Riboz - Ribose - Wikipedia
| |||
| |||
| İsimler | |||
|---|---|---|---|
| IUPAC adı (2R,3R,4S,5R) -5- (hidroksimetil) oksolan-2,3,4-triol | |||
| Diğer isimler d-Ribose | |||
| Tanımlayıcılar | |||
3 boyutlu model (JSmol ) |
| ||
| ChEMBL | |||
| ChemSpider |
| ||
| DrugBank | |||
| EC Numarası |
| ||
PubChem Müşteri Kimliği | |||
| UNII | |||
| |||
| |||
| Özellikleri[1][2] | |||
| C5H10Ö5 | |||
| Molar kütle | 150.13 | ||
| Görünüm | Beyaz katı | ||
| Erime noktası | 95 ° C (203 ° F; 368 K) | ||
| 100 g / L (25 ° C, 77 ° F) | |||
Kiral rotasyon ([α]D) | −21.5 ° (H2Ö) | ||
| Bağıntılı bileşikler | |||
İlişkili aldopentozlar | Arabinose Ksiloz Lyxose | ||
Bağıntılı bileşikler | Deoksiriboz | ||
Aksi belirtilmedikçe, veriler kendi içlerindeki malzemeler için verilmiştir. standart durum (25 ° C'de [77 ° F], 100 kPa). | |||
| Bilgi kutusu referansları | |||

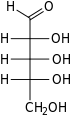


Riboz bir basit şeker ve karbonhidrat ile Moleküler formül C5H10Ö5 ve doğrusal form bileşimi H− (C = O) - (CHOH)4−H. Doğal olarak oluşan form, d-ribose, bir bileşenidir ribonükleotidler olan RNA inşa edilmiştir ve bu nedenle bu bileşik, kodlama, kod çözme, düzenleme ve ifade nın-nin genler. Bir yapısal analog, deoksiriboz benzer şekilde temel bir bileşen olan DNA. l-Ribose ilk olarak tarafından hazırlanan doğal olmayan bir şekerdir. Emil Fischer ve Oscar Piloty 1891'de.[3] 1909 yılına kadar Phoebus Levene ve Walter Jacobs bunu kabul etti d-ribose bir doğal ürün, enantiyomer Fischer ve Piloty'nin ürünü ve önemli bir bileşeni nükleik asitler.[4][5][6] Fischer, başka bir şekerin adının kısmen yeniden düzenlenmesi olduğu için "riboz" adını seçti. arabinoz ribozun bir epimer 2 'karbonda; her iki isim de ilgili Arap sakızı hangi arabinozun ilk izole edildiği ve hangisinden hazırlandığı l-ribose.[6][7]

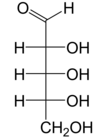

Sağ: Fischer projeksiyonu of açık zincir biçimleri d- ve l-ribose
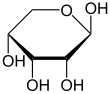
Çoğu şeker gibi riboz, döngüsel formlar içinde denge Doğrusal biçimiyle ve bunlar, özellikle sulu çözelti.[8] "Riboz" adı, biyokimya ve biyolojide tüm bu formlara atıfta bulunmak için kullanılır, ancak gerektiğinde her biri için daha spesifik isimler kullanılır. Doğrusal formunda riboz, pentoz tümüyle şeker hidroksil fonksiyonel gruplar aynı tarafta Fischer projeksiyonu. d-Ribose bu hidroksil gruplarına sağ tarafta sahiptir ve sistematik isim (2R,3R,4R) -2,3,4,5-tetrahidroksipentanal,[9] İken lribozun hidroksil grupları bir Fischer projeksiyonunda sol tarafta görünür. Ribozun siklizasyonu, yarı asetal saldırı nedeniyle oluşum aldehit C4 'hidroksil grubu tarafından bir furanoz oluşturmak için C5 'hidroksil grubu veya formu piranoz form. Her durumda, α- ve β- olarak adlandırılan ve olarak bilinen iki olası geometrik sonuç vardır. anomerler, bağlı olarak stereokimya hemiasetal karbon atomunda ("anomerik karbon"). Oda sıcaklığında, yaklaşık% 76 d-riboz, piranoz formlarında bulunur[8]:228 (α: β = 1: 2)[10] ve furanoz formlarında% 24[8]:228 (α: β = 1: 3),[10] doğrusal formun yalnızca yaklaşık% 0,1'i mevcut.[11][12]
ribonükleositler adenozin, sitidin, guanozin, ve üridin hepsi türevler / β-d- ribofuranose. Metabolik açıdan önemli içeren türler fosforile riboz içerir ADP, ATP, koenzim A,[8]:228–229 ve NADH. kamp ve cGMP bazı sinyal yollarında ikincil haberciler olarak görev yapar ve ayrıca riboz türevleridir. Riboz parça antibiyotikler dahil bazı farmasötik ajanlarda görülür neomisin ve paromomisin.[10]
Sentez ve kaynaklar
5-fosfat esteri olarak riboz, tipik olarak glikozdan üretilir. pentoz fosfat yolu. En azından bazı archaea'da alternatif yollar tanımlanmıştır.[13]
Riboz kimyasal olarak sentezlenebilir, ancak ticari üretim glikozun fermantasyonuna dayanır. Genetiği değiştirilmiş suşların kullanılması B. subtilis90 g / litre riboz, 200 g glikozdan üretilebilir. Dönüşüm, glukonat ve ribulozun ara olmasını gerektirir.[14]
Riboz tespit edildi göktaşları.[15][16]
Yapısı
Riboz bir aldopentoz (beş içeren bir monosakkarit karbon atomlar) açık zincir form, bir aldehit fonksiyonel grup sondan birinci. Monosakkaritler için geleneksel numaralandırma şemasında, karbon atomları C1 'den (aldehit grubunda) C5' e numaralandırılır. deoksiriboz DNA'da bulunan türev, ribozdan farklıdır. hidrojen yerine atom hidroksil grup C2 '. Bu hidroksil grubu, RNA ekleme.
"d- "adına d-ribose, stereokimya of kiral aldehit grubundan (C4 ') en uzaktaki karbon atomu. İçinde d-hepsi olduğu gibi d-şekerler, bu karbon atomunun konfigürasyonu şu şekildedir: d-gliseraldehit.

α-dRibopiranoz
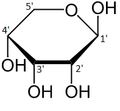
β-dRibopiranoz

α-d-Ribofuranose

β-d-Ribofuranose




Çözeltideki riboz formlarının göreceli bolluğu: β-d-ribopiranoz (% 59), α-d-ribopiranoz (% 20), β-d-ribofuranose (% 13), α-d- ribofuranoz (% 7) ve açık zincir (% 0.1).[11]
İçindeki riboz kalıntıları için nükleositler ve nükleotid bağları kapsayan dönme için burulma açıları, ilgili nükleosit ve nükleotidin konfigürasyonunu etkiler. ikincil yapı bir nükleik asidin 7'sinin dönüşü ile belirlenir. burulma açıları.[17] Büyük miktarda burulma açısına sahip olmak, daha fazla esneklik sağlar.
Kapalı halka nervürlerinde, yukarıda bahsedilen gözlenen esneklik gözlenmez çünkü halka döngüsü yapıdaki olası burulma açılarının sayısına bir sınır getirir.[17] Kapalı form ribozların konformatörleri, ne kadar yalnız olduğuna göre farklılık gösterir. oksijen molekülde, göreceli olarak konumlandırılmıştır. azotlu baz (olarak da bilinir nükleobaz veya sadece bir baz) riboza tutturulmuştur. Bir karbon tabana doğru bakıyorsa riboz endo olarak etiketlenir. Bir karbon tabandan uzağa bakıyorsa, riboz ekso olarak etiketlenir. Kapalı döngü ribozun 2 'karbonuna bağlı bir oksijen molekülü varsa, o zaman ekso onayı daha kararlıdır çünkü oksijenin bazla etkileşimini azaltır.[17] Farkın kendisi oldukça küçüktür, ancak bütün bir RNA zincirine bakıldığında, bu küçük fark büyük bir etkiye neden olur.
- Ribose'un bazı büzülme konfigürasyonları
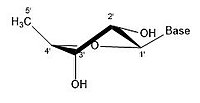
2 'endo
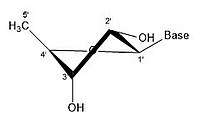
2 'endo 3' exo
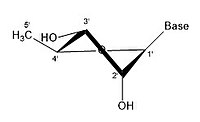
3 'endo 2' exo

3 'endo




Bir riboz molekülü tipik olarak kağıt üzerinde düzlemsel bir molekül olarak temsil edilir. Buna rağmen, doğası gereği tipik olarak düzlemsel değildir. Hidrojen atomları arasında bile, bir riboz molekülü üzerindeki birçok bileşen sterik engel ve aralarında gerginlik. Bu kalabalığı gidermek ve halka gerginliği halka büzülür, yani düzlemsel olmaz.[18] Bu büzülme, bir atomu düzlemden uzaklaştırarak, gerginliği hafifleterek ve daha kararlı bir konfigürasyon sağlayarak elde edilir.[17] Şeker halkası konformasyonu (özellikle riboz şekeri) olarak bilinen buruşma, büzüşmenin yanı sıra büzüşmenin genliği ile tanımlanabilir. sözde dönme açı. Sözde dönüş açısı "kuzey (N)" veya "güney (S)" aralığı olarak tanımlanabilir. Her iki aralık da çift sarmallarda bulunurken, kuzey aralığı genellikle RNA ve Bir çeşit DNA. Bunun aksine, güney bölgesi, B formu DNA. Z-DNA hem kuzey hem de güneyde şekerler içerir.[19] Yalnızca tek bir atom yer değiştirdiğinde, buna "zarf" büzüşmesi denir. İki atom yer değiştirdiğinde, zikzak oryantasyonuna atıfta bulunularak "bükülme" büzülmesi olarak adlandırılır.[20] Bir "endo" büzüşmede, atomların ana yer değiştirmesi β-yüzünde, C4'-C5 'bağı ve taban ile aynı taraftadır. Bir "ekso" büzülmesinde, atomların ana yer değiştirmesi, halkanın karşı tarafında, α-yüzündedir. Ribozun başlıca formları, 3'-endo büzüştürücü (genellikle RNA ve A-form DNA tarafından benimsenir) ve 2'-endo büzüştürücüdür (genellikle B-form DNA tarafından benimsenir).[21] Bu halka büzüşmeleri, halka burulma açılarındaki değişikliklerden geliştirilmiştir; sonsuz sayıda açı kombinasyonu vardır, bu nedenle, her biri farklı aktivasyon enerjileri ile ayrılmış sonsuz sayıda yer değiştirebilir büzülme şekli vardır.
Fonksiyonlar
ATP, ribozdan türetilir; bir riboz içerir, üç fosfat gruplar ve bir adenin taban. ATP, hücresel solunum itibaren adenozin difosfat (Bir fosfat grubu eksik olan ATP).
Sinyal yolları
Riboz, ikincil sinyal moleküllerinde bir yapı taşıdır. siklik adenozin monofosfat (cAMP) ATP'den türetilmiştir. CAMP'nin kullanıldığı belirli bir durum, cAMP'ye bağlı sinyalleşme yolları. CAMP sinyal yollarında, uyarıcı veya inhibe edici bir hormon reseptörü, bir sinyal molekülü. Bu reseptörler, uyarıcı veya inhibe edici bir düzenleyici ile bağlantılıdır. G-proteini. Uyarıcı bir G-proteini aktive edildiğinde, adenilil siklaz katalizler Mg kullanarak cAMP'ye ATP2+ veya Mn2+. İkincil bir haberci olan cAMP, daha sonra etkinleştirmeye devam eder protein kinaz A, hangisi bir enzim hücreyi düzenleyen metabolizma. Protein kinaz A, metabolik enzimleri şu yolla düzenler: fosforilasyon Bu, orijinal sinyal molekülüne bağlı olarak hücrede bir değişikliğe neden olur. Bunun tersi, inhibe edici bir G-proteini etkinleştirildiğinde meydana gelir; G-proteini adenilil siklazı inhibe eder ve ATP, cAMP'ye dönüştürülmez.

Metabolizma
Riboz, hücre içi enerji transferlerindeki rolü nedeniyle "moleküler para birimi" olarak anılır.[kaynak belirtilmeli ] Örneğin, nikotinamid adenin dinükleotid (NAD), flavin adenin dinükleotid (FAD) ve nikotinamid adenin dinükleotid fosfat (NADP) tümü şunları içerir: d- ribofuranose parça. Her biri olabilir elde edilen d- dönüştürüldükten sonra ribose d-riboz 5-fosfat enzim tarafından ribokinaz.[22][23] NAD, FAD ve NADP, biyokimyasal alanda elektron alıcısı olarak hareket eder redoks dahil olmak üzere ana metabolik yolaklardaki reaksiyonlar glikoliz, sitrik asit döngüsü, mayalanma, ve elektron taşıma zinciri.

Nükleotid biyosentezi
Nükleotidler kurtarma yoluyla sentezlenir veya de novo sentezi.[24] Nükleotid kurtarma önceden yapılmış nükleotidlerin parçalarını kullanır ve bunları ileride kullanılmak üzere yeniden sentezler. De novo olarak, amino asitler, karbon dioksit, folat türevleri ve fosforibosil pirofosfat (PRPP) nükleotidleri sentezlemek için kullanılır.[24] Hem de novo hem de kurtarma, ATP'den ve riboz 5-fosfattan sentezlenen PRPP'yi gerektirir. PRPP sentetaz.[24]
Değişiklikler
Doğadaki değişiklikler
Ribokinaz dönüşümünü katalize eder d-bozmak d-riboz 5-fosfat. Dönüştürüldükten sonra, d-ribose-5-fosfat, amino asitler triptofan ve histidin veya kullanım için pentoz fosfat yolu. Emilimi dİnce bağırsaklarda riboz% 88-100'dür (200 mg / kg · saate kadar).[25]
Riboz molekülünün C2 'konumunda önemli bir değişiklik meydana gelir. Ekleyerek O-alkil grubu, RNA'nın nükleer direnci, ilave stabilize edici kuvvetler nedeniyle artar. Bu kuvvetler, artış nedeniyle dengeleniyor. molekül içi hidrojen bağı ve artış glikosidik bağ istikrar.[26] Ortaya çıkan direnç artışı, yarı ömür nın-nin siRNA ve hücrelerde ve hayvanlarda potansiyel terapötik potansiyel.[27] metilasyon belirli bölgelerdeki riboz, bağışıklık uyarılmasındaki bir azalma ile ilişkilidir.[28]
Sentetik modifikasyonlar
Fosforilasyon ile birlikte ribofuranoz molekülleri oksijenlerini selenyum ve kükürt sadece 4 'konumunda değişen benzer şekerler üretmek için. Bu türevler daha fazla lipofilik orijinal molekülden daha fazla. Artan lipofiliklik, bu türleri aşağıdaki gibi tekniklerde kullanım için daha uygun hale getirir. PCR, RNA aptameri değişiklik sonrası, antisens teknolojisi ve aşamalı olarak X-ışını kristalografik veri.[27]
Doğadaki 2 'modifikasyonlarına benzer şekilde, ribozun sentetik bir modifikasyonu, aşağıdakilerin eklenmesini içerir: flor 2 'konumunda. Bu florlanmış riboz, metillenmiş riboza benzer şekilde hareket eder, çünkü DNA ipliğindeki ribozun konumuna bağlı olarak bağışıklık stimülasyonunu baskılayabilir.[26] Metilasyon ve florlama arasındaki büyük fark, ikincisinin yalnızca sentetik modifikasyonlar yoluyla gerçekleşmesidir. Florin ilavesi, glikosidik bağın stabilizasyonunda bir artışa ve molekül içi hidrojen bağlarında bir artışa yol açar.[26]
Tıbbi kullanımlar
d-ribose, yönetiminde kullanım için önerilmiştir konjestif kalp yetmezliği[29] (yanı sıra diğer kalp hastalığı türleri) ve kronik yorgunluk sendromu (CFS), açık etiketli kör olmayan, randomize olmayan ve çapraz olmayan öznel bir çalışmada miyaljik ensefalomiyelit (ME) olarak da adlandırılır.[30]
Tamamlayıcı d-ribose, pentoz fosfat yolu üretmek için enerji üreten bir yol d-riboz-5-fosfat. Enzim glikoz-6-fosfat-dehidrojenaz (G-6-PDH) genellikle hücrelerde yetersizdir, ancak daha çok hastalıklı dokularda olduğu gibi miyokardiyal kalp hastalığı olan hastalarda hücreler. Arzı d-de riboz mitokondri doğrudan ATP üretimi ile ilişkilidir; azaldı d-ribose arz, üretilen ATP miktarını azaltır. Çalışmalar, takviyenin ddoku iskemisini takiben riboz (örneğin miyokardiyal iskemi) miyokardiyal ATP üretimini ve dolayısıyla mitokondriyal fonksiyonu arttırır. Esasen, tamamlayıcı yönetimi d- riboz, alternatif bir 5-fosfo kaynağı sağlayarak pentoz fosfat yolundaki enzimatik bir adımı atlar.d-ribose 1-pirofosfat ATP üretimi için. Tamamlayıcı driboz, insanlarda ve diğer hayvanlarda hücresel hasarı azaltırken ATP seviyelerinin iyileşmesini artırır. Bir çalışma, tamamlayıcı kullanımının d-ribose örneğini azaltır anjina, göğüs ağrısı teşhis edilen erkeklerde koroner arter hastalığı.[31] d-Ribose birçok kişiyi tedavi etmek için kullanılmıştır. patolojik kronik yorgunluk sendromu gibi durumlar, fibromiyalji ve miyokardiyal disfonksiyon. Aynı zamanda egzersiz sonrası kramp, ağrı, sertlik vb. Semptomları azaltmak ve atletik performansı iyileştirmek için kullanılır.[kaynak belirtilmeli ].
Referanslar
- ^ Merck Endeksi: Kimyasallar, İlaçlar ve Biyolojik Ürünler Ansiklopedisi (11. baskı), Merck, 1989, ISBN 091191028X, 8205
- ^ Weast, Robert C., ed. (1981). CRC El Kitabı Kimya ve Fizik (62. baskı). Boca Raton, FL: CRC Press. s. C-506. ISBN 0-8493-0462-8.
- ^ Fischer, Emil; Pilotluk, Oscar (1891). "Ueber eine neue Pentonsäure und die zweite inaktif Trioxyglutarsäure" [Yeni bir pentonik asit ve ikinci inaktif trioksiglutarik asit hakkında]. Berichte der deutschen chemischen Gesellschaft (Almanca'da). 24 (2): 4214–4225. doi:10.1002 / cber.189102402322.
- ^ Levene, P.A.; Jacobs, W. A. (1909). "Über Inosinsäure" [İnosik asit hakkında]. Berichte der deutschen chemischen Gesellschaft (Almanca'da). 42 (1): 1198–1203. doi:10.1002 / cber.190904201196.
- ^ Levene, P.A.; Jacobs, W. A. (1909). "Über die Pentose in den Nucleinsäuren" [Nükleik asitlerdeki pentoz hakkında]. Berichte der deutschen chemischen Gesellschaft (Almanca'da). 42 (3): 3247–3251. doi:10.1002 / cber.19090420351.
- ^ a b Jeanloz, Roger W.; Fletcher, Hewitt G. (1951). "Riboz Kimyası". İçinde Hudson, Claude S.; Cantor, Sidney M. (editörler). Karbonhidrat Kimyasındaki Gelişmeler. 6. Akademik Basın. s. 135–174. doi:10.1016 / S0096-5332 (08) 60066-1. ISBN 9780080562650. PMID 14894350.
- ^ Nechamkin Howard (1958). "Kimyasal terminolojinin bazı ilginç etimolojik türevleri". Bilim eğitimi. 42 (5): 463–474. Bibcode:1958SciEd..42..463N. doi:10.1002 / sce.3730420523.
- ^ a b c d Dewick, Paul M. (2013). "Bir Nükleofil Olarak Oksijen: Hemisetaller, Hemiketaller, Asetaller ve Ketaller". Organik Kimyanın Temelleri: Eczacılık, Tıbbi Kimya ve Biyolojik Kimya Öğrencileri İçin. John Wiley & Sons. s. 224–234. ISBN 9781118681961.
- ^ Leigh, Jeffery (Temmuz – Ağustos 2012). "IUPAC Dışı İsimlendirme Sistemleri". Kimya Uluslararası. Uluslararası Temel ve Uygulamalı Kimya Birliği. 34 (4). Alındı 15 Aralık 2019.
- ^ a b c Butani, S.P. (2019). "Aldopentozlar - Nükleik Asitlerin Şekerleri". Biyomoleküllerin Kimyası (2. baskı). CRC Basın. s. 63–65. ISBN 9781000650907.
- ^ a b Drew, Kenneth N .; Zajicek, Jaroslav; Bondo, Gail; Bose, Bidisha; Serianni, Anthony S. (Şubat 1998). "13C-etiketli aldopentozlar: heteronükleer 1D ve 2D NMR spektroskopisi ile siklik ve asiklik formların tespiti ve miktar tayini ". Karbonhidrat Araştırması. 307 (3–4): 199–209. doi:10.1016 / S0008-6215 (98) 00040-8.
- ^ de Wulf, P .; Vandamme, E.J. (1997). "ᴅ-Ribozun Mikrobiyal Sentezi: Metabolik Deregülasyon ve Fermantasyon Süreci". Uygulamalı Mikrobiyolojideki Gelişmeler. 44: 167–214. doi:10.1016 / S0065-2164 (08) 70462-3. ISBN 9780120026449.
- ^ Tumbula, D. L .; Teng, Q .; Bartlett, M. G .; Whitman, W. B. (1997). "Riboz biyosentezi ve Methanococcus maripaludis'te ortak aromatik amino asit yolunda alternatif bir ilk adım için kanıt". Bakteriyoloji Dergisi. 179 (19): 6010–6013. doi:10.1128 / jb.179.19.6010-6013.1997. PMC 179501. PMID 9324245.
- ^ Wulf, P. De; Vandamme, E.J. (1997). "Fermantasyon yoluyla d -riboz üretimi". Uygulamalı Mikrobiyoloji ve Biyoteknoloji. 48 (2): 141–148. doi:10.1007 / s002530051029. PMID 9299771. S2CID 34340369.
- ^ Steigerwald, Bill; Jones, Nancy; Furukawa, Yoshihiro (18 Kasım 2019). "Meteorlarda Şekerlerin İlk Tespiti Hayatın Kökeni İle İlgili İpuçları Veriyor". NASA. Alındı 18 Kasım 2019.
- ^ Furukawa, Yoshihiro; Chikaraishi, Yoshito; Ohkouchi, Naohiko; Ogawa, Nanako O .; Glavin, Daniel P .; Dworkin, Jason P .; Abe, Chiaki; Nakamura, Tomoki (2019). "Dünya dışı riboz ve ilkel göktaşlarındaki diğer şekerler". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 116 (49): 24440–24445. Bibcode:2019PNAS..11624440F. doi:10.1073 / pnas.1907169116. PMC 6900709. PMID 31740594.
- ^ a b c d Bloomfield, Victor; Crothers, Donald; Tinoco, Ignacio (2000). Nükleik Asitler: Yapılar, Özellikler ve Fonksiyonlar. Üniversite Bilim Kitapları. pp.19 –25.
- ^ Voet, Donald; Voet Judith (2011). Biyokimya. John Wiley & Sons, Inc. s.1152, 1153. ISBN 978-0470570951.
- ^ Foloppe, Nicolas; MacKerell, Alexander D. (Ağustos 1998). "Nükleik Asitlerin Deoksiriboz ve Riboz Gruplarının Konformasyonel Özellikleri: Bir Kuantum Mekanik Çalışma". Fiziksel Kimya B Dergisi. 102 (34): 6669–6678. doi:10.1021 / jp9818683. ISSN 1520-6106.
- ^ "Nükleik asit mimarisi". fbio.uh.cu. Alındı 8 Ekim 2019.
- ^ Neidle Stephen (2008). "DNA ve RNA'nın Yapı Taşları". Neidle'de, Stephen (ed.). Nükleik Asit Yapısının Prensipleri. Akademik Basın. pp.20 –37. doi:10.1016 / B978-012369507-9.50003-0. ISBN 9780123695079.
- ^ Bork, Peer; Sander, Chris; Valencia, Alfonso (1993). "Farklı protein kıvrımlarında benzer enzimatik fonksiyonun yakınsak evrimi: Şeker kinazların heksokinaz, ribokinaz ve galaktokinaz aileleri". Protein Bilimi. 2 (1): 31–40. doi:10.1002 / pro.5560020104. PMC 2142297. PMID 8382990.
- ^ Park, Jae; Gupta, Radhey S. (2008). "Adenosin kinaz ve ribokinaz - RK protein ailesi". Hücresel ve Moleküler Yaşam Bilimleri. 65 (18): 2875–2896. doi:10.1007 / s00018-008-8123-1. PMID 18560757. S2CID 11439854.
- ^ a b c Puigserver, Pere (2018). "Sinyal İletimi ve Metabolomik". Hoffman, Ronald'da; Benz, Edward J .; Silberstein, Leslie E .; Heslop, Helen E. (editörler). Hematoloji (7. baskı). Elsevier. s. 68–78. doi:10.1016 / B978-0-323-35762-3.00007-X. ISBN 9780323357623.
- ^ "Bitkisel İlaçlar, Takviyeler A-Z Endeksi". PDRHealth.com. PDR, LLC. Arşivlenen orijinal 11 Ekim 2008.
- ^ a b c Hamlow, Lucas; O, Chenchen; Fan, Lin; Wu, Ranran; Yang, Bo; Rodgers, M. T .; Berden, Giel; Oomens, J. (Haziran 2015). Irmpd Eylem Spektroskopisi ile Belirlenen Sitidin 2'-Riboz Modifikasyonlarının Yapısal Etkileri. 70. Uluslararası Moleküler Spektroskopi Sempozyumu. Urbana-Champaign'deki Illinois Üniversitesi. Bibcode:2015isms.confEMI13H. doi:10.15278 / isms.2015.MI13.
- ^ a b Evich, Marina; Spring-Connell, Alexander M .; Germann, Markus W. (27 Ocak 2017). "Modifiye riboz şekerlerin nükleik asit yapısı ve işlevi üzerindeki etkisi". Heterosiklik İletişim. 23 (3): 155–165. doi:10.1515 / hc-2017-0056. ISSN 2191-0197. S2CID 91052034.
- ^ Peacock, Hayden; Fucini, Raymond V .; Jayalath, Prasanna; Ibarra-Soza, José M .; Haringsma, Henry J .; Flanagan, W. Michael; Willingham, Aarron; Beal, Peter A. (2011). "Nükleobaz ve Riboz Modifikasyonları, MicroRNA-122-mimetik RNA ile İmmünostimülasyonu Kontrol Ediyor". Amerikan Kimya Derneği Dergisi. 133 (24): 9200–9203. doi:10.1021 / ja202492e. PMC 3116021. PMID 21612237.
- ^ Omran, Heyder; McCarter, Dean; St Cyr, John; Lüderitz, Berndt (2004). "ᴅ-Riboz, konjestif kalp yetmezliği hastalarına yardımcı olur". Deneysel ve Klinik Kardiyoloji. Yaz (9 (2)): 117–118. PMC 2716264. PMID 19641697.
- ^ Teitelbaum, Jacob E .; Johnson, Clarence; St Cyr, John (26 Kasım 2006). "Kronik yorgunluk sendromu ve fibromiyaljide ᴅ-riboz kullanımı: bir pilot çalışma". Alternatif ve Tamamlayıcı Tıp Dergisi. 12 (9): 857–862. CiteSeerX 10.1.1.582.4800. doi:10.1089 / acm.2006.12.857. PMID 17109576.
- ^ "Riboz". wa.kaiserpermanente.org. Alındı 7 Ekim 2019.
Türleri karbonhidratlar | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genel | |||||||||||||||
| Geometri | |||||||||||||||
| Monosakkaritler |
| ||||||||||||||
| Çoklu |
| ||||||||||||||
| |||||||||||||||