Kalıtılabilirlik - Heritability

Kalıtılabilirlik bir istatistik alanlarında kullanılan üreme ve genetik derecesini tahmin eden varyasyon içinde fenotipik özellik içinde nüfus bunun sebebi genetik çeşitlilik o popülasyondaki bireyler arasında.[1] Çevresel faktörlerin varyasyonunun aksine, bir özelliğin varyasyonunun ne kadarının genetik faktörlerin varyasyonuna atfedilebileceğini ölçer. Kalıtılabilirlik kavramı aşağıdaki soru şeklinde ifade edilebilir: "Belirli bir özellikteki varyasyonun bir popülasyon içindeki oranı nedir? değil çevre mi yoksa rastgele şansla mı açıklanıyor? "[2]
Bir özellikte ölçülen varyasyonun diğer nedenleri şu şekilde karakterize edilir: çevresel faktörler, dahil olmak üzere gözlemsel hata. İnsanların kalıtımla ilgili araştırmalarında, bunlar genellikle aynı hanede yetişmiş kişilerin, olmayan kişilere aşağı yukarı benzer olmalarına neden olup olmadıklarına bağlı olarak "paylaşılan çevre" ve "paylaşılmayan ortam" dan faktörlere paylaştırılır.
Kalıtılabilirlik, bir popülasyondaki ilgili bireyler arasındaki bireysel fenotipik varyasyonun karşılaştırılmasıyla, bireysel fenotip ve genotip verileri arasındaki ilişki incelenerek tahmin edilir.[3][4] hatta genom çapında ilişkilendirme çalışmasından (GWAS) özet düzeydeki verileri modelleyerek.[5] Kalıtım, önemli bir kavramdır. nicel genetik, Özellikle de seçici yetiştirme ve davranış genetiği (Örneğin, ikiz çalışmalar ). Teknik tanımının genel olarak anlaşılan halk tanımından farklı olması nedeniyle çok fazla kafa karışıklığının kaynağıdır. Bu nedenle kullanımı, davranışsal özelliklerin "kalıtsal" olduğu veya özellikle genler aracılığıyla aktarıldığı yanlış izlenimini taşır.[6] Davranışsal genetikçiler ayrıca, genlerin ve ortamların davranışsal özelliklere ayrı, ilave bir şekilde katkıda bulunduğu varsayımına dayanarak kalıtsallık analizleri yaparlar.[7]
Genel Bakış
Kalıtılabilirlik, fenotip değişkenliği fraksiyonunu ölçer. genetik çeşitlilik. Bu, bireysel bir fenotipin bu fraksiyonunun genetikten kaynaklandığını söylemekle aynı şey değildir. Örneğin, kişilik özelliklerinin kalıtsallığı yaklaşık 0,6 olduğu için, bu kişiliğinizin% 60'ının ebeveynlerinizden miras kaldığı ve% 40'ının çevreden geldiğini söylemek yanlıştır. Ek olarak, kalıtılabilirlik, çevrenin daha fazla çeşitliliğe katkıda bulunmaya başlaması gibi herhangi bir genetik değişiklik meydana gelmeden değişebilir. Örnek olarak, her ikisinin de genler ve çevrenin zekayı etkileme potansiyeli vardır. Genetik çeşitlilik artarsa kalıtılabilirlik artabilir, bu da bireylerin farklı zeka seviyeleri gösterme gibi daha fazla fenotipik varyasyon göstermesine neden olabilir. Öte yandan, çevresel çeşitlilik azalırsa kalıtılabilirlik de artabilir, bu da bireylerin daha benzer düzeylerde zeka göstermesi gibi daha az fenotipik varyasyon göstermesine neden olur. Genetik daha fazla varyasyona katkıda bulunduğunda veya genetik olmayan faktörler daha az varyasyona katkıda bulunduğundan kalıtılabilirlik artar; önemli olan göreceli katkıdır. Kalıtılabilirlik, belirli bir ortamdaki belirli bir popülasyona özgüdür. Sonuç olarak, bir özelliğin yüksek kalıtsallığı, özelliğin çevresel etkilere çok duyarlı olmadığı anlamına gelmez.[8] Kalıtım aynı zamanda çevredeki değişiklikler, göç, akraba veya araştırılan popülasyonda kalıtsallığın kendisinin ölçülme şekli.[9] Bir özelliğin kalıtılabilirliği, söz konusu özelliğin bir bireyde genetik olarak belirlenme derecesinin bir ölçüsü olarak yorumlanmamalıdır.[10][11]
Fenotipin çevreye bağımlılığının derecesi, ilgili genlerin bir işlevi de olabilir. Kalıtımla ilgili konular karmaşıktır çünkü genler yönlendirmek ortaya çıkan tüm ortamlarda ifadesini neredeyse kaçınılmaz kılan bir fenotip. Aynı genotipe sahip bireyler, adı verilen bir mekanizma aracılığıyla farklı fenotipler de sergileyebilirler. fenotipik esneklik, bu da kalıtımın bazı durumlarda ölçülmesini zorlaştırır. Moleküler biyolojideki son kavrayışlar, transkripsiyonel çevresel değişikliklerle ilişkili bireysel genlerin aktivitesi. Bununla birlikte, çevreden etkilenmeyen çok sayıda gen vardır.[12]
Kalıtım kullanımının tahminleri istatistiksel analizler bireyler arasındaki farklılıkların nedenlerini belirlemeye yardımcı olmak. Kalıtım varyansla ilgili olduğu için, zorunlu olarak bir popülasyondaki bireyler arasındaki farklılıkların bir açıklamasıdır. Kalıtım olabilir tek değişkenli - tek bir özelliği - veya çok değişkenli - birden çok özellik arasındaki genetik ve çevresel ilişkileri aynı anda incelemek. Bu, farklı fenotipler arasındaki genetik örtüşme testine izin verir: örneğin saç rengi ve göz rengi. Çevre ve genetik de etkileşime girebilir ve kalıtım analizleri bu etkileşimleri test edebilir ve inceleyebilir (GxE modelleri).
Kalıtım analizleri için bir ön koşul, hesaba katılması gereken bazı popülasyon varyasyonlarının olmasıdır. Bu son nokta, kalıtımın popülasyonda değişmez olan faktörlerin etkisini hesaba katamayacağı gerçeğini vurgulamaktadır. Faktörler, yoksa ve popülasyonda mevcut değilse, örneğin kimsenin belirli bir antibiyotik veya her yerde mevcut oldukları için, sanki herkes içki içiyormuş gibi Kahve. Uygulamada, tüm insan davranış özellikleri değişir ve neredeyse tüm özellikler bir miktar kalıtım gösterir.[13]
Tanım
Herhangi bir belirli fenotip olabilir modellenmiş genetik ve çevresel etkilerin toplamı olarak:[14]
- Fenotip (P) = Genotip (G) + Çevre (E).
Benzer şekilde, özellikteki fenotipik varyans - Var (P) - aşağıdaki gibi etkilerin toplamıdır:
- Var (P) = Var (G) + Var (E) + 2 Cov (G,E).
Planlı bir deneyde Cov (G,E) 0'da kontrol edilebilir ve tutulabilir. Bu durumda kalıtım, olarak tanımlanır:[15]


H2 geniş anlamda kalıtsallıktır. Bu, katkı maddesi dahil olmak üzere bir popülasyonun fenotipik varyansına tüm genetik katkıları yansıtır. baskın, ve epistatik (çok genli etkileşimler) ve anne ve baba etkileri, bireylerin ebeveynlerinin fenotipinden doğrudan etkilendiği durumlarda, örneğin Süt memelilerde üretim.
Genetik varyansın özellikle önemli bir bileşeni, ek varyans, Var (A) 'dır, bu varyansın ortalama etkilerinden (toplamsal etkiler) aleller. Her ebeveyn, başına tek bir alel geçtiği için mahal her bir yavru için, ebeveyn-çocuk benzerliği, tek alellerin ortalama etkisine bağlıdır. Bu nedenle, toplamsal varyans, ebeveyn-çocuk benzerliğinden sorumlu varyansın genetik bileşenini temsil eder. Fenotipik varyansın ilave genetik kısmı, Dar anlamda kalıtım olarak bilinir ve şu şekilde tanımlanır:

Büyük harf H2 geniş anlam ve küçük harf belirtmek için kullanılır h2 dar anlamda.
Ek bir ayak parmağı veya belirli hastalıklar gibi sürekli olmayan ancak ikiye bölünmüş özellikler için, çeşitli alellerin katkısı, bir eşiği aşan bir özellik olarak kendini gösteren bir toplam olarak kabul edilebilir. sorumluluk eşiği modeli kalıtımın tahmin edilebildiği ve seçimin modellenebildiği.
Katkı varyansı aşağıdakiler için önemlidir: seçim. Hayvancılığın iyileştirilmesi gibi seçici bir baskı uygulanırsa, özelliğin tepkisi doğrudan dar anlamda kalıtsallıkla ilgilidir. Özelliğin ortalaması, seçilen ebeveynlerin ortalamasının, seçilen ebeveynlerin seçildiği nüfusun ortalamasından ne kadar farklı olduğunun bir fonksiyonu olarak gelecek nesilde artacaktır. Gözlenen seçime cevap dar anlamda kalıtsallığın tahminine yol açar ( Kalıtımın farkına varıldı). Temelde yatan prensip budur yapay seçim veya üreme.
Misal
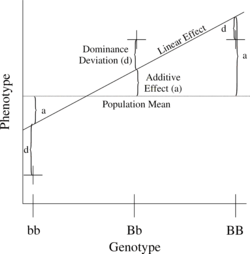
En basit genetik model, bir kantitatif fenotipi etkileyen iki alleli (b ve B) içeren tek bir lokusu içerir.
Sayısı B aleller 0, 1 veya 2 arasında değişebilir. Herhangi bir genotip için BbenBjBeklenen fenotip daha sonra genel ortalamanın, doğrusal bir etkinin ve bir baskınlık sapmasının toplamı olarak yazılabilir:
- = Nüfus ortalama + Katkı Etkisi () + Hakimiyet Sapması ().



Bu lokustaki toplamsal genetik varyans, ağırlıklı ortalama Katkı etkilerinin karelerinin sayısı:

nerede

Hakimiyet sapmalarının varyansı için benzer bir ilişki vardır:

nerede

doğrusal regresyon Genotip üzerindeki fenotipin etkisi Şekil 1'de gösterilmektedir.
Varsayımlar
İnsan özelliklerinin toplam kalıtımsallığı tahminleri, "eklenebilirlik varsayımı" olarak adlandırılan epistazın olmadığını varsayar. Bazı araştırmacılar bu tür tahminlere "kalıtımın olmaması "bilinen genetik lokuslar tarafından hesaba katılmayan, toplamsallık varsayımı bu tahminleri geçersiz kılabilir.[16] Ayrıca ergen zekası ve ergen zekası ve davranış genetiği araştırmalarında toplamsallık varsayımının sıklıkla ihlal edildiğine dair bazı ampirik kanıtlar vardır akademik başarı.[17]
Kalıtımın tahmin edilmesi
Sadece beri P doğrudan gözlemlenebilir veya ölçülebilir, kalıtılabilirlik, genetik veya çevresel benzerlik seviyelerinde değişiklik gösteren deneklerde gözlemlenen benzerliklerden tahmin edilmelidir. istatistiksel tahmin etmek için gerekli analizler genetik ve çevre varyansın bileşenleri örnek özelliklerine bağlıdır. Kısaca, çok farklı düzeylerde genetik ilişkiye sahip bireylerden elde edilen veriler kullanılarak daha iyi tahminler elde edilir - örneğin ikizler, daha uzaktan ilgili (ve dolayısıyla daha az benzer) konulardan ziyade kardeşler, ebeveynler ve çocuklar. standart hata kalıtım için tahminler, büyük örneklem büyüklükleriyle geliştirilmiştir.
İnsan olmayan popülasyonlarda, bilgilerin kontrollü bir şekilde toplanması genellikle mümkündür. Örneğin, çiftlik hayvanları arasında bir boğanın çok sayıda inekten yavru üretmesi ve ortamları kontrol etmesi kolaydır. Böyle deneysel kontrol doğal olarak oluşan ilişkilere ve ortamlara dayanarak insan verilerini toplarken genellikle mümkün değildir.
Klasik nicel genetikte, kalıtsallığın tahminine ilişkin iki düşünce ekolü vardı.
Bir düşünce okulu tarafından geliştirilmiştir Sewall Wright -de Chicago Üniversitesi ve daha da popüler hale geldi C. C. Li (Chicago Üniversitesi ) ve J. L. Lush (Iowa Eyalet Üniversitesi ). Korelasyonların analizine ve buna bağlı olarak regresyona dayanır. Yol Analizi tarafından geliştirilmiştir Sewall Wright kalıtsallığı tahmin etmenin bir yolu olarak.
İkincisi başlangıçta tarafından geliştirilmiştir R. A. Fisher ve genişletildi Edinburgh Üniversitesi, Iowa Eyalet Üniversitesi, ve Kuzey Karolina Eyalet Üniversitesi yanı sıra diğer okullar. Dayanmaktadır varyans analizi akrabaların sınıf içi korelasyonunu kullanarak ıslah çalışmalarının. Varyans bileşenlerini (ve dolayısıyla kalıtımı) tahmin etmenin çeşitli yöntemleri ANOVA bu analizlerde kullanılmaktadır.
Bugün, kalıtım, genel soyağacından tahmin edilebilir. doğrusal karışık modeller ve den genomik ilişki genetik belirteçlerden tahmin edilmektedir.
İnsan kalıtımına ilişkin araştırmalar, genellikle benimseme çalışması tasarımlarını kullanır. tek yumurta ikizi hayatlarının erken dönemlerinde ayrılmış ve farklı ortamlarda büyümüş olan. Bu tür bireyler aynı genotiplere sahiptir ve genotip ve çevrenin etkilerini ayırmak için kullanılabilir. Bu tasarımın bir sınırı, ortak doğum öncesi ortamı ve nispeten düşük sayıda ikizlerin ayrı yetiştirilmesidir. İkinci ve daha yaygın bir tasarım, ikiz çalışma kalıtımı tahmin etmek için özdeş ve çift yumurta ikizlerinin benzerliğinin kullanıldığı. Bu çalışmalar, tek yumurta ikizlerinin tamamen genetik olarak aynı değil, potansiyel olarak kalıtımın eksik tahmin edilmesine neden olur.
İçinde Gözlemsel çalışmalar veya çağrıştırıcı etkiler nedeniyle (bir genom, üzerlerindeki etkisiyle ortamları çağrıştırdığında), G ve E şu şekilde değişebilir: gen ortamı korelasyonu. Kalıtımı tahmin etmek için kullanılan yöntemlere bağlı olarak, genetik faktörler ile paylaşılan veya paylaşılmayan ortamlar arasındaki korelasyonlar, kalıtımla karıştırılabilir veya karıştırılmayabilir.[18]
Regresyon / korelasyon tahmin yöntemleri
İlk tahmin okulu, kalıtsallığı tahmin etmek için regresyon ve korelasyonu kullanır.
Yakın akrabaların karşılaştırılması
Akrabaların karşılaştırmasında genel olarak şunu görüyoruz:

nerede r olarak düşünülebilir ilişki katsayısı, b regresyon katsayısı ve t korelasyon katsayısıdır.
Ebeveyn-çocuk regresyonu

Kalıtım, ebeveyn ve yavru özellikleri karşılaştırılarak tahmin edilebilir (Şekil 2'deki gibi). Çizginin eğimi (0.57), yavru değerleri ebeveynlerdeki ortalama özelliğe göre gerilediğinde, özelliğin kalıtsallığına yaklaşır. Yalnızca bir ebeveynin değeri kullanılırsa, kalıtılabilirlik eğimin iki katıdır. (Bunun "terimin kaynağı olduğunu unutmayın"gerileme, "çünkü yavru değerleri her zaman ortalamaya gerilemek nüfus için değer, yanieğim her zaman birden azdır). Bu regresyon etkisi aynı zamanda DeFries – Fulker yöntemi etkilenen bir üye için seçilen ikizleri analiz etmek için.[19]
Kardeş karşılaştırması
Tam Sib tasarımları kullanılarak kalıtımsallığa temel bir yaklaşım benimsenebilir: Hem biyolojik bir anne hem de babayı paylaşan kardeşler arasındaki benzerliğin karşılaştırılması.[20] Yalnızca ilave gen eylemi olduğunda, bu kardeş fenotipik korelasyon, aşinalık - ilave genetik varyansın yarısı artı ortak ortamın tam etkisinin toplamı. Böylelikle, ek kalıtılabilirliğe tam Sib fenotipik korelasyonunun iki katı kadar bir üst sınır koyar. Yarı Sib tasarımları, bir ebeveyni diğer kardeş gruplarıyla paylaşan kardeşlerin fenotipik özelliklerini karşılaştırır.
İkiz çalışmaları
İnsanlardaki özelliklerin kalıtım derecesi en çok ikizler arasındaki benzerlikler karşılaştırılarak tahmin edilir. "İkiz çalışmaların avantajı, toplam varyansın genetik, paylaşılan veya ortak çevresel ve benzersiz çevresel bileşenlere bölünerek kalıtımın doğru bir tahminini mümkün kılmasıdır".[21] Kardeş veya dizigotik (DZ) ikizler, ortalama olarak genlerinin yarısını paylaşırlar ( çeşitli çiftleşme özellik için) ve çok özdeş veya monozigotik (MZ) ikizler, genetik olarak DZ ikizlerinden iki kat daha benzerdir. Öyleyse, kalıtımın kaba bir tahmini, aşağıdaki farkın yaklaşık iki katıdır ilişki MZ ve DZ ikizleri arasında, yani Falconer'ın formülü H2= 2 (r (MZ) -r (DZ)).
Paylaşılan ortamın etkisi, c2, yetiştirildikleri ortamın ortaklığı nedeniyle kardeşler arasındaki benzerliğe katkıda bulunur. Paylaşılan çevre, DZ ikizlerinin aynı genleri paylaşma derecesi olan yarı kalıtsallık eksi DZ korelasyonu ile yaklaşık olarak hesaplanır, c2= DZ-1/2h2. Benzersiz çevresel varyans, e2, birlikte yetiştirilen tek yumurta ikizlerinin farklı olma derecesini yansıtır, e2= 1-r (MZ).
Varyans tahmin yöntemlerinin analizi
İkinci kalıtım tahmin metotları grubu ANOVA ve varyans bileşenlerinin tahminini içerir.
Temel model
Kempthorne'un temel tartışmasını kullanıyoruz.[14] Sadece en temel genetik modelleri göz önünde bulundurarak, tek bir lokusun genotipli nicel katkısına bakabiliriz. Gben gibi

nerede genotipin etkisidir Gben ve çevresel etkidir.


Bir grup babayla ve onların soylarıyla rastgele barajlardan gelen bir deney düşünün. Soy genlerinin yarısını babadan ve yarısını (rastgele) annelerinden aldığından, soy denklemi

Sınıf içi korelasyonlar
Yukarıdaki deneyi düşünün. Karşılaştırabileceğimiz iki soy grubumuz var. Birincisi, tek bir baba için çeşitli soyları karşılaştırmaktır ( efendi grubu içinde). Varyans, genetik varyans için terimleri (hepsi aynı genotipe sahip olmadıkları için) ve çevresel varyansı içerecektir. Bu bir hata terim.
İkinci nesil grubu, yarı kardeşlerin ortalamalarının birbirleriyle karşılaştırılmasıdır ( efendi grubu arasında). Buna ek olarak hata terimi baba gruplarında olduğu gibi, yarım kardeşlerin farklı vasıtaları arasındaki farklılıklar nedeniyle ek bir terimimiz var. Sınıf içi korelasyon
- ,

çünkü çevresel etkiler birbirinden bağımsızdır.
ANOVA
İle bir deneyde beyler ve baba başına döl, aşağıdaki ANOVA'yı kullanarak hesaplayabiliriz genetik varyans olarak ve çevresel varyans olarak:




| Kaynak | d.f. | Ortalama Kare | Beklenen Ortalama Kare |
|---|---|---|---|
| Baba grupları arasında | |||
| Baba grupları içinde |






terim sınıf içi korelasyon üvey kardeşler arasında. Kolayca hesaplayabiliriz . Beklenen Ortalama Kare, bireylerin ilişkilerinden (örneğin, bir baba içindeki soyun hepsi yarı kardeştir) ve sınıf içi korelasyonların anlaşılmasından hesaplanır.


Kalıtılabilirliği hesaplamak için ANOVA'nın kullanılması, genellikle gen-çevre etkileşimleri, çünkü ANOVA çok daha düşük istatistiksel güç doğrudan etkiler yerine etkileşim etkilerini test etmek için.[22]
Katkı ve baskınlık terimleriyle model
Toplamsal ve baskınlık terimlerine sahip ancak diğerlerine sahip olmayan bir model için, tek bir lokus için denklem

nerede
i'nin toplamsal etkisidirinci alel j'nin toplamsal etkisidirinci alel ij için hakimiyet sapmasıinci genotip ve çevre.


Deneyler, Tablo 1'de verilene benzer bir kurulumla çalıştırılabilir. Farklı ilişki grupları kullanarak, farklı sınıf içi korelasyonları değerlendirebiliriz. Kullanma ilave genetik varyans olarak ve baskınlık sapma varyansı olarak, sınıf içi korelasyonlar doğrusal fonksiyonlar Bu parametrelerin. Genel olarak,


- Sınıf içi korelasyon

nerede ve olarak bulunur

P [ aleller ilişki çiftinden rastgele çizilmiş iniş ile aynı ], ve

P [ genotipler ilişki çiftinden rastgele çizilmiş iniş ile aynı ].

Bazı ortak ilişkiler ve katsayıları Tablo 2'de verilmiştir.
| İlişki | ||
|---|---|---|
| Tek yumurta ikizi | ||
| Ebeveyn-Çocuk | ||
| Yarım Kardeşler | ||
| Tam Kardeşler | ||
| İlk kuzenler | ||
| Çifte Birinci Kuzenler |






Doğrusal karışık modeller
Literatürde doğrusal karma modelleri kullanan çok çeşitli yaklaşımlar bildirilmiştir. Bu yöntemler aracılığıyla fenotipik varyans, kalıtsallığı tahmin etmek için genetik, çevresel ve deneysel tasarım varyanslarına bölünür. Çevresel varyans, bireyleri geniş bir çevre yelpazesinde inceleyerek açık bir şekilde modellenebilir, ancak fenotipik ve çevresel varyanstan genetik varyansın çıkarımı, bir özelliği etkileyen çevresel etkinin tüm aralığını yakalama zorluğundan dolayı kalıtımın eksik tahmin edilmesine yol açabilir. Kalıtımın hesaplanması için diğer yöntemler, genom çapında ilişkilendirme çalışmaları bir özellik üzerindeki etkiyi genetik faktörlerle tahmin etmek, bu, varsayılan olarak ilişkili genetik lokusların hızı ve etkisiyle yansıtılır (genellikle tek nükleotid polimorfizmleri ) özellik üzerinde. Ancak bu, kalıtımın hafife alınmasına yol açabilir. Bu tutarsızlık "eksik kalıtsallık" olarak adlandırılır ve kalıtım modellerindeki hem genetik hem de çevresel varyansı doğru bir şekilde modellemenin zorluğunu yansıtır.[23]
Büyük, karmaşık bir soy ağacı veya yukarıda belirtilen başka bir veri türü mevcut olduğunda, kalıtılabilirlik ve diğer nicel genetik parametreler aşağıdaki yöntemlerle tahmin edilebilir: sınırlı maksimum olasılık (REML) veya Bayesci yöntemler. işlenmemiş veri genellikle her birey için üç veya daha fazla veri noktasına sahip olacaktır: baba için bir kod, anne için bir kod ve bir veya birkaç özellik değeri. Farklı özellik değerleri, farklı özellikler veya farklı ölçüm zaman noktaları için olabilir.
Şu anda popüler olan metodoloji, baba ve barajın kimlikleri konusunda yüksek derecede kesinliğe dayanmaktadır; baba kimliğini olasılıkla ele almak yaygın değildir. Metodoloji yabani popülasyonlara nadiren uygulandığından (birkaç yabani toynaklı ve kuş popülasyonu için kullanılmış olmasına rağmen) ve babalar her zaman üreme programlarında çok yüksek derecede kesinlik ile bilindiğinden, bu genellikle bir sorun değildir. Ayrıca belirsiz babalığı açıklayan algoritmalar da vardır.
Soy ağacı, Pedigree Viewer gibi programlar kullanılarak görüntülenebilir. [1] gibi programlarla analiz edildi ve ASReml, VCE [2], WOMBAT [3], R ortamında MCMCglmm [4] ya da BLUPF90 programlar ailesi [5].
Soy ağacı modelleri, aşağıdaki gibi karışıklıkları çözmek için faydalıdır: Ters nedensellik, anne etkileri benzeri doğum öncesi ortam ve kafa karıştırıcı genetik hakimiyet, paylaşılan ortam ve maternal gen etkileri.[24][9]
Genomik kalıtım
Genom çapında genotip verileri ve büyük popülasyon örneklerinden fenotipler mevcut olduğunda, bireyler arasındaki ilişkileri genotiplerine göre tahmin edebilir ve genetik belirteçler tarafından açıklanan varyansı tahmin etmek için doğrusal bir karma model kullanılabilir. Bu, ortak genetik varyantlar tarafından yakalanan varyansa dayalı bir genomik kalıtım tahmini verir.[4] Alel frekansı için farklı ayarlamalar yapan birden fazla yöntem vardır ve Bağlantı dengesizliği. Özellikle, Yüksek Çözünürlüklü Olabilirlik (HDL) olarak adlandırılan yöntem, genomik kalıtımı yalnızca GWAS özet istatistiklerini kullanarak tahmin edebilir,[5] Bu, çeşitli GWAS meta analizlerinde mevcut olan büyük örneklem boyutunun dahil edilmesini kolaylaştırır.
Seçime yanıt
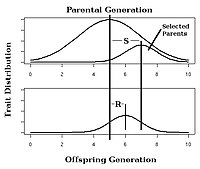
İçinde seçici yetiştirme bitki ve hayvanlarda, bilinen dar anlamda kalıtsallığa sahip bir özelliğin seçimine beklenen yanıt kullanılarak tahmin edilebilir yetiştiricinin denklemi:[25]


Bu denklemde, Seçime Yanıt (R), ebeveyn nesil ile sonraki nesil arasındaki gerçekleşen ortalama fark olarak tanımlanır ve Seçim Farklılığı (S), ebeveyn nesil ile seçilen ebeveyn arasındaki ortalama fark olarak tanımlanır.[14]:1957[26]
Örneğin, bir bitki yetiştiricisinin, mısır başağı başına çekirdek sayısını artırmak amacıyla seçici bir yetiştirme projesine dahil olduğunu hayal edin. Tartışma adına, ebeveyn nesildeki ortalama mısır başağının 100 tane çekirdeğe sahip olduğunu varsayalım. Ayrıca seçilen ebeveynlerin kulak başına ortalama 120 tane mısır ürettiklerini de varsayalım. Eğer h2 0.5'e eşitse, sonraki nesil ortalama 0.5 (120-100) = kulak başına 10 ek dane ile mısır üretecektir. Bu nedenle, mısır başağı başına düşen toplam tane sayısı ortalama 110'a eşit olacaktır.
Yapay bir seçim deneyinde seçime verilen yanıtı gözlemlemek, Şekil 4'teki gibi gerçekleşen kalıtımın hesaplanmasına izin verecektir.
Yukarıdaki denklemdeki kalıtsallığın orana eşit olduğuna dikkat edin sadece genotip ve çevresel gürültü takip ederse Gauss dağılımları.

Tartışmalar
 | Bu bölüm olabilir dengesiz belirli bakış açılarına doğru. (Ağustos 2016) |
Kalıtılabilirlik tahminlerinin öne çıkan eleştirmenleri, örneğin Steven Rose,[27] Jay Joseph,[28] ve Richard Bentall, büyük ölçüde kalıtım tahminlerine odaklanın Davranış bilimleri ve sosyal Bilimler. Bentall, bu tür kalıtsallık puanlarının tipik olarak sayısal olarak yüksek puanlar elde etmek için mantıksız olarak hesaplandığını, kalıtımın şu şekilde yanlış yorumlandığını iddia etmiştir: genetik belirleme ve bu iddia edilen önyargı, daha sonra psikoza neden olan çocukluk istismarı gibi araştırmaların nedensel olarak daha önemli bulduğu diğer faktörlerden uzaklaştırır.[29][30] Kalıtılabilirlik tahminleri de doğası gereği sınırlıdır çünkü genlerin veya çevrenin, incelenen özelliğin gelişiminde daha büyük bir rol oynayıp oynamadığına ilişkin herhangi bir bilgi vermezler. Bu yüzden, David Moore ve David Shenk Davranış genetiği bağlamında "kalıtsallık" terimini "... bilim tarihindeki en yanıltıcılardan biri" olarak tanımlar ve çok nadir durumlar dışında hiçbir değeri olmadığını savunur.[31] Karmaşık insan özelliklerini incelerken, genlerin ve çevrenin göreceli katkılarını belirlemek için kalıtsallık analizini kullanmak imkansızdır, çünkü bu özellikler birbiriyle etkileşim halinde olan birden fazla nedenden kaynaklanır.[32] Bununla birlikte, bazı araştırmacılar bu ikisini çözmenin mümkün olduğunu iddia ediyor.[33]
Kalıtım tahminlerine ilişkin tartışma, büyük ölçüde ikiz çalışmalar. Kıt başarı moleküler genetik bunu doğrulayan çalışmalar popülasyon-genetik çalışmaların sonuçları kalıtımın olmaması sorun.[34] Eric Turkheimer, daha yeni moleküler yöntemlerin ikiz çalışmalarının geleneksel yorumunu doğruladığını iddia etti:[34] Genler ve davranışlar arasındaki ilişkilerin nasıl açıklanacağı çoğunlukla belirsiz olsa da.[35] Turkheimer'a göre, hem genler hem de çevre kalıtsaldır, genetik katkı çevreye göre değişir ve kalıtılabilirliğe odaklanma, diğer önemli faktörlerden uzaklaşır.[36] Ancak genel olarak kalıtım yaygın olarak uygulanabilen bir kavramdır.[9]
Ayrıca bakınız
Referanslar
- ^ Wray N, Visscher P (2008). "Kalıtım Özelliklerinin Tahmin Edilmesi". Doğa Eğitimi. 1 (1): 29. Arşivlendi 2 Ağustos 2015 tarihinde orjinalinden. Alındı 24 Temmuz 2015.
- ^ Gazzaniga MS, Heatherton TF, Halpern DF. Psikolojik bilim (5. baskı). New York. ISBN 978-0-393-26313-8. OCLC 908409996.
- ^ Yang J, Lee SH, Goddard ME, Visscher PM (Ocak 2011). "GCTA: genom çapında karmaşık özellik analizi için bir araç". Amerikan İnsan Genetiği Dergisi. 88 (1): 76–82. doi:10.1016 / j.ajhg.2010.11.011. PMC 3014363. PMID 21167468.
- ^ a b Yang J, Zeng J, Goddard ME, Wray NR, Visscher PM (Ağustos 2017). "SNP'ye dayalı kalıtımın kavramları, tahmini ve yorumu" (PDF). Doğa Genetiği. 49 (9): 1304–1310. doi:10.1038 / ng.3941. PMID 28854176. S2CID 8790524. Arşivlendi (PDF) 2020-10-05 tarihinde orjinalinden. Alındı 2020-09-06.
- ^ a b Ning Z, Pawitan Y, Shen X (Haziran 2020). "İnsan karmaşık özellikleri arasındaki genetik korelasyonların yüksek tanımlı olasılık çıkarımı". Doğa Genetiği. 52 (8): 859–864. doi:10.1038 / s41588-020-0653-y. hdl:10616/47311. PMID 32601477. S2CID 220260262.
- ^ Stoltenberg SF (Haziran 1997). "Kalıtımla uzlaşmak". Genetica. 99 (2–3): 89–96. doi:10.1007 / BF02259512. hdl:2027.42/42804. PMID 9463077. S2CID 18212219.
- ^ Wahlsten D (1994). "Kalıtımın zekası" (PDF). Kanada Psikolojisi. 35 (3): 244–260. doi:10.1037/0708-5591.35.3.244. ISSN 1878-7304. Arşivlendi (PDF) 2018-10-24 tarihinde orjinalinden. Alındı 2019-12-05.
- ^ Maccoby EE (Şubat 2000). "Ebeveynlik ve çocuklar üzerindeki etkileri: okuma ve davranış genetiğini yanlış okuma üzerine". Yıllık Psikoloji İncelemesi. 51 (1): 1–27. doi:10.1146 / annurev.psych.51.1.1. PMID 10751963.
- ^ a b c Visscher PM, Hill WG, Wray NR (Nisan 2008). "Genomik çağında kalıtılabilirlik - kavramlar ve yanlış kanılar" (PDF). Doğa Yorumları. Genetik. 9 (4): 255–66. doi:10.1038 / nrg2322. PMID 18319743. S2CID 690431. Arşivlendi (PDF) 2016-03-24 tarihinde orjinalinden. Alındı 2015-08-28.
- ^ Sos B, Matzel LD (Ocak 2018). "Zeka paradoksu: Kalıtım ve şekillendirilebilirlik, gizli gen-çevre etkileşiminde bir arada var olur". Psikolojik Bülten. 144 (1): 26–47. doi:10.1037 / bul0000131. PMC 5754247. PMID 29083200.
- ^ Blok N (Ağustos 1995). "Kalıtımın ırk konusunda nasıl yanıltıcı olduğu". Biliş. 56 (2): 99–128. doi:10.1016 / 0010-0277 (95) 00678-r. PMID 7554794. S2CID 204981536.
- ^ Wills C (2007). "Popülasyon Genetiğinin İlkeleri, 4. baskı" (PDF). Kalıtım Dergisi (Kitap incelemesi). 98 (4): 382. doi:10.1093 / jhered / esm035. Arşivlendi (PDF) 2020-10-05 tarihinde orjinalinden. Alındı 2018-04-20.
- incelemesi: Hartl DL, Clark AG (2007). Popülasyon Genetiğinin İlkeleri. Sunderland, MA: Sinauer and Associates. s. xv + 652. ISBN 978-0-87893-308-2.
- ^ Turkheimer E (Ekim 2000). "Davranış Genetiğinin Üç Yasası ve Ne Anlama Geldikleri" (PDF). Psikolojik Bilimde Güncel Yönler. 9 (5): 160–164. doi:10.1111/1467-8721.00084. ISSN 0963-7214. S2CID 2861437. Arşivlendi (PDF) 19 Ekim 2013 tarihinde orjinalinden. Alındı 29 Ekim 2013.
- ^ a b c Kempthorne O (1957). Genetik istatistiklere giriş (1. baskı). Ames, Iowa: Iowa Eyalet Üniv. Basın. OCLC 422371269.
- ^ Stephen Downes ve Lucas Matthews. "Kalıtım". Stanford Felsefe Ansiklopedisi. Stanford Üniversitesi. Arşivlendi 2020-02-25 tarihinde orjinalinden. Alındı 2020-02-20.
- ^ Zuk O, Hechter E, Sunyaev SR, Lander ES (Ocak 2012). "Kalıtımın kaybolmasının gizemi: Genetik etkileşimler hayali kalıtım yaratır". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 109 (4): 1193–8. doi:10.1073 / pnas.1119675109. PMC 3268279. PMID 22223662.
- ^ Daw J, Guo G, Harris KM (Temmuz 2015). "Doğanın beslenmesi: Akademik başarıda ve sözel zekada paylaşılan ortamların rolünün yeniden değerlendirilmesi". Sosyal Bilimler Araştırması. 52: 422–39. doi:10.1016 / j.ssresearch.2015.02.011. PMC 4888873. PMID 26004471.
- ^ Cattell RB (Kasım 1960). "Çoklu soyut varyans analizi denklemleri ve çözümleri: sürekli değişkenler üzerinde doğa-yetiştirme araştırması için". Psikolojik İnceleme. 67 (6): 353–72. doi:10.1037 / h0043487. PMID 13691636.
- ^ DeFries JC, Fulker DW (Eylül 1985). "İkiz verilerin çoklu regresyon analizi". Davranış Genetiği. 15 (5): 467–73. doi:10.1007 / BF01066239. PMID 4074272. S2CID 1172312.
- ^ Falconer DS, Mackay TF (Aralık 1995). Kantitatif Genetiğe Giriş (4. baskı). uzun adam. ISBN 978-0582243026.
- ^ Gielen M, Lindsey PJ, Derom C, Smeets HJ, Souren NY, Paulussen AD, Derom R, Nijhuis JG (Ocak 2008). "Kalıtımı artırmak ve doğum ağırlığı için aday genlerin tanımlanmasını kolaylaştırmak için genetik ve çevresel faktörlerin modellenmesi: bir ikiz çalışması". Davranış Genetiği. 38 (1): 44–54. doi:10.1007 / s10519-007-9170-3. PMC 2226023. PMID 18157630.
- ^ Wahlsten, Douglas (Mart 1990). "Varyans analizinin kalıtım-çevre etkileşimine duyarsızlığı" (PDF). Davranış ve Beyin Bilimleri. 13 (1): 109–120. doi:10.1017 / S0140525X00077797. ISSN 1469-1825.
- ^ Heckerman D, Gurdasani D, Kadie C, Pomilla C, Carstensen T, Martin H, Ekoru K, Nsubuga RN, Ssenyomo G, Kamali A, Kaleebu P, Widmer C, Sandhu MS (Temmuz 2016). "Çevresel çeşitliliği açıkça ele alan kalıtsallık tahmini için doğrusal karma model". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 113 (27): 7377–82. doi:10.1073 / pnas.1510497113. PMC 4941438. PMID 27382152.
- ^ Hill WG, Goddard ME, Visscher PM (Şubat 2008). MacKay TF, Goddard ME (editörler). "Veriler ve teori, esas olarak karmaşık özellikler için ilave genetik varyansa işaret ediyor". PLOS Genetiği. 4 (2): e1000008. doi:10.1371 / journal.pgen.1000008. PMC 2265475. PMID 18454194.

- ^ Plomin R, DeFries JC, McClearn GE, McGuffin P (2017). Davranışsal Genetik: Bir Başlangıç (2. baskı). New York: W.H. Özgür adam. ISBN 978-0-7167-2056-0.
- ^ Falconer DS, Mackay TF (1998). Kantitatif genetiğe giriş (4. baskı). Essex: Longman. ISBN 978-0-582-24302-6.
- ^ Gül SP (Haziran 2006). "Yorum: kalıtım tahminleri - son satış tarihleri çoktan geçmiş". Uluslararası Epidemiyoloji Dergisi. 35 (3): 525–7. doi:10.1093 / ije / dyl064. PMID 16645027.
- ^ Joseph J (2004). "Bölüm 5". Gen Yanılsaması. New York: Algora. s. 141. ISBN 978-1-898059-47-9. Arşivlendi 2017-07-19 tarihinde orjinalinden. Alındı 2016-04-02.
- ^ Bentall RP (2009). Zihin Doktoru: Mevcut Akıl Hastalığına Uyguladığımız Tedavimiz Gerçekten Yararlı mı?. New York: New York University Press. s. 123–127. ISBN 978-0-8147-8723-6. Arşivlendi 2020-10-05 tarihinde orjinalinden. Alındı 2016-04-02.
- ^ McGrath M (5 Temmuz 2009). "Doctoring the Mind: Review". Telgraf. Arşivlendi 4 Temmuz 2012 tarihinde orjinalinden. Alındı 4 Nisan 2018.
- ^ Moore DS, Shenk D (Ocak 2017). "Kalıtım yanılgısı". Wiley Disiplinlerarası İncelemeler: Bilişsel Bilimler. 8 (1–2): e1400. doi:10.1002 / wcs.1400. PMID 27906501.
- ^ Feldman MW, Ramachandran S (Nisan 2018). "Neye kıyasla eksik? Kalıtılabilirliği, genleri ve kültürü yeniden ziyaret etmek". Londra Kraliyet Cemiyeti'nin Felsefi İşlemleri. Seri B, Biyolojik Bilimler. 373 (1743): 20170064. doi:10.1098 / rstb.2017.0064. PMC 5812976. PMID 29440529.
... tüm karmaşık insan özellikleri, çeşitli nedenlerin birleşiminden kaynaklanır. Bu nedenler etkileşime girerse, bir özelliğin her birine bağlı olarak niceliksel değerleri atamak imkansızdır, tıpkı bir dikdörtgenin alanının ne kadarının iki boyutunun her birine ayrı ayrı bağlı olduğunu söyleyemeyeceğimiz gibi. Bu nedenle, karmaşık insan fenotiplerinin analizinde ... aslında "fenotipin belirlenmesinde genlerin ve çevrenin göreceli önemini" bulamayız.
- ^ Tredoux, Gavan. "Dikdörtgenlerin Doğası ve Doğası." (2019).
- ^ a b Turkheimer E (2011). "Hala kayıp". İnsani Gelişmede Araştırma. 8 (3–4): 227–241. doi:10.1080/15427609.2011.625321. S2CID 14737438.
- ^ Turkheimer E (2015). "Genetik Tahmin". Hastings Center Raporu. 45 (5 Ek): S32–8. doi:10.1002 / hast.496. PMID 26413946.
- ^ Joseph J (2014). İkiz Çalışmalarıyla İlgili Sorun: Sosyal ve Davranış Bilimlerinde İkiz Araştırmalarının Yeniden Değerlendirilmesi (PDF). New York: Routledge. s. 81. ISBN 978-1-317-60590-4. Arşivlendi (PDF) 2016-04-04 tarihinde orjinalinden. Alındı 2016-04-02.
daha fazla okuma
- Lynch M, Walsh B (1998). Kantitatif özelliklerin genetiği ve analizi. Sunderland, Mass .: Sinauer Doç. ISBN 978-0-87893-481-2.
- Johnson W, Penke L, Spinath FM (2011). "Kalıtımı Anlamak: Nedir ve Ne Değildir". Avrupa Kişilik Dergisi. 25 (4): 287–294. doi:10.1002 / başına.835. ISSN 0890-2070.
Dış bağlantılar
- Stanford Encyclopedia of Philosophy girişi Kalıtım ve Kalıtım üzerine
- Lynch ve Walsh'un iki ciltlik kitabını içeren Quantitative Genetics Resources web sitesi. Serbest erişim
| Nicel Genetikte Kavramlar | |
|---|---|
| İlgili konular | |
| |
| Lisenkoistler | |
|---|---|
| Muhalifler | |
| Kavramlar | |
| Sovyet politikaları | |