Stegoceras - Stegoceras
| Stegoceras | |
|---|---|
 | |
| Yeniden inşa edilmiş iki S. validum UALVP 2 örneğine dayalı iskeletler, Royal Tyrrell Müzesi | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Sipariş: | †Ornithischia |
| Aile: | †Pachycephalosauridae |
| Cins: | †Stegoceras Lambe, 1902 |
| Türler | |
| †Stegoceras validum Lambe, 1902 | |
| Türler | |
| |
| Eş anlamlı | |
Liste
| |
Stegoceras bir cins nın-nin pachycephalosaurid (kubbe başlı) Dinozor şimdi ne yaşadı Kuzey Amerika esnasında Geç Kretase dönem, yaklaşık 77,5 ila 74 milyon yıl önce (mya). İlk örnekler Alberta, Kanada, 1902'de tanımlandı ve türler Stegoceras validum bu kalıntılara dayanıyordu. Genel ad "boynuz çatı" anlamına gelir ve özel ad "güçlü" anlamına gelir. Yıllar içinde bu cinse birkaç başka tür de yerleştirilmiştir, ancak bunlar o zamandan beri başka cinslere taşınmış veya küçük eş anlamlılar. Şu anda sadece S. validum ve S. novomexicanum, 2011 yılında bulunan fosillerden seçildi Yeni Meksika, kalmak. Son türlerin geçerliliği de tartışılmaktadır.
Stegoceras küçüktü iki ayaklı yaklaşık 2 ila 2,5 metre (6,6 ila 8,2 ft) uzunluğunda ve yaklaşık 10 ila 40 kilogram (22 ila 88 lb) ağırlığındaydı. Sert bir Omurga ve sertleşmiş bir kuyruk. Pelvik bölge, muhtemelen genişletilmiş bir bağırsak nedeniyle genişti. Kafatası kabaca üçgen şeklindeydi ve kısa bir burnu vardı ve tepesinde kalın, geniş ve nispeten pürüzsüz bir kubbe vardı. Kafatasının arkasında kalın bir "raf" vardı. oksiput ve gözlerin üzerinde kalın bir çıkıntı vardı. Kafatasının çoğu tüberküller (veya yuvarlak "büyümeler") ve düğümler (veya "topuzlar"), çoğu sıra halinde ve en büyüğü rafta oluşturulmuş küçük boynuzlar. Dişler küçük ve tırtıklıydı. Kafatasının genç hayvanlarda düz olduğu ve yaşla birlikte bir kubbe haline geldiği düşünülmektedir.
Başlangıçta sadece kafatası kubbelerinden biliniyor, Stegoceras bilinen ilk pachycephalosaur'lardan biriydi ve bu ilk kalıntıların eksik olması, bu grubun yakınlıkları hakkında birçok teoriye yol açtı. Tam Stegoceras iskeletin ilişkili parçalarıyla birlikte kafatası 1924'te bulundu ve bu hayvanlara daha fazla ışık tuttu. Pachycephalosaurlar bugün boynuzlu Ceratopsia'lılar grupta Marginocephalia. Stegoceras kendisi düşünüldü baz alınan (veya "ilkel") diğer pachycephalosaurlarla karşılaştırıldığında. Stegoceras büyük ihtimalle otoburdu ve muhtemelen iyi bir koku alma duyusuna sahipti. Kubbenin işlevi tartışıldı ve rakip teoriler arasında özel iç savaş (kafa veya yan-dikme), cinsel görüntü veya tür tanıma. S. validum dan bilinmektedir Dinozor Parkı Oluşumu ve Oldman Formasyonu, buna karşılık S. novomexicanum dan Fruitland ve Kirtland Formasyonu.
Keşif tarihi

Bilinen ilk kalıntıları Stegoceras Kanadalı tarafından toplandı paleontolog Lawrence Lambe -den Belly River Grubu, içinde Red Deer Nehri Alberta bölgesi Kanada. Bu kalıntılar iki kısmi kafatası kubbesinden oluşuyordu (örnek CMN 515 ve CMN 1423 Kanada Doğa Müzesi ) 1898'de toplanan farklı boyutlardaki iki hayvandan ve 1901'de toplanan üçüncü bir kısmi kubbeden (CMN 1594). Lambe, bu örneklere dayanarak, yeniyi tanımladı ve adlandırdı. tek tip cins ve türler Stegoceras validus içinde 1902.[1][2] Genel isim Stegoceras dan geliyor Yunan stegè/ στέγη, "çatı" anlamına gelir ve Keras/ κέρας "boynuz" anlamına gelir. Özel isim validus Latince'de muhtemelen kalın kafatası çatısına atıfta bulunulduğunda "güçlü" anlamına gelir.[3] Çünkü türler birden fazla örneğe dayanıyordu (a syntype serisi ), CMN 515, ders türü örneği tarafından John Bell Hatcher 1907'de.[4][2]
Daha önce bölgede benzer kalıntılar bulunmadığından, Lambe onların ne tür bir dinozor olduklarından ve bir veya daha fazla türü temsil edip etmediklerinden emin değildi; kubbelerin "prenasal" olduğunu ileri sürdü. burun kemikleri başın orta hattında ve burun boynuzuna benzerliklerini kaydetti. Triceratops örnek.[1] 1903'te Macar paleontolog Franz Nopcsa von Felső-Szilvás parçalı kubbelerin Stegoceras aslında ön ve nazal kemiklerdi ve bu nedenle hayvanın tek, eşleşmemiş bir boynuzu olacaktı. Lambe, Nopscsa'nın makalesinin 1903 tarihli incelemesinde yeni bir tür "tek boynuzlu dinozor" fikrine sempati duyuyordu. Şu anda, hangi dinozor grubu hakkında hala belirsizlik vardı. Stegoceras ikisine de aitti Ceratopsia'lılar (boynuzlu dinozorlar) ve stegosaurlar (kaplanmış dinozorlar) yarışmacı olarak.[5][6] Hatcher şüpheli Stegoceras örnekler aynı türe ait olup dinozor olup olmadıkları ve kubbelerin kafatasının ön, oksipital ve paryetal kemiklerinden oluştuğunu öne sürüyordu.[2] 1918'de Lambe, başka bir kubbeyi (CMN 138) S. validusve yeni bir türe isim verdi, S. brevis, CMN 1423 örneğine göre (başlangıçta dahil ettiği S. validus). Bu zamana kadar, bu hayvanları Stegosauria'nın üyeleri olarak gördü (daha sonra her iki zırhlı dinozor ailesinden oluşuyordu, Stegosauridae ve Ankylosauridae ), Psalisauridae adını verdiği yeni bir ailede (adı tonozlu veya kubbe şeklindeki kafatası çatısı).[7]

1924'te Amerikalı paleontolog Charles W. Gilmore tam bir kafatasını tanımladı S. validus ilişkili postkraniyal kalıntılarla (UALVP 2 örneği Alberta Üniversitesi Omurgalı Paleontolojisi Laboratuvarı), o zamana kadar kubbe başlı bir dinozorun en eksiksiz kalıntıları. Bu bulgu, Hatcher'ın kubbelerin kafatasının frontoparietal bölgesinden oluştuğu şeklindeki yorumunu doğruladı. UALVP 2, küçük, dezartiküle kemik elementlerle bulundu, daha sonra gastralya diğerlerinde bilinmeyen (abdominal kaburga) ornitorik dinozorlar (iki ana dinozor grubundan biri). Gilmore, dişlerin S. validus türlerinkine çok benziyordu Troodon formozusu (1856'da adlandırıldı ve o zamana kadar sadece izole dişlerden biliniyordu) ve bulunduğu yerin yakınında bulunan bir kafatası kubbesini tanımladı. Troodon bulundu. Bu nedenle Gilmore, Stegoceras geçersiz küçük eşanlamlı nın-nin Troodon, dolayısıyla yeniden adlandırmak S. validus içine T. validusve iki türün bile aynı olabileceğini öne sürdü. Dahası, buldu S. brevis özdeş olmak S. validusve bu nedenle ikincisinin küçük eşanlamlısı. Ayrıca bu türleri yeni aileye yerleştirdi. Troodontidae (Lambe, Psalisauridae için bir tür cins seçmediği için) ornitopod dinozorlar.[8][9] Kafatası, "ilkel" görünümlü iskelete kıyasla çok özel göründüğünden, Nopcsa bu parçaların gerçekten birbirine ait olup olmadığından şüphe etti ve kafatasının bir kafatasına ait olduğunu öne sürdü. nodosaur, iskelet ornitopod ve sözde gastralia (karın kaburga) bir balıktır. Bu iddia 1930'larda Gilmore ve Loris S. Russell tarafından çürütüldü.[10]

Gilmore'un sınıflandırması Amerikalı paleontologlar tarafından desteklendi. Barnum Brown ve Erich Maren Schlaikjer o zamanlar 46 kafatasından bilinen kubbe başlı dinozorlar hakkındaki 1943 incelemelerinde. Brown ve Schlaikjer bu örneklerden yeni türlere isim verdi. T. sternbergi ve T. edmontonensis (hem Alberta'dan) hem de büyük türleri hareket ettirmek T. wyomingensis (1931'de adlandırıldı) yeni cinse Pachycephalosaurus, diğer iki türle birlikte. Onlar buldular T. validus farklı T. formosusama dikkate alındı S. brevis dişi formu T. validusve bu nedenle küçük bir eşanlamlı. Bu zamana kadar, kubbe başlı dinozorlar ya ornitopodların ya da ankilozorların akrabaları olarak görülüyordu.[10] 1945'te, T. formosus ve S. validus Amerikan paleontolog Charles M. Sternberg ikisi arasındaki farklılıkları gösterdi ve bunun yerine şunu önerdi: Troodon bir Theropod dinozor ve kubbe başlı dinozorların kendi ailelerine yerleştirilmesi gerektiğini söyledi. Rağmen Stegoceras bu ailenin isimlendirilen ilk üyesi oldu, Sternberg grubu seçti Pachycephalosauridae ikinci cinsten sonra, bu adı ("kalın başlı kertenkele" anlamına gelir) daha açıklayıcı bulmuştur. O da düşündü T. sternbergi ve T. edmontonensis üyeleri Stegoceras, bulundu S. brevis geçerli ve yeni bir tür olarak adlandırıldı, S. lambeiönceden atıfta bulunulan bir örneğe göre S. validus.[3][11] Ayrılık Troodon 1948'de Russell tarafından desteklendi ve dişleri neredeyse aynı olan bir theropod diş hekimini tanımladı. T. formosus.[12]
1953'te, Birger Bohlin isimli Troodon Bexelli Çin'den bir parietal kemiğe dayanmaktadır.[13] 1964'te, Oskar Kuhn bunu tartışmasız bir tür olarak kabul etti Stegoceras; S. bexelli.[14] 1974'te Polonyalı paleontologlar Teresa Maryańska ve Halszka Osmólska "gastralia" nın Stegoceras pachycephalosaur'un kuyruğundaki bu tür yapıları belirledikten sonra kemikleşmiş tendonlardı Homalocephale.[9] 1979'da William Patrick Wall ve Peter Galton yeni türlere isim verdi Stegoceras brownidaha önce dişi olarak tanımlanan düzleştirilmiş bir kubbeye dayanan S. validus 1971'de Galton tarafından. Özel isim, holotip örneği (örnek AMNH 5450, Amerikan Doğa Tarihi Müzesi ) Alberta'da.[15] 1983'te Galton ve Hans-Dieter Sues taşındı S. browni kendi cinsine, Ornatotholus (Ornatus Latince "süslü" anlamına gelir ve tholus "kubbe" için) ve daha önce Asya'da bilinen "düz başlı" pachycephalosaurlar grubunun bilinen ilk Amerikalı üyesi olarak kabul edildi.[16] 1987'de pachycephalosaurs üzerine yapılan bir incelemede, Sues ve Galton özel adı değiştirdiler. validus -e geçerli, daha sonra bilimsel literatürde kullanılmıştır. Bu yazarlar eş anlamlı S. brevis, S. sternbergi, ve S. lambei ile S. validum, bulundu S. bexelli farklı Stegoceras çeşitli özelliklerde ve belirsiz bir pachycephalosaur olarak kabul edildi.[3][4]

Goodwin ve arkadaşları 1998'de Ornatotholus bir çocuk S. validum, bu nedenle küçük bir eşanlamlı.[17] 2000 yılında Robert M. Sullivan, S. edmontonensis ve S. brevis cinse Prenocephale (o zamana kadar sadece Moğol türlerinden biliniyordu P. prenes) ve daha muhtemel buldum S. bexelli ye ait Prenocephale daha Stegocerasama bunu bir nomen dubium (karakterleri ayırt etmeksizin şüpheli isim) eksikliğinden dolayı ve holotip örneğinin kaybolmuş gibi göründüğünü kaydetti.[18] 2003 yılında Thomas E. Williamson ve Thomas Carr düşünülen Ornatotholus a nomen dubiumveya belki bir çocuk Stegoceras.[19] 2003 revizyonunda StegocerasSullivan bunu kabul etti Ornatotholus küçük eşanlamlıydı Stegoceras, taşındı S. lambei yeni cinse Colepiocephale, ve S. sternbergi -e Hanssuesia. Cins olduğunu belirtti Stegoceras olmuştu çöp sepeti taksonu o noktaya kadar küçük ve orta boy Kuzey Amerika pachycephalosaurları için. Bu zamana kadar düzinelerce örnek gönderilmişti. S. validumolarak tanımlanamayacak kadar eksik birçok alan dahil Stegoceras kesinlikle. UALVP 2, hala en eksiksiz örnektir. Stegoceras, cinsin en bilimsel anlayışının dayandığı.[4]
2002'de Williamson ve Carr bir kubbe tanımladılar (örnek NMMNH P-33983, New Mexico Doğa Tarihi ve Bilim Müzesi ) itibaren San Juan Havzası, Yeni Meksika belirsiz türlerin bir juvenil pachycephalosaur'u olarak kabul ettikleri (belki de Sphaerotholus goodwini). 2006'da Sullivan ve Spencer G.Lucas bunu çocuk olarak kabul etti S. validumtürlerin yelpazesini önemli ölçüde genişletecek.[20][21] İçinde 2011, Steven E. Jasinski ve Sullivan, örneği bir yetişkin olarak kabul ettiler ve onu yeni türlerin holotipi yaptılar. Stegoceras novomexicanum, diğer iki örnekle (SMP VP-2555 ve SMP VP-2790) paratip olarak.[22] Bir 2011 Filogenetik analiz Watabe ve meslektaşları tarafından ikisini yerleştirmedi Stegoceras birbirine yakın türler.[23] 2016 yılında Williamson ve Stephen L. Brusatte holotipini yeniden denedi S. novomexicanum ve paratiplerin holotip ile aynı taksona ait olmadığını ve ilgili tüm örneklerin yavrular olduğunu buldu. Dahası, holotip örneğinin farklı türleri temsil edip etmediğini belirleyemediler. S. novomexicanumya da çocuklardan biri ise S. validum veya Sphaerotholus goodwiniveya daha önce bilinen başka bir pachycephalosaur.[24] 2016 yılında Jasinski ve Sullivan, S. novomexicanum; türleri teşhis etmek için kullanılan bazı özelliklerin bir alt yetişkin aşamasının göstergesi olduğunu, ancak holotipte türleri ayırt eden ek teşhis özellikleri sunduğunu kabul ettiler. Ayrıca bazı yetişkin özelliklerine de işaret ettiler. heterokroni (zamanlamadaki fark ontogenetik türlerdeki ilgili taksonlar arasındaki değişiklikler). Paratiplerin ve diğer atanmış örneklerin, daha yüksek kubbeli kafataslarına sahip olmaları bakımından holotipten farklı olduğunu kabul ettiler, bunun yerine onlara cf. S. novomexicanum (tanımlanması zor), ancak kısıtlı olması nedeniyle hepsinin aynı taksona (atanan örnekler yetişkinler) ait olma olasılığını bulmuşlardır. stratigrafik aralık ve coğrafi aralık.[25]
Açıklama

Stegoceras Kuzey Amerika'nın en çok bilinen pachycephalosaurlarından biridir ve bilinen birkaç tanesinden biridir. postkraniyal kalıntılar; S. validum örnek UALVP 2 en eksiksiz olanıdır Stegoceras bugüne kadar bilinen kişi. Uzunluğunun yaklaşık 2 ila 2,5 metre (6,6 ila 8,2 ft) olduğu tahmin edilmektedir. keçi.[26][27][28] Ağırlığın yaklaşık 10 ila 40 kilogram (22 ila 88 lb) olduğu tahmin edilmektedir.[29] Stegoceras diğer pachycephalosaurlara kıyasla küçük ila orta büyüklükteydi.[3] S. novomexicanum daha küçük görünüyor S. validum, ancak bilinen örneklerin (eksik kafatasları) yetişkin mi yoksa genç mi olduğu tartışmalıdır.[22][24]
Omurga nın-nin Stegoceras tam olarak bilinmiyor. Arasındaki eklemlenme zigagofizler Arka arkaya gelen dorsal (arka) omurların (eklem süreçleri), vertebral kolonun yana doğru hareketini engellemiş gibi görünmektedir, bu da onu çok sert hale getirmiştir ve daha da güçlenmiştir. kemikleşmiş tendonlar.[3] Boyun omurları bilinmemekle birlikte, oksipital kondil (ilk boyun omuruyla ifade edilen), çoğu dinozor boynunun "S" veya "U" şeklinde olduğu gibi boynun kavisli bir duruşta tutulduğunu gösterir.[30] Konumlarına göre HomalocephaleUALVP 2 ile bulunan kemikleşmiş tendonlar karmaşık bir yapı oluşturmuş olabilir "kuyruk sepeti"kuyrukta, paralel sıralardan oluşan, her tendonun ekstremiteleri birbirini izleyen bir sonraki ile temas eder. Bu tür yapılar Myorhabdoive aksi halde yalnızca teleost balık; özellik, pachycephalosaurlara özgüdür. dört ayaklı (dört kollu) hayvanlar ve kuyruğu sertleştirme işlevi görmüş olabilir.[9]

kürek kemiği (kürek kemiği) daha uzundu humerus (üst kol kemiği); bıçağı ince ve dardı ve kaburgaların dış hatlarını izleyerek hafifçe bükülmüştü. Kürek kemiği üst uçta genişlemedi, ancak tabanda çok genişledi. korakoid çoğunlukla ince ve tabak gibiydi. Humerus ince bir şafta sahipti, uzunluğu boyunca hafifçe bükülmüş ve hafifçe eğilmişti. deltopektoral tepe (nerede deltoid ve pektoral kaslar) zayıf bir şekilde geliştirildi. Sonları ulna genişletilmiş ve şaft boyunca sırtlar uzanmıştır. yarıçap sıradışı olan ulnadan daha sağlamdı. Yukarıdan bakıldığında, pelvik kuşak için çok genişti iki ayaklı Archosaur ve arka kısma doğru genişledi. Pelvik bölgenin genişliği, bağırsağın arka uzantısını barındırmış olabilir. ilium uzamıştı ve ischium uzun ve inceydi. Rağmen kasık bilinmiyor, muhtemelen boyutu küçültüldü Homalocephale. uyluk (uyluk kemiği) ince ve içe doğru kıvrımlıydı, tibia ince ve bükülmüştü ve fibula üst uçta ince ve genişti. metatars ayağın dar olduğu ve bilinen tek toynaklı Bir ayak parmağının (pençe kemiği) ince ve hafif kavisliydi.[3] Uzuvları olsa da Stegoceras tam olarak bilinmemekle birlikte, beş parmaklı ellere ve dört ayak parmağına sahip olmaları büyük olasılıkla diğer pachycephalosaurlara benziyorlardı.[31]
Kafatası ve diş yapısı

Kafatası Stegoceras yandan bakıldığında nispeten kısa bir burun ile kabaca üçgen şeklindeydi. önden ve parietal kemikler çok kalındı ve yükseltilmiş bir kubbe oluşturdu. dikiş bu iki element arasında yok edildi (sadece bazı örneklerde zayıf bir şekilde görülebiliyordu) ve topluca "frontoparietal" olarak adlandırılıyorlar. Frontoparietal kubbe genişti ve nispeten pürüzsüz bir yüzeye sahipti, sadece yanları buruşuktu (buruşuk). Yukarıda ve arasında daraltıldı orbita (göz yuvaları). Arkada daralmış frontoparietal, arasına sıkışmıştı. skuamozal kemikler ve üzerinde bir depresyonla sona erdi oksiput kafatasının arkasında. Parietal ve skuamozal kemikler oksiput üzerinde, kapsamı örnekler arasında değişen parietoskuamozal şelf olarak adlandırılan kalın bir raf oluşturdu. Skuamozal büyüktü, kubbenin bir parçası değildi ve arka kısmı şişmişti. Düzensiz aralıklarla süslenmiştir. tüberküller (veya yuvarlak büyümeler) ve üst kenarları boyunca uzanan ve kafatasının arkasında her iki tarafta sivri bir tüberkül (veya küçük boynuz) ile biten bir sıra düğüm (topuz). Daha küçük tüberküllerin bir iç sırası, büyük olanla paralel uzanıyordu. Kubbenin üst yüzeyi dışında kafatasının büyük bir kısmı düğümlerle süslenmişti ve çoğu sıra halinde dizilmişti.[3]
Büyük yörünge kusurlu bir elips şeklindeydi (önden arkaya en uzun eksene sahip) ve yana doğru ve biraz ileriye dönüktü. zamansız fenestra gözün arkası dar ve arkaya doğru eğimliydi ve supratemporal fenestra frontoparietalın kalınlaşması nedeniyle kafatasının arka yüzünün boyutu çok küçüldü. bazikranyum (kat Braincase) yörüngelerin altındaki ve çevresindeki bölgelerden kısaltıldı ve mesafelendi. damak. Oksiput geriye ve aşağıya doğru eğimliydi ve oksipital kondil aynı yöne doğru eğildi. gözyaşı kemiği yörüngenin alt ön kenarını oluşturdu ve yüzeyinde düğüm benzeri süslemeler vardı. prefrontal ve palpebral kemikler kaynaşmış ve yörüngenin üzerinde kalın bir sırt oluşturmuştur. Nispeten büyük jugal kemik çene eklemine doğru ileriye ve aşağıya doğru uzanan yörüngenin alt kenar boşluğunu oluşturdu. Yayılan bir düzenlemede sırtlar ve düğümlerle süslenmiştir.[3]
Burun açıklıkları büyüktü ve öne doğru bakıyordu. burun kemiği kalın, ağır yontulmuş ve dışbükey bir profile sahipti. Ön kemikle birlikte kafatasının orta tepesinde bir çıkıntı (kalkan) oluşturdu. Ön alt premaksilla (üst çenenin ön kemiği) buruşmuş ve kalınlaşmıştı. Küçük foramen (delik) premaksilla arasındaki sütürde mevcuttu ve burun boşluğu ve muhtemelen Jacobson organı (bir koku alma duyu organı). Maksilla kısa ve derindi ve muhtemelen bir sinüs. Maksilla, oradaki her diş pozisyonuna karşılık gelen bir dizi foraminaya sahipti ve bunlar, yedek dişlerin sürmesi için geçitler olarak işlev görüyordu. Çene, yörüngenin arkasının altındaki kafatası ile eklemlenmiştir. Alt çenenin diş taşıyan kısmı uzundu, arkasındaki kısım oldukça kısaydı. Korunmamış olsa da, bir ön kemik alt çenenin ön tarafındaki fasetlerle gösterilir.[3] Diğer pachycephalosaurlar gibi, küçük bir gagası olacaktı.[31]

Stegoceras dişleri vardı heterodont (farklılaştırılmış) ve Thecodont (yuvalara yerleştirilmiş). Nispeten küçük dişlerden oluşan marjinal sıralara sahipti ve sıralar düz bir kesme kenarı oluşturmuyordu. Dişler, çenelerin uzunluğu boyunca eğik olarak yerleştirildi ve önden arkaya hafifçe üst üste geldi. Her iki tarafta, en eksiksiz numunenin (UALVP 2) premaksillada üç, üst çene (üst çenenin her ikisi de) ve on yedi diş hekimi alt çenenin. Premaksilladaki dişler, maksilladaki dişlerden kısa bir süre ile ayrıldı. diastema (boşluk) ve premaksilladaki iki sıra önde dişsiz bir boşlukla ayrıldı. Üst çenenin ön kısmındaki (premaksilla) ve ön alt çenenin dişleri benzerdi; bunların daha uzun, daha sivri ve kıvrık kronları ve arkada bir "topuğu" vardı. Alt çenenin ön dişleri, üst çenenin dişlerinden daha büyüktü. Kronların ön kenarlarında sekiz delik vardı dişler (çentikler) ve arka kenar dokuz ila on bir arasındaydı. Üst (maksilla) ve alt çenenin arkasındaki dişler yan görünümde üçgen ve önden bakıldığında sıkıştırılmıştı. Kesiti oval olan uzun kökleri vardı ve kronlarda belirgin cingulum üslerinde. Buradaki dişler sıkıştırıldı ve kronların tepesine doğru yönlendirildi. Hem dış hem de iç tarafı diş kronları delik emaye ve her iki taraf da bir sırt ile dikey olarak bölünmüştür. Her kenarda yaklaşık yedi veya sekiz diş eti vardı ve ön kenarı genellikle en fazla olana sahipti.[3]
Kafatası Stegoceras Belirgin parietosquamosal şelfi (yaşla birlikte küçülse de), frontoparientalinin "başlangıç" kubbesi (kubbesi yaşla birlikte artmış olsa da), şişirilmiş burun kemikleri, skuamozal kemiklerin yanlarında ve arkasında tüberküller, her bir skuamozalin üst tarafında altıya kadar tüberkülden oluşan sıralar ve parietalin geriye doğru çıkıntısında en fazla iki düğüm. Ayrıca burun süslemesi olmaması ve diyasteminin azalması açısından da farklıdır.[4][32] Kafatası S. novomexicanum bundan ayırt edilebilir S. validum Parietal kemiğin geriye doğru genişlemesinin daha küçültülmesi ve üçgen olması, daha büyük supratemporal fenestraya sahip olması (bunun nedeni örneklerin olası gençlik durumundan kaynaklanıyor olabilir) ve skuamozal ve parietal arasında kabaca paralel sütür temaslarının olması gibi özelliklerde. Aynı zamanda daha küçük bir ön patrona sahip gibi görünüyor. S. validum,[22][24] ve genel olarak daha zarif görünüyor.[25]
Sınıflandırma

1970'lerde, grup hakkında daha fazla bilgi sağlayan Asya fosillerinden daha fazla pachycephalosaur cinsi tanımlandı. 1974'te Maryańska ve Osmólska, pachycephalosaurların kendi türlerini garanti edecek kadar farklı oldukları sonucuna vardılar. alttakım Ornithischia, Pachycephalosauria içinde. 1978'de Çinli paleontolog Dong Zhiming Pachycephalosauria'yı iki aileye ayırdı; kubbe başlı Pachycephalosauridae (dahil Stegoceras) ve düz başlı Homalocephalidae (orijinal olarak Homalocephaleridae olarak yazılmıştır).[33] Wall ve Galton, 1979'da haklı gösterilen pachycephalosaurlar için alt sıra durumu bulamadı.[15] 1980'lerde, Ornithischia'daki pachycephalosaurların yakınlıkları çözülmemişti. Ana rakip görüşler, grubun ornitopodlara veya ceratopsianlara en yakın olduğu yönündeydi; ikinci görüş, iskelet arasındaki benzerlikler nedeniyle Stegoceras ve "ilkel" ceratopsian Protoceratops. 1986'da Amerikalı paleontolog Paul Sereno pachycephalosaurs ve ceratopsians arasındaki ilişkiyi destekledi ve onları grupta birleştirdi Marginocephalia, oksiput üzerindeki "raf" yapısı gibi benzer kraniyal özelliklere dayalıdır. Bu gruplama için kanıtların çok güçlü olmadığını, ancak grubun geçerliliğinin 1987'de Sues ve Galton tarafından desteklendiğini kabul etti.[3]
21. yüzyılın başlarında, postkraniyal kalıntılardan birkaç pachycephalosaur cinsi biliniyordu ve birçok takson yalnızca kubbelerden biliniyordu, bu da grup içinde sınıflandırmayı zorlaştırıyordu. Pachycephalosaurlar, bu nedenle esas olarak, düz ila kubbeli frontoparietal, postorbital ve skuamozal kemikler boyunca geniş ve düzleştirilmiş çubuk ve oksiputta derin plakalar olan skuamozal kemikler gibi kafatası özellikleriyle tanımlanır.[28] 1986'da Sereno, pachycephalosaurları kafataslarının kubbelerinin boyutuna göre farklı gruplara ayırdı ("Tholocephalidae" ve "Domocephalinae" gibi artık geçersiz taksonlarda gruplandı) ve 2000 yılında "kısmen" kubbeli olarak değerlendirdi. Stegoceras Sözde "ilkel" düz başlı ve gelişmiş "tam" kubbeli cinsler arasında bir geçiş (örneğin Pachycephalosaurus).[34] Pachycephalosaurların kubbe başlı / düz başlı bölümü, yassı kafalar düşünüldüğü için sonraki yıllarda terk edilmiştir. pomorfik (çocuk benzeri) veya çoğu revizyonda türetilmiş özellikler, ancak cinsel olarak dimorfik kişisel özellik. 2006 yılında Sullivan, pachycephalosaurlar arasındaki taksonomik yakınlıkların belirlenmesinde kubbe boyutunun yararlı olduğu fikrine karşı çıktı.[28] 2003 yılında Sullivan buldu Stegoceras kendisi daha fazlası baz alınan (veya "ilkel"), Pachycephalosaurinae alt ailesinin "tam kubbeli" üyelerinden daha iyidir, Sereno'nun 1986'da ulaştığı sonuçları detaylandırır.[4]
Bir 2013 Filogenetik analiz Evans ve meslektaşları, bazı düz başlı pachycephalosaur cinslerinin "tam" kubbeli taksonlarla "eksik" kubbeli taksonlarla daha yakından ilişkili olduğunu bulmuşlardır. StegocerasBu, kubbeli taksonların yavrularını temsil ettiklerini ve düz kafaların taksonomik yakınlıkları göstermediğini göstermektedir.[35] kladogram aşağıda yerleşimi göstermektedir Stegoceras Pachycephalosauridae'de Schott ve diğerleri, 2016'ya göre:[36]

| Pachycephalosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
biyocoğrafya ve pachycephalosaurların erken evrimsel tarihi tam olarak anlaşılamamıştır ve ancak yeni keşiflerle açıklığa kavuşturulabilir. Pachycephalosaurlar fosil kayıtlarında aniden ortaya çıkarlar ve hem Kuzey Amerika'da hem de Asya'da bulunurlar, bu nedenle ilk ne zaman ortaya çıktıkları ve hangi yönden dağıldıkları bilinmemektedir. Grubun bilinen en eski üyeleri (örneğin Akrotol ) "tam kubbelidir" ve Santoniyen Geç Kretase dönemi aşaması (yaklaşık 84 milyon yıl önce). Bu, sözde daha ilkel olandan öncedir Stegoceras -den Orta Kampaniyen (77 milyon yıl önce) ve Homalocephale -den Erken Maastrihtiyen (70 milyon yıl önce), bu nedenle kafatasının kubbesi homoplastik bir özellik olabilir (bir tür yakınsak evrim ). İlgili ceratopsia'lılara kıyasla pachycephalosaurların geç ortaya çıkması, hayalet soy (fosil kayıtlarından çıkarılan ama fosil kayıtlarından eksik), 66 milyon yıla yayılıyor. Geç Jura Kretase'ye. Pakycephalosaurlar çoğunlukla küçük olduklarından, bunun nedeni tafonomik önyargı; daha küçük hayvanların fosilleşme yoluyla korunma olasılığı daha düşüktür. Daha hassas kemiklerin de korunma olasılığı daha düşüktür, bu yüzden pachycephalosaurlar esas olarak sağlam kafataslarından bilinirler.[4][35]
Paleobiyoloji
Besleme mekaniği

Pachycephalosaurların ne yediği tam olarak belli değil; çok küçük, çıkıntılı dişlere sahip oldukları için, sert, lifli bitkileri aynı dönemin diğer dinozorları kadar etkili bir şekilde çiğneyemezlerdi. Keskin, tırtıklı dişlerinin yaprak, tohum, meyve ve böceklerden oluşan karışık bir diyet için ideal olduğu varsayılmaktadır.[37] Stegoceras diş kronlarının diş kronlarınınkilere benzer olması nedeniyle tamamen otçul bir diyet yapmış olabilir. Iguanid kertenkeleler. Premaksiller dişler, ön kemikle temastan kaynaklanan yıpranma yüzeylerini gösterir ve maksiller dişler, diğer ornitorik dinozorlarda görülenlere benzer şekilde çift aşınma yüzlerine sahiptir. UALVP 2'nin her üç çene dişinden biri sürüyor yedek dişler ve diş replasmanı sıralı üçlülerde geriye doğru ilerlemede gerçekleşti. Oksipital bölge Stegoceras kas bağlanması için sınırları iyi belirlenmiş ve çene hareketinin Stegoceras ve diğer pachycephalosaurlar çoğunlukla yukarı ve aşağı hareketlerle sınırlıydı ve çene rotasyonu için sadece küçük bir yetenek vardı. Bu, çene yapısına ve diş mikro yıpranmasına dayanır ve dişlerin aşınma yüzleri, ısırma kuvvetinin kırmadan çok kesme için kullanıldığını gösterir.[3][38]
Ontogenetik değişiklikler

Kafataslarında görülen varyasyon için tarihsel olarak birkaç açıklama önerilmiştir. Stegoceras ve diğer pachycephalosaurlar. Brown ve Schlaikjer, cinsel dimorfizm kubbe derecesinde ve AMNH 5450 gibi düz başlı örneklerin hipotezinde bulundu (Ornatotholus) kadın biçimini temsil etti Stegoceras. Bu fikir bir 1981 tarafından desteklendi morfometrik Champan ve meslektaşlarının yaptığı çalışma, erkeklerin daha büyük ve daha kalın kubbelere sahip olduğunu buldu.[32][39] Diğer düz başlı pachycephalosaurlar keşfedildikten sonra, kubbe derecesinin taksonomik öneme sahip bir özellik olduğu önerildi ve bu nedenle AMNH 5450, 1979'dan itibaren ayrı bir takson olarak kabul edildi. 1998'de Goodwin ve meslektaşları bunun yerine kubbenin şişmesinin yaşla değişen ontogenetik bir özellik olduğunu öne sürdüler. histolojik bir çalışma S. validum Kubbeyi gösteren kafatası, vasküler, hızlı büyüyen kemikten oluşuyordu, bu da yaşla birlikte kubbenin artmasıyla tutarlıydı. Bu yazarlar, sözde farklı özelliklerinin Ornatotholus kolayca ontogeninin sonucu olabilir.[32][17]
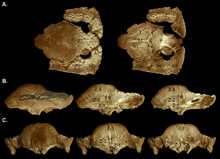
2003 yılında, Williamson ve Carr varsayımsal bir büyüme serisi yayınladılar. S. validum, gösteriliyor Ornatotholus gençlik aşaması olarak. Gençlerin düz, kalınlaştırılmış bir frontoparietal çatı, daha büyük supratemporal fenestrae ile karakterize edildiğini ve birbirine yakın tüberkül ve düğümlerle süslendiğini öne sürdüler. Parietoskuamosal rafın boyutu küçülmedi ve frontoparietal sütür açıktı. Alt yetişkinler, tepenin arka kısmı ve kafatası-çatının düz olduğu höyük benzeri kubbelere sahipti. Supratemporal fenestrae boyut olarak asimetri gösterdi ve frontoparietal sütürün kapanması değişkendi. Büyüme sırasında kubbe genişledikçe düğümler gerildi ya da neredeyse tamamen yok edildi, geriye mozaik kaplı bir yüzey kaldı. Desen genellikle maksimum genişlemenin meydana geldiği kubbenin en yüksek noktasında (tepe) yok edildi. Kafatasındaki tüberküller farklı yönlere gerilmiş ve parietoskuamozal şelfin kenarındakiler hipertrofik (büyütülmüş) tüberküller. Alt yetişkin ve yetişkin kafataslarının arka ve yanları daha az modifiye edilmiş tüberküllerle süslenmiştir. Genişleyen kubbeye dahil edilmeden önce, kafatası kemikleri genişledi ve bu kemikler arasında bağlantılar oluştu. Yetişkin kubbesi geniş ve dışbükeydi ve rafın çoğunu birleştirdi, boyut olarak küçültüldü ve oksiputun üzerine kalın bir "dudak" olarak sarktı. Supratempooral fenestrae kapatıldı, ancak frontoparietal ve bağlantılı kafatası kemikleri arasındaki sütür yetişkinlerde ve alt yetişkinlerde her zaman kapalı değildi.[19]
2011'de Ryan Schott ve arkadaşları, kraniyal kubbe ontogenisinin daha kapsamlı bir analizini yaptı. S. validum. Çalışma, parietoskuamozal şelfin, büyüme boyunca süslemenin düzenini koruduğunu ve frontoparietal kubbelerin vaskülaritesinin boyutla birlikte azaldığını buldu. Ayrıca kubbe şekli ve boyutunun büyüme ile güçlü bir şekilde ilişkili olduğunu ve büyümenin allometrik (kıyasla eş ölçülü ) düzden kubbeye, destekleyici Ornatotholus çocuk olarak Stegoceras. Ayrıca, gençlikten yetişkine dramatik değişiklikler içeren bu kubbe büyümesi modelinin, pachycephalosaurların ortak gelişimsel yörüngesi olduğunu varsaydılar. Bu araştırmacılar, Williamson ve Carr'ın supratemporal fenestranın yaşla kapandığına dair gözlemi genel olarak doğru olsa da, frontoparietal boyutuna bakılmaksızın bu fenestranın boyutunda hala yüksek derecede bireysel varyasyon olduğunu ve bu özelliğin ontogeniden bağımsız olmuştur.[32]
Schott ve David C. Evans tarafından 2012 yılında yapılan bir çalışmada, incelenen kişinin skuamozal rafındaki tek tek düğümlerin sayısı ve şeklinin S. validum kafatasları önemli ölçüde farklıydı ve bu değişkenliğin ontojenik değişikliklerle ilişkili görünmediği, ancak bireysel varyasyondan kaynaklandığı görülüyor. Bu araştırmacılar supratemporal fenestranın genişliği ile skuamozalın boyutu arasında bir ilişki bulamadılar.[40]
Kubbe işlevi

Pachycephalosaur kubbelerin işlevi tartışıldı ve Stegoceras çeşitli çalışmalarda deneysel model olarak kullanılmıştır. Kubbe, esas olarak bir silah olarak yorumlanmıştır. özel iç savaş, bir cinsel görüntü yapı veya bir araç tür tanıma.[41][42] Kafataslarının kubbeli olduğu hipotezi Stegoceras ve diğer pachycephalosaurlar kafaları kırmak için kullanıldılar ilk olarak Amerikalı paleontolog tarafından önerildi Edwin Colbert Galton, 1970 ve 1971'de bu fikir üzerinde durdu ve kubbenin basitçe dekoratif olması durumunda daha az yoğun olacağını ve yapının kuvvete direnmek için ideal olduğunu savundu. Galton şunu önerdi: Stegoceras kafatasını boyuna dik olarak tuttuğunda, kafatasından kuvvet iletilirdi ve kafatasının yerinden çıkma ihtimali çok düşüktü ve bu nedenle kubbe bir koç olarak kullanılabilirdi. Kubbenin kendisinde sivri uçlar bulunmadığından ve parietoskuamosal sahanlığın "etkisiz" bir konumda olduğundan, bunun esas olarak yırtıcı hayvanlara karşı savunma olarak kullanılmasının olası olmadığına inanıyordu, ancak onu özel bir rekabetle uyumlu buldu. Galton, vertebral kolon yatay konumda tutulurken kubbelerin birbirine vurulduğunu hayal etti. Bu, ya darbelerle karşı karşıya gelirken ya da başları alçaltılmış olarak birbirlerine yüklenirken yapılabilir (modern koyun ve keçilere benzer). Ayrıca, kafayı bu amaçla kullanırken sırtın sertliğinin faydalı olacağını da belirtti. 1978'de Sues, Galton ile pachycephalosaurların anatomisinin, kubbeden kubbeye etki stresi iletme ile tutarlı olduğu konusunda hemfikirdi. pleksi cam modeller. Darbe, boyun ve vücut tarafından emilecek ve boyun bağları ve kasları, darbelere göz atarak yaralanmaları önleyecektir (modern büyük boynuzlu koyun ). Sues ayrıca, hayvanların birbirlerinin kanatlarını kıstırabileceklerini öne sürdü.[30][43][44]
1997'de Amerikalı paleontolog Kenneth Carpenter pachycephalosaur'un arkasındaki dorsal omurların Homalocephale sırtın boynun hemen önünde aşağı doğru kıvrıldığını (korunmamış) ve boyun yukarı kıvrılmadıkça başın yere işaret edeceğini gösterin. Bu nedenle, boyunlarının Stegoceras ve diğer pachycephalosaurlar kavisli bir duruşta tutuldu (dinozorlarda olduğu gibi) ve bu nedenle başlarını, boynunu ve vücutlarını yatay olarak hizalayamayacakları için stresi iletmek için gerekli olacaktı. Their necks would have to be held below the level of the back, which would have risked damaging the spinal cord on impact. Modern bighorn sheep and bizon overcome this problem by having strong ligaments from the neck to the tall neural spines over the shoulders (which absorb the force of impact), but such features are not known in pachycephalosaurs. These animals also absorb the force of impact through sinus chambers at the base of their horns, and their foreheads and horns form a broad contact surface, unlike the narrow surface of pachycephalosaur domes. Because the dome of Stegoceras was rounded, it would have given a very small area for potential impact, and the domes would have glanced off each other (unless the impact was perfectly centred). Combating pachycephalosaurs would have had difficulty seeing each other while their heads were lowered, due to the bony ridges above the eyes.[30]

Because of the problems he found with the head-butting hypothesis, Carpenter instead suggested the domes were adaptations for flank-butting (as seen in some large African mammals); he imagined that two animals would stand parallel, facing each other or the same direction, and direct blows to the side of the opponent. The relatively large body width of pachycephalosaurs may consequently have served to protect vital organs from harm during flank-butting. Bu mümkündür Stegoceras and similar pachycephalosaurs would have delivered the blows with a movement of the neck from the side and a rotation of the head. The upper sides of the dome have the greatest surface area, and may have been the point of impact. The thickness of the dome would have increased the power behind a blow to the sides, and this would ensure that the opponent felt the force of the impact, without being seriously injured. The bone rim above the orbit may have protected the aggressor's eye when making a blow. Carpenter suggested that the pachycephalosaurs would have first engaged in tehdit ekranı by bobbing and presenting their heads to show the size of their domes (intimidation), and thereafter delivered blows to each other, until one opponent signalled submission.[30]
In 1987, J. K. Rigby and colleagues suggested that pachycephalosaur domes were heat-exchange organs used for termoregülasyon, based on their internal "radiating structures" (Trabeküller ). This idea was supported by a few other writers in the mid-1990s.[42] In 1998, Goodwin and colleagues considered the lack of sinuses in the skull of Stegoceras ve "bal peteği "-like network of vascular bone in the dome ill-suited for head-butting, and pointed out that the bones adjacent to the dome risked fracture during such contact. Building on the idea that the ossified tendons that stiffened the tails of Stegoceras and other pachycephalosaurs enabled them to take a tripodal stance (first suggested by Maryańska and Osmólska in 1974), Goodwin et al. suggested these structures could have protected the tail against flank-butting, or that the tail itself could have been used as a weapon.[17] In 2004, Goodwin and colleagues studied the cranial histology of pachycephalosaurs, and found that the vascularity (including the trabeculae) of the domes decreased with age, which they found inconsistent with a function in either head-butting or heat-exchange. They also suggested that a dense layer of Sharpey's fibers near the surface of the dome indicated that it had an external covering in life, which makes it impossible to know the shape of the dome in a living animal. These researchers instead concluded that the domes were mainly for species recognition and communication (as in some African teklifler ) and that use in sexual display was only secondary. They further speculated that the external covering of the domes was brightly coloured in life, or may have changed colour seasonally.[42]

In 2008, Eric Snively and Adam Cox tested the performance of 2D and 3D pachycephalosaur skulls through sonlu elemanlar analizi, and found that they could withstand considerable impact; greater vaulting of the domes allowed for higher forces of impact. They also considered it likely that pachycephalosaur domes were covered in keratin, a strong material that can withstand much energy without being permanently damaged (like the osteodermler nın-nin timsahlar ), and therefore incorporated keratin into their test formula.[45] In 2011, American palaeontologists Kevin Padian ve John R. Horner proposed that "bizarre structures" in dinosaurs in general (including domes, frills, horns, and crests) were primarily used for species recognition, and dismissed other explanations as unsupported by evidence. Among other studies, these authors cited Goodwin et al.'s 2004 paper on pachycephalosaur domes as support of this idea, and they pointed out that such structures did not appear to be sexually dimorphic.[46] In a response to Padian and Horner the same year, Rob J. Knell and Scott D. Sampson argued that species recognition was not unlikely as a secondary function for "bizarre structures" in dinosaurs, but that cinsel seçim (used in display or combat to compete for mates) was a more likely explanation, due to the high cost of developing them, and because such structures appear to be highly variable within species.[47] In 2013, the British palaeontologists David E. Hone and Darren Naish criticized the "species recognition hypothesis", and argued that no extant animals use such structures primarily for species recognition, and that Padian and Horner had ignored the possibility of mutual sexual selection (where both sexes are ornamented).[48]

In 2011, Snively and Jessica M. Theodor conducted a finite element analysis by simulating head-impacts with CT scanned skulls of S. validum (UALVP 2), Prenocephale prenes and several extant head-butting artiodaktiller. They found that the correlations between head-striking and skull morphologies found in the living animals also existed in the studied pachycephalosaurs. Stegoceras ve Prenocephale both had skull shapes similar to the bighorn sheep with süngerimsi kemik protecting the brain. They also shared similarities in the distribution of compact and cancellous regions with the bighorn sheep, beyaz karınlı duiker ve zürafa. The white-bellied duiker was found to be the closest morphological analogue to Stegoceras; this head-butting species has a dome which is smaller but similarly rounded. Stegoceras was better capable of dissipating force than artiodactyls that butt heads at high forces, but the less vascularized domes of older pachycephalosaurs, and possibly diminished ability to heal from injuries, argued against such combat in older individuals. The study also tested the effects of a keratinous covering of the dome, and found it to aid in performance. Rağmen Stegoceras lacked the pnömatik sinuses that are found below the point of impact in the skulls of head-striking artiodactyls, it instead had vascular struts which could have similarly acted as braces, as well as conduits to feed the development of a keratin covering.[49]

In 2012, Schott and Evans suggested that the regularity in squamosal ornamentation throughout the ontogeny of Stegoceras was consistent with species recognition, but the change from flat to domed frontoparietals in late age suggests that the function of this feature changed through ontogeny, and was perhaps sexually selected, possibly for intra-specific combat.[40] In 2012, Caleb M. Brown and Anthony P. Russell suggested that the stiffened tails were probably not used as defence against flank-butting, but may have enabled the animals to take a tripodal stance during intra-specific combat, with the tail as support. Brown and Russell found that the tail could thereby help in resisting compressive, tensile, and torsional loading when the animal delivered or received blows with the dome.[9]
A 2013 study by Joseph E. Peterson and colleagues identified lesions in skulls of Stegoceras and other pachycephalosaurs, which were interpreted as infections caused by trauma. Lesions were found on 22% of sampled pachycephalosaur skulls (a frequency consistent across genera), but were absent from flat-headed specimens (which have been interpreted as juveniles or females), which is consistent with use in intra-specific combat (for territory or mates). The distribution of lesions in these animals tended to concentrate at the top of the dome, which supports head-butting behaviour. Flank-butting would probably result in fewer injuries, which would instead be concentrated on the sides of the dome. These observations were made while comparing the lesions with those on the skulls and flanks of modern sheep skeletons. The researchers noted that modern head-butting animals use their weapons for both combat and display, and that pachycephalosaurs could therefore also have used their domes for both. Displaying a weapon and willingness to use it can be enough to settle disputes in some animals.[41]
Nasal passages
In 1989, Emily B. Griffin found that Stegoceras and other pachycephalosaurs had a good sense of smell (olfaction), based on the study of cranial endokastlar that showed large koku soğanları beyinde.[50] In 2014, Jason M. Bourke and colleagues found that Stegoceras would have needed cartilaginous nasal turbinates in the front of the nasal passages for airflow to reach the olfactory region. Evidence for the presence of this structure is a bony ridge to which it could have attached. The size of the olfactory region also indicates that Stegoceras had a keen sense of smell. The researchers found that the dinosaur could have had either a scroll-shaped turbinate (like in a Türkiye ) or a branched one (as in an devekuşu ) as both could have directed air to the olfactory region. The blood vessel system in the passages also suggest that the turbinates served to cool down warm arterial blood from the body that was heading to the brain. Kafatası S. validum specimen UALVP 2 was suited for a study of this kind due to its exceptional preservation; it has ossified soft tissue in the nasal cavity, which would otherwise be kıkırdaklı and therefore not preserved through mineralization.[51]
Paleoekoloji

S. validum is known from the late Geç Kretase Belly River Group (the Canadian equivalent to the Judith River Group in the US), and specimens have been recovered from the Dinozor Parkı Oluşumu (late Campanian, 76.5 to 75 mya) in Dinozor İl Parkı (including the lectotype specimen), and the Oldman Formasyonu (middle Campanian, 77.5 to 76.5 mya) of Alberta, Canada. The pachycephalosaurs Hanssuesia ve Foraminacephale are also known from both formations.[4] S. novomexicanum dan bilinmektedir Fruitland (late Campanian, about 75 mya) and lower Kirtland Formasyonu (geç Kampaniyen, about 74 mya) of New Mexico, and if this species correctly belongs in Stegoceras, the genus would have had a broad geographic distribution.[22] The presence of similar pachycephalosaurs in both the west and north of North America during the latest Cretaceous shows that they were an important part of the dinosaur faunas there.[24] It has traditionally been suggested that pachycehalosaurs inhabited mountain environments; wear of their skulls was supposedly a result of them having been rolled by water from upland areas, and comparisons with bighorn sheep reinforced the theory. In 2014, Jordan C. Mallon and Evans disputed this idea, as the wear and original locations of the skulls is not consistent with having been transported in such a way, and they instead proposed that North American pachycephalosaurs inhabited alüvyon (associated with water) and kıyı düzlüğü ortamlar.[52]
The Dinosaur Park Formation is interpreted as a low-relief setting of nehirler ve taşkın yatakları that became more swampy ve etkilenen deniz conditions over time as the Batı İç Denizyolu aşılmış batıya.[53] iklim was warmer than present-day Alberta, without don, but with wetter and drier seasons. İğne yapraklılar were apparently the dominant gölgelik plants, with an alt hikaye nın-nin eğrelti otları, ağaç eğrelti otları, ve anjiyospermler.[54] Dinosaur Park is known for its diverse community of herbivores. Hem de Stegoceras, the formation has also yielded fossils of the ceratopsians Erboğa, Styracosaurus ve Chasmosaurus, hadrosauridler Prosaurolophus, Lambeosaurus, Gryposaurus, Corythosaurus, ve Parasaurolophus, ve Ankylosaurlar Edmontoni ve Euoplocephalus. Theropods present include the tyrannosaurids Gorgosaurus ve Daspletosaurus.[55] Other dinosaurs known from the Oldman Formation include the hadrosaur Brachylophosaurus, the ceratopsians Coronosaurus ve Albertaceratops, ornitomimidler, Therizinosaurlar and possibly ankylosaurs. Theropods included Troodontidler, Oviraptorozorlar, dromaeosaurid Saurornitholestes and possibly an albertosaurine tyrannosaur.[56]
Ayrıca bakınız
Referanslar
- ^ a b Lambe, L. M. (1902). "New genera and species from the Belly River Series (mid-Cretaceous)". Geological Survey of Canada, Contributions to Canadian Palaeontology. 3: 68.
- ^ a b c Hatcher, J.B.; Lull, R.S.; Marsh, O.C.; Osborn, H. F. (1907). "The Ceratopsia". Monographs of the United States Geological Survey. XLIX. doi:10.5962/bhl.title.60500.
- ^ a b c d e f g h ben j k l Sues, H. D. & Galton, P. M. (1987). "Anatomy and classification of the North American Pachycephalosauria (Dinosauria: Ornithischia)". Palaeontographica Abteilung A. 198: 1–40.
- ^ a b c d e f g Sullivan, R. M. (2003). "Dinozorun revizyonu Stegoceras Lambe (Ornithischia, Pachycephalosauridae) ". Omurgalı Paleontoloji Dergisi. 23 (1): 181–207. doi:10.1671/0272-4634(2003)23[181:rotdsl]2.0.co;2.
- ^ Nopcsa, F. (1903). "Über Stegoceras und Stereocephalus". Centralblatt für Mineralogie, Geologie und Paläontologie. 1903: 266–267.
- ^ Lambe, L.M. (1903). "Recent Zoopaleontology". Bilim. 18 (445): 60. Bibcode:1903Sci....18...60L. doi:10.1126/science.18.445.60. JSTOR 1631645. PMID 17746863.
- ^ Lambe, L. M. (1918). "The Cretaceous genus Stegoceras typifying a new family referred provisionally to the Stegosauria". Kanada Kraliyet Cemiyeti'nin İşlemleri. 12 (4): 23–36.
- ^ Gilmore, C. W., 1924. On Troodon validus, an orthopodous dinosaur from the Belly River Cretaceous of Alberta, Canada. Department of Geology, University of Alberta Bulletin 1:1–43
- ^ a b c d Brown, C. M .; Russell, A. P.; Farke, A. A. (2012). "Homology and Architecture of the Caudal Basket of Pachycephalosauria (Dinosauria: Ornithischia): The First Occurrence of Myorhabdoi in Tetrapoda". PLOS ONE. 7 (1): e30212. Bibcode:2012PLoSO...730212B. doi:10.1371/journal.pone.0030212. PMC 3260247. PMID 22272307.
- ^ a b Brown, B .; E. M., Schlaikjer (1943). "A study of the troödont dinosaurs, with the description of a new genus and four new species". AMNH Bülteni. 82. hdl:2246/387.
- ^ Sternberg, C. M. (1945). "Pachycephalosauridae Proposed for Dome-Headed Dinosaurs, Stegoceras lambei, n. sp., Described". Paleontoloji Dergisi. 19 (5): 534–538. JSTOR 1299007.
- ^ Russell, L. S. (1948). "The Dentary of Troödon, a Genus of Theropod Dinosaurs". Paleontoloji Dergisi. 22 (5): 625–629. JSTOR 1299599.
- ^ Bohlin, B., 1953. Fossil reptiles from Mongolia and Kansu. Reports from the Scientific Expedition to the North-western Provinces of China under Leadership of Dr. Sven Hedin. VI. Vertebrate Palaeontology 6. The Sino-Swedish Expedition Publication 37:1–113
- ^ Kuhn, O., 1964, Fossilium Catalogus I: Animalia Pars 105. Ornithischia (Supplementum I), IJsel Pers, Deventer, 80 pp
- ^ a b Wall, W. P.; Galton, P. M. (1979). "Notes on pachycephalosaurid dinosaurs (Reptilia: Ornithischia) from North America, with comments on their status as ornithopods". Kanada Yer Bilimleri Dergisi. 16 (6): 1176–1186. Bibcode:1979CaJES..16.1176W. doi:10.1139/e79-104.
- ^ Galton, P. M .; Sues, H.-D. (1983). "New data on pachycephalosaurid dinosaurs (Reptilia: Ornithischia) from North America". Kanada Yer Bilimleri Dergisi. 20 (3): 462–472. Bibcode:1983CaJES..20..462G. doi:10.1139/e83-043.
- ^ a b c Goodwin, M. B.; Buchholtz, E. A.; Johnson, R. E. (1998). "Cranial anatomy and diagnosis of Stygimoloch spinifer (Ornithischia: Pachycephalosauria) with comments on cranial display structures in agonistic behavior". Omurgalı Paleontoloji Dergisi. 18 (2): 363–375. doi:10.1080/02724634.1998.10011064.
- ^ Sullivan, Robert M. (2000). "Prenocephale edmontonensis (Brown and Schlaikjer) new comb. ve P. brevis (Lambe) new comb. (Dinosauria: Ornithischia: Pachycephalosauria) from the Upper Cretaceous of North America". New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 17: 177–90.
- ^ a b Williamson, T. E.; Carr, T. D. (2003). "A new genus of derived pachycephalosaurian from western North America". Omurgalı Paleontoloji Dergisi. 22 (4): 779–801. doi:10.1671/0272-4634(2002)022[0779:ANGODP]2.0.CO;2.
- ^ Sullivan, R. M.; Lucas, S. G. (2006). "The pachycephalosaurid dinosaur Stegoceras validum from the Upper Cretaceous Fruitland Formation, San Juan Basin, New Mexico". New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 35: 329–330.
- ^ Williamson, T. E.; Carr, T. D. (2002). "A juvenile pachycephalosaur (Dinosauria: Pachycephalosauridae) from the Fruitland Formation". New Mexico: New Mexico Geology. 24: 67–68.
- ^ a b c d Jasinski, S. E.; Sullivan, R. M. (2011). "Re-evaluation of pachycephalosaurids from the Fruitland-Kirtland transition (Kirtlandian, late Campanian), San Juan Basin, New Mexico, with a description of a new species of Stegoceras ve yeniden değerlendirme Texascephale langstoni" (PDF). Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin. 53: 202–215.
- ^ Watabe, M .; Tsogtbaatar, K .; Sullivan, R. M. (2011). "A new pachycephalosaurid from the Baynshire Formation (Cenomanian-late Santonian), Gobi Desert, Mongolia" (PDF). Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin. 53: 489–497.
- ^ a b c d Williamson, T. E.; Brusatte, S. L. (2016). "Pachycephalosaurs (Dinosauria: Ornithischia) from the Upper Cretaceous (upper Campanian) of New Mexico: A reassessment of Stegoceras novomexicanum". Kretase Araştırmaları. 62: 29–43. doi:10.1016/j.cretres.2016.01.012.
- ^ a b Jasinski, S. E.; Sullivan, R. M (2016). "The validity of the Late Cretaceous pachycephalosaurid Stegoceras novomexicanum (Dinosauria: Pachycephalosauridae)". New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 74 (107).
- ^ Glut, D. F. (1997). Dinozorlar: Ansiklopedi. Jefferson, North Carolina: McFarland & Co. pp. 834–838. ISBN 978-0-89950-917-4.
- ^ Lambert, D. (1993). Nihai Dinozor Kitabı. New York: Dorling Kindersley. s.155. ISBN 978-1-56458-304-8.
- ^ a b c Sullivan, R. M. (2006). "A taxonomic review of the Pachycephalosauridae (Dinosauria: Ornithischia)". New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 35: 347–365.
- ^ Peczkis, J. (1995). "Implications of Body-Mass Estimates for Dinosaurs". Omurgalı Paleontoloji Dergisi. 14 (4): 520–533. doi:10.1080/02724634.1995.10011575. JSTOR 4523591.
- ^ a b c d Carpenter, K. (1997). "Agonistic behavior in pachycephalosaurs (Ornithischia: Dinosauria): a new look at head-butting behavior" (pdf). Jeolojiye Katkılar. 32 (1): 19–25.
- ^ a b Paul, G. S. (2010). Princeton Dinozorlar Saha Rehberi. Princeton University Press. pp.241–242. ISBN 978-0-691-13720-9.
- ^ a b c d Schott, R.K .; Evans, D. C .; Goodwin, M. B.; Horner, J. R.; Brown, C. M .; Longrich, N. R. (2011). "Cranial ontogeny in Stegoceras validum (Dinosauria: Pachycephalosauria): a quantitative model of pachycephalosaur dome growth and Variation". PLOS ONE. 6 (6): e21092. Bibcode:2011PLoSO...621092S. doi:10.1371/journal.pone.0021092. PMC 3126802. PMID 21738608.
- ^ Perle, A.; Osmólska, H. (1982). "Goyocephale lattimorei gen. et sp. n., a new flat-headed pachycephalosaur (Ornlthlschia, Dinosauria) from the Upper Cretaceous of Mongolia – Acta Palaeontologica Polonica". Acta Palaeontologica Polonica. 27: 115–127.
- ^ Sereno, P. C., 2000. The fossil record, systematics and evolution of pachycephalosaurs and ceratopsians from Asia. 480–516 in Benton, M.J., M.A. Shishkin, D.M. Unwin & E.N. Kurochkin (eds.), Rusya ve Moğolistan'da Dinozorlar Çağı. Cambridge University Press, Cambridge.
- ^ a b Evans, D. C .; Schott, R.K .; Larson, D. W .; Brown, C. M .; Ryan, M.J. (2013). "En eski Kuzey Amerika pachycephalosaurid ve küçük gövdeli ornithischian dinozorlarının gizli çeşitliliği". Doğa İletişimi. 4: 1828. Bibcode:2013NatCo ... 4.1828E. doi:10.1038 / ncomms2749. PMID 23652016.
- ^ Schott, R.K .; Evans, D. C. (2016). "Cranial variation and systematics of Foraminacephale brevis gen. kas. and the diversity of pachycephalosaurid dinosaurs (Ornithischia: Cerapoda) in the Belly River Group of Alberta, Canada". Linnean Society'nin Zooloji Dergisi. doi:10.1111/zoj.12465.
- ^ Maryańska, T .; Chapman, R. E.; Weishampel, D. B. (2004). "Pachycephalosauria". Weishampel, D. B .; Dodson, P.; Osmólska, H. (editörler). Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. pp.464 –477. ISBN 978-0-520-24209-8.
- ^ Nabavizadeh, A. (2016). "Evolutionary Trends in the Jaw Adductor Mechanics of Ornithischian Dinosaurs". Anatomik Kayıt. 299 (3): 271–294. doi:10.1002/ar.23306. PMID 26692539.
- ^ Chapman, R. E.; Galton, Pe. M .; Sepkoski, J. J.; Wall, W. P. (1981). "A Morphometric Study of the Cranium of the Pachycephalosaurid Dinosaur Stegoceras". Paleontoloji Dergisi. 55 (3): 608–618. JSTOR 1304275.
- ^ a b Schott, R.K .; Evans, D. C. (2012). "Squamosal Ontogeny and Variation in the Pachycephalosaurian Dinosaur Stegoceras validum Lambe, 1902, from the Dinosaur Park Formation, Alberta". Omurgalı Paleontoloji Dergisi. 32 (4): 903–913. doi:10.1080/02724634.2012.679878. JSTOR 23251281. S2CID 84526252.
- ^ a b Peterson, J. E.; Dischler, C.; Longrich, N. R .; Dodson, P. (2013). "Distributions of Cranial Pathologies Provide Evidence for Head-Butting in Dome-Headed Dinosaurs (Pachycephalosauridae)". PLOS ONE. 8 (7): e68620. Bibcode:2013PLoSO...868620P. doi:10.1371/journal.pone.0068620. PMC 3712952. PMID 23874691.
- ^ a b c Goodwin, M. B.; Horner, J. R. (2004). "Cranial Histology of Pachycephalosaurs (Ornithischia: Marginocephalia) Reveals Transitory Structures Inconsistent with Head-Butting Behavior". Paleobiyoloji. 30 (2): 253–267. doi:10.1666/0094-8373(2004)030<0253:chopom>2.0.co;2. JSTOR 4096846.
- ^ Galton, P. M. (1971). "A Primitive Dome-Headed Dinosaur (Ornithischia: Pachycephalosauridae) from the Lower Cretaceous of England and the Function of the Dome of Pachycephalosaurids". Paleontoloji Dergisi. 45 (1): 40–47. JSTOR 1302750.
- ^ Sues, H. D. (1978). "Functional morphology of the dome in pachycephalosaurid dinosaurs". Neues Jahrbuch für Geologie und Paläontologie - Monatshefte. 8: 459–472.
- ^ Snively, E .; Cox, A. (2008). "Structural Mechanics of Pachycephalosaur Crania Permitted Head-butting Behavior". Paleontoloji Electronica. 11: 1–17.
- ^ Padian, K .; Horner, J. R. (2011). "The evolution of 'bizarre structures' in dinosaurs: biomechanics, sexual selection, social selection or species recognition?". Zooloji Dergisi. 283 (1): 3–17. doi:10.1111/j.1469-7998.2010.00719.x.
- ^ Knell, R. J.; Sampson, S. (2011). "Bizarre structures in dinosaurs: species recognition or sexual selection? A response to Padian and Horner" (PDF). Zooloji Dergisi. 283 (1): 18–22. doi:10.1111/j.1469-7998.2010.00758.x.
- ^ Hone, D. W. E .; Naish, D. (2013). "The 'species recognition hypothesis' does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs". Zooloji Dergisi. 290 (3): 172–180. doi:10.1111/jzo.12035.
- ^ Snively, E .; Theodor, J. M. (2011). "Common Functional Correlates of Head-Strike Behavior in the Pachycephalosaur Stegoceras validum (Ornithischia, Dinosauria) and Combative Artiodactyls". PLOS ONE. 6 (6): e21422. Bibcode:2011PLoSO...621422S. doi:10.1371/journal.pone.0021422. PMC 3125168. PMID 21738658.
- ^ Giffin, E. B. (1989). "Pachycephalosaur Paleoneurolagy (Archosauria: Ornithischia)". Omurgalı Paleontoloji Dergisi. 9 (1): 67–77. doi:10.1080/02724634.1989.10011739. JSTOR 4523238.
- ^ Bourke, J. M.; Porter, Wm. R .; Ridgely, R. C.; Lyson, T. R.; Schachner, E. R .; Bell, P. R.; Witmer, L. M. (2014). "Breathing life into dinosaurs: tackling challenges of soft-tissue restoration and nasal airflow in extinct species" (PDF). Anatomik Kayıt. 297 (11): 2148–2186. doi:10.1002/ar.23046. PMID 25312371. S2CID 4660680. Arşivlenen orijinal (PDF) 2016-08-09 tarihinde.
- ^ Mallon, J. C.; Evans, D. C. (2014). "Taphonomy and habitat preference of North American pachycephalosaurids (Dinosauria, Ornithischia)". Lethaia. 47 (4): 567–578. doi:10.1111/let.12082.
- ^ Eberth, David A. (2005). "The Geology". Currie'de, Philip J .; Koppelhus, Eva Bundgaard (eds.). Dinozor İl Parkı. pp.54–82. ISBN 978-0-253-34595-0.
- ^ Braman, Dennis R .; Koppelhus, Eva B. (2005). "Campanian palynomorphs". Currie'de, Philip J .; Koppelhus, Eva Bundgaard (eds.). Dinozor İl Parkı. pp.101–30. ISBN 978-0-253-34595-0.
- ^ Weishampel, D. B.; Barrett, P. M .; Coria, R. A .; Le Loeuff, J .; Xu Xing; Z. X.; Sahni, A .; Gomani M.P., Elizabeth; Noto, C. R. (2004). "Dinosaur Distribution". Weishampel, D. B .; Dodson, P.; Osmólska, H. (editörler). Dinosauria (2. baskı). California Üniversitesi Yayınları. pp.517 –606. ISBN 978-0-520-24209-8.
- ^ Eberth, D. A. (1997). "Judith River Wedge". İçinde Currie, Philip J.; Padian, Kevin (eds.). Dinozorlar Ansiklopedisi. San Diego: Akademik Basın. pp.199 –204. ISBN 978-0-12-226810-6.
Dış bağlantılar
 İle ilgili medya Stegoceras Wikimedia Commons'ta
İle ilgili medya Stegoceras Wikimedia Commons'ta İle ilgili veriler Stegoceras Wikispecies'de
İle ilgili veriler Stegoceras Wikispecies'de
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||





