Oviraptor - Oviraptor - Wikipedia
| Oviraptor | |
|---|---|
 | |
| Holotip örneği Oviraptor | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Üst aile: | †Caenagnathoidea |
| Aile: | †Oviraptoridae |
| Cins: | †Oviraptor Osborn, 1924 |
| Türler | |
| †Oviraptor philoceratops Osborn, 1924 | |
| Eş anlamlı | |
| |
Oviraptor ("yumurta ele geçiren" veya "yumurta hırsızı" anlamına gelir) cins nın-nin oviraptorid Dinozor yaşadı Asya esnasında Geç Kretase dönem. İlk kalıntılar Djadokhta Formasyonu nın-nin Moğolistan 1923'te liderliğindeki bir paleontolojik keşif sırasında Roy Chapman Andrews ve ertesi yıl cins ve tür türler Oviraptor philoceratops tarafından adlandırıldı Henry Fairfield Osborn. Cins adı, yumurta çalma alışkanlıklarına ilişkin ilk düşünceye atıfta bulunur ve özel ad, bu görüşü güçlendirmek için bir tercih olduğunu gösterir. Ceratopsian yumurtalar. Bu cinse çok sayıda örnek atıfta bulunulmasına rağmen, Oviraptor sadece tek bir kısmi iskeletten bilinmektedir. holotip yanı sıra yuva yaklaşık on beş yumurta ve bir yavrudan birkaç küçük parça.
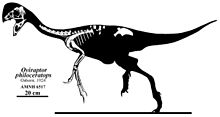
Oviraptor oldukça küçüktü tüylü oviraptorid, uzunluğu 1,6 m (5,2 ft), ağırlığı 33–40 kg (73–88 lb) arasındadır. Kafatasına sahip geniş bir alt çenesi ve muhtemelen tepesi vardı. Hem üst hem de alt çeneler dişsiz ve bir azgın geliştirdi gaga, alt çenelerin sağlam morfolojisi boyunca beslenme sırasında kullanıldı. Kollar iyi gelişmiş ve uzamış, kavisli pençeli üç parmakla son bulmuştur. Diğer oviraptoridler gibi, Oviraptor ilk ayak parmağıyla dört parmaklı ayakları olan uzun arka ayakları vardı indirgenmiş. Kuyruk muhtemelen çok uzun değildi ve Pygostyle büyük tüyleri destekleyen.
İlk ilişkileri Oviraptor o zamanlar yeterince anlaşılmamıştı ve ilgisiz Ornithomimidae orijinal tanımlayıcı Henry Osborn tarafından. Ancak, yeniden yapılan incelemeler Rinchen Barsbold Kanıtlandı Oviraptor ayrı bir aileyi garanti edecek kadar farklıydı, Oviraptoridae. İlk tarif edildiğinde, Oviraptor holotipin bir dinozor yuvasıyla yakın ilişkisi göz önüne alındığında, yumurta hırsızı, yumurta yiyen bir dinozor olarak yorumlandı. Bununla birlikte, yuvalama pozlarındaki çok sayıda oviraptorozorun bulguları, bu örneğin aslında kara kara düşünen yuva ve çalmak ya da yumurtaları beslememek. Dahası, holotip numunesi ile ilişkili olarak küçük bir yavru veya yavruya ait kalıntıların keşfi rapor edilmiştir. ebeveyn bakımı.
Keşif tarihi

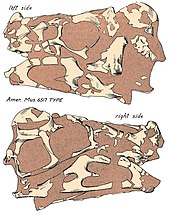
İlk kalıntıları Oviraptor kırmızımsı üzerinde keşfedildi kumtaşları of Geç Kretase Djadokhta Formasyonu nın-nin Moğolistan Bayn Dzak mevkiinde (aynı zamanda Alevli Kayalıklar ), 1923 yılında bir paleontolojik keşif gezisi sırasında. Bu sefer, Kuzey Amerikalı doğa bilimci Roy Chapman Andrews ve bilimde yeni olan üç theropod fosil kalıntısının keşfiyle sona erdi - bunlar da dahil Oviraptor. Bunlar resmen Kuzey Amerika tarafından tanımlandı paleontolog Henry Fairfield Osborn 1924'te, yeni materyalin temelinde cins adını veren Oviraptor, Saurornithoides ve Velociraptor. Belirli cins Oviraptor tip türler ile dikildi O. philoceratops holotipe dayalı AMNH 6517, iskeletin arkasına sahip olmayan ancak kötü bir şekilde ezilmiş olan kısmi bir birey kafatası, kısmi servikal ve sırt omurları dahil pektoral elemanlar furcula sol kol ve kısmi ellerle, sol ilium ve bazı pirzola. Buna göre, bu örnek bir yuva yaklaşık 15 yumurtalar - AMNH 6508 olarak kataloglanmış bir yuva - kafatasının yumurtalardan yalnızca 10 cm (100 mm) tortu. Her iki numunenin yakınlığı göz önüne alındığında, Osborn yorumladı Oviraptor yumurta yeme alışkanlığına sahip bir dinozor olarak ve jenerik adının, Oviraptor, dır-dir Latince fosillerin birleşmesi nedeniyle "yumurta ele geçiren" veya "yumurta hırsızı" için. Özel isim, Philoceratops, aynı zamanda yuva ile ilgili ilk düşüncenin bir sonucu olarak verilen "ceratopsian yumurtalarına düşkünlük" olarak tasarlanmıştır. Protoceratops veya başkası Ceratopsian. Ancak Osborn, adın Oviraptor bu dinozorla ilgili yanlış bir algıyı yansıtabilir. Dahası, Osborn şunu buldu: Oviraptor dişsiz çeneleri temel alan ve ilgili olmayan - o sırada ilişkili olduğu düşünülen - hızlı hareket eden ornitomimidlere benzer olması ve Oviraptor için Ornithomimidae.[1] Osborn daha önce taksonu şu şekilde rapor etmişti: "Fenestrosaurus philoceratops",[2] ama bu daha sonra itibarını yitirdi.[1]

1976'da Moğol paleontoloğu Rinchen Barsbold taksonomik yerleşimiyle ilgili bazı tutarsızlıklar kaydetti Oviraptor ve anatomik özelliklere göre bu taksonun ornitomimidlerden oldukça farklı olduğu sonucuna varmıştır. Bu düşüncenin altında, Oviraptoridae içermek Oviraptor ve yakın akrabalar.[3] Osborn'un ilk açıklamasından sonra Oviraptorholotip ile ilişkili yumurta yuvasının ait olduğu kabul edildi. Protoceratops,[4][5] ve oviraptoridlerin büyük ölçüde yumurta yiyen terapodlar olduğu düşünülüyordu.[6] Bununla birlikte, 1990'larda, çok sayıda yuvalama ve yuva yapan oviraptorid örneğinin keşfi, Osborn'un adıyla ilgili ihtarında haklı olduğunu kanıtladı. Oviraptor. Bu bulgular, oviraptoridlerin kara kara kara kara kara düşündüklerini ve üzerlerine çömelerek yuvalarını koruduklarını gösterdi. Bu yeni kanıt çizgisi, yuvanın holotipiyle ilişkili olduğunu gösterdi. Oviraptor ona aitti ve numune aslında ölüm anında yumurtaları avlamak yerine kuluçkaya yatıyordu.[7][8][9]
Yönlendirilen örnekler

Oviraptoridae'nin 1976'da adlandırılmasından sonra Barsbold, altı ek örneği daha OviraptorMPC-D 100/20 ve 100/21 numarası altında iki özel numune dahil.[3] 1986'da Barsbold, son ikisinin cinse ait olmadığını ve bunun yerine yeni bir oviraptoridi temsil ettiklerini fark etti: Conchoraptor.[10] Diğer örneklerin çoğunun da ait olma olasılığı düşüktür. Oviraptor kendisi ve diğer oviraptoridlere atanmıştır.[11][12] Kısmi bir birey de yumurtaları ile Bayan Mandahu Formasyonu Moğolistan, 1996 yılında Dong Zhiming ve Philip J. Currie IVPP V9608 örneği.[9] Ancak, 2010'da Nicholas R. Longrich ve son iki paleontolog, el falanks oranları gibi çeşitli anatomik farklılıklar olduğu için bu sevkle ilgili belirsizliklerini ifade etmişlerdir. Bu örneğin, bu taksona atfedilemeyen farklı ve belirsiz bir tür olduğu sonucuna vardılar.[13] 1981'de Barsbold, MPC-D 100/42 örneğini Oviraptor, Djadokhta Formasyonundan çok iyi korunmuş ve oldukça eksiksiz bir birey.[14] Bilinen unsurlarından beri Oviraptor MPC-D 100/42, diğer üyelere kıyasla o kadar parçalıydı ki, bu taksonun ana referansı / tasviri oldu. Oviraptor philoceratops içinde Bilimsel edebiyat.[15][11]
Bu anlayış, tarafından reddedildi James M. Clark ve 2002 yılında, bu uzun tepeli örneğin kafatasının daha fazla özelliğine sahip olduğunu kaydeden meslektaşları, Citipati ile yaptığından Oviraptor- aslında, bir sorguç korumaz - ve birincisinin ikinci bir türünü veya tamamen yeni bir cinsi temsil edebilir.[16] 1986'da Barsbold, ikinci bir tür Oviraptor, "O. mongoliensis", MPC-D 100 / 32a örneğine göre, Nemegt Oluşumu.[10] Bununla birlikte, 1997'de Barsbold tarafından yapılan bir yeniden inceleme, bu örnekte yeni cinsi adlandırmaya yetecek kadar farklılık buldu. Rinchenia ama bunu formaliteyle tanımlamadı ve bu yeni oviraptorid bir nomen dubium.[17] Bu, tarafından değiştirildi Lehçe paleontolog Halszka Osmólska ve taksonu resmi olarak adlandırarak 2004'te Rinchenia mongoliensis.[18] Kuzey Amerikalı paleontolog Mark A. Norell ve meslektaşları 2018'de yeni bir örnek bildirdi Oviraptor: Bir tibia ve bir yavru veya çok küçük bir gencin iki metatarsalından oluşan AMNH 33092. AMNH 33092, holotip ile ilişkili bulundu ve muhtemelen yuvanın bir parçasıydı. Oviraptor artık ilişkili yumurtalarla birlikte holotipten ve bir yavru / yuvadan bilinmektedir.[19]
Açıklama
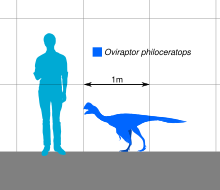
Holotip numunesinin uzunluğu 1,6 m (5,2 ft) ve ağırlığı 33 ila 40 kg (73 ila 88 lb) arasında tahmin edilmiştir.[20][21][22] Holotip, iskeletin arka bölgesinden büyük ölçüde yoksun olsa da, muhtemelen Oviraptor üç işlevsellikle sonuçlanan iki iyi gelişmiş arka ayağı vardı ayak parmakları ilki olan körelmiş yanı sıra nispeten azaltılmış kuyruk. İlgili olarak kanıtlandığı gibi Oviraptoridler kollar uzamış tüyler ve kuyruk bir Pygostyle, bir tüy hayranını desteklediği biliniyor.[23][24]
kafatası nın-nin Oviraptor derin ve kısaltılmıştı Fenestrae (açıklıklar) diğer dinozorlara kıyasla ve korunduğu sürece yaklaşık 17,9 cm (179 mm) boyutundadır. Holotip kafatasının premaksilla gibi birkaç bölgeden yoksun olduğu göz önüne alındığında, gerçek uzunluk aslında daha uzun olabilir. Holotip kafatasının neredeyse tamamı bir tepeden yoksundur, ancak kaynaşmış paryetal ve ön kemiklerin üst yüzeyleri, kafatasının desteklediği, muhtemelen iyi gelişmiş bir tepeye sahip olduğunu gösterir. burun ve premaksilla kemikleri (esas olarak ikincisi) kürsü. Oviraptor diğer oviraptoridlerin oldukça tıknaz çenelerine kıyasla daha uzun bir burun ile sonuçlanabilecek uzun bir maksilla ve diş dişine sahipti. damak rijittir, çene hattının altına uzanır ve premaksilladan oluşur, kusmuklar, ve üst çene. Diğer oviraptoridlerde olduğu gibi, damakta aşağıya doğru yönlendirilmiş bir çift diş benzeri çıkıntıya sahip olabilir. Diğer oviraptoridlerde olduğu gibi, burun delikleri (dış burun delikleri) nispeten küçük olacak ve kafatasının yukarısına yerleştirilmiş olacaktı. Oviraptor vardı dişsiz sağlam biten çeneler, papağan -sevmek Rhamphotheca (azgın gaga). Diş ucunun eğriliği aşağıya dönmüştü ancak diğer oviraptoridlerden daha az belirgindi. Citipati. Genel olarak alt çene, 19,5 cm'yi (195 mm) kaplayan kısa ve derin bir kemiktir.[16]

Çoğu oviraptorda olduğu gibi, holotipin sinir dikenleri boyun omurları omurlar kısaydı ve sinir kemerleri X şeklindeydi. Ancak dikenler arka omurlarda daha belirgin hale gelir. İlk servikal vertebranın zigapofizleri birbirine paralel olarak yapılandırılmıştır ve postzygapophyses, çoğunlukla benzer şekilde orta hattan önemli ölçüde farklı görünmemektedir. Citipati. servikal kaburga holotipte omurlarla kaynaşmıştır.[25][19] Sinir dikenleri ön dizinin dikdörtgendir. sırt omurları yandan görüldüğünde ve servikallerin dikenlerinden daha büyük olduğunda. En öndeki dorsal omurda, aşağıdakilere benzer birkaç pleurocoels (küçük hava aralıklı delikler) bulunabilir. Khaan.[25]
furcula nın-nin Oviraptor diğer oviraptoridlerden hipokleidyumun ön yüzeyinde orta hat omurgasına sahip olması bakımından çok farklıdır - furkulanın merkezinde aşağıya doğru bir çıkıntı. Bu kemik V şeklindedir, enine kesiti yuvarlatılmıştır, uzun sivri uç benzeri bir hipokleidyumu korur ve interklaviküler açı yaklaşık 90 ° 'dir.[26] skapulocoracoid holotipte kaynaşmıştır, ancak korakoid ağır hasar görmüş. kürek kemiği hafif eğimli olup 23 cm (230 mm) uzunluğundadır. Oviraptor şunlardan oluşan nispeten uzun bir kolu vardı humerus, yarıçap, ulna, ve Manus. Falangeal formülü Oviraptor diğer birçok theropod ve oviraptoridde görüldüğü gibi 2-3-4 idi. Eli Oviraptor üç sıska ve kuş benzeri parmağı vardı, her parmak yan yana düzleştirilmiş ve kıvrık toynaklılar (pençe kemik). Bazı oviraptoridlerin aksine, Oviraptor Birincisine göre ikinci ve üçüncü parmakta bir azalma olmadı.[1][25][27]
Saygın çocuk Oviraptor AMNH 33092, metatars III ve IV ile bir sağ kaval kemiği içeren arka bacak materyalini korur. Kaval kemiği 5.87 cm (58.7 mm) uzunluğundadır ve holotipten önemli ölçüde daha küçük bir bireyi gösterir.[19] Yuva AMNH 6508 korur uzatma taşı her yumurtanın 14 cm (140 mm) uzunluğunda olduğu yumurtalar (bazıları eksiktir). Bununla birlikte, tafonomik kırmanın onları 2 cm'ye (20 mm) kadar sıkıştırmış olma ihtimali vardır.[7][28]
Sınıflandırma
Oviraptor başlangıçta müttefikti ornitomimidler Dişsiz gagasından dolayı Osborn tarafından. Osborn ayrıca şunlarla benzerlikler buldu: Chirostenotes hala yakın akrabası olarak kabul edilen Oviraptor.[1] 1976'da, Barsbold içermek için yeni bir aile kurdu Oviraptor ve yakın akrabası Oviraptor tip cins of Oviraptoridae.[3] Holotip kafatasının 2002'de Clark ve meslektaşları tarafından yeniden tanımlanması sırasında, Oviraptor nispeten uzun bir maksilla ve diş dişine sahipti. Bu özellikler türetilmiş oviraptoridlerde daha az belirgindir ve şunu önerir: Oviraptor Oviraptoridae'nin yakın tabanına aittir.[16]
kladogram aşağıda bir analizi takip eder Gregory F. Funston ve meslektaşları 2020'de:[29]
| Oviraptoridae |
| ||||||||||||||||||||||||
Paleobiyoloji
Besleme

İlk kez 1924'te Osborn tarafından tanımlandığında, Oviraptor başlangıçta bir yumurtalık - yumurta bazlı bir diyete sahip bir organizma - holotipin ait olduğu düşünülen bir yuva ile ilişkisine dayanan yaşam tarzı Protoceratops.[1] 1977'de Barsbold, ezici bir çene hipotezi önerdi. Sağlam alt çenelerin gücünün ve muhtemelen Rhamphotheca (azgın gaga) kabuğunu kıracak kadar güçlüydü yumuşakçalar gibi istiridye ile aynı jeolojik oluşumda bulunan Oviraptor. Bu kemikler ana üst çene kemiğinin bir parçasını oluşturur veya üst çene, ortada birleşerek bir çift çatal oluşturur. Rhamphotheca ve alt çeneler, birkaç kemiğin uzamasıyla birlikte damak, delici bir alet yapardı. Barsbold ayrıca oviraptoridlerin bir yarı sulu yumuşakça temelli diyet, burun boşluklarının yüksek konumu, kuyrukta artırılmış kas yapısı ve ilk kılavuzun daha büyük boyutuna dayalı yaşam tarzı hane.[30] 1990 konferans özetinde, David K.Smith osteolojik bir yeniden değerlendirmeyi sundu. Oviraptor Barsbold tarafından yapılan açıklamaları reddettiği yerde. Uzmanlaşmış bir ön ayağı gösteren hiçbir kanıt bulamadı. suda hareket ve çeneler, bir kırma mekanizmasını korumak yerine kesme yüzeylerini korur. Kafatası olduğu gibi dişsiz Smith, hafif yapılı ve birkaç güçlü kas ekleme alanından yoksun, yapraklar diyetin önemli bir parçası olabilir Oviraptor.[31] Ancak, 1995'te Norell ve meslektaşları, bir kertenkele holotip numunesinin vücut boşluğunda, Oviraptor kısmen etobur.[32]

2008 yılında, Stig Olav K. Jansen, birkaç oviraptorid türünün kafatasını, kuşlar ve kaplumbağalar hangi özelliklerin bir rhamphothecayı öngörebileceğini araştırmak. Oviraptoridlerin alt çenelerinin, papağanlar ve üst çenelerin kaplumbağalarınkine daha çok benzemesi. Bu gözlemlere dayanarak Jansen, oviraptoridlerin her yerde yaşayan keskin gelişmiş rhamphotheca, öne çıkan ön ayaklarla birlikte küçük yakalamak ve yırtmak için uyarlanmış olacağından Av. Dahası, damağın sivri çıkıntıları avı tutmaya katkıda bulunurdu. Jansen, tamamen otçul Oviraptoridlerdeki diyet, düz ve geniş olmadıkları için olası görünmemektedir. Tomia (çenelerin kenarlarını keser) çiğnemek ve alt çeneleri yana doğru hareket ettiremedi. Bununla birlikte, alt çenelerin yumurta gibi en azından ezilmiş unsurlara sahip olacak kadar güçlü olduğunu düşünüyordu. Fındık veya diğer zor tohumlar.[33]
Longrich ve meslektaşları 2010 yılında da bir durofajlı (kabuk kırma uygulayan hayvanlar) hipotezi, bu tür hayvanların tipik olarak geliştiği göz önüne alındığında diş geniş kırma yüzeyleri ile. Alt çenelerdeki diş kemiklerinin sivri şekli, oviraptoridlerin bunun yerine yiyecekleri kesmek için kullanılan keskin kenarlı bir ramphotheca'ya sahip olduğunu göstermektedir. sempatik Diş hekiminin ön tarafındaki (kemik birleşmesi) bölge ezme için bir miktar yetenek vermiş olabilir, ancak bu nispeten küçük bir alan olduğu için muhtemelen çenelerin ana işlevi değildi. Yumuşakçaları yiyenlerin aleyhine olan bir başka argüman da, çoğu oviraptorid'in sedimanlar Çoğunlukla kurak veya yarı kurak ortamları temsil ettiği şeklinde yorumlananlar, örneğin Oviraptor içinde Djadokhta Formasyonu. Ekip ayrıca oviraptoridlerin ve dinnodontlar kısa, derin ve dişsiz çeneler gibi kafatası özelliklerini paylaşmak; uzatılmış diş hekimliği semptomları; uzun mandibular açıklıklar; ve sivri bir damak. Oviraptoridlere benzeyen çeneli modern hayvanlar arasında papağanlar ve kaplumbağa; ikinci grup ayrıca palatal bölgede diş benzeri çıkıntılara sahiptir. Longrich ve meslektaşları, oviraptoridler ve otçul hayvanlar arasındaki benzerlikler nedeniyle, diyetlerinin büyük bir kısmının büyük olasılıkla bitki materyalinden oluştuğu sonucuna vardılar. Oviraptoridlerin çeneleri, aşağıdakiler gibi yiyecekleri işlemek için uzmanlaşmış olabilir: kserofitik bitki örtüsü - az suya sahip ortamlar için adapte edilmiş bir bitki örtüsü - kurak ortamlarında büyümüş olabilir, ancak paleoflora hakkında çok az şey bilindiği için bunu göstermek mümkün değildir. Gobi Çölü.[13]
Ancak 2018'de Funston ve meslektaşları ezici çene hipotezini destekledi. Tıknaz olduğunu belirttiler kürsü ve oviraptoridlerin sağlam alt çeneleri, aslında, papağanlarınkine oldukça benzeyen güçlü ve kıstırıcı bir ısırığa işaret eder. Funston ve meslektaşları, oviraptoridlerin bu anatomik özelliklerinin bir meyveli fındık ve tohum içeren diyet.[34]
Üreme


Tanımından beri embriyonik Citipati 1994'te oviraptoridler daha anlaşılır hale geldi: yumurta yiyen hayvanlar olmak yerine, aslında kara kara kara kara kara kara kara kara düşündüler ve yuvalar. Bu örnek, holotipinin Oviraptor muhtemelen bir cinsel olgun ölen birey kuluçka ilişkili yuva yumurtalar.[7] Oviraptoridler üzerindeki bu yeni davranış, yetişkin bir yuvalama örneğinin raporu ve kısa açıklamasıyla daha net hale geldi. Citipati 1995'te Norell ve arkadaşları tarafından. Örnek üstte bulundu yumurta manşonları arka ayakları yuvanın her iki yanında simetrik olarak çömelmiş ve ön ayakları yuvanın çevresini kaplamıştır. Bu derin düşünme duruşu bugün yalnızca modern kuş dinozorları ve ikinci grup arasında davranışsal bir bağı destekler ve kuş olmayan dinozorlar.[8] 1996'da Dong ve Currie, yeni bir yuvalama oviraptorid örneğini tanımladı. Bayan Mandahu Formasyonu. Yaklaşık 6 yumurtanın oluşturduğu bir yuvanın üzerinde korunmuş halde bulunmuş ve bunlar, dairesel desenli höyük şeklinde bir yapıya yerleştirilmiştir. Numune, ön ayakları yumurtaları kaplayan ve kısmen korunmuş arka ayağı yuvanın merkezine yakın bir şekilde yuvanın üzerinde bulunduğundan, Dong ve Currie yuvanın bir yuva tarafından yakalanıp gömüldüğünü öne sürdüler. kum fırtınası inkübasyon sırasında. Oviraptoridlerin yumurta hırsızı olma olasılığını dışladılar, çünkü yuvayı bir kum fırtınasına veya başka bir şeye gömülmeden çok önce tüketecek veya içgüdüsel olarak terk edeceklerdi. meteorolojik fenomen.[9]
1999'da Clark ve ekibi daha önce bildirilen Citipati yuvalama örneği ve kısaca holotip örneğini tartıştı Oviraptor ve yuva AMNH 6508 ile ilişkisi. Holotipin yuva üzerinde bulunduğu kesin konumun, aralarında ayrıldıkları için belirsiz olduğuna işaret ettiler. hazırlık ve yuva, ikisi hasar görmüş yaklaşık 15 yumurta ile tam olarak tamamlanmış görünmüyor. Dahası, yuvanın yarım daire şeklindeki düzenlemesi, yumurtaların çiftler halinde ve en az üç halka halinde yerleştirildiğini ve bu yuvanın orijinal olarak dairesel olduğunu, bir höyük.[28]
2004'te Thomas P. Hopp ve Mark J. Orsen, kanat ve kuyruğun uzama ve gelişim nedenlerini değerlendirmek için, oviraptoridler de dahil olmak üzere, soyu tükenmiş ve mevcut dinozor türlerinin kuluçka davranışlarını analiz ettiler. tüyler. En eksiksiz oviraptorid yuvalama örneğinin - o zamanki 1995 Citipati yuva örneği — çok kuş benzeri bir duruşta, ön ayakları yakın katlanmış bir duruşta ve göğüs bölgesi, göbek ve ayaklar yumurtalarla temas halinde bulundu, Hopp ve Orsen, uzun tüyler ve bir tüy kaplama büyük olasılıkla hayatta mevcuttu. Oviraptoridlerin "kanatları" ve kuyruğu, yumurtalar için koruma sağlardı ve yavrular gibi iklim faktörlerine karşı Güneş ışığı, rüzgar, ve yağmur yağıyor. Bununla birlikte, bu örneğin kolları, bazı modern kuşlarda olduğu gibi aşırı derecede katlanmamıştı, bunun yerine, daha büyük olanı andıran daha genişler. uçamayan kuşlar gibi devekuşu. Kolun uzatılmış konumu, oviraptoridler gibi büyük kavramalarda yuva yaptığı bilinen bu kuşun kuluçka davranışına da benzer. Hopp ve Orsen, yuva yapan oviraptoridlerin ön ayakları pozisyonuna dayanarak, kanat ve kuyruk tüyü uzamasının arkasındaki atasal neden olarak kuluçka yapmayı önerdi, çünkü optimal sağlamaya daha fazla ihtiyaç vardı. koruma yumurtalar ve yavrular için.[35]

2005 yılında Tamaki Sato ve ekibi, olağandışı bir oviraptorid örneğini bildirdi. Nanxiong Oluşumu. Bu yeni örneğin içinde iki yumurta bulunan pelvik bölgeyi korurken bulundu ve bu nedenle kadın. Yumurtaların boyutu ve konumu, oviraptoridlerin iki işlevselliğini koruduğunu göstermektedir. yumurta kanalları ama yumurta sayısını azaltmıştı yumurtlanmış yumurta kanalı başına bir.[36] David J.Varricchio ve meslektaşları, 2008'de yumurtaların nispeten büyük yumurta kavrama boyutunun oviraptoridlerin ve Troodontidler en çok pratik yapan modern kuşlarınkine benzer çok eşli çiftleşme ve geniş erkek ebeveyn bakımı, gibi ratit benzer alışkanlıklar öneren kuşlar. Bu üreme sistemi modern kuşlar için atadan kalma durumu temsil etme olasılığı yüksektir, iki ebeveynin bakımı (her iki ebeveynin de katıldığı) daha sonraki bir gelişmedir.[37] 2014 yılında W. Scott Persons ve meslektaşları, oviraptorosaurların ikincil olarak uçamadığını ve kuyruklarındaki bazı özelliklerin bir eğilimin göstergesi olabileceğini öne sürdü. görüntüleme davranışı, gibi kur gösterimi. Birkaç oviraptorosaur ve oviraptorid'in kuyruğu, pijamalar Kuyruğun ucunda, en azından modern kuşlarda bir tüy yelpazesini desteklemek için kullanılan kemikli bir yapı. Dahası, kuyruk özellikle kaslıydı ve kur hareketlerine yardımcı olabilecek belirgin bir esnekliğe sahipti.[24]

2018'de Tzu-Ruei Yang ve meslektaşları, kütikül oviraptoridler de dahil olmak üzere maniraptoran dinozorlarının birkaç yumurta kabuğunun katmanları. Bu belirli katmanlar şunlardan oluşur: proteinler, polisakkaritler ve pigmentler, ama esas olarak lipidler ve hidroksiapatit. Modern kuşlarda yumurtaları dehidrasyon ve işgali mikroorganizmalar. Çoğu oviraptorid örneğinin oluşumlarında bulunduğu gibi kaliş Yang ve meslektaşları, kütikül kaplı yumurtaların bu tür kurak iklimlerde ve ortamlarda kuluçka başarılarını artırmak için uyarlanmış bir üreme stratejisi olabileceğini öne sürdüler.[38]
2019'da Yang ve meslektaşları hipotezini yeniden değerlendirdiler. termoregülasyon Nanxiong Formasyonundan alınan tam oviraptorid yuvaları kullanarak temaslı inkübasyon ve oviraptorid kavramanın yapısının ayrıntılı bir yeniden inşasını sağladı. Yuva ile ilişkili yetişkin oviraptorid örneklerinin, yumurtlama sürecinde bir dişiyi temsil edebildikleri için yumurtaları inkübe etmeleri gerekmediğini ve çoklu halkalı kavramanın yeterince önlediğini belirttiler. ısı transferi ebeveynden yumurtaların iç halkalarına kadar. Ortalama bir oviraptorid yuva, oldukça organize bir mimariye sahip, hafif eğimli bir höyük olarak inşa edildi: yumurtalar muhtemelen pigmentliydi ve her bir çift üç ila dört eliptik halka şeklinde düzenlenmiş çiftler halinde düzenlenmişti. Ebeveyn muhtemelen yuva merkezinden çalıştığı için, bu bölgede yumurta yoktu. Yang ve meslektaşları, oviraptorid yuvalama tarzının modern analoglardan yoksun olacak kadar benzersiz olduğu sonucuna vardılar, bu nedenle oviraptorid üremesini kullanmak, kuş üreme stratejilerinin evrimi hakkında bilgi vermek için en iyi örnek olmayabilir. Bununla birlikte, ekip, gencin çocuk olup olmadığını belirleyemedi. Oviraptor AMNH 33092, holotip ile ilişkili yuvadan çıkmıştı.[39]
Paleoçevre
Oviraptor Bayn Dzak yöresinden bilinmektedir. Djadokhta Formasyonu Moğolistan'da, geçmişi Geç Kretase yaklaşık 71 milyon ila 75 milyon yıl önce.[40] Djadokhta Formasyonunun paleo ortamı, bir yarı kurak iklim ile kumul ve alüvyon modern ile benzer ayarlar Gobi Çölü. Yarı kurak bozkır manzara tarafından kurutuldu aralıklı akışlar ve bazen tozdan etkilenmiş ve kum fırtınası ve nem mevsimseldi.[41][42] Bu oluşumun büyük ölçüde oldukça kurak ortamları koruduğu düşünülmesine rağmen, birkaç kısa ömürlü su kütleleri Ukhaa Tolgod bölgesinden bildirilmiştir. akarsu sedimantasyon.[43] Ayrıca, daha sonra Kampaniyen yaş ve içine Maastrihtiyen iklim, daha nemli akarsu ortamına kayacaktır. Nemegt Oluşumu.[42]
Djadokhta Formasyonu bir alt Bayn Dzak Üyesi ve bir üst Turgrugyin Üyesi olarak ayrılmıştır. Bilinen kalıntıları Oviraptor dinozorları da veren Bayn Dzak üyesi tarafından üretilmiştir. Bainoceratops, Pinacosaurus, Protoceratops, Saurornithoides, Velociraptor,[40] ve Halszkaraptor.[44] Bu üyeden gelen diğer dinozor faunası, Ukhaa Tolgod yöresinin faunasını içerir. Apsaravis, Byronosaurus, Citipati, Gobipteryx, Khaan, Khol, Shuuvuia, Tsaagan,[43] ve Minotaurasaurus.[45]
Tafonomi
Holotipinin pozu Oviraptor yumurtaların birlikteliğiyle birlikte, bir süre boyunca yuvanın üzerinde hapsolduğunu öne sürün. kum fırtınası, ve cenaze töreni Vücudun tamamen dağınık olma fırsatı olmadığı göz önüne alındığında nispeten hızlıydı veya atılmış avcılar tarafından.[1][9] Paleontolog Kenneth Carpenter Kum fırtınalarının tortularda bulunan yumurtaların gömülmesinin en olası olayı olabileceği konusunda da hemfikirdi.[46] Elementler arasında, kafatası özellikle düzleşmiş ve şekil değiştirmiştir. fosilleşme süreci.[16]
Ayrıca bakınız
Referanslar
- ^ a b c d e f Osborn, H.F. (1924). "Üç yeni Theropoda, Protoceratops bölgesi, orta Moğolistan". Amerikan Müzesi Novitates (144): 1−12. hdl:2246/3223. OCLC 40272928.
- ^ Osborn, H.F. (1924). "Bilinmeyen bir kıtanın keşfi". Doğal Tarih. 24 (2): 133−149.
- ^ a b c Barsbold, R. (1976). "О новом позднемеловом семей- стве мелких теропод Oviraptoridae fam. Nov Монголии" [Yeni bir Geç Kretase ailesi küçük theropodlar Oviraptoridae n. dostum. Moğolistan'da]. Doklady Akademii Nauk SSSR. 226 (3): 685−688.
- ^ Coombs, W. P. (1989). "Dinozor yuvaları ve ebeveyn davranışları için modern analoglar". Farlow, J. O. (ed.). Dinozorların paleobiyolojisi. Amerika Jeoloji Derneği Özel Kağıt 238. Colorado: Boulder. sayfa 21−54. doi:10.1130 / SPE238-p21.
- ^ Thulborn, R.A. (1992). "Protoceratops dinozor yuvası". Lethaia. 25 (2): 145−149. doi:10.1111 / j.1502-3931.1992.tb01379.x.
- ^ Currie, P. J .; Godfrey, S. J .; Nessov, L.A. (1993). "Kuzey Amerika ve Asya'nın Üst Kretase'sinden yeni caenagnathid (Dinosauria: Theropoda) örnekleri". Kanada Yer Bilimleri Dergisi. 30 (10): 2255−2272. Bibcode:1993CaJES..30.2255C. doi:10.1139 / e93-196.
- ^ a b c Norell, M. A .; Clark, J. M .; Dashzeveg, D .; Barsbold, R .; Chiappe, L. M .; Davidson, A. R .; McKenna, M. C .; Altangerel, P .; Novacek, M.J. (1994). "Bir theropod dinozor embriyosu ve Flaming Cliffs Dinozor yumurtalarının yakınlıkları". Bilim. 266 (5186): 779−782. Bibcode:1994Sci ... 266..779N. doi:10.1126 / science.266.5186.779. JSTOR 2885545. PMID 17730398.
- ^ a b Norell, M. A .; Clark, J. M .; Chiappe, L. M .; Dashzeveg, D. (1995). "Yuvalanan bir dinozor". Doğa. 378 (6559): 774−776. Bibcode:1995Natur.378..774N. doi:10.1038 / 378774a0.
- ^ a b c d Dong, Z .; Currie, P. J. (1996). "Çin Halk Cumhuriyeti, İç Moğolistan, Bayan Mandahu'da bir yumurta yuvasında bir oviraptorid iskeletinin keşfedilmesi üzerine". Kanada Yer Bilimleri Dergisi. 33 (4): 631−636. Bibcode:1996CaJES..33..631D. doi:10.1139 / e96-046.
- ^ a b Barsbold, R. (1986). "Хищные динозавры овирапторы" [Etçil dinozorlar oviraptors]. Vorobyeva, E. I. (ed.). Moğol Halk Cumhuriyeti'nde herpetolojik araştırma (Rusça). Hayvanların Evrimsel Morfolojisi ve Ekolojisi Enstitüsü: Doklady Akademii Nauk SSSR. s. 210–223.
- ^ a b Maryańska, T .; Osmólska, H .; Wolsan, M. (2002). "Oviraptorosauria için Avialan statüsü" (PDF). Acta Palaeontologica Polonica. 47 (1): 97−116.
- ^ Kundrát, M .; Janáček, J. (2007). "Moğolistan'ın Geç Kretase'sinden oviraptorid dinozor Conchoraptor gracilis'in (Theropoda, Maniraptora) kraniyal pnömatizasyon ve işitsel algıları". Naturwissenschaften. 94 (9): 769−778. Bibcode:2007NW ..... 94..769R. doi:10.1007 / s00114-007-0258-7.
- ^ a b Longrich, N. R .; Currie, P. J .; Dong, Z. (2010). "İç Moğolistan, Bayan Mandahu'nun Üst Kretase'sinden yeni bir oviraptorid (Dinosauria: Theropoda)". Paleontoloji. 53 (5): 945−960. doi:10.1111 / j.1475-4983.2010.00968.x.
- ^ Barsbold, R. (1981). "Bedene çürüyen hayvanlar Монголии" [Moğolistan'ın dişsiz etobur dinozorları]. İşlemler, Ortak Sovyet-Moğol Paleontoloji Seferi (Rusça). 15: 28−39. S2CID 195060070. Çeviri kağıt
- ^ Barsbold, R .; Maryańska, T .; Osmólska, H. (1990). "Oviraptorosauria". Weishampel, D. B .; Osmolska, H .; Dodson, P. (editörler). Dinosauria (1. baskı). Berkeley: California Üniversitesi Yayınları. sayfa 249−258. ISBN 9780520067271.
- ^ a b c d Clark, J. M .; Norell, M. A .; Rowe, T. (2002). "Citipati osmolskae'nin (Theropoda, Oviraptorosauria) Kraniyal Anatomisi ve Oviraptor philoceratops Holotipinin Yeniden Yorumlanması" (PDF). Amerikan Müzesi Novitates (3364): 1−24. doi:10.1206 / 0003-0082 (2002) 364 <0001: CAOCOT> 2.0.CO; 2. hdl:2246/2853.
- ^ Barsbold, 1997. Oviraptorosauria. Currie ve Padian'da (editörler). Dinozorlar Ansiklopedisi. 505-509.
- ^ Osmólska, H .; Currie, P. J .; Barsbold, R. (2004). "Oviraptorosauria". Weishampel, D. B .; Dodson, P .; Osmólska, H. (editörler). Dinosauria (2. baskı). Berkeley: California Üniversitesi Yayınları. s. 165−183. ISBN 0520242092.
- ^ a b c Norell, M. A .; Balanoff, A. M .; Barta, D. E .; Erickson, G.M. (2018). "Ukhaa Tolgod, Omnogov Aimag, Moğolistan'dan bir yumurta yuvasıyla ilişkili ikinci bir Citipati osmolskae örneği". Amerikan Müzesi Novitates (3899): 1−44. hdl:2246/6858.
- ^ Werner, J .; Griebeler, E.M. (2013). "Kuş Dışı Dinozorların Üremesine ve Bunların Evrimsel ve Ekolojik Etkilerine İlişkin Yeni Görüşler: Fosil Kanıtlarını Mevcut Yakın Akrabaların Alometrilerine Bağlamak". PLOS ONE. 8 (8): e72862. doi:10.1371 / journal.pone.0072862. PMC 3749170. PMID 23991160.
- ^ Paul, G. S. (2016). Princeton Dinozorlar Saha Rehberi (2. baskı). Princeton, New Jersey: Princeton University Press. s. 178. ISBN 9780691167664.
- ^ Campione, N. E .; Evans, D.C. (2020). "Kuş olmayan dinozorlarda vücut kütle tahmininin doğruluğu ve kesinliği". Biyolojik İncelemeler. doi:10.1111 / brv.12638.
- ^ Fanti, F .; Currie, P. J .; Badamgarav, D .; Lalueza-Fox, C. (2012). "Moğolistan'ın Baruungoyot ve Nemegt Formasyonlarından (Geç Kretase) Nemegtomaia'nın yeni örnekleri". PLOS ONE. 7 (2): e31330. Bibcode:2012PLoSO ... 731330F. doi:10.1371 / journal.pone.0031330. PMC 3275628. PMID 22347465.
- ^ a b Persons, W. S .; Currie, P. J .; Norell, M.A. (2014). "Oviraptorosaur kuyruğu formları ve işlevleri". Acta Palaeontologica Polonica. doi:10.4202 / app.2012.0093.
- ^ a b c Balanoff, A. M .; Norell, M.A. (2012). "Khaan mckennai (Oviraptorosauria, Theropoda) Osteolojisi". Amerikan Doğa Tarihi Müzesi Bülteni (372): 1−77. doi:10.1206/803.1. hdl:2246/6413. OCLC 884738416.
- ^ Nesbitt, S. J .; Turner, A. H .; Spaulding, M .; Conrad, J. L .; Norell, M.A. (2009). "Theropod furcula". Morfoloji Dergisi. 270 (7): 856−879. doi:10.1002 / jmor.10724. PMID 19206153.
- ^ Lü, J .; Currie, P. J .; Xu, L .; Zhang, X .; Pu, H .; Jia, S. (2013). "Orta Çin'den tavuk büyüklüğünde oviraptorid dinozorlar ve bunların ontogenetik etkileri" (PDF). Naturwissenschaften. 100 (2): 165−175. Bibcode:2013NW .... 100..165L. doi:10.1007 / s00114-012-1007-0. PMID 23314810.
- ^ a b Clark, J. M .; Norell, M. A .; Chiappe, L.M. (1999). "Moğolistan'daki Ukhaa Tolgod'un Geç Kretase'sinden bir oviraptorid iskeleti, bir oviraptorid yuva üzerinde kuş benzeri bir kuluçka pozisyonunda korunmuş". Amerikan Müzesi Novitates (3265): 1−36. hdl:2246/3102.
- ^ Funston, G. F .; Tsogtbaatar, C .; Tsogtbaatar, K .; Kobayashi, Y .; Sullivan, C .; Currie, P.J. (2020). "İki parmaklı yeni bir dinozor, Oviraptorosauria'nın radyasyonuna ışık tutuyor". Royal Society Açık Bilim. 7 (10): 201184. doi:10.1098 / rsos.201184.
- ^ Barsbold, R. (1977). "Oviraptorların (Theropoda, Saurischia) çene aparatının kinetik ve tuhaflığı". Sovyet-Moğol Paleontoloji Seferi, Trudy. 4: 37−47. Çeviri kağıt
- ^ Smith, D. K. (1990). "Oviraptor philoceratops Osteolojisi, Moğolistan'ın Üst Kretase'sinden olası bir otçul theropod". Omurgalı Paleontoloji Dergisi. 10 (ek. 003): 42A. doi:10.1080/02724634.1990.10011841.
- ^ Norell, M. A .; Gaffney, E. S .; Dingus, L. (1995). Amerikan Doğa Tarihi Müzesi'nde Dinozorları Keşfetmek. Knopf Inc. s. 225.
- ^ Jansen, S. O. K. (2008). Mevcut kuşlar ve kaplumbağalara dayalı oviraptoridlerde gaga morfolojisi (Tez). Oslo Üniversitesi. s. 1−48. hdl:10852/11785.
- ^ Funston, G. F .; Mendonca, S. E .; Currie, P. J .; Barsbold, R .; Barsbold, R. (2018). "Nemegt Havzasında Oviraptorosaur anatomisi, çeşitliliği ve ekolojisi". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 494: 101−120. Bibcode:2018PPP ... 494..101F. doi:10.1016 / j.palaeo.2017.10.023.
- ^ Hopp, T. P .; Orsen, M.J. (2004). "Dinozor Düşünme Davranışı ve Uçuş Tüylerinin Kökeni" (PDF). Currie, P. J .; Koppelhus, E. B .; Shugar, M. A .; Wright, J. L. (editörler). Tüylü ejderhalar: dinozorlardan kuşlara geçiş üzerine araştırmalar. Bloomington: Indiana University Press. sayfa 234−250.
- ^ Sato, T .; Cheng, Y.-N .; Wu, X.-C .; Zelenitsky, D. K .; Hsiao, Y.-F. (2005). "Bir Dişi Dinozorun İçinde Bir Çift Kabuklu Yumurta" (PDF). Bilim. 308 (5720): 375. doi:10.1126 / science.1110578. PMID 15831749. S2CID 19470371.
- ^ Varricchio, D. J .; Moore, J. R .; Erickson, G. M .; Norell, M. A .; Jackson, F. D .; Borkowski, J. J. (2008). "Avian babalık bakımı dinozor kökenlidir". Bilim. 322 (5909): 1826−1828. Bibcode:2008Sci ... 322.1826V. doi:10.1126 / science.1163245. PMID 19095938.
- ^ Yang, T.-R .; Chen, Y.-H .; Wiemann, J .; Spiering, B .; Sander, P.M. (2018). "Fosil yumurta kabuğu kütikülü, dinozor yuvalama ekolojisini aydınlatır". PeerJ. 6: e5144. doi:10.7717 / peerj.5144. PMC 6037156. PMID 30002976.
- ^ Yang, T.-R .; Wiemann, J .; Xu, L .; Cheng, Y.-N .; Wu, X.-C .; Sander, P.M. (2019). "Oviraptorid kavramaların yeniden yapılandırılması, benzersiz yuvalama biyolojilerini aydınlatıyor". Acta Palaeontologica Polonica. 466: 581−596. doi:10.4202 / app.00497.2018.
- ^ a b Dashzeveg, D .; Dingus, L .; Loope, D. B .; Swisher III, C.C .; Dulam, T .; Sweeney, M.R. (2005). "Üst Kretase Djadokhta Formasyonu için Yeni Stratigrafik Alt Bölüm, Çökelme Ortamı ve Yaş Tahmini, Güney Ulan Nur Havzası, Moğolistan" (PDF). Amerikan Müzesi Novitates (3498): 1−31. doi:10.1206 / 0003-0082 (2005) 498 [0001: NSSDEA] 2.0.CO; 2. hdl:2246/5667.
- ^ Jerzykiewicz, T. (1997). "Djadokhta Oluşumu". Currie, P. J .; Padian, K. (editörler). Dinozorlar Ansiklopedisi. San Diego: Akademik Basın. pp.188 −191. ISBN 978-0-12-226810-6.
- ^ a b Chinzorig, T .; Kobayashi, Y .; Tsogtbaatar, K .; Currie, P. J .; Watabe, M .; Barsbold, R. (2017). "Moğolistan, Tögrögiin Shiree'nin Üst Kretase Djadokhta Formasyonundan İlk Ornitomimid (Theropoda, Ornithomimosauria)". Bilimsel Raporlar. 7 (5835). Bibcode:2017NatSR ... 7.5835C. doi:10.1038 / s41598-017-05272-6. PMC 5517598. PMID 28724887.
- ^ a b Dingus, L .; Loope, D. B .; Dashzeveg, D .; Swisher III, C.C .; Minjin, C .; Novacek, M. J .; Norell, M.A. (2008). "Ukhaa Tolgod Jeolojisi (Djadokhta Formasyonu, Üst Kretase, Nemegt Havzası, Moğolistan)" (PDF). Amerikan Müzesi Novitates (3616): 1−40. doi:10.1206/442.1. hdl:2246/5916.
- ^ Cau, A .; Beyrand, V .; Voeten, D. F. A. E .; Fernandez, V .; Tafforeau, P .; Stein, K .; Barsbold, R .; Tsogtbaatar, K .; Currie, P. J .; Godefroit, P. (2017). "Senkrotron taraması, kuş benzeri dinozorlardan oluşan yeni bir sınıfta amfibi ekomorfolojiyi ortaya çıkarıyor". Doğa. 552 (7685): 395−399. Bibcode:2017Natur.552..395C. doi:10.1038 / nature24679. PMID 29211712.
- ^ Alicea, J .; Loewen, M. (2013). "Djodokta Formasyonundan yeni Minotaurasaurus materyali takson için yeni taksonomik ve stratigrafik kriterler oluşturmaktadır". Omurgalı Paleontoloji Dergisi. Program ve Özetler: 76.
- ^ Carpener, K. (1999). "Fosil Olarak Yumurtalar". Yumurtalar, Yuvalar ve Yavru Dinozorlar: Dinozorların Üremesine Bir Bakış (Geçmişin Hayatı). Bloomington: Indiana University Press. s. 111. ISBN 9780253334978.
Dış bağlantılar
 İle ilgili medya Oviraptor Wikimedia Commons'ta
İle ilgili medya Oviraptor Wikimedia Commons'ta İle ilgili veriler Oviraptor Wikispecies'de
İle ilgili veriler Oviraptor Wikispecies'de- Oviraptor yuva AMNH 6508 fotoğrafları AMNH
- Oviraptor holotip kafatası fotoğrafı AMNH
















