yapma problemi karmaşıktır çünkü farklı türlerde bulunan çeşitli uyku modlarını içermesi gerekir.
Semptomatik düzeyde uyku, tepkisizlik ile karakterizedir. duyusal girişler, düşük motor çıktı, bilinç azaldı farkındalık ve hızlı geri dönüşlü uyanıklık.[16] Ancak, bunları biyolojik bir tanıma dönüştürmek zordur çünkü beyindeki hiçbir yol uykunun oluşmasından ve düzenlenmesinden sorumlu değildir. İlk önerilerden biri, uykuyu, uykunun devre dışı bırakılması olarak tanımlamaktı. beyin zarı ve talamus[17] uyku sırasında duyusal girdilere neredeyse hiç cevap vermemesi nedeniyle. Ancak bu, uykunun bazı aşamalarında her iki bölge de aktif olduğu için geçersiz kılındı. Aslında, talamusun yalnızca duyusal bilgiyi kortekse iletme anlamında devre dışı bırakıldığı görülmektedir.[18]
Uyku ile ilgili diğer gözlemlerden bazıları, sempatik aktivite ve artış parasempatik aktivite non-REM uykusunda ve kalp atış hızı ve kan basıncında artışla birlikte homeostatik yanıt ve kas tonusu sırasında REM uykusu.[19][20] Bununla birlikte, bu semptomlar uyku durumlarıyla sınırlı değildir ve belirli fizyolojik tanımlarla eşleşmez.
Daha yakın zamanlarda, tanım problemi, genel beyin aktivitesinin karakteristik EEG modelleri şeklinde gözlemlenmesiyle ele alınmıştır.[21] Her biri uyku evresi ve uyanıklığın karakteristik bir modeli vardır. EEG uyku evresini belirlemek için kullanılabilir. Uyanma genellikle şu özelliklere sahiptir: beta (12–30 Hz) ve gama (25–100 Hz) huzurlu veya stresli bir aktivite olup olmamasına bağlı olarak.[22] Uyku başlangıcı, bu frekansın yavaşlayarak uyuşukluğa dönüşmesini içerir. alfa (8-12 Hz) ve son olarak teta (4-10 Hz) Aşama 1 NREM uykusu.[23] Bu frekans, NREM ve REM uykusunun daha yüksek aşamalarında giderek azalır. Öte yandan, uyanıklık sırasında uyku dalgalarının genliği en düşüktür (10-30μV) ve uykunun çeşitli aşamalarında giderek artan bir artış gösterir. Aşama 2 aşağıdakilerle karakterize edilir: uyku iğleri (sigma frekansında aralıklı dalga kümeleri, yani 12-14 Hz) ve K kompleksleri (keskin yukarı sapma ve ardından daha yavaş aşağı sapma). Evre 3 uykusunda daha fazla uyku mili vardır.[24][25][26] Aşama 3 ve 4 çok yüksek genliğe sahiptir delta dalgaları (0-4 Hz) ve yavaş dalga uykusu olarak bilinirler.[27] REM uykusu, düşük genlikli, karışık frekanslı dalgalarla karakterizedir. Genellikle bir testere dişi dalga paterni mevcuttur.[28]
Uyku bireyoluşu ve soyoluşu

Hayvanlar aleminde uykunun nasıl evrimleştiği ve insanlarda nasıl geliştiği soruları özellikle önemlidir çünkü bunlar sırasıyla uykunun işlevlerine ve mekanizmalarına ipucu verebilir.
Uyku evrimi
Farklı uyku düzenlerinin evrimi, bir dizi seçici basınçlar vücut büyüklüğü, göreceli metabolik hız, avlanma, besin kaynaklarının türü ve konumu ve bağışıklık işlevi dahil.[29][30][31][32] Uyku (özellikle derin SWS ve REM ) zor bir davranıştır çünkü hızla artar yırtıcılık risk. Bu, uykunun gelişmesi için uyku işlevlerinin içerdiği riske göre önemli bir avantaj sağlaması gerektiği anlamına gelir. Aslında, farklı organizmalarda uyku üzerinde çalışmak, onların kısmi uyku mekanizmaları geliştirerek veya koruyucu habitatlara sahip olarak bu riski nasıl dengelediklerini gösterir. Bu nedenle, uykunun evrimini incelemek, sadece gelişimsel yönler ve mekanizmalar hakkında değil, aynı zamanda bir uyarlanabilir uyku için gerekçe.
Uyku evrimini araştıran bir zorluk, yeterli uyku bilgisinin yalnızca iki hayvan grubu için bilinmesidir. kordata ve eklembacaklı.[7] Mevcut verilerle, uykunun nasıl geliştiğini belirlemek için karşılaştırmalı çalışmalar yapılmıştır. Bilim adamlarının bu araştırmalar aracılığıyla yanıtlamaya çalıştıkları bir soru, uykunun yalnızca bir kez mi yoksa birden çok kez mi evrimleştiği. Bunu anlamak için, evrimsel geçmişleri oldukça iyi bilinen farklı hayvan sınıflarındaki uyku modellerine bakarlar ve benzerliklerini ve farklılıklarını incelerler.
İnsanlar hem yavaş dalga hem de REM uykusuna sahiptirler, her iki aşamada her iki göz de kapalıdır ve her ikisi de yarım küreler dahil beyin. Uyku da kaydedildi memeliler insanlardan başka. Bir çalışma gösterdi ki ekidnalar sadece yavaş dalga uykusuna sahiptir (REM olmayan). Bu, REM uykusunun evrimde ancak sonra ortaya çıktığını gösteriyor gibi görünüyor. Therians.[33] Ancak bu daha sonra, ekidnada uykunun her iki modu da tek bir uyku durumunda birleştirdiğini iddia eden araştırmalarla itiraz edildi.[34] Diğer araştırmalar tuhaf bir uyku şekli göstermiştir. odontocetes (sevmek yunuslar ve domuzbalıkları ). Bu denir unihemispherical yavaş dalga uykusu (USWS). Bu uyku modu sırasında herhangi bir zamanda, bir beyin yarım küresinin EEG'si uykuyu gösterirken diğerininki uyanıklığa eşdeğerdir. Bazı durumlarda ilgili göz açıktır. Bu, hayvanın avcı riskini azaltmasına ve suda yüzerken uyumasına izin verebilir, ancak hayvan dinlenirken de uyuyabilir.[35][36]
Memelilerde bulunan uykunun bağıntıları, kuşlar aynı zamanda, yani kuş uykusu memelilere çok benzer ve hem SWS hem de REM uykusunu içerir, her iki gözün kapanması, kas tonusunun düşmesi vb.[37] Bununla birlikte, kanatlılarda REM uykusunun oranı çok daha düşüktür. Ayrıca, ortamda yüksek avlanma riski varsa, bazı kuşlar tek gözleri açık uyuyabilirler.[38][39] Bu, uçuş sırasında uyku olasılığını doğurur; Uykunun çok önemli olduğu ve bazı kuş türlerinin haftalarca sürekli uçabildiği düşünüldüğünde, bunun bariz sonuç olduğu görülmektedir. Ancak, uçuşta uyku kaydedilmedi ve şu ana kadar EEG verileri tarafından desteklenmiyor. Daha fazla araştırma, kuşların uçuş sırasında uyuyup uymadığını veya uyku yokken uzun uçuşlarda sağlıklı kalmalarını sağlayan başka mekanizmalar olup olmadığını açıklayabilir.
Kuşların aksine, çok az tutarlı uyku özelliği bulunmuştur. sürüngen Türler. Tek yaygın gözlem, sürüngenlerin REM uykusuna sahip olmamasıdır.[7]
Bazılarında uyu omurgasızlar ayrıca kapsamlı bir şekilde çalışılmıştır, örneğin, uyku meyve sinekleri (Drosophila)[40] ve bal arıları.[41] Bu hayvanlardaki bazı uyku mekanizmaları keşfedilirken, diğerleri oldukça belirsiz kalmıştır. Uykuyu tanımlayan özellikler çoğunlukla tanımlanmıştır ve memeliler gibi, bu, duyusal girdiye karşı azalmış tepkiyi, şu şekilde motor tepkisinin eksikliğini içerir. anten hareketsizlik vb.
Her iki uyku formunun da memelilerde ve kuşlarda bulunup sürüngenlerde bulunmaması (ki bunlar ara aşama olarak kabul edilir), uykunun her ikisinde de ayrı ayrı evrimleşmiş olabileceğini gösterir. Bunun doğrulanmasının ardından, uykunun EEG bağıntılarının işlevlerine dahil olup olmadığı veya sadece bir özellik olup olmadığı konusunda daha fazla araştırma yapılabilir. Bu, uykunun uzun vadeli esneklikte rolünü anlamada daha da yardımcı olabilir.
Tsoukalas'a (2012) göre, REM uykusu, iyi bilinen bir savunma mekanizmasının evrimsel bir dönüşümüdür. tonik hareketsizlik refleks. Hayvan hipnozu veya ölüm taklidi olarak da bilinen bu refleks, saldıran bir yırtıcıya karşı son savunma hattı olarak işlev görür ve hayvanın tamamen hareketsiz kalmasından oluşur: hayvan ölü görünür (çapraz başvuru "keseli sıçan oynamak"). Bu reaksiyonun nörofizyolojisi ve fenomenolojisi, derin bir evrimsel akrabalığa ihanet eden bir gerçek olan REM uykusuna çarpıcı benzerlikler göstermektedir. Örneğin, her iki reaksiyon da beyin sapı kontrolü, felç, sempatik aktivasyon ve ısı düzenleyici değişiklikler sergiler. Bu teori, önceki birçok bulguyu birleşik ve evrimsel olarak iyi bilgilendirilmiş bir çerçeveye entegre ediyor.[42][43]
Uyku gelişimi ve yaşlanma
Uyku ontogenisi, bir türün farklı yaş gruplarında, özellikle gelişme ve yaşlanma. Memeliler arasında bebekler en uzun süre uyur.[44] İnsan bebekler ortalama olarak 8 saat REM uykusu ve 8 saat NREM uykusuna sahiptir. Her uyku modunda harcanan zaman yüzdesi gelişimin ilk birkaç haftasında büyük ölçüde değişir ve bazı çalışmalar bunu çocuğun erken dönemlik derecesi ile ilişkilendirmiştir.[45] Doğum sonrası gelişimden sonraki birkaç ay içinde, REM uykusunda geçirilen saat yüzdesinde belirgin bir azalma olur. Çocuk yetişkin olduğunda, yaklaşık 6-7 saat NREM uykusunda ve yaklaşık bir saat REM uykusunda geçirir.[46][47] Bu sadece insanlar için değil, yiyecek için ebeveynlerine bağımlı birçok hayvan için de geçerlidir.[48] Gelişimin ilk aşamalarında REM uykusunun yüzdesinin çok yüksek olduğu gözlemi, REM uykusunun erken beyin gelişimini kolaylaştırabileceği hipotezine yol açtı.[45] Bununla birlikte, bu teori diğer çalışmalarla tartışılmıştır.
Uyku davranışı sırasında önemli değişikliklere uğrar. Gençlik. Bu değişikliklerin bazıları insanlarda toplumsal olabilir, ancak diğer değişiklikler hormonaldir. Bir diğer önemli değişiklik, çocukluk dönemine göre uyku saatlerinin azalması ve yavaş yavaş bir yetişkinle özdeşleşmesidir. Ayrıca speküle ediliyor ki homeostatik düzenleme mekanizmalar ergenlik döneminde değişebilir. Bunun dışında ergenlerin değişen rutinlerinin biliş ve dikkat gibi diğer davranışlar üzerindeki etkisi henüz araştırılmamıştır.[49][50] Örneğin Ohayon ve arkadaşları, çocukluktan ergenliğe kadar toplam uyku süresindeki düşüşün biyolojik özellikten çok çevresel faktörlerle ilişkili göründüğünü belirtmişlerdir.[51]
Yetişkinlikte uyku mimarisi, uyku gecikmesinin ve NREM 1. ve 2. aşamalarında geçirilen sürenin yaşla artabileceğini, REM ve SWS uykusunda harcanan sürenin azaldığını gösteriyor.[51] Bu değişiklikler sıklıkla beyin atrofisi, bilişsel bozukluk ve yaşlılıkta nörodejeneratif bozukluklarla ilişkilendirilmiştir.[51][52][53][54][55][56] Örneğin, Backhaus ve arkadaşları, orta yaştaki bildirimsel bellek konsolidasyonundaki düşüşün (deneylerinde: 48 ila 55 yaş arası), daha düşük miktarda SWS'ye bağlı olduğunu ve bunun 30 yaş civarında azalmaya başlayabileceğini belirtmişlerdir. .[52] Mander ve arkadaşlarına göre, medial prefrontal kortekste (mPFC) gri cevherdeki atrofi, NREM uykusu sırasında yavaş aktivitede, yaşlı erişkinlerde hafıza konsolidasyonunu bozabilecek bozulmanın bir göstergesidir.[53] Ve gündüz aşırı uykululuk ve gece gibi uyku bozuklukları uykusuzluk hastalığı, sıklıkla ilerleyici fonksiyonel bozukluğun faktör riski olarak anılmıştır. Alzheimer hastalığı (AD) veya Parkinson hastalığı (PD).[54][56]
Bu nedenle, yaşlanmada uyku aynı derecede önemli bir başka araştırma alanıdır. Yaygın bir gözlem, birçok yaşlı yetişkinin uykuya dalamama ve uyku verimliliğinde belirgin bir düşüş yaşayarak uyku başladıktan sonra yatakta uyanık zaman geçirmesidir.[57] Ayrıca bazı değişiklikler olabilir sirkadiyen ritimler.[58] Bu değişikliklere neyin sebep olduğu ve yaşlı yetişkinlerin rahat uykusunu sağlamak için bunların nasıl azaltılabileceği konusunda çalışmalar devam etmektedir.
Uyku sırasında beyin aktivitesi


Uyku sırasında beynin farklı bölümlerinin aktivitesini anlamak, uykunun işlevleri hakkında bir ipucu verebilir. Zihinsel aktivitenin, beynin farklı bölgelerinden de olsa, uykunun tüm aşamalarında mevcut olduğu gözlemlenmiştir. Bu yüzden, popüler anlayışın aksine beyin uyku sırasında asla tamamen kapanmaz. Ayrıca belirli bir bölgenin uyku yoğunluğu homeostatik olarak Uyumadan önce ilgili aktivite miktarı ile ilgili.[59][60] PET ve fMRI gibi görüntüleme yöntemlerinin EEG kayıtları ile birlikte kullanılması, hangi beyin bölgelerinin karakteristik dalga sinyallerini oluşturmaya katıldığına ve işlevlerinin neler olabileceğine dair bir ipucu veriyor.
Aşama modelinin tarihsel gelişimi
Uykunun aşamaları ilk olarak 1937'de Alfred Lee Loomis ve farklı olanları ayıran iş arkadaşları elektroensefalografi Uyanıklıktan derin uykuya kadar olan spektrumu temsil eden beş seviyeye (A'dan E'ye) uykunun (EEG) özellikleri.[61] 1953'te, REM uykusunun farklı olduğu keşfedildi ve böylece William C. Dement ve Nathaniel Kleitman uykuyu dört NREM aşaması ve REM olarak yeniden sınıflandırdı.[21] Evreleme kriterleri 1968'de Allan Rechtschaffen ve "R&K uyku puanlama el kitabında" Anthony Kales.[62][63]
R&K standardında NREM uykusu, 3. ve 4. aşamaları içeren yavaş dalga uykusu ile dört aşamaya bölündü. 3. aşamada, delta dalgaları toplam dalga modellerinin% 50'sinden daha azını oluştururken, bunlar% 50'den fazlasını oluşturuyordu. 4. aşamada ayrıca, REM uykusu bazen 5. aşama olarak adlandırılıyordu. 2004'te AASM, R&K puanlama sistemini gözden geçirmek için AASM Görsel Puanlama Görev Gücü'nü görevlendirdi. İnceleme birkaç değişiklikle sonuçlandı, en önemlisi Aşama 3 ve 4'ün Aşama N3 ile birleşimiydi. Gözden geçirilmiş puanlama 2007 yılında şu şekilde yayınlandı: Uyku ve İlişkili Olayların Puanlanması için AASM Kılavuzu.[64] Uyarılma, solunum, kalp ve hareket olayları da eklendi.[65][66]
NREM uyku aktivitesi
NREM uykusu azalmış küresel ve bölgesel serebral kan akışı. Yetişkin insanlarda tüm uykunun ~% 80'ini oluşturur.[67] Başlangıçta, beyin sapı uyarılmayla ilişkilendirilen etkisiz olacaktır, ancak bunun daha sonra PET çalışmalarının düşük çözünürlüğünden kaynaklandığı bulundu ve beyin sapında da bir miktar yavaş dalga aktivitesi olduğu gösterildi. Bununla birlikte, beynin diğer bölümleri de dahil olmak üzere Precuneus, bazal önbeyin ve Bazal ganglion uyku sırasında devre dışı bırakılır. Korteksin birçok bölgesi de hareketsizdir, ancak farklı seviyelerdedir. Örneğin, ventromedial prefrontal korteks en az aktif alan olarak kabul edilirken birincil korteks, en az devre dışı bırakılan.[25][68]
NREM uykusu yavaş salınımlarla karakterizedir, iğ ve delta dalgaları. Beynin diğer bölümlerindeki lezyonlar onları etkilemediği, ancak korteksteki lezyonlar etkilediği için, yavaş salınımların korteksten geldiği gösterilmiştir.[69] Delta dalgalarının karşılıklı olarak bağlanmış talamik ve kortikal sinir devreleri tarafından oluşturulduğu gösterilmiştir. Uyku sırasında talamus, beyne duyusal bilgi aktarmayı durdurur, ancak kortikal projeksiyonlarına gönderilen sinyaller üretmeye devam eder. Bu dalgalar talamusta korteksin yokluğunda bile üretilir, ancak kortikal çıktının büyük nöron grupları tarafından eşzamanlı ateşlemede rol oynadığı görülmektedir.[70] talamik retiküler çekirdek uyku iğlerinin pacemaker'ı olarak kabul edilir. Bu, talamusun ritmik uyarılmasının sekonder artışın artmasına yol açmasıyla daha da doğrulanmıştır. depolarizasyon kortikal nöronlarda, bu da ateşleme genliğinin artmasına neden olarak kendi kendine devam eden aktiviteye neden olur. Uyku iğlerinin, korteksi duyusal girdiden ayırmada ve kalsiyum iyonlarının hücrelere girmesine izin vermede rol oynadığı ve böylece potansiyel olarak bir rol oynadığı tahmin edilmektedir. Plastisite.[71][72]
NREM 1
NREM Aşama 1 (N1 - hafif uyku, uyku hali, uykulu uyku - yetişkinlerde toplam uykunun% 5-10'u): Bu, genellikle uyku ve uyanıklık arasında ortaya çıkan ve bazen daha derin uyku dönemleri ile REM dönemleri arasında ortaya çıkan bir uyku aşamasıdır. Kaslar aktiftir ve gözler yavaşça yuvarlanır, orta derecede açılıp kapanır. Beyin, alfa dalgaları 8-13 sıklıktaHz (uyanık durumda yaygın) teta dalgaları 4–7 Hz frekansa sahip. Ani seğirmeler ve hipnik gerizekalı pozitif olarak da bilinir miyoklonus, N1 sırasında uykunun başlamasıyla ilişkili olabilir. Bazı insanlar da deneyimleyebilir hipnogojik halüsinasyonlar bu aşamada. Non-REM1 sırasında, insanlar bazılarını kaybeder. kas tonusu ve dış çevrenin en bilinçli farkındalığı.
NREM 2
NREM Evre 2 (N2 - yetişkinlerde toplam uykunun% 45-55'i[73]): Bu aşamada, teta aktivitesi gözlemlenir ve uyuyanların uyanması giderek zorlaşır; alfa dalgaları önceki aşamanın, adı verilen ani faaliyetle kesintiye uğraması uyku iğleri (veya talamokortikal iğler) ve K kompleksleri.[74] Uyku milleri 11 ile 16 Hz arasındadır (en yaygın olarak 12-14 Hz). Bu aşamada, EMG ile ölçülen kas aktivitesi azalır ve dış çevreye ilişkin bilinçli farkındalık kaybolur.
NREM 3
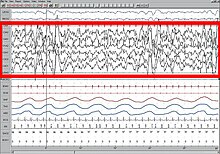
NREM Evre 3 (N3 - yetişkinlerde toplam uykunun% 15–25'i): Eskiden 3. ve 4. aşamalara ayrılmış olan bu aşama yavaş dalga uykusu (SWS) veya derin uyku. SWS, preoptik alan ve oluşur delta aktivitesi 3,5 Hz'den düşük yüksek genlikli dalgalar. Uyuyan çevreye daha az duyarlıdır; birçok çevresel uyarı artık herhangi bir tepki üretmez. Yavaş dalga uykusunun, öznel uykululuk duygularını en çok rahatlatan ve vücudu geri yükleyen en dinlendirici uyku hali olduğu düşünülmektedir.[75]
Bu aşama, minimum% 20 varlığı ile karakterize edilir. delta dalgaları 0,5–2 Hz aralığında değişen ve tepeden tepeye amplitüdü> 75 μV olan. (EEG standartları, delta dalgalarını 0 ila 4 Hz olarak tanımlar, ancak her iki orijinal R&K modelinde uyku standartları (Allan Rechtschaffen ve "R&K uyku puanlama kılavuzundaki" Anthony Kales.),[62][63] Yeni 2007 AASM kılavuzlarının yanı sıra 0,5–2 Hz aralığı vardır.) Bu, aşağıdaki gibi parasomnilerin görüldüğü aşamadır. gece terörü, yatak ıslatma, uyurgezerlik, ve somniloquy meydana gelir. Birçok çizim ve açıklama hala% 20-50 delta dalgaları olan bir evre N3'ü ve% 50'den fazla delta dalgaları olan bir aşama N4'ü göstermektedir; bunlar aşama N3 olarak birleştirilmiştir.[73]
REM uyku aktivitesi

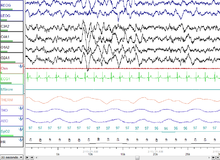
REM Aşaması (REM Uykusu - yetişkinlerde toplam uykunun% 20-25'i[76]): REM uykusu, çoğu kasın felç olduğu ve kalp atış hızı, solunum ve vücut ısısının düzensiz hale geldiği yerdir. REM uykusu şu şekilde açılır: asetilkolin salgılama ve dahil olmak üzere monoaminleri salgılayan nöronlar tarafından engellenir serotonin. REM ayrıca paradoksal uyku çünkü uyuyan kişi, uyanma durumuna benzer yüksek frekanslı EEG dalgaları sergilemesine rağmen, uyandırılması diğer herhangi bir uyku evresinden daha zordur.[74] Hayati belirtiler uyarılmayı ve beyin tarafından oksijen tüketiminin uyuyan kişinin uyanık olduğu zamandan daha yüksek olduğunu gösterir.[77] REM uykusu, uyanıklıkla karşılaştırılabilen yüksek küresel beyin kan akışı ile karakterizedir.[78] Aslında, korteksteki pek çok alanın, REM uykusu sırasında uyanıklıktan bile daha fazla kan akışına sahip olduğu kaydedilmiştir. hipokamp, geçici -oksipital alanlar, korteksin bazı kısımları ve bazal önbeyin. limbik ve paralimbik sistem dahil amigdala REM uykusu sırasında diğer aktif bölgelerdir.[78][79] REM uykusu sırasındaki beyin aktivitesi uyanıklığa çok benziyor gibi görünse de, REM ve uyanıklık arasındaki temel fark şudur: uyarılma REM'de daha etkili bir şekilde engellenir. Bu, sanal sessizliğiyle birlikte monoaminerjik Beyindeki nöronların REM'i karakterize ettiği söylenebilir.[80]
Yeni doğmuş bir bebek günde 8-9 saatini sadece REM uykusunda geçirir. Beş yaş civarında, KEP'de sadece iki saatten biraz fazla zaman harcanır.[81] REM uykusunun işlevi belirsizdir, ancak eksikliği karmaşık görevleri öğrenme yeteneğini bozar. Kas kaynaklı fonksiyonel felç atonia REM'de, bu aşamada meydana gelen genellikle canlı rüyalardan sahneleri fiziksel olarak canlandırarak organizmaları kendine zarar vermekten korumak gerekli olabilir.
EEG kayıtlarında, REM uykusu yüksek frekans, düşük genlikli aktivite ve kendiliğinden meydana gelen beta ve gama dalgaları. Bu hızlı frekans dalgalarının oluşumu için en iyi adaylar, kortikotalamik devrelerde hızlı ritmik patlama yapan nöronlardır. Yavaş dalga uykusundan farklı olarak, hızlı frekans ritimleri, talamokortikal ve neokortikal alanlar arasındaki belirli yerel devrelerde sınırlı alanlar üzerinde senkronize edilir. Bunların oluşturduğu söyleniyor kolinerjik beyin sapı yapılarından süreçler.
Bunun dışında amigdala, REM uykusunun modülasyonunda rol oynar ve REM uykusunun dahili bilgi işlemeye izin verdiği hipotezini destekler. Yüksek amigdalar aktivitesi ayrıca rüyalar sırasında duygusal tepkilere neden olabilir.[82] Benzer şekilde, rüyaların tuhaflığı, azalan aktiviteden kaynaklanıyor olabilir. prefrontal bilginin entegre edilmesine dahil olan bölgeler Bölümsel hafıza.
Ponto-genikülo-oksipital dalgalar
REM uykusu aynı zamanda ateşleme ile de ilgilidir. ponto-genikülo-oksipital dalgalar (fazik aktivite veya PGO dalgaları olarak da adlandırılır) ve kolinerjik yükselen uyarılma sistemindeki aktivite. PGO dalgaları, yanal genikülat çekirdek ve oksipital korteks REM öncesi dönemde ve rüya içeriğini temsil ettiği düşünülmektedir. LG kortikal kanaldaki daha yüksek sinyal-gürültü oranı, rüyalardaki görsel görüntülerin REM uykusunun tam olarak gelişmesinden önce ortaya çıkabileceğini göstermektedir, ancak bu henüz doğrulanmamıştır. PGO dalgaları ayrıca beynin gelişimi ve yapısal olgunlaşmasında rol oynayabilir. uzun vadeli güçlendirme olgunlaşmamış hayvanlarda, gelişimsel beyinde uyku sırasında yüksek PGO aktivitesi olduğu gerçeğine dayanmaktadır.[83][84]
Ağ yeniden etkinleştirme
Uyku sırasındaki diğer aktivite şekli yeniden aktivasyondur. Bazı elektrofizyolojik çalışmalar, uykudan önce bir öğrenme görevi sırasında bulunan nöronal aktivite modellerinin uyku sırasında beyinde yeniden etkinleştirildiğini göstermiştir.[85] Bu, aktif alanların hafızadan sorumlu alanlarla çakışmasıyla birlikte, uykunun bazı hafıza konsolidasyon işlevlerine sahip olabileceği teorisine yol açtı. Bu ilişkide, bazı çalışmalar, sıralı bir motor görevinden sonra, motor öncesi ve görsel korteks ilgili alanlar en çok REM uykusu sırasında aktiftir, ancak NREM sırasında değildir. Benzer şekilde, hipokampal Uzamsal öğrenme görevlerinde yer alan alanlar NREM uykusunda yeniden etkinleştirilir, ancak REM'de etkinleştirilmez. Bu tür çalışmalar, belirli bellek türlerinin sağlamlaştırılmasında uykunun bir rolüne işaret ediyor. Bununla birlikte, diğer bellek türlerinin de bu mekanizmalarla konsolide edilip edilmediği hala belirsizdir.[86]
Hipokampal neokortikal diyalog
Hipokampal neokortikal diyalog, sıradaki çok yapılandırılmış etkileşimleri ifade eder. SWS topluluk olarak adlandırılan nöron grupları arasında hipokamp ve neokorteks.[87][88] Keskin dalga paternleri (SPW), SWS sırasında hipokampa hakimdir ve hipokampustaki nöron popülasyonları bu aşamada organize patlamalara katılır. Bu, korteksteki (AŞAĞI / YUKARI durumu) durum değişiklikleri ile eşzamanlı olarak yapılır ve korteksteki yavaş salınımlar ile koordine edilir. Bu gözlemler, hipokampusun kısa ve orta vadeli hafızada rol oynadığı, korteksin ise uzun süreli hafızada rol oynadığı bilgisiyle birleştiğinde, hipokampal neokortikal diyaloğun hipokampusun bilgi aktarımını sağlayan bir mekanizma olabileceği hipotezine yol açmıştır. kortekse. Bu nedenle, hipokampal neokortikal diyaloğun hafıza konsolidasyonunda rol oynadığı söyleniyor.[89]
Uyku düzenleme
Uyku düzenlemesi bir organizmanın ne zaman uyku ile uyanıklık arasında geçiş yaptığının kontrolünü ifade eder.[90] Buradaki temel sorular, beynin hangi bölümlerinin uyku başlangıcına dahil olduğunu ve bunların etki mekanizmalarının ne olduğunu belirlemektir.[91] İnsanlarda ve çoğu hayvanda uyku ve uyanıklık bir elektronik takla model, yani her iki durum da kararlıdır, ancak ara durumlar değildir.[92][93] Elbette, flip-flop'tan farklı olarak, uyku durumunda, uyanma dakikasından uzaklaşan bir zamanlayıcı var gibi görünüyor, böylece belirli bir süre sonra kişi uyumak zorunda kalıyor ve böyle bir durumda uyanmak bile dengesiz bir duruma dönüşüyor. . Bunun tersi de daha az ölçüde doğru olabilir.
Uyku başlangıcı
Uykudaki lezyonların ortaya çıkmasıyla uykuya başlayan mekanizmalara biraz ışık tutuldu. preoptik alan ve anterior hipotalamus yol açmak uykusuzluk hastalığı posterior hipotalamusta olanlar ise uykululuğa neden olur.[94][95] Bu, daha da daraltıldı ve merkezi orta beyin tegmentum kortikal aktivasyonda rol oynayan bölgedir. Bu nedenle, uyku başlangıcı, ön hipotalamusun aktivasyonunun yanı sıra arka bölgelerin ve merkezi orta beyin tegmentumunun inhibisyonundan kaynaklanıyor gibi görünmektedir. Daha fazla araştırma, hipotalamik bölgenin adı verildiğini göstermiştir. ventrolateral preoptik çekirdek inhibe edici nörotransmitteri üretir GABA uyku başlangıcı sırasında uyarılma sistemini engeller.[92]
Uyku düzenleme modelleri
Uyku iki paralel mekanizma ile düzenlenir, homeostatik düzenleme ve sirkadiyen düzenleme hipotalamus tarafından kontrol edilir ve üst kiyazmatik çekirdek (SCN), sırasıyla. Uyku sürüşünün kesin doğası bilinmemekle birlikte, uyanıklık sırasında homeostatik basınç oluşur ve bu kişi uyuyana kadar devam eder. Adenozin Bunda kritik bir rol oynadığı düşünülmektedir ve birçok kişi, basınç artışının kısmen adenozin birikiminden kaynaklandığını öne sürmüştür. Ancak bazı araştırmacılar, tek başına birikimin bu fenomeni tam olarak açıklamadığını göstermiştir. Sirkadiyen ritim, çevresel ipuçlarının yokluğunda bile devam ettiği gösterilen vücutta 24 saatlik bir döngüdür. Bu, SCN'den beyin sapına olan projeksiyonlardan kaynaklanır.
Bu iki süreç modeli ilk olarak 1982'de Borbely tarafından önerildi,[96] onlara sırasıyla Süreç S (homeostatik) ve Süreç C (Sirkadiyen) adını veren. Yavaş dalga yoğunluğunun gece boyunca nasıl arttığını ve günün başlangıcında sirkadiyen ritim sinüzoid gibi iken nasıl düştüğünü gösterdi. İkisi arasındaki fark en yüksek olduğunda uyku basıncının maksimum olduğunu öne sürdü.
1993'te rakip süreç modeli adı verilen farklı bir model[97] önerildi. Bu model, Borbely'nin modeline karşı, bu iki sürecin uyku üretmek için birbirine zıt olduğunu açıkladı. Bu modele göre sirkadiyen ritimde yer alan SCN, uyanıklığı artırır ve homeostatik ritme karşı çıkar. Karşıt olarak, hipotalamusta bir anahtar gibi davranan ve uyarılma sistemini kapatan karmaşık bir multisinaptik yolla düzenlenen homeostatik ritim vardır. Her iki etki birlikte, uyku ve uyanıklığın testere benzeri bir etkisi yaratır.[80] Daha yakın zamanlarda, her iki modelin de kendileri için bir miktar geçerliliği olduğu öne sürülürken, yeni teoriler NREM uykusunun REM tarafından engellenmesinin de bir rol oynayabileceğini savunuyor.[98] Her durumda, iki süreç mekanizması basit sirkadiyen ritime esneklik katar ve uyarlanabilir bir ölçü olarak gelişebilirdi.
Talamik düzenleme
Uykudaki beyin aktivitesinin çoğu, talamus ve talamusun SWS'de kritik bir rol oynayabileceği görülmektedir. İki ana salınım yavaş dalga uykusu, delta ve yavaş salınım, hem talamus hem de korteks tarafından oluşturulabilir. Bununla birlikte, uyku iğleri yalnızca talamus tarafından üretilebilir ve bu da rolünü çok önemli hale getirir. Talamik pacemaker hipotezi[99] bu salınımların talamus tarafından üretildiğini, ancak aynı anda ateşlenen birkaç talamik nöron grubunun senkronizasyonunun, talamus ile talamik etkileşime bağlı olduğunu kabul eder. korteks. Talamus ayrıca, tonik moddan fazik moda değiştiğinde uyku başlangıcında kritik bir rol oynar, böylece hem merkezi hem de merkezi olmayan öğeler için bir ayna görevi görür ve aktivitelerini koordine etmek için korteksin uzak kısımlarını birbirine bağlar.[100][101][102]
Yükselen retiküler aktivasyon sistemi
artan retiküler aktivasyon sistemi bir dizi nöral alt sistemden oluşur. talamik çekirdek ve bir dizi dopaminerjik, noradrenerjik, serotonerjik, histaminerjik, kolinerjik, ve glutamaterjik beyin çekirdekleri.[103][104][105][106] Uyanık olduğunda, her türlü spesifik olmayan duyusal bilgiyi alır ve bunları kortekse iletir. Aynı zamanda savaş veya kaç tepkilerini de modüle eder ve bu nedenle motor sistemiyle bağlantılıdır. Uyku başlangıcı sırasında, iki yolla hareket eder: talamus yoluyla kortekse uzanan bir kolinerjik yol ve hipotalamus yoluyla kortekse uzanan bir dizi monoaminerjik yol. NREM uykusu sırasında bu sistem, GABAerjik nöronlar tarafından engellenir. ventrolateral preoptik alan ve parafasiyal bölge farklı beyin bölgelerindeki diğer uykuyu teşvik eden nöronların yanı sıra.
Uyku fonksiyonu
Uyku ihtiyacı ve işlevi uyku araştırmalarında en az anlaşılan alanlar arasındadır. 50 yıllık bir araştırmadan sonra, insanların uyku nedenleri hakkında ne bildiği sorulduğunda, William C. Dement, kurucusu Stanford Üniversitesi 's Sleep Research Center, answered, "As far as I know, the only reason we need to sleep that is really, really solid is because we get sleepy."[107] It is likely that sleep evolved to fulfill some primeval function and took on multiple functions over time[108] (analogous to the gırtlak, which controls the passage of food and air, but indi over time to develop speech capabilities).
The multiple hypotheses proposed to explain the function of sleep reflect the incomplete understanding of the subject. While some functions of sleep are known, others have been proposed but not completely substantiated or understood. Some of the early ideas about sleep function were based on the fact that most (if not all) external activity is stopped during sleep. Initially, it was thought that sleep was simply a mechanism for the body to "take a break" and reduce wear. Later observations of the low metabolik rates in the brain during sleep seemed to indicate some metabolic functions of sleep.[109] This theory is not fully adequate as sleep only decreases metabolism by about 5–10%.[110][111] With the development of EEG, it was found that the brain has almost continuous internal activity during sleep, leading to the idea that the function could be that of reorganization or specification of neuronal circuits or strengthening of connections.[112][113] These hypotheses are still being explored. Other proposed functions of sleep include- maintaining hormonal balance, temperature regulation and maintaining heart rate.
According to a recent sleep disruption and insomnia review study[114], there are short-term and long-term negative consequences on healthy individuals. The short term consequences include increased stress responsivity and psychosocial issues such as impaired cognitive or academic performance and depression. Experiments indicated that, in healthy children and adults, episodes of fragmented sleep or insomnia increased sympathetic activation, which can disrupt mood and cognition. The long term consequences include metabolic issues such as glucose homeostasis disruption and even tumor formation and increased risks of cancer.
Koruma
The "Preservation and Protection" theory holds that sleep serves an adaptive function. It protects the animal during that portion of the 24-hour day in which being awake, and hence roaming around, would place the individual at greatest risk.[115] Organisms do not require 24 hours to feed themselves and meet other necessities. From this perspective of adaptation, organisms are safer by staying out of harm's way, where potentially they could be prey to other, stronger organisms. They sleep at times that maximize their safety, given their physical capacities and their habitats.
This theory fails to explain why the brain disengages from the external environment during normal sleep. However, the brain consumes a large proportion of the body's energy at any one time and preservation of energy could only occur by limiting its sensory inputs. Another argument against the theory is that sleep is not simply a passive consequence of removing the animal from the environment, but is a "drive"; animals alter their behaviors in order to obtain sleep.
Therefore, circadian regulation is more than sufficient to explain periods of activity and quiescence that are adaptive to an organism, but the more peculiar specializations of sleep probably serve different and unknown functions. Moreover, the preservation theory needs to explain why carnivores like lions, which are on top of the besin zinciri and thus have little to fear, sleep the most. It has been suggested that they need to minimize energy expenditure when not hunting.
Waste clearance from the brain
During sleep, metabolik atık gibi ürünler immünoglobulinler, protein fragments or intact proteins like beta-amiloid, may be cleared from the interstitium aracılığıyla glifatik sistem nın-nin lenf -like channels coursing along perivascular spaces ve astrosit network of the brain.[116][117][118] According to this model, hollow tubes between the blood vessels and astrocytes act like a savak allowing drainage of Beyin omurilik sıvısı carrying wastes out of the brain into systemic blood.[117][118] Such mechanisms, which remain under preliminary research as of 2017, indicate potential ways in which sleep is a regulated maintenance period for brain bağışıklık functions and clearance of beta-amyloid, a risk faktörü için Alzheimer hastalığı.[117]
Restorasyon
Yara iyileşmesi has been shown to be affected by sleep.[119]
It has been shown that sleep deprivation affects the bağışıklık sistemi.[120] It is now possible to state that "sleep loss impairs immune function and immune challenge alters sleep," and it has been suggested that sleep increases white blood cell counts.[121] A 2014 study found that depriving mice of sleep increased cancer growth and dampened the immune system's ability to control cancers.[122]
The effect of sleep duration on somatik growth is not completely known. One study recorded growth, height, and weight, as correlated to parent-reported time in bed in 305 children over a period of nine years (age 1–10). It was found that "the variation of sleep duration among children does not seem to have an effect on growth."[123] It is well established that slow-wave sleep affects büyüme hormonu levels in adult men.[12] During eight hours' sleep, Van Cauter, Leproult, and Plat found that the men with a high percentage of SWS (average 24%) also had high growth hormone secretion, while subjects with a low percentage of SWS (average 9%) had low growth hormone secretion.[46]
There is some supporting evidence of the restorative function of sleep. The sleeping brain has been shown to remove metabolic waste products at a faster rate than during an awake state.[124] While awake, metabolism generates reactive oxygen species, which are damaging to cells. In sleep, metabolic rates decrease and reactive oxygen species generation is reduced allowing restorative processes to take over. It is theorized that sleep helps facilitate the synthesis of molecules that help repair and protect the brain from these harmful elements generated during waking.[125] The metabolic phase during sleep is anabolic; anabolic hormones such as growth hormones (as mentioned above) are secreted preferentially during sleep.
Energy conservation could as well have been accomplished by resting quiescent without shutting off the organism from the environment, potentially a dangerous situation. A sedentary nonsleeping animal is more likely to survive predators, while still preserving energy. Sleep, therefore, seems to serve another purpose, or other purposes, than simply conserving energy. Another potential purpose for sleep could be to restore signal strength in synapses that are activated while awake to a "baseline" level, weakening unnecessary connections that to better facilitate learning and memory functions again the next day; this means the brain is forgetting some of the things we learn each day.[126]
Endokrin işlevi
The secretion of many hormonlar is affected by sleep-wake cycles. Örneğin, melatonin, a hormonal timekeeper, is considered a strongly sirkadiyen hormone, whose secretion increases at dim light and peaks during nocturnal sleep, diminishing with bright light to the eyes.[127] In some organisms melatonin secretion depends on sleep, but in humans it is independent of sleep and depends only on light level. Of course, in humans as well as other animals, such a hormone may facilitate coordination of sleep onset. Benzer şekilde, kortizol ve tiroid uyarıcı hormon (TSH) are strongly circadian and diurnal hormones, mostly independent of sleep.[128] In contrast, other hormones like growth hormone (GH) & prolaktin are critically sleep-dependent, and are suppressed in the absence of sleep.[129] GH has maximum increase during SWS while prolactin is secreted early after sleep onset and rises through the night. In some hormones whose secretion is controlled by light level, sleep seems to increase secretion. Almost in all cases, sleep deprivation has detrimental effects. For example, cortisol, which is essential for metabolism (it is so important that animals can die within a week of its deficiency) and affects the ability to withstand noxious stimuli, is increased by waking and during REM sleep.[130] Similarly, TSH increases during nocturnal sleep and decreases with prolonged periods of reduced sleep, but increases during total acute sleep deprivation.[131][132]
Because hormones play a major role in energy balance and metabolism, and sleep plays a critical role in the timing and amplitude of their secretion, sleep has a sizable effect on metabolism. This could explain some of the early theories of sleep function that predicted that sleep has a metabolic regulation role.
Memory processing
According to Plihal & Born, sleep generally increases recalling of previous learning and experiences, and its benefit depends on the phase of sleep and the type of memory.[133] For example, studies based on declarative and procedural memory tasks applied over early and late nocturnal sleep, as well as wakefulness controlled conditions, have been shown that declarative memory improves more during early sleep (dominated by SWS) while procedural memory during late sleep (dominated by REM sleep).[134][135]
Regarding to declarative memory, the functional role of SWS has been associated with hippocampal replays of previously encoded neural patterns that seem to facilitate long-term memories consolidation.[134][135] This assumption is based on the active system consolidation hypothesis, which states that repeated reactivations of newly encoded information in hippocampus during slow oscillations in NREM sleep mediate the stabilization and gradually integration of declarative memory with pre-existing knowledge networks on the cortical level.[136] It assumes the hippocampus might hold information only temporarily and in fast-learning rate, whereas the neocortex is related to long-term storage and slow-learning rate.[134][135][137][138][139] This dialogue between hippocampus and neocortex occurs in parallel with hippocampal sharp-wave ripples ve thalamo-cortical spindles, synchrony that drives the formation of spindle-ripple event which seems to be a prerequisite for the formation of long-term memories.[135][137][139][140]
Reactivation of memory also occurs during wakefulness and its function is associated with serving to update the reactivated memory with new encoded information, whereas reactivations during SWS are presented as crucial for memory stabilization.[135] Based on targeted memory reactivation (TMR) experiments that use associated memory cues to triggering memory traces during sleep, several studies have been reassuring the importance of nocturnal reactivations for the formation of persistent memories in neocortical networks, as well as highlighting the possibility of increasing people’s memory performance at declarative recalls.[134][140][138][139][141]
Furthermore, nocturnal reactivation seems to share the same neural oscillatory patterns as reactivation during wakefulness, processes which might be coordinated by teta aktivitesi.[142] During wakefulness, theta oscillations have been often related to successful performance in memory tasks, and cued memory reactivations during sleep have been showing that theta activity is significantly stronger in subsequent recognition of cued stimuli as compared to uncued ones, possibly indicating a strengthening of memory traces and lexical integration by cuing during sleep.[143] However, the beneficial effect of TMR for memory consolidation seems to occur only if the cued memories can be related to prior knowledge.[144]
Other studies have been also looking at the specific effects of different stages of sleep on different types of memory. For example, it has been found that sleep deprivation does not significantly affect recognition of faces, but can produce a significant impairment of temporal memory (discriminating which face belonged to which set shown). Sleep deprivation was also found to increase beliefs of being correct, especially if they were wrong. Another study reported that the performance on free recall of a list of nouns is significantly worse when sleep deprived (an average of 2.8 ± 2 words) compared to having a normal night of sleep (4.7 ± 4 words). These results reinforce the role of sleep on Bildirimsel bellek oluşumu. This has been further confirmed by observations of low metabolic activity in the Prefrontal korteks ve geçici ve parietal loblar for the temporal learning and verbal learning tasks respectively. Data analysis has also shown that the neural assemblies during SWS correlated significantly more with templates than during waking hours or REM sleep. Also, post-learning, post-SWS reverberations lasted 48 hours, much longer than the duration of novel object learning (1 hour), indicating uzun vadeli güçlendirme.
Moreover, observations include the importance of uyuklayan: improved performance in some kinds of tasks after a 1-hour afternoon nap; studies of performance of shift workers, showing that an equal number of hours of sleep in the day is not the same as in the night. Current research studies look at the molecular and physiological basis of bellek konsolidasyonu during sleep. These, along with studies of genes that may play a role in this phenomenon, together promise to give a more complete picture of the role of sleep in memory.
Renormalizing the synaptic strength
Sleep can also serve to weaken synaptic connections that were acquired over the course of the day but which are not essential to optimal functioning. In doing so, the resource demands can be lessened, since the upkeep and strengthening of synaptic connections constitutes a large portion of energy consumption by the brain and tax other cellular mechanisms such as protein sentezi for new channels.[126][145] Without a mechanism like this taking place during sleep, the metabolic needs of the brain would increase over repeated exposure to daily synaptic strengthening, up to a point where the strains become excessive or untenable.
Behavior change with sleep deprivation
One approach to understanding the role of sleep is to study the deprivation of it.[146] Uyku eksikliği is common and sometimes even necessary in modern societies because of occupational and domestic reasons like round-the-clock service, security or media coverage, cross-time-zone projects etc. This makes understanding the effects of sleep deprivation very important.
Many studies have been done from the early 1900s to document the effect of sleep deprivation. The study of REM deprivation began with William C. Dement more than fifty years ago. He conducted a sleep and dream research project on eight subjects, all male. For a span of up to 7 days, he deprived the participants of REM sleep by waking them each time they started to enter the stage. He monitored this with small electrodes attached to their scalp and temples. As the study went on, he noticed that the more he deprived the men of REM sleep, the more often he had to wake them. Afterwards, they showed more REM sleep than usual, later named REM rebound.[147][148]
neurobehavioral basis for these has been studied only recently. Sleep deprivation has been strongly correlated with increased probability of accidents and industrial errors.[149][150] Many studies have shown the slowing of metabolic activity in the brain with many hours of uyku borcu.[131] Some studies have also shown that the Dikkat network in the brain is particularly affected by lack of sleep,[151] and though some of the effects on attention may be masked by alternate activities (like standing or walking) or caffeine consumption,[152] attention deficit cannot be completely avoided.
Sleep deprivation has been shown to have a detrimental effect on cognitive tasks, especially involving divergent functions or multitasking.[149][153][154] It also has effects on mood and emotion, and there have been multiple reports of increased tendency for rage, fear or depression with sleep debt.[155][156] However, some of the higher cognitive functions seem to remain unaffected albeit slower.[153] Many of these effects vary from person to person[157] i.e. while some individuals have high degrees of cognitive impairment with lack of sleep, in others, it has minimal effects. The exact mechanisms for the above are still unknown and the exact neural pathways and cellular mechanisms of sleep debt are still being researched.
Uyku bozuklukları
A sleep disorder, or somnipathy, is a medical disorder of the sleep patterns of a person or animal. Polisomnografi is a test commonly used for diagnosing some sleep disorders. Uyku bozuklukları genel olarak şöyle sınıflandırılır: dyssomnias, parasomniler, sirkadiyen ritim uyku bozuklukları (CRSD), and other disorders including ones caused by medical or psychological conditions and uyku hastalığı. Some common sleep disorders include uykusuzluk hastalığı (chronic inability to sleep), uyku apnesi (abnormally low breathing during sleep), narkolepsi (excessive sleepiness at inappropriate times), katapleksi (sudden and transient loss of muscle tone), and sleeping sickness (disruption of sleep cycle due to infection). Other disorders that are being studied include uyurgezerlik, sleep terror ve bed wetting.
Studying sleep disorders is particularly useful as it gives some clues as to which parts of the brain may be involved in the modified function. This is done by comparing the imaging and histological patterns in normal and affected subjects. Treatment of sleep disorders typically involves behavioral and psikoterapötik methods though other techniques may also be used. The choice of treatment methodology for a specific patient depends on the patient's diagnosis, medical and psychiatric history, and preferences, as well as the expertise of the treating clinician. Often, behavioral or psychotherapeutic and farmakolojik approaches are compatible and can effectively be combined to maximize therapeutic benefits.
Frequently, sleep disorders have been also associated with neurodegenerative diseases, mainly when they are characterized by abnormal accumulation of alfa-sinüklein, gibi çoklu sistem atrofisi (MSA), Parkinson hastalığı (PD) ve Lewy vücut hastalığı (LBD).[158][54][159][56][160][161][162] Örneğin, Parkinson hastalığı teşhisi konan kişiler genellikle farklı türde uyku kaygıları sergilediler. uykusuzluk hastalığı (PD popülasyonunun yaklaşık% 70'i), hipersomnia (PD popülasyonunun% 50'sinden fazlası) ve REM uyku davranış bozukluğu (RBD) - PH popülasyonunun yaklaşık% 40'ını etkileyebilir ve artan motor semptomlarla ilişkilidir.[158][54][159][56][160][162] Furthermore, RBD has been also highlighted as a strong precursor of future development of those neurodegenerative diseases over several years in prior, which seems to be a great opportunity for improving treatments.[54][56]
Uyku bozuklukları da gözlenmiştir. Alzheimer hastalığı (AD), nüfusunun yaklaşık% 45'ini etkiler.[54][56][161] Dahası, bakıcı raporlarına göre bu yüzde daha da yüksektir, yaklaşık% 70.[163] PD popülasyonunda olduğu gibi, uykusuzluk hastalığı ve hipersomnia are frequently recognized in AD patients, which are associated with accumulation of Beta-amiloid, sirkadiyen ritim uyku bozuklukları (CRSD) ve melatonin değişiklik.[54][56][163] Ek olarak, değişiklikler uyku mimarisi AD'de de görülmektedir.[54][56][161] Yaşlanma ile uyku yapısı doğal olarak değişiyor gibi görünse de, AD hastalarında ağırlaşmaktadır. SWS potansiyel olarak azalır (bazen tamamen yok), iğler ve REM uykusunda geçirilen süre de azalırken gecikme süresi artar.[163] AD'de zayıf uyku başlangıcı aynı zamanda rüya ile ilişkili halüsinasyon, artan huzursuzluk, gezinme ve ajitasyon ile ilişkilendirilmiştir. gün batımı - hastalıkta sunulan tipik bir kronobiyolojik fenomen.[56][163]
The neurodegenerative conditions are commonly related to brain structures impairments, which might disrupt the states of sleep and wakefulness, circadian rhythm, motor or non motor functioning.[54][56] Öte yandan, uyku bozuklukları sıklıkla hastanın bilişsel işlevlerinin, duygusal durumunun ve yaşam kalitesinin kötüleşmesi ile ilgilidir.[56][162][163] Dahası, bu anormal davranışsal semptomlar, yakınlarının ve bakıcılarının bunaltılmasına olumsuz katkıda bulunur.[56][162][163] Bu nedenle, uyku bozuklukları ve nörodejeneratif hastalıklar arasındaki ilişkinin daha derinlemesine anlaşılması, esas olarak onunla ilgili sınırlı araştırma ve artan yaşam beklentisi dikkate alındığında son derece önemli görünmektedir.[54][163]
A related field is that of uyku ilacı which involves the diagnosis and therapy of sleep disorders and sleep deprivation, which is a major cause of accidents. This involves a variety of diagnostic methods including polysomnography, uyku günlüğü, çoklu uyku gecikme testi, etc. Similarly, treatment may be behavioral such as bilişsel davranışçı terapi or may include pharmacological medication or bright light therapy.
Rüya görmek

Dreams are successions of images, ideas, emotions, and sensations that occur involuntarily in the mind during certain stages of sleep (mainly the REM stage). The content and purpose of dreams are not yet clearly understood though various theories have been proposed. Rüyaların bilimsel çalışmasına denir tekiroloji.
There are many theories about the neurological basis of dreaming. Bu şunları içerir: activation synthesis theory —the theory that dreams result from brain stem activation during REM sleep; the continual activation theory—the theory that dreaming is a result of activation and synthesis but dreams and REM sleep are controlled by different structures in the brain; and dreams as excitations of long term memory—a theory which claims that long term memory excitations are prevalent during waking hours as well but are usually controlled and become apparent only during sleep.
There are multiple theories about dream function as well. Some studies claim that dreams strengthen semantic memories. This is based on the role of hippocampal neocortical dialog and general connections between sleep and memory. One study surmises that dreams erase junk data in the brain. Emotional adaptation ve mood regulation are other proposed functions of dreaming.
Bir evrimsel standpoint, dreams might simulate and rehearse threatening events, that were common in the organism's ancestral environment, hence increasing a persons ability to tackle everyday problems and challenges in the present. For this reason these threatening events may have been passed on in the form of genetic memories.[164][165] This theory accords well with the claim that REM sleep is an evolutionary transformation of a well-known defensive mechanism, the tonic immobility reflex.[42][43]
Most theories of dream function appear to be conflicting, but it is possible that many short-term dream functions could act together to achieve a bigger long-term function.[7] It may be noted that evidence for none of these theories is entirely conclusive.
The incorporation of waking memory events into dreams is another area of active research and some researchers have tried to link it to the declarative memory consolidation functions of dreaming.[166][167]
A related area of research is the neuroscience basis of kabuslar. Many studies have confirmed a high prevalence of nightmares and some have correlated them with high stres seviyeleri.[168] Multiple models of nightmare production have been proposed including neo-Freudyen models as well as other models such as image contextualization model, boundary thickness model, threat simulation model etc.[169] Nörotransmiter imbalance has been proposed as a cause of nightmares, as also affective network dysfunction- a model which claims that nightmare is a product of dysfunction of circuitry normally involved in dreaming.[170][171] As with dreaming, none of the models have yielded conclusive results and studies continue about these questions.
Ayrıca bakınız
Referanslar
- ^ "A brief history of sleep research".
- ^ "NCBI Sleep Guide". Arşivlenen orijinal on 2007-01-10.
- ^ Cirelli C, Shaw PJ, Rechtschaffen A, Tononi G (September 1999). "No evidence of brain cell degeneration after long-term sleep deprivation in rats". Beyin Araştırması. 840 (1–2): 184–93. doi:10.1016/s0006-8993(99)01768-0. PMID 10517970. S2CID 592724.
- ^ Stickgold R, Valker MP (22 May 2010). The Neuroscience of Sleep. s. xiii. ISBN 9780123757227. Alındı 18 Temmuz 2015.
- ^ Konnikova M (8 July 2015). "The Work We Do While We Sleep". The New Yorker. Alındı 17 Temmuz 2015.
The Harvard sleep researcher Robert Stickgold has recalled his former collaborator J. Allan Hobson joking that the only known function of sleep is to cure sleepiness.
- ^ Kilduff TS, Lein ES, de la Iglesia H, Sakurai T, Fu YH, Shaw P (November 2008). "New developments in sleep research: molecular genetics, gene expression, and systems neurobiology". Nörobilim Dergisi. 28 (46): 11814–8. doi:10.1523/JNEUROSCI.3768-08.2008. PMC 2628168. PMID 19005045.
- ^ a b c d Stickgold, Robert (2009). The Neuroscience of Sleep. Amsterdam: Boston : Academic Press/Elsevier. pp.61 –86. ISBN 9780123750730.
- ^ J. Alan Hobson, Edward F. Pace-Scott, & Robert Stickgold (2000), "Dreaming and the brain: Toward a cognitive neuroscience of conscious states", Davranış ve Beyin Bilimleri 23.
- ^ Kahverengi, pp. 1100–1102.
- ^ Kahverengi, pp. 1118–1119: "Compared with wakefulness, sleep reduces brain energy demands, as suggested by the 44% reduction in the cerebral metabolic rate (CMR) of glucose (791) and a 25% reduction in the CMR of O2 (774) during sleep."
- ^ Siegel JM (April 2008). "Do all animals sleep?". Sinirbilimlerindeki Eğilimler. 31 (4): 208–13. doi:10.1016/j.tins.2008.02.001. PMID 18328577. S2CID 6614359.
- ^ a b Eve Van Cauter & Karine Spiegel (1999). "Circadian and Sleep Control of Hormonal Secretions", in Turek & Zee (eds.), Regulation of Sleep and Circadian Rhythms, pp. 397–425.
- ^ Hobson JA, McCarley RW (December 1977). "The brain as a dream state generator: an activation-synthesis hypothesis of the dream process". Amerikan Psikiyatri Dergisi. 134 (12): 1335–48. doi:10.1176 / ajp.134.12.1335. PMID 21570.
- ^ "Online Free Medical Dictionary".
- ^ Anch, A. Michael (1988). Sleep : a scientific perspective. Englewood Kayalıkları, NJ: Prentice Hall. ISBN 9780138129187.
- ^ Thorpy MJ, Yager J (2001). The encyclopedia of sleep and sleep disorders (2. baskı). New York: Dosyadaki Gerçekler. ISBN 978-0-8160-4089-6.
- ^ Magnin M, Rey M, Bastuji H, Guillemant P, Mauguière F, Garcia-Larrea L (February 2010). "Thalamic deactivation at sleep onset precedes that of the cerebral cortex in humans". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 107 (8): 3829–33. Bibcode:2010PNAS..107.3829M. doi:10.1073/pnas.0909710107. PMC 2840430. PMID 20142493.
- ^ Jones EG (March 2009). "Synchrony in the interconnected circuitry of the thalamus and cerebral cortex". New York Bilimler Akademisi Yıllıkları. 1157 (1): 10–23. Bibcode:2009NYASA1157...10J. doi:10.1111/j.1749-6632.2009.04534.x. PMID 19351352. S2CID 19140386.
- ^ Somers VK, Dyken ME, Mark AL, Abboud FM (February 1993). "Sympathetic-nerve activity during sleep in normal subjects". New England Tıp Dergisi. 328 (5): 303–7. doi:10.1056/NEJM199302043280502. PMID 8419815.
- ^ Tononi G, Cirelli C (February 2006). "Sleep function and synaptic homeostasis". Uyku Tıbbı Yorumları. 10 (1): 49–62. doi:10.1016/j.smrv.2005.05.002. PMID 16376591.
- ^ a b Dement W, Kleitman N (November 1957). "Cyclic variations in EEG during sleep and their relation to eye movements, body motility, and dreaming". Elektroensefalografi ve Klinik Nörofizyoloji. 9 (4): 673–90. doi:10.1016/0013-4694(57)90088-3. PMID 13480240.
- ^ Hughes JR (July 2008). "Gamma, fast, and ultrafast waves of the brain: their relationships with epilepsy and behavior". Epilepsi ve Davranış. 13 (1): 25–31. doi:10.1016/j.yebeh.2008.01.011. PMID 18439878. S2CID 19484309.
- ^ Barker W, Burgwin S (Nov–Dec 1948). "Brain wave patterns accompanying changes in sleep and wakefulness during hypnosis". Psikosomatik Tıp. 10 (6): 317–26. doi:10.1097/00006842-194811000-00002. PMID 18106841. S2CID 31249127.
- ^ Jankel WR, Niedermeyer E (January 1985). "Sleep spindles". Klinik Nörofizyoloji Dergisi. 2 (1): 1–35. doi:10.1097/00004691-198501000-00001. PMID 3932462. S2CID 44570089.
- ^ a b Hofle N, Paus T, Reutens D, Fiset P, Gotman J, Evans AC, Jones BE (June 1997). "Regional cerebral blood flow changes as a function of delta and spindle activity during slow wave sleep in humans". Nörobilim Dergisi. 17 (12): 4800–8. doi:10.1523/JNEUROSCI.17-12-04800.1997. PMC 6573353. PMID 9169538.
- ^ Oswald I, Taylor AM, Treisman M (September 1960). "Discriminative responses to stimulation during human sleep". Beyin. 83 (3): 440–53. doi:10.1093/brain/83.3.440. PMID 13731563.
- ^ Loomis AL, Harvey EN, Hobart GA (1938). "Distribution of disturbance-patterns in the human electroencephalogram with special reference to sleep". Nörofizyoloji Dergisi. 1 (5): 413–430. doi:10.1152/jn.1938.1.5.413.
- ^ Dumermuth G, Walz W, Scollo-Lavizzari G, Kleiner B (1 January 1972). "Spectral analysis of EEG activity in different sleep stages in normal adults". Avrupa Nörolojisi. 7 (5): 265–96. doi:10.1159/000114432. PMID 4339034.
- ^ McNamara, P., R. A. Barton, and C. L. Nunn. 2010, Evolution of sleep: Phylogenetic and functional perspectives. Cambridge University Press, Cambridge.
- ^ Capellini I, Nunn CL, McNamara P, Preston BT, Barton RA (October 2008). "Yırtıcı değil, enerjik kısıtlamalar memelilerde uyku düzeninin evrimini etkiler". Fonksiyonel Ekoloji. 22 (5): 847–853. doi:10.1111 / j.1365-2435.2008.01449.x. PMC 2860325. PMID 20428321.
- ^ Acerbi A, McNamara P, Nunn CL (May 2008). "To sleep or not to sleep: the ecology of sleep in artificial organisms". BMC Ekolojisi. 8: 10. doi:10.1186/1472-6785-8-10. PMC 2396600. PMID 18479523.
- ^ Preston, B. T., I. Capellini, P. McNamara, R. A. Barton, and C. L. Nunn. 2009. Parasite resistance and the adaptive significance of sleep. Bmc Evolutionary Biology 9.
- ^ Allison T, Van Twyver H, Goff WR (July 1972). "Electrophysiological studies of the echidna, Tachyglossus aculeatus. I. Waking and sleep". Archives Italiennes de Biologie. 110 (2): 145–84. PMID 4342268.
- ^ Siegel JM, Manger PR, Nienhuis R, Fahringer HM, Pettigrew JD (May 1996). "The echidna Tachyglossus aculeatus combines REM and non-REM aspects in a single sleep state: implications for the evolution of sleep". Nörobilim Dergisi. 16 (10): 3500–6. doi:10.1523/JNEUROSCI.16-10-03500.1996. PMC 6579141. PMID 8627382.
- ^ Mukhametov, Lev M. (1987). "Unihemispheric slow-wave sleep in the Amazonian dolphin, Inia geoffrensis". Sinirbilim Mektupları. 79 (1–2): 128–132. doi:10.1016/0304-3940(87)90684-7. PMID 3670722. S2CID 30316737.
- ^ Lyamin OI, Mukhametov LM, Siegel JM, Nazarenko EA, Polyakova IG, Shpak OV (February 2002). "Unihemispheric slow wave sleep and the state of the eyes in a white whale". Davranışsal Beyin Araştırması. 129 (1–2): 125–9. doi:10.1016/S0166-4328(01)00346-1. PMID 11809503. S2CID 27009685.
- ^ Roth TC, Lesku JA, Amlaner CJ, Lima SL (December 2006). "A phylogenetic analysis of the correlates of sleep in birds". Uyku Araştırmaları Dergisi. 15 (4): 395–402. doi:10.1111/j.1365-2869.2006.00559.x. PMID 17118096.
- ^ Mascetti GG, Bobbo D, Rugger M, Vallortigara G (August 2004). "Monocular sleep in male domestic chicks". Davranışsal Beyin Araştırması. 153 (2): 447–52. doi:10.1016/j.bbr.2003.12.022. PMID 15265641. S2CID 19793594.
- ^ Rattenborg NC, Lima SL, Amlaner CJ (November 1999). "Facultative control of avian unihemispheric sleep under the risk of predation". Davranışsal Beyin Araştırması. 105 (2): 163–72. doi:10.1016/S0166-4328(99)00070-4. PMID 10563490. S2CID 8570743.
- ^ Shaw, P. J. (2000). "Correlates of Sleep and Waking in Drosophila melanogaster". Bilim. 287 (5459): 1834–1837. Bibcode:2000Sci...287.1834S. doi:10.1126/science.287.5459.1834. PMID 10710313.
- ^ Sauer S, Kinkelin M, Herrmann E, Kaiser W (August 2003). "The dynamics of sleep-like behaviour in honey bees". Karşılaştırmalı Fizyoloji Dergisi A. 189 (8): 599–607. doi:10.1007/s00359-003-0436-9. PMID 12861424. S2CID 13603649.
- ^ a b Tsoukalas, Ioannis (2012). "The origin of REM sleep: A hypothesis". Rüya görmek. 22 (4): 253–283. doi:10.1037/a0030790.
- ^ a b Vitelli, R. (2013). Exploring the Mystery of REM Sleep. Bugün Psikoloji, On-line blog, 25 March
- ^ Parmelee AH, Wenner WH, Schulz HR (1 October 1964). "Infant sleep patterns: From birth to 16 weeks of age". Pediatri Dergisi. 65 (4): 576–582. doi:10.1016/S0022-3476(64)80291-2. PMID 14216645.
- ^ a b Gertner S, Greenbaum CW, Sadeh A, Dolfin Z, Sirota L, Ben-Nun Y (July 2002). "Sleep-wake patterns in preterm infants and 6 month's home environment: implications for early cognitive development". Erken İnsan Gelişimi. 68 (2): 93–102. doi:10.1016/S0378-3782(02)00018-X. PMID 12113995.
- ^ a b Van Cauter E, Leproult R, Plat L (August 2000). "Age-related changes in slow wave sleep and REM sleep and relationship with growth hormone and cortisol levels in healthy men". JAMA. 284 (7): 861–8. doi:10.1001/jama.284.7.861. PMID 10938176.
- ^ Roffwarg HP, Muzio JN, Dement WC (April 1966). "Ontogenetic development of the human sleep-dream cycle". Bilim. 152 (3722): 604–19. Bibcode:1966Sci...152..604R. doi:10.1126/science.152.3722.604. PMID 17779492.
- ^ Ibuka, Nobuo (1984). "Ontogenesis of circadian sleep-wakefulness rhythms and developmental changes of sleep in the altricial rat and in the precocial guinea pig". Davranışsal Beyin Araştırması. 11 (3): 185–196. doi:10.1016/0166-4328(84)90210-9. PMID 6721913. S2CID 11458572.
- ^ Carskadon MA, Wolfson AR, Acebo C, Tzischinsky O, Seifer R (December 1998). "Adolescent sleep patterns, circadian timing, and sleepiness at a transition to early school days". Uyku. 21 (8): 871–81. doi:10.1093 / uyku / 21.8.871. PMID 9871949.
- ^ Wolfson AR, Carskadon MA (August 1998). "Sleep schedules and daytime functioning in adolescents". Çocuk Gelişimi. 69 (4): 875–87. doi:10.1111 / j.1467-8624.1998.tb06149.x. PMID 9768476.
- ^ a b c Ohayon M, Carskadon M, Guilleminault C, Vitiello M (2004). "Meta-analysis of quantitative sleep parameters from childhood to old age in healthy individuals: Developing normative sleep values across the human lifespan". Uyku. 27 (7): 1255–73. doi:10.1093/sleep/27.7.1255. PMID 15586779.
- ^ a b Backhaus J, Born J, Hoeckesfeld R, Fokuhl S, Hohagen F, Junghanns K (2007). "Midlife Decline in Declarative Memory Consolidation Is Correlated with a Decline in Slow Wave Sleep". Öğrenme ve Hafıza. 14 (5): 336–341. doi:10.1101/lm.470507. PMC 1876757. PMID 17522024.
- ^ a b Mander, B., Rao, V., Lu, B., Saletin, J., Lindquist, J., Ancoli-Israel, S., . . . Walker, M. (2013). Prefrontal atrophy, disrupted NREM slow waves and impaired hippocampal-dependent memory in aging. 357 - 364.
- ^ a b c d e f g h ben j Zhong N, Rogers, Lewis (2011). "Sleep–wake disturbances in common neurodegenerative diseases: A closer look at selected aspects of the neural circuitry". Nörolojik Bilimler Dergisi. 307 (1–2): 9–14. doi:10.1016/j.jns.2011.04.020. PMID 21570695. S2CID 44744844.
- ^ Cordi M, Schlarb A, Rasch B (2014). "Deepening sleep by hypnotic suggestion". Uyku. 37 (6): 1143–52. doi:10.5665/sleep.3778. PMC 4015388. PMID 24882909.
- ^ a b c d e f g h ben j k l Malkani R, Attarian H (2015). "Sleep in Neurodegenerative Disorders". Current Sleep Medicine Reports. 1 (2): 81–90. doi:10.1007/s40675-015-0016-x.
- ^ Weitzman ED, Czeisler CA, Coleman RM, Spielman AJ, Zimmerman JC, Dement W, Richardson G, Pollak CP (July 1981). "Delayed sleep phase syndrome. A chronobiological disorder with sleep-onset insomnia". Genel Psikiyatri Arşivleri. 38 (7): 737–46. doi:10.1001/archpsyc.1981.01780320017001. PMID 7247637.
- ^ Myers BL, Badia P (1995). "Changes in circadian rhythms and sleep quality with aging: mechanisms and interventions". Nörobilim ve Biyodavranışsal İncelemeler. 19 (4): 553–71. doi:10.1016/0149-7634(95)00018-6. PMID 8684716. S2CID 23651761.
- ^ Borbély AA (1 January 1998). "Processes underlying sleep regulation" (PDF). Hormon Araştırması. 49 (3–4): 114–7. doi:10.1159/000023156. PMID 9550110. S2CID 15651050.
- ^ Mazoyer B, Houdé O, Joliot M, Mellet E, Tzourio-Mazoyer N (September 2009). "Regional cerebral blood flow increases during wakeful rest following cognitive training". Beyin Araştırmaları Bülteni. 80 (3): 133–8. doi:10.1016/j.brainresbull.2009.06.021. PMID 19589374. S2CID 22696285.
- ^ Loomis AL, Harvey EN, Hobart GA (1937). "III Cerebral states during sleep, as studied by human brain potentials". J. Exp. Psychol. 21 (2): 127–44. doi:10.1037/h0057431.
- ^ a b Kahverengi, pp. 1108–1109.
- ^ a b Rechtschaffen A, Kales A, eds. (1968). A Manual of Standardized Terminology, Techniques and Scoring System for Sleep Stages of Human Subjects (PDF). Washington: Public Health Service, US Government Printing Office.
- ^ Iber C, Ancoli-Israel S, Chesson A, Quan SF, American Academy of Sleep Medicine (2007). The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications. Westchester: American Academy of Sleep Medicine.
- ^ "Stages of Sleep" (PDF). Psychology World. 1998. Alındı 15 Haziran 2008.
(includes illustrations of "sleep spindles" and "K-complexes")
- ^ Schulz H (April 2008). "Rethinking sleep analysis". Klinik Uyku Tıbbı Dergisi. 4 (2): 99–103. doi:10.5664/jcsm.27124. PMC 2335403. PMID 18468306.
- ^ Parmeggiani (2011), Systemic Homeostasis and Poikilostasis in Sleep, Passim.
- ^ Braun, A. (1997). "Regional cerebral blood flow throughout the sleep-wake cycle. An H2(15)O PET study". Beyin. 120 (7): 1173–1197. doi:10.1093/brain/120.7.1173. PMID 9236630.
- ^ McGinty DJ, Sterman MB (June 1968). "Sleep suppression after basal forebrain lesions in the cat". Bilim. 160 (3833): 1253–5. Bibcode:1968Sci...160.1253M. doi:10.1126/science.160.3833.1253. PMID 5689683. S2CID 24677928.
- ^ Hutt, Axel, ed. (2011-07-12). Sleep and anesthesia : neural correlates in theory and experiment. New York: Springer. ISBN 9781461401728.
- ^ Steriade, M (1997). "Synchronized activities of coupled oscillators in the cerebral cortex and thalamus at different levels of vigilance [published erratum appears in Cereb Cortex 1997 Dec;7(8):779]". Beyin zarı. 7 (6): 583–604. doi:10.1093/cercor/7.6.583. PMID 9276182.
- ^ Contreras D, Destexhe A, Sejnowski TJ, Steriade M (February 1997). "Spatiotemporal patterns of spindle oscillations in cortex and thalamus". Nörobilim Dergisi. 17 (3): 1179–96. doi:10.1523/JNEUROSCI.17-03-01179.1997. PMC 6573181. PMID 8994070.
- ^ a b Fuller PM, Gooley JJ, Saper CB (December 2006). "Neurobiology of the sleep-wake cycle: sleep architecture, circadian regulation, and regulatory feedback". Biyolojik Ritimler Dergisi. 21 (6): 482–93. doi:10.1177/0748730406294627. PMID 17107938. S2CID 36572447.
- ^ a b Daniel L S, Gilbert DT, Wegner DM (2009). Psikoloji. Worth Yayıncıları. ISBN 978-1-4292-0615-0.
- ^ Waterhouse J, Fukuda Y, Morita T (March 2012). "Daily rhythms of the sleep-wake cycle". Journal of Physiological Anthropology. 31: 5. doi:10.1186/1880-6805-31-5. PMC 3375033. PMID 22738268.
- ^ Myers DG (22 September 2003). Psychology, Seventh Edition, in Modules (High School Version). Macmillan. s. 268–. ISBN 978-0-7167-8595-8. Alındı 22 Ağustos 2012.
- ^ Saladin, Kenneth S. (2012). Anatomy and Physiology: The Unity of Form and Function, 6th Edition. McGraw-Hill. s. 537. ISBN 978-0-07-337825-1.
- ^ a b Reivich M, Isaacs G, Evarts E, Kety S (April 1968). "The effect of slow wave sleep and REM sleep on regional cerebral blood flow in cats". Nörokimya Dergisi. 15 (4): 301–6. doi:10.1111/j.1471-4159.1968.tb11614.x. PMID 5641651. S2CID 27688214.
- ^ Maquet P, Péters J, Aerts J, Delfiore G, Degueldre C, Luxen A, Franck G (September 1996). "Functional neuroanatomy of human rapid-eye-movement sleep and dreaming" (PDF). Doğa. 383 (6596): 163–6. Bibcode:1996Natur.383..163M. doi:10.1038/383163a0. PMID 8774879. S2CID 19628239. Arşivlenen orijinal (PDF) 2010-07-05 tarihinde. Alındı 2012-07-28.
- ^ a b Saper CB, Scammell TE, Lu J (October 2005). "Hypothalamic regulation of sleep and circadian rhythms". Doğa. 437 (7063): 1257–63. Bibcode:2005Natur.437.1257S. doi:10.1038 / nature04284. PMID 16251950. S2CID 1793658.
- ^ Siegel, Jerome M (1999). "Uyku". Encarta Ansiklopedisi. Microsoft. Arşivlenen orijinal 14 Aralık 2007'de. Alındı 25 Ocak 2008.
- ^ Siegel JM (June 2006). "The stuff dreams are made of: anatomical substrates of REM sleep". Doğa Sinirbilim. 9 (6): 721–2. doi:10.1038/nn0606-721. PMID 16732200. S2CID 7951497.
- ^ Nelson JP, McCarley RW, Hobson JA (October 1983). "REM sleep burst neurons, PGO waves, and eye movement information". Nörofizyoloji Dergisi. 50 (4): 784–97. doi:10.1152/jn.1983.50.4.784. PMID 6631463.
- ^ Hobson JA, Stickgold R, Pace-Schott EF (February 1998). "The neuropsychology of REM sleep dreaming". NeuroReport. 9 (3): R1–14. doi:10.1097/00001756-199802160-00033. PMID 9512371.
- ^ Wilson MA, McNaughton BL (July 1994). "Reactivation of hippocampal ensemble memories during sleep". Bilim. 265 (5172): 676–9. Bibcode:1994Sci ... 265..676W. doi:10.1126 / science.8036517. PMID 8036517.
- ^ Stickgold R, Walker MP (June 2007). "Sleep-dependent memory consolidation and reconsolidation". Uyku Tıbbı. 8 (4): 331–43. doi:10.1016/j.sleep.2007.03.011. PMC 2680680. PMID 17470412.
- ^ Buzsáki, G. (1 Ocak 1996). "Hipokampo-Neokortikal Diyalog". Beyin zarı. 6 (2): 81–92. doi:10.1093 / cercor / 6.2.81. PMID 8670641.
- ^ BuzsÁk, GyÖrgy (1 Haziran 1998). "Uyku sırasında hafıza konsolidasyonu: nörofizyolojik bir bakış açısı". Uyku Araştırmaları Dergisi. 7 (S1): 17–23. doi:10.1046 / j.1365-2869.7.s1.3.x. PMID 9682189.
- ^ Ferrara M, Moroni F, De Gennaro L, Nobili L (1 Ocak 2012). "Hipokampal uyku özellikleri: insan hafıza fonksiyonuyla ilişkiler". Nörolojide Sınırlar. 3: 57. doi:10.3389 / fneur.2012.00057. PMC 3327976. PMID 22529835.
- ^ "Harvard Tıp Fakültesi Uyku Temelleri".
- ^ Siegel, Jerome H. (2002). Uykunun ve Uyanmanın Sinir Kontrolü. Springer. ISBN 9780387955360.
- ^ a b Saper CB, Chou TC, Scammell TE (Aralık 2001). "Uyku anahtarı: uyku ve uyanıklığın hipotalamik kontrolü". Sinirbilimlerindeki Eğilimler. 24 (12): 726–31. doi:10.1016 / S0166-2236 (00) 02002-6. PMID 11718878. S2CID 206027570.
- ^ Lu J, Sherman D, Devor M, Saper CB (Haziran 2006). "REM uykusunun kontrolü için varsayılan bir flip-flop anahtarı". Doğa. 441 (7093): 589–94. Bibcode:2006Natur.441..589L. doi:10.1038 / nature04767. PMID 16688184. S2CID 4408007.
- ^ Sallanon M, Denoyer M, Kitahama K, Aubert C, Gay N, Jouvet M (1989). "Preoptik nöron lezyonlarının neden olduğu uzun süreli uykusuzluk ve kedide posterior hipotalamusa muscimol enjeksiyonu ile geçici geri dönüşü". Sinirbilim. 32 (3): 669–83. doi:10.1016/0306-4522(89)90289-3. PMID 2601839. S2CID 5791711.
- ^ Swett CP, Hobson JA (Eylül 1968). "Posterior hipotalamik lezyonların kedilerde uyku ve uyanmanın davranışsal ve elektrografik belirtileri üzerindeki etkileri". Arşivler Italiennes de Biologie. 106 (3): 283–93. PMID 5724423.
- ^ Borbély AA (1982). "İki süreçli uyku düzenlemesi modeli". İnsan Nörobiyolojisi. 1 (3): 195–204. PMID 7185792.
- ^ Edgar DM, Dement WC, Fuller CA (Mart 1993). "Sincap maymunlarında SCN lezyonlarının uyku üzerindeki etkisi: uyku-uyanıklık düzenlemesindeki rakip süreçler için kanıt". Nörobilim Dergisi. 13 (3): 1065–79. doi:10.1523 / JNEUROSCI.13-03-01065.1993. PMC 6576589. PMID 8441003.
- ^ Birendra N. Mallick; ve diğerleri, eds. (2011-07-14). Hızlı göz hareketi uykusu: düzenleme ve işlev. Cambridge, İngiltere: Cambridge University Press. ISBN 9780521116800.
- ^ McCormick DA, Bal T (1 Mart 1997). "Uyku ve uyarılma: talamokortikal mekanizmalar". Yıllık Nörobilim İncelemesi. 20 (1): 185–215. doi:10.1146 / annurev.neuro.20.1.185. PMID 9056712.
- ^ Sforza E, Montagna P, Tinuper P, Cortelli P, Avoni P, Ferrillo F, Petersen R, Gambetti P, Lugaresi E (Haziran 1995). "Ölümcül ailesel uykusuzlukta uyku-uyanıklık döngüsü anormallikleri. Talamusun uyku düzenlemesindeki rolünün kanıtı". Elektroensefalografi ve Klinik Nörofizyoloji. 94 (6): 398–405. doi:10.1016 / 0013-4694 (94) 00318-F. PMID 7607093.
- ^ Tinuper P, Montagna P, Medori R, Cortelli P, Zucconi M, Baruzzi A, Lugaresi E (Ağustos 1989). "Talamus uyku-uyanma döngüsünün düzenlenmesine katılır. Ölümcül ailevi talamik dejenerasyonda kliniko-patolojik bir çalışma". Elektroensefalografi ve Klinik Nörofizyoloji. 73 (2): 117–23. doi:10.1016/0013-4694(89)90190-9. PMID 2473878.
- ^ Coulon P, Budde T, Pape HC (Ocak 2012). "Uyku rölesi - talamusun merkezi ve merkezi olmayan uyku düzenlemesindeki rolü". Pflügers Arşivi. 463 (1): 53–71. doi:10.1007 / s00424-011-1014-6. PMID 21912835. S2CID 15459459.
- ^ Iwańczuk W, Guźniczak P (2015). "Uyku, uyarılma, farkındalık ve bilinç olaylarının nörofizyolojik temelleri. Bölüm 1". Anesteziyoloji Yoğun Terapi. 47 (2): 162–7. doi:10.5603 / AIT.2015.0015. PMID 25940332.
Yükselen retiküler aktivasyon sistemi (ARAS), sürekli bir uyanıklık durumundan sorumludur. Spinoretiküler yollar ve kraniyal sinirler (trigeminal sinir - polimodal yollar, koku alma siniri, optik sinir ve vestibülokoklear sinir - monomodal yollar) yoluyla iletilen çeşitli modalitelerin duyusal reseptörlerinden bilgi alır. Bu yollar talamusa doğrudan veya dolaylı olarak retiküler oluşum çekirdeklerinin (magnoselüler çekirdekler ve pontin tegmentumun retiküler çekirdekleri) medial kolonu yoluyla ulaşırlar. Retiküler aktive edici sistem, arka orta beynin ve ön ponsun dorsal kısmında başlar, diensefalonda devam eder ve daha sonra talamusa ve hipotalamusa ulaşan iki kısma ayrılır ve bunlar daha sonra serebral kortekse taşınır (Şekil 1). Talamik projeksiyonda, pons ve orta beyin (PPT) pedunculopontine tegmental çekirdeğinden ve pons ve orta beyin (LDT) nükleuslarının laterodorsal tegmental çekirdeğinden kaynaklanan kolinerjik nöronlar hakimdir [17, 18]. Hipotalamik projeksiyon, locus coeruleus'un (LC) noradrenerjik nöronlarını ve lateral hipotalamustan geçen ve histaminerjik tubero-mamillary nükleusun (TMN) aksonlarına ulaşan dorsal ve medyan raphe çekirdeklerinin (DR) serotoninerjik nöronlarını içerir. ön beyin, korteks ve hipokampusa uzanan yol. Kortikal uyarılma ayrıca substantia nigra (SN), ventral tegmenti alanı (VTA) ve periakuaduktal gri alanın (PAG) dopaminerjik nöronlarından yararlanır. Pons ve orta beyindeki daha az kolinerjik nöron, talamusu atlayarak ventral yol boyunca ön beyine projeksiyonlar gönderir [19, 20].
- ^ Malenka RC, Nestler EJ, Hyman SE (2009). "Bölüm 12: Uyku ve Uyandırma". Sydor A, Brown RY (editörler). Moleküler Nörofarmakoloji: Klinik Nörobilim Vakfı (2. baskı). New York, ABD: McGraw-Hill Medical. s. 295. ISBN 9780071481274.
RAS, dört monoaminerjik yolu içeren birkaç farklı devreden oluşan karmaşık bir yapıdır ... Norepinefrin yolu, lokus seruleus (LC) ve ilgili beyin sapı çekirdeklerinden kaynaklanır; serotonerjik nöronlar, beyin sapındaki raphe çekirdeklerinden de kaynaklanır; dopaminerjik nöronlar ventral tegmental alandan (VTA) kaynaklanır; ve histaminerjik yol, arka hipotalamusun tüberomamiller çekirdeğindeki (TMN) nöronlardan kaynaklanır. Bölüm 6'da tartışıldığı gibi, bu nöronlar, kısıtlı hücre gövdesi koleksiyonlarından beyin boyunca geniş bir alana yayılır. Norepinefrin, serotonin, dopamin ve histamin, karmaşık düzenleyici işlevlere sahiptir ve genel olarak uyanıklığı teşvik eder. Beyin sapındaki PT de ARAS'ın önemli bir bileşenidir. PT kolinerjik nöronların (REM-on hücreler) aktivitesi, REM uykusunu destekler. Uyanma sırasında, REM-on hücreleri, ARAS norepinefrin ve REM-off hücreleri olarak adlandırılan serotonin nöronlarının bir alt kümesi tarafından inhibe edilir.
- ^ Brudzynski SM (Temmuz 2014). "Yükselen mezolimbik kolinerjik sistem - olumsuz duygusal durumların başlangıcında yer alan retiküler aktive edici sistemin belirli bir bölümü". Moleküler Sinirbilim Dergisi. 53 (3): 436–45. doi:10.1007 / s12031-013-0179-1. PMID 24272957. S2CID 14615039.
ARAS'ın uyanma ve uyanıklığı sürdürme işlevlerinin anlaşılması, beyin sapı retiküler çekirdeğin içinden çıkan yükselen yollarla, farklı vericileri sentezleyen ve onları geniş alanlara salan pontomesensefalik çekirdeklerle birlikte çok sayıda nöron grubunun nörokimyasal keşifleriyle daha da karmaşık hale gelmiştir. beyin ve tüm neokorteks içinde (gözden geçirme için bkz. Jones 2003; Lin ve diğerleri 2011). Glutamaterjik, kolinerjik, noradrenerjik, dopaminerjik, serotonerjik, histaminerjik ve oreksinerjik sistemleri içerirler (inceleme için, Lin ve ark. 2011'e bakınız). ... ARAS, orta hat ve intralaminar talamik çekirdekler boyunca çalışan, tüm neokorteksin aktivitesini değiştirebilen yaygın, spesifik olmayan yolları temsil etti ve bu nedenle, bu sistem başlangıçta doğal uyaranlara ve altında yatan kritik sisteme genel bir uyarılma sistemi olarak önerildi. uyanıklık (Moruzzi ve Magoun 1949; Lindsley ve diğerleri 1949; Starzl ve diğerleri 1951, Şekil 1'de noktalı alana bakınız). ... Sıçanda yapılan yakın tarihli bir çalışmada, uyanıklık durumunun çoğunlukla parabrakial çekirdek ve precoeruleus bölgelerinden bazal ön beyine yükselen glutamaterjik projeksiyonla korunduğu ve ardından serebral kortekse aktarıldığı bulundu (Fuller ve ark.2011) ). ... Anatomik çalışmalar, kolinerjik hücre gruplarının bulunduğu alanlardan kaynaklanan ve biri talamustan, diğeri ise hipotalamus ve preoptik alan boyunca ventral olarak seyahat eden ve limbik sistemle karşılıklı olarak bağlantılı iki ana yol olduğunu göstermiştir (Nauta ve Kuypers 1958; Siegel 2004). ... Talamik retiküler çekirdekle olan kolinerjik bağlantılarda sayıldığı gibi ...
- ^ Schwartz MD, Kilduff TS (Aralık 2015). "Uyku ve Uyanıklığın Nörobiyolojisi". Kuzey Amerika Psikiyatri Klinikleri. 38 (4): 615–44. doi:10.1016 / j.psc.2015.07.002. PMC 4660253. PMID 26600100.
Bu yükselen retiküler aktive edici sistem (ARAS), kolinerjik laterodorsal ve pedunculopontine tegmentum (LDT / PPT), noradrenergic locus coeruleus (LC), serotonerjik (5-HT) Raphe çekirdekleri ve dopaminerjik ventral tegmental alan (VTA), substantia nigra'dan oluşur. ) ve korteksi talamus, hipotalamus ve BF yoluyla doğrudan ve dolaylı olarak uyaran periakuaduktal gri projeksiyonlar.6, 12-18 Bu aminerjik ve katekolaminerjik popülasyonlar, sisteme fonksiyonel fazlalık ve esneklik kazandıran çok sayıda ara bağlantıya ve paralel projeksiyonlara sahiptir.6, 13, 19 ... Daha yakın zamanlarda, fasiyal sinire bitişik medüller parafasiyal bölge (PZ) anatomik, elektrofizyolojik ve kemo- ve optogenetik çalışmalara dayanarak uykuyu teşvik eden bir merkez olarak tanımlandı.23, 24 GABAerjik PZ nöronları, BF'ye projeksiyon yapan glutamaterjik parabrakial (PB) nöronları inhibe eder,25 böylece uyanıklık ve REM uykusu pahasına NREM uykusunu teşvik eder. ... Hcrt nöronları beyin ve omurilik boyunca geniş bir alana yayılır92, 96, 99, 100 TM'nin HA hücreleri gibi uyanmayı destekleyen hücre gruplarına büyük projeksiyonlar dahil,101 dorsal Raphe çekirdeklerinin (DRN) 5-HT hücreleri,101 LC'nin noradrenerjik hücreleri,102 ve LDT, PPT ve BF'deki kolinerjik hücreler.101, 103 ... Hcrt, LC de dahil olmak üzere uyanma ve uyarılma ile ilgili hücresel sistemleri doğrudan uyarır,102, 106, 107 DRN,108, 109 TM,110-112 LDT,113, 114 kolinerjik BF,115 ve VTA'da hem dopamin (DA) hem de DA olmayan nöronlar.116, 117
- ^ Max DT (Mayıs 2010). "Uykunun Sırları". National Geographic Dergisi.
- ^ Krueger JM, Rector DM, Roy S, Van Dongen HP, Belenky G, Panksepp J (Aralık 2008). "Nöronal toplulukların temel bir özelliği olarak uyku". Doğa Yorumları. Sinirbilim. 9 (12): 910–9. doi:10.1038 / nrn2521. PMC 2586424. PMID 18985047.
- ^ Wolstenholme GE, O'Connor M (1961). Ciba Vakfı Uykunun Doğası Sempozyumu. Boston: Küçük, Kahverengi. ISBN 978-0-470-71922-0.
- ^ "Uyku Müfredatı. B. Uykunun Filogeni". Uyku Araştırmaları Derneği, Eğitim Komitesi. Arşivlenen orijinal 2005-03-18 tarihinde. Alındı 26 Eylül 2010.
- ^ "Uykunun İşlevi.". Scribd.com. Erişim tarihi: 1 Aralık 2011.
- ^ Krueger JM, Obál F, Fang J (Haziran 1999). "Neden uyuyoruz: uyku fonksiyonunun teorik bir görünümü". Uyku Tıbbı Yorumları. 3 (2): 119–29. doi:10.1016 / S1087-0792 (99) 90019-9. PMID 15310481.
- ^ Krueger JM, Obál F (Haziran 1993). "Bir nöronal grup uyku fonksiyonu teorisi". Uyku Araştırmaları Dergisi. 2 (2): 63–69. doi:10.1111 / j.1365-2869.1993.tb00064.x. PMID 10607073.
- ^ Medic G, Wille M, Hemels ME (2017-05-19). "Uyku bozukluğunun kısa ve uzun vadeli sağlık sonuçları". Doğa ve Uyku Bilimi. 9: 151–161. doi:10.2147 / nss.s134864. PMC 5449130. PMID 28579842.
- ^ Choi, Charles Q. (25 Ağustos 2009) Yeni Teori Soruları Neden Uyuuyoruz, LiveScience.com.
- ^ Xie L, Kang H, Xu Q, Chen MJ, Liao Y, Thiyagarajan M, O'Donnell J, Christensen DJ, Nicholson C, Iliff JJ, Takano T, Deane R, Nedergaard M (Ekim 2013). "Uyku, yetişkin beynindeki metabolitlerin temizlenmesini sağlar". Bilim. 342 (6156): 373–7. Bibcode:2013Sci ... 342..373X. doi:10.1126 / science.1241224. PMC 3880190. PMID 24136970.
- ^ a b c Nedergaard M, Goldman SA (2016). "Beyin göçü". Bilimsel amerikalı. 314 (Mart): 44–49. Bibcode:2016SciAm.314c..44N. doi:10.1038 / bilimselamerican0316-44. PMC 5347443. PMID 27066643.
- ^ a b Strazielle N, Ghersi-Egea JF (Mayıs 2013). "Küçük bileşiklerin ve makromoleküllerin beyin düzeniyle ilişkili olarak kan-beyin arayüzlerinin fizyolojisi". Moleküler Eczacılık. 10 (5): 1473–91. doi:10.1021 / mp300518e. PMID 23298398.
- ^ Gümüştekín K, Seven B, Karabulut N, Aktaş O, Gürsan N, Aslan S, Keleş M, Varoglu E, Dane S (Kasım 2004). "Uykusuzluk, nikotin ve selenyumun sıçanlarda yara iyileşmesi üzerindeki etkileri". The International Journal of Neuroscience (Gönderilen makale). 114 (11): 1433–42. doi:10.1080/00207450490509168. PMID 15636354. S2CID 30346608.
- ^ Zager A, Andersen ML, Ruiz FS, Antunes IB, Tufik S (Temmuz 2007). "Akut ve kronik uyku kaybının sıçanların immün modülasyonu üzerindeki etkileri". Amerikan Fizyoloji Dergisi. Düzenleyici, Bütünleştirici ve Karşılaştırmalı Fizyoloji. 293 (1): R504–9. doi:10.1152 / ajpregu.00105.2007. PMID 17409265.
- ^ Opp MR (Ocak 2009). "Bağışıklık sistemini beslemek için uyumak: memeli uykusu ve parazitlere karşı direnç". BMC Evrimsel Biyoloji. 9: 8. doi:10.1186/1471-2148-9-8. PMC 2633283. PMID 19134176.
- ^ Peres, Judy (14 Mart 2012) Zzz'lerinizi almak için iyi bir neden Chicago Tribune Health, 26 Mart 2014'te alındı
- ^ Jenni OG, Molinari L, Caflisch JA, Largo RH (Ekim 2007). "1 ila 10 yaş arası uyku süresi: büyümeye kıyasla değişkenlik ve istikrar". Pediatri. 120 (4): e769–76. doi:10.1542 / peds. 2006-3300. PMID 17908734. S2CID 16754966.
- ^ "Beyin uyku sırasında toksinleri dışarı atabilir". Ulusal Sağlık Enstitüleri. Alındı 25 Ekim 2013.
- ^ Siegel JM (Ekim 2005). "Memeli uykusunun işlevlerine dair ipuçları". Doğa. 437 (7063): 1264–71. Bibcode:2005Natur.437.1264S. doi:10.1038 / nature04285. PMID 16251951. S2CID 234089.
- ^ a b Cirelli C, Tononi G (Ağustos 2013). "Muhtemelen Budama". Bilimsel amerikalı. 309 (2): 34–39. Bibcode:2013SciAm.309b..34T. doi:10.1038 / bilimselamerican0813-34. PMID 23923204.
- ^ Lewy AJ, Wehr TA, Goodwin FK, Newsome DA, Markey SP (Aralık 1980). "Işık, insanlarda melatonin salgılanmasını baskılar". Bilim. 210 (4475): 1267–9. Bibcode:1980Sci ... 210.1267L. doi:10.1126 / science.7434030. PMID 7434030.
- ^ Leproult R, Colecchia EF, L'Hermite-Balériaux M, Van Cauter E (Ocak 2001). "Sabahları loştan parlak ışığa geçiş, kortizol seviyelerinin aniden yükselmesine neden olur". Klinik Endokrinoloji ve Metabolizma Dergisi. 86 (1): 151–7. doi:10.1210 / jcem.86.1.7102. PMID 11231993.
- ^ Van Cauter E, Kerkhofs M, Caufriez A, Van Onderbergen A, Thorner MO, Copinschi G (Haziran 1992). "Normal insanda büyüme hormonu salgılanmasının nicel bir tahmini: tekrarlanabilirlik ve uyku ve günün saati ile ilişkisi". Klinik Endokrinoloji ve Metabolizma Dergisi. 74 (6): 1441–50. doi:10.1210 / jcem.74.6.1592892. PMID 1592892.
- ^ Kern W, Dodt C, Born J, Fehm HL (Ocak 1996). "Yaşlanma sürecinde gece uykusu sırasında kortizol ve büyüme hormonu salgısındaki değişiklikler". Gerontology Dergileri. Seri A, Biyolojik Bilimler ve Tıp Bilimleri. 51 (1): M3–9. doi:10.1093 / gerona / 51A.1.M3. PMID 8548511.
- ^ a b Knutson KL, Spiegel K, Penev P, Van Cauter E (Haziran 2007). "Uykusuzluğun metabolik sonuçları". Uyku Tıbbı Yorumları. 11 (3): 163–78. doi:10.1016 / j.smrv.2007.01.002. PMC 1991337. PMID 17442599.
- ^ Spiegel K, Leproult R, Van Cauter E (Ekim 1999). "Uyku borcunun metabolik ve endokrin fonksiyon üzerindeki etkisi". Lancet. 354 (9188): 1435–9. doi:10.1016 / S0140-6736 (99) 01376-8. PMID 10543671. S2CID 3854642.
- ^ Plihal W, J doğdu (1997). "Erken ve geç gece uykusunun bildirimsel ve prosedürel bellek üzerindeki etkileri". Bilişsel Sinirbilim Dergisi. 9 (4): 534–547. doi:10.1162 / jocn.1997.9.4.534. PMID 23968216. S2CID 3300300.
- ^ a b c d Rasch B, Büchel C, Gais S, Born J (2007). "Yavaş dalga uyku istemi bildirimsel bellek konsolidasyonu sırasında koku ipuçları". Bilim. 315 (5817): 1426–9. Bibcode:2007Sci ... 315.1426R. doi:10.1126 / science.1138581. PMID 17347444. S2CID 19788434.
- ^ a b c d e J, Wilhelm I (2012) doğdu. "Uyku sırasında belleğin sistem konsolidasyonu". Psikolojik Araştırma. 76 (2): 192–203. doi:10.1007 / s00426-011-0335-6. PMC 3278619. PMID 21541757.
- ^ Diekelmann S, J doğdu (2010). "Uykunun hafıza işlevi". Doğa Yorumları Nörobilim. 11 (2): 114–126. doi:10.1038 / nrn2762. PMID 20046194. S2CID 1851910.
- ^ a b Rasch B, J doğdu (2013). "Uykunun hafızadaki rolü hakkında". Fizyolojik İncelemeler. 93 (2): 681–766. doi:10.1152 / physrev.00032.2012. PMC 3768102. PMID 23589831.
- ^ a b Schreiner T, Rasch B (2015). "Uyku Sırasında Sözlü İşaretle Kelime Öğrenmeyi Geliştirme". Beyin zarı. 25 (11): 4169–4179. doi:10.1093 / cercor / bhu139. PMID 24962994.
- ^ a b c Schreiner, Rasch (2017). "Uyku sırasında dil öğrenimi için hafızanın yeniden etkinleştirilmesinin yararlı rolü: Bir inceleme" (PDF). Beyin ve Dil. 167: 94–105. doi:10.1016 / j.bandl.2016.02.005. PMID 27036946. S2CID 3377186.
- ^ a b Ngo H, Martinetz T, Doğan J, Mölle M (2013). "Uykunun İşitsel Kapalı Döngü Stimülasyonu Yavaş Salınım Belleği Geliştirir". Nöron. 78 (3): 545–553. doi:10.1016 / j.neuron.2013.03.006. PMID 23583623.
- ^ Klinzing J, Kugler S, Soekadar S, Rasch B, Doğan J, Diekelmann S (2018). "Yavaş dalga uykusu sırasında koku uyarısı, düşük kolinerjik tondan bağımsız olarak hafızaya fayda sağlar". Psikofarmakoloji. 235 (1): 291–299. doi:10.1007 / s00213-017-4768-5. PMC 5748395. PMID 29119218.
- ^ Schreiner T, Doeller C, Jensen O, Rasch B, Staudigl T (2018). "Theta Faz-Koordineli Bellek Yeniden Aktivasyonu, NREM Uykusu Sırasında Yavaş Salınımlı Bir Ritmde Yeniden Gerçekleşiyor". Hücre Raporları. 25 (2): 296–301. doi:10.1016 / j.celrep.2018.09.037. PMC 6198287. PMID 30304670.
- ^ Schreiner T, Göldi M, Rasch B (2015). "Uyku sırasında kelime bilgisi, sonraki tanıma testi sırasında teta aktivitesini artırır". Psikofizyoloji. 52 (11): 1538–1543. doi:10.1111 / psyp.12505. PMID 26235609.
- ^ Groch, S., Schreiner, T., Rasch, B., Huber, R. ve Wilhelm, I. (2017). Uyku sırasında hedeflenen hafıza yeniden aktivasyonunun yararlı etkisi için önceden bilgi gereklidir. Bilimsel Raporlar, 7
- ^ Tononi G (2011). Uyku fonksiyonu ve sinaptik homeostaz (Konferans konuşması). Allen Enstitüsü.
- ^ Carlson NR, Miller HL, Heth DS, Donahoe JW, Martin GN (2010). Psikoloji Davranış Bilimi, Kitaplar A La Carte Edition. Pearson College Div. ISBN 978-0-205-76223-1.
- ^ Hock RR (2013). "Uyumak, şüphesiz rüya görmek ..." Psikolojiyi Değiştiren Kırk Araştırma (7. baskı). Upper Saddle River, NJ: Pearson Education. s. 42–49. ISBN 978-0-205-91839-3.
- ^ Dement W (1966). "Bölüm 12: Rüya Yoksunluğunun Etkisi: Her gece belirli bir miktarda rüya görme ihtiyacı, son deneylerde önerilmektedir". Brian Fizyolojisi ve Psikolojisi. California Üniversitesi Yayınları.
- ^ a b Van Dongen HP, Maislin G, Mullington JM, Dinges DF (Mart 2003). "Ek uyanıklığın kümülatif maliyeti: kronik uyku kısıtlaması ve toplam uyku yoksunluğundan nörodavranışsal işlevler ve uyku fizyolojisi üzerindeki doz-yanıt etkileri". Uyku. 26 (2): 117–26. doi:10.1093 / uyku / 26.2.117. PMID 12683469.
- ^ Connor J, Norton R, Ameratunga S, Robinson E, Civil I, Dunn R, Bailey J, Jackson R (Mayıs 2002). "Sürücünün uykulu hali ve araçtaki yolcuların ciddi şekilde yaralanma riski: nüfus temelli vaka kontrol çalışması". BMJ. 324 (7346): 1125. doi:10.1136 / bmj.324.7346.1125. PMC 107904. PMID 12003884.
- ^ Lim J, Dinges DF (1 Mayıs 2008). "Uykusuzluk ve tetikte dikkat". New York Bilimler Akademisi Yıllıkları. 1129 (1): 305–22. Bibcode:2008NYASA1129..305L. doi:10.1196 / annals.1417.002. PMID 18591490. S2CID 14220859.
- ^ Wright KP, Badia P, Myers BL, Plenzler SC (Mart 1997). "Uzun süreli uyku yoksunluğu sırasında bozulmuş uyanıklık ve performansa karşı bir önlem olarak parlak ışık ve kafein kombinasyonu". Uyku Araştırmaları Dergisi. 6 (1): 26–35. doi:10.1046 / j.1365-2869.1997.00022.x. PMID 9125696. S2CID 6615362.
- ^ a b Durmer JS, Dinges DF (Mart 2005). "Uyku yoksunluğunun nörobilişsel sonuçları". Nörolojide Seminerler. 25 (1): 117–29. CiteSeerX 10.1.1.580.1922. doi:10.1055 / s-2005-867080. PMID 15798944.
- ^ Drummond SP, Gillin JC, Brown GG (Haziran 2001). "Uykusuzluktan sonra bölünmüş bir dikkat görevi sırasında artan beyin tepkisi". Uyku Araştırmaları Dergisi. 10 (2): 85–92. doi:10.1046 / j.1365-2869.2001.00245.x. PMID 11422722.
- ^ van der Helm E, Gujar N, Walker MP (Mart 2010). "Uyku yoksunluğu, insan duygularının doğru bir şekilde tanınmasını engeller". Uyku. 33 (3): 335–42. doi:10.1093 / uyku / 33.3.335. PMC 2831427. PMID 20337191.
- ^ Tsuno N, Besset A, Ritchie K (Ekim 2005). "Uyku ve depresyon". Klinik Psikiyatri Dergisi. 66 (10): 1254–69. doi:10.4088 / JCP.v66n1008. PMID 16259539.
- ^ Chee MW, Chuah LY (Ağustos 2008). "Uyku ve uykusuzluğun hafızayı ve bilişi nasıl etkilediğine dair işlevsel beyin görüntüleme içgörüler". Nörolojide Güncel Görüş. 21 (4): 417–23. doi:10.1097 / WCO.0b013e3283052cf7. PMID 18607201. S2CID 15785066.
- ^ a b Aurora R, Zak R, Maganti R, Auerbach S, Casey K, Chowdhuri S, Morgenthaler T (2010). "REM uyku davranış bozukluğunun (RBD) tedavisi için en iyi uygulama kılavuzu". Klinik Uyku Tıbbı Dergisi. 6 (1): 85–95. doi:10.5664 / jcsm.27717. PMC 2823283. PMID 20191945.
- ^ a b Bjørnarå D, Toft (2013). "Parkinson hastalığında REM uykusu davranış bozukluğu - Cinsiyet farkı var mı?". Parkinsonizm ve İlgili Bozukluklar. 19 (1): 120–122. doi:10.1016 / j.parkreldis.2012.05.027. PMID 22726815.
- ^ a b Bjørnarå K, Dietrichs E, Toft M (2015). "Parkinson hastalığında olası hızlı göz hareketi uyku davranış bozukluğunun boylamsal değerlendirmesi". Avrupa Nöroloji Dergisi. 22 (8): 1242–1244. doi:10.1111 / ene.12723. S2CID 5237488.
- ^ a b c Wang P, Wing YK, Xing J, vd. (2016). "Olası Alzheimer hastalığı olan hastalarda hızlı göz hareketi uyku davranış bozukluğu". Yaşlanma Clin Exp Res. 28 (5): 951. doi:10.1007 / s40520-015-0382-8. PMID 26022447. S2CID 29014852.
- ^ a b c d McCarter S, Howell J (2017). "Alzheimer Dışı Demanslarda REM Uyku Davranışı Bozukluğu ve Diğer Uyku Bozuklukları". Güncel Uyku Tıbbı Raporları. 3 (3): 193–203. doi:10.1007 / s40675-017-0078-z. S2CID 148800517.
- ^ a b c d e f g Dick-Muehlke, C. (2015). Alzheimer hastalığında bireyin değişen bakış açılarının psikososyal çalışmaları (Premier Referans Kaynağı). Hershey, PA: Tıbbi Bilgi Bilimi Referansı.
- ^ Revonsuo A (Aralık 2000). "Rüyaların yeniden yorumlanması: rüya görme işlevinin evrimsel bir hipotezi". Davranış ve Beyin Bilimleri. 23 (6): 877–901, tartışma 904–1121. doi:10.1017 / s0140525x00004015. PMID 11515147.
- ^ Montangero J (2000). "Rüya işlevi hakkında daha genel bir evrimsel hipotez". Davranış ve Beyin Bilimleri. 23 (6): 972–973. doi:10.1017 / s0140525x00664026.
- ^ Stickgold R, Hobson JA, Fosse R, Fosse M (Kasım 2001). "Uyku, öğrenme ve rüyalar: çevrimdışı bellek yeniden işleme". Bilim. 294 (5544): 1052–7. Bibcode:2001Sci ... 294.1052S. doi:10.1126 / science.1063530. PMID 11691983. S2CID 8631140.
- ^ Greenberg R, Katz H, Schwartz W, Pearlman C (1992). "Psikanalitik rüya kuramının araştırmaya dayalı yeniden değerlendirilmesi". Amerikan Psikanaliz Derneği Dergisi. 40 (2): 531–50. doi:10.1177/000306519204000211. PMID 1593084. S2CID 24985171.
- ^ Sadeh A, Keinan G, Daon K (Eylül 2004). "Stresin uyku üzerindeki etkileri: başa çıkma tarzının ılımlı rolü". Sağlıklı psikoloji. 23 (5): 542–5. CiteSeerX 10.1.1.476.8304. doi:10.1037/0278-6133.23.5.542. PMID 15367075.
- ^ Nielsen T, Levin R (Ağustos 2007). "Kabuslar: yeni bir nörobilişsel model". Uyku Tıbbı Yorumları. 11 (4): 295–310. doi:10.1016 / j.smrv.2007.03.004. PMID 17498981.
- ^ Pagel JF (2010). "Uyuşturucular, Rüyalar ve Kabuslar". Uyku Tıbbı Klinikleri. 5 (2): 277–287. doi:10.1016 / j.jsmc.2010.01.007.
- ^ Levin R, Nielsen T (1 Nisan 2009). "Kabuslar, Kötü Rüyalar ve Duygu Düzensizliği: Bir Gözden Geçirme ve Yeni Bir Nörobilişsel Rüya Modeli". Psikolojik Bilimde Güncel Yönler. 18 (2): 84–88. doi:10.1111 / j.1467-8721.2009.01614.x. S2CID 41082725.