Enantiornithes - Enantiornithes
| Enantiornithes | |
|---|---|
 | |
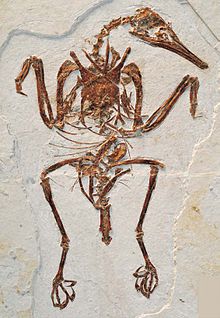
| Bir fosil örneği bohaiornithid (Zhouornis hani ) | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Ornitotoras |
| Clade: | †Enantiornithes Walker, 1981 |
| Alt gruplar | |
ve metni gör | |
Enantiornithes nesli tükenmiş bir grup Avialanlar (geniş anlamıyla "kuşlar"), bilinen en bol ve çeşitli grup Mesozoik dönem.[3][4][5] Neredeyse tüm dişler her kanatta tutulmuş ve parmakları pençeli, ancak bunun dışında dışarıdan modern kuşlara çok benziyordu. 80'den fazla enantiornithe türü adlandırılmıştır, ancak bazı isimler yalnızca tek kemikleri temsil eder, bu nedenle büyük olasılıkla hepsi geçerli değildir. Enantiornitheans'ın nesli tükendi Kretase-Paleojen sınırı, ile birlikte Hesperornithidler ve diğer tüm kuş olmayan dinozorlar.
Keşif ve adlandırma
Keşfedilecek ilk enantiornitheanslar, yanlışlıkla modern kuş gruplarına yönlendirildi. Örneğin, bilinen ilk enantiornithean, Gobipteryx minuta, başlangıçta bir Paleognath ile ilgili devekuşları ve Tinamou.[6] Enantiornitheans ilk olarak kuşların ayrı bir soy veya "alt sınıfı" olarak kabul edildi. Cyril A. Walker 1981'de. Walker bu keşfi, son dönemlerden kalan bazı kısmi kalıntılara dayanarak yaptı. Kretase dönem şimdi ne Arjantin, yeni bir cinse atadığı, Enantiornis, tüm gruba adını veriyor. 1990'lardan bu yana, daha birçok tam enantiornitheans keşfedildi ve daha önce birkaç tanesinin "kuşlar" (ör. Iberomesornis, Cathayornis, ve Sinornis ) ayrıca enantiornitheans idi.
"Enantiornithes" adı "zıt kuşlar" anlamına gelir. Antik Yunan enantios (ἐνάντιος) "zıt" + orniitler (ὄρνιθες) "kuşlar". Adı icat edildi Cyril Alexander Walker grubu kuran dönüm noktası makalesinde.[7] Walker makalesinde "tersi" ile ne demek istediğini açıkladı:
Belki de Enantiornithes ile diğer tüm kuşlar arasındaki en temel ve karakteristik fark, kuşların arasındaki eklemlenmenin doğasındadır. kürek kemiği [...] ve korakoid 'normal' koşulun tamamen tersine döndüğü yer.[7]
Bu anatomik bir özelliği ifade eder - omuz kemikler - modern kuşların tersi olan içbükey dışbükey soket eklemi vardır. Spesifik olarak, enantiornitheans'ta, kürek kemiği (kürek kemiği) karşılar korakoid (memeliler dışındaki omurgalılarda omuz kuşağının birincil kemiği) dışbükey topuz ve omuz bıçağı üzerindeki ilgili nokta içbükey ve tabak şeklindedir. Modern kuşlarda, eklemin eklemlenme biçimi tersine çevrilmiştir.[8]
Walker, makalesinin etimoloji bölümünde bu ismi vermesinin nedenleri konusunda net değildi ve bu belirsizlik daha sonraki araştırmacılar arasında bazı karışıklıklara yol açtı. Örneğin, Alan Feduccia 1996'da belirtilmiştir:
Kuşlar, pek çok ayırt edici özellik arasında benzersiz bir triosseal kanal oluşumu olduğu ve metatarsalların proksimal olarak distal olarak kaynaştığı için bu şekilde adlandırılmıştır, modern kuşların tersi.[9]
Feduccia'nın tarsometatarsus (kombine üst ayak ve ayak bileği kemiği) doğru, ancak Walker bu mantığı orijinal makalesinde kullanmadı. Walker, tarsometatarsus füzyonunu hiçbir zaman zıt olarak değil, "Yalnızca kısmi" olarak tanımladı. Ayrıca, enantiornithe'ların triosseal kanallara sahip oldukları kesin değildir, çünkü hiçbir fosil bu özelliğini korumaz.[3]
Bir grup olarak Enantiornithes'e genellikle "enantiornitinler" adı verilir. Bununla birlikte, birkaç bilim adamı bunun yanlış olduğunu belirtmiştir, çünkü hayvan gruplarının adlarını oluşturmak için standart kuralları takip etmek, yalnızca alt aileye atıfta bulunmak anlamına gelir. Enantiornithinae. Modern kuşlar ve nesli tükenmiş gruplar için kullanılan adlandırma kurallarının ardından, doğru terimin "enantiornithean" olduğu belirtildi.[10]
Kökeni ve aralığı
Enantiornitheanslar hariç her kıtada bulundu Antarktika. Bu gruba atfedilebilecek fosiller, yalnızca Kretase yaşta ve Enantiornitheans'ların, avialan olmayanlarıyla aynı zamanda neslinin tükendiğine inanılıyor. Dinozor akraba. Bilinen en eski enantiornitheanslar, Erken Kretase nın-nin ispanya (Örneğin. Noguerornis ) ve Çin (Örneğin. Protopteryx ) ve en son Geç Kretase Kuzey ve Güney Amerika (ör. Avisaurus ve Enantiornis ). Bu grubun yaygın bir şekilde ortaya çıkması, en azından bazı enantiornitelerin okyanusları kendi güçleri altında geçebildiklerini göstermektedir; küresel bir dağılıma sahip bilinen ilk avialan soyudur.
Açıklama

Çoğu enantiornithean fosili çok parçalıdır ve bazı türler yalnızca tek bir kemik parçasından bilinmektedir. Tam artikülasyonda ve yumuşak doku korumalı hemen hemen tüm numuneler, Las Hoyas içinde Cuenca, ispanya ve Jehol grubu içinde Liaoning (Çin ). Enantiornithean fosilleri, hem iç hem de deniz sedimanlarında bulunmuş ve bu da onların ekolojik olarak çeşitli bir grup olduklarını düşündürmektedir. Enantiornitheans, kuşları, yüzücüler, granivorlar, böcekçiller, balıkçılar ve yırtıcı kuşları içeriyor gibi görünüyor. Enantiornithean türlerinin büyük çoğunluğu küçüktü. serçe ve bir sığırcık,[11] bu sınıfın en büyük üyeleri ise Pengornis houi,[12] Xiangornis shenmi[13] ve Zhouornis hani.[11] Yalnızca ayak izlerinden bilinen potansiyel olarak turna boyutundaki türler de dahil olmak üzere çok daha büyük birkaç tür var olmuş olabilir. Eumeralla Formasyonu (ve muhtemelen aynı zamanda Wonthaggi Oluşumu tek bir furcula ), bir enantiornithean'a ait olabilir.[14] Bir takson, Mirarce, boyut olarak modern ile benzer olarak tanımlanır hindi,[15] önceki "enantiornitheans" olarak tanımlanırken karatavuk boyutlu.[12]
Olağanüstü enantiornithean kalıntıları da korunmuştur. Birman kehribar mevduatlar 99 milyon yıl öncesine aittir. Bu kalıntılar, herhangi bir mezozoik dinozor arasında en iyi korunmuş olanlar arasındadır. Keşfedilen ilk kehribar kaplı enantiornithean kalıntıları, 2016'da açıklanan iki kanattı (aşağıya bakınız).[16] Bir enantiyornitenin neredeyse tamamı 2017'de tanımlandı.[17] ve 2018'de başka bir yavru tanımlandı.[18] 2019'da bir kanatla birlikte büyük ölçüde tam bir ayak tanımlandı.[19] 2020'de büyük bir taksonun kanadı tanımlandı.[20]
Kafatası

Çok çeşitli yaşam alanları ve diyetleri göz önüne alındığında, enantiornitheans kafatasları türler arasında önemli ölçüde değişiklik gösterdi. Enantiornithean kafatasları benzersiz bir ilkel ve gelişmiş özellikler paketini birleştirdi. Daha ilkel avialanlar gibi Archæopteryx, birkaç ayrı kafatası kemiği tuttular, küçük premaksilla (burun ucu kemikleri) ve çoğu türün dişsiz gagalar yerine dişlek çeneleri vardı. Sadece birkaç tür, örneğin Gobipteryx minuta, tamamen dişsiz ve gagalıydı. Ayrıca basitti kemikleri dördül her birini ayıran tam bir çubuk yörünge (göz deliği) her birinden antorbital fenestra ve çatallı arka uçları olmayan diş dişleri (alt çenenin ana dişli kemikleri). Bir skuamozal kemik belirsiz bir genç numunede korunurken, poztorbital içinde korunur Shenqiornis ve Pengornis. Modern kuşlarda bu kemikler kafatasına asimile edilir. Bazı enantiornitheans'ların geçici fenestra (başın yan tarafındaki delikler), postorbitallerin mevcut olmaması veya açıklıkları bölmeye yetecek kadar uzun olmaması nedeniyle modern kuşlarda olduğu gibi yörüngelerle birleşmiştir.[21] Bir kuadratojugal kemik modern kuşlarda jugal ile kaynaşmış olan, Pterygornis.[22] Kafatasının bu ilkel özelliklerinin varlığı, enantiornitheans'ları yalnızca sınırlı kafatası kinesis (çeneyi kafatasından bağımsız olarak hareket ettirebilme yeteneği).[23]
Kanat

Çok büyük bir kuş grubu olarak, enantiornitheans, ekoloji ve beslenmedeki farklılıklara dayanan yüksek çeşitlilikte farklı vücut planları sergilediler, eşit çeşitlilikte kanat formları, modern kuşlarda görülen farklı yaşam tarzlarına birçok paralel uyum sağladı. Genel olarak, enantiornitheans'ın kanatları, daha ilkel avialanlara kıyasla daha gelişmişti. Archæopteryxve modern kuşlara giden soyda bulunanlara benzer uçuşla ilgili bazı özellikler sergiledi. Ornithuromorpha. Enantiornithe'ların çoğunun en azından bazı parmaklarında pençeleri varken, birçok türün ellerini kısaltması, oldukça hareketli bir omuz eklemi ve modern kuşlara benzer şekilde kanat kemiklerinde orantılı değişiklikler vardı. Modern kuşlar gibi, enantiornitheans'ın alulas veya "piç kanatlar", havada daha yüksek manevra kabiliyeti sağlayan ve hassas inişlere yardımcı olan, birinci basamaktaki tüylerin ileriye dönük küçük düzenlemeleri.[24]
Korunmuş tüylere sahip birkaç kanat, Birman kehribar. Bunlar ilk tamamlananlar Mesozoik Dinozor bu şekilde korunmuş kalıntılar (başka türlü biliniyor, herhangi bir türe atanmamış birkaç izole tüy var) ve bilinen en zarif şekilde korunmuş dinozor fosillerinden biri.[25] Korunan kanatlar, tüy pigmentinde farklılıklar gösterir ve enantiornithe'ların dikenler, kılçıklar ve kancalar gibi tamamen modern tüylere ve uzun uçuş tüyleri, kısa örtüler, büyük bir alula ve kuş tüyü astarı dahil modern bir kanat tüyü düzenlemesine sahip olduklarını kanıtlar.[16]
Bir enantiornithean fosili, bacaklarında kanat benzeri tüy öbekleri gösterir. Archæopteryx. Bacak tüyleri de dört kanatlı dinozoru andırıyor Microraptor Bununla birlikte, enantiyorianda tüyler daha kısadır, daha düzensizdir (açıkça bir kanat oluşturmaz) ve ayak boyunca değil, sadece ayak bileğine kadar uzanır.[26]
Kuyruk
Clarke et al. (2006), o zaman bilinen tüm enantiornithean fosillerini araştırdı ve hiçbirinin modern kuşlarda olduğu gibi asansör üreten bir yelpaze oluşturan kuyruk tüylerinin korunmadığı sonucuna vardı. Dışındaki tüm avialanların Euornithes (aradıkları sınıf Ornithurae ) korunmuş kuyruk tüyleri ile sadece kısa örtüler veya uzun çift kuyruk tüyleri vardı. Geliştirilmesini önerdiler. Pygostyle in enantiornitheans, modern bir kuyruk tüyü anatomisinin gelişimi değil, kuyruk kısaltmasının bir işlevi olmalıydı. Bu bilim adamları, kuyruk tüyleri hayranının ve ilgili kasların onları kontrol etmek için gerekli olduğunu ileri sürdüler. dikdörtgen ampul, enantiornitheans gibi daha ilkel avialanlardaki uzun, çubuk veya hançer şeklindeki pygostyles yerine, modern kuşlardakine benzer kısa, üçgen bir pygostyle ile birlikte gelişti. Bir kuş tüyü yelpaze yerine, çoğu enantiornithe'nin soyu tükenmiş olanlara benzer bir çift uzun özel tüyleri vardı. Konfüçyüs ve kesin cennet Kuşları.[27]
Bununla birlikte, daha ileri keşifler, en azından ilkel enantiorniteliler arasında kuyruk anatomisinin daha önce düşünülenden daha karmaşık olduğunu gösterdi. Bir enantiornithean, Shanweiniao, başlangıçta birbiriyle örtüşen en az dört uzun kuyruk tüyüne sahip olarak yorumlandı[28] euornitheans'ın kuyruk fanlarına benzer bir kaldırma oluşturan yüzey oluşturmuş olabilir,[29] daha sonraki bir çalışma şunu gösteriyor Shanweiniao sahip olma olasılığı daha yüksekti Rachis -de bulunan tüylere benzer hakim kuyruk tüyleri Paraprotopteryx.[30] Chiappeavis, ilkel pengornithid enantiornithean, daha ilkel avialanlarinkine benzer kuyruk tüyleri yelpazesine sahipti. Sapeornis, bu durumun atalardan kalma bir durum olabileceğini, küçük tüylerin, erken dönem avialanlarda sergileme amacıyla birkaç kez geliştirilmiş bir özellik olduğunu düşündürmektedir.[30] Başka bir enantiornithean, Feitianius, ayrıca ayrıntılı bir kuyruk tüyleri yelpazesine sahipti. Daha da önemlisi, kuyruğun etrafında korunan yumuşak doku, dikdörtgen bir ampulün kalıntıları olarak yorumlandı ve bu özelliğin aslında modern görünümlü pijamalı türlerle sınırlı olmadığını, daha önce düşünüldüğünden çok daha erken evrilmiş olabileceğini ve birçoğunda mevcut olabileceğini düşündürdü. enantiornitheans.[31] Enantiornithean'ın en az bir cinsi, Cruralispennia, modern görünümlü bir pygostyle'a sahipti ama bir kuyruk yelpazesi yoktu.[32]
Biyoloji
Diyet
Enantiornitliler arasında kafatası şeklinin geniş çeşitliliği göz önüne alındığında, grup içinde birçok farklı beslenme uzmanlığı mevcut olmalıdır. Bazıları Shenqiornis, sert kabuklu omurgasızları yemeye uygun büyük, sağlam çenelere sahipti. İçinde Longipterygids Burunlar uzun ve inceydi, dişleri çenelerin ucuyla sınırlıydı ve muhtemelen çamur-sondalar (küçük dişli türler) ve balıkçılar (büyük dişli türler) idi. Kısa, kör dişleri Pengornis muhtemelen yumuşak gövdeli eklembacaklılarla beslenmek için kullanıldı.[21] Güçlü kancalı pençeleri bohaiornithids küçük ve orta büyüklükteki omurgalıların avcıları olduklarını öne sürüyorlar, ancak sağlam dişleri bunun yerine sert kabuklu hayvanlardan oluşan bir diyet öneriyor.[1]
Birkaç örnek, gerçek mide içeriğini korur. Ne yazık ki bunların hiçbiri kafatasını korumaz, bu nedenle bilinen diyetleri ile burun / diş şekli arasında doğrudan bir ilişki kurulamaz. Eoalulavis kalıntılarına sahip olduğu bulundu dış iskeletler sudan kabuklular sindirim sisteminde korunmuş,[33] ve Enantiophoenix fosilleşmiş kemikler arasında korunmuş kehribar gövdeleri, bu hayvanın modern gibi ağaç özüyle beslendiğini düşündürür. Sapsuckers ve diğer kuşlar. Bitki özü fosilleşir ve kehribar olur.[34] Bununla birlikte, son zamanlarda özün ölüm sonrası hareket ettiği, dolayısıyla gerçek mide içeriğini temsil etmediği öne sürülmüştür. Varsayılan balıkla birleştirildi peletler nın-nin Piscivorenantiornis balık dışkısı olduğu ortaya çıktı, bazı türlerin garip mide içeriği yumurtalıklar ve sözde mide taşı Bohaiornis rastgele mineral olmak, sadece Eoalulavis gerçek mide içeriğini gösterir.[35]
Paravian sindirim sistemleri üzerine yapılan bir araştırma, bilinen enantiornitelerin bir mahsul ve bir taşlıktan yoksun olduğunu, mide taşı kullanmadığını ve pelletleri çıkarmadığını gösteriyor. Bu, farklı diş ve kafatası şekillerinin ima ettiği yüksek diyet çeşitliliği ile çelişkili olarak kabul edilir.[36] bazı modern kuşlar taşlığı kaybetmiş ve yalnızca güçlü mide asitlerine güvenmiş olsa da.[37]
Predasyon
Sanz tarafından bildirilen İspanya'dan bir fosil et al. 2001 yılında, üç farklı türe ait dört tane enantiyornithean iskeletinin kalıntılarını içeriyordu. Bunlar büyük ölçüde eksiksizdir, çok sıkı bir şekilde ilişkilidir ve kısmi sindirimi gösteren kemiklerin yüzeyinde oyuklaşma gösterirler. Yazarlar, bu ilişkinin geri akan bir pelet olduğu ve sindirim ve boyut ayrıntılarına göre, yavruların bir bütün olarak yutulduğu sonucuna varmışlardır. pterosaur ya da küçük Theropod Dinozor. Bu ilk kanıttı Mesozoik avialanlar av hayvanlarıydı ve bazı Mesozoyik pan-kuşları Bugün baykuşlar gibi kusan peletler.[38]
Hayat hikayesi

Bilinen enantiornithean fosilleri arasında yumurtalar,[39][40] embriyolar,[41] ve yavrular.[42] Hâlâ yumurtası içinde kıvrılmış olan bir enantiornithean embriyo, Yixian Formasyonu.[43] Juvenil örnekler, bir faktör kombinasyonu ile tanımlanabilir: Ölüm anında hala kıkırdaktan yapılmış kısımları gösteren kemik uçlarının pürüzlü dokusu, nispeten küçük göğüs kemikleri, büyük kafatasları ve gözler ve henüz birbirine kaynaşmamış kemikler .[44] Bazı kuluçka örneklerine resmi isimler verilmiştir.Liaoxiornis delicatus "; ancak, Luis Chiappe ve meslektaşları, yavrulara dayalı olarak yeni türlerin isimlendirilmesi uygulamasının enantiornitheans araştırmalarına zarar verdiğini düşündüler, çünkü belirli bir genç örneğin hangi yetişkin türlere ait olduğunu belirlemek neredeyse imkansızdır, bu da kuluçka holotipine sahip herhangi bir türü yapar. nomen dubium.[44]
Moğol'un kuluçkalık örnekleriyle birlikte Gobipteryx[45] ve Gobipipus,[46][47] Bu buluntular, enantiyornithean yavrularının iskelet kemikleşmesine, iyi gelişmiş kanat tüylerine ve büyük beyinlere sahip olduğunu göstermektedir. erken dönem veya süper precocial günümüz kuşlarında gelişim modelleri. Başka bir deyişle, enantiornitheans muhtemelen zaten iyi gelişmiş ve koşmaya, yem aramaya ve hatta muhtemelen birkaç günlükken uçmaya hazır yumurtadan çıkmışlardır.[44]
Bu hayvanların büyüme oranlarını belirlemek için enantiornithe kemik histolojisinin analizleri yapılmıştır. 2006 yılı çalışması Concornis kemikler, modern kuşlardan farklı bir büyüme modeli gösterdi; büyüme yumurtadan çıktıktan sonraki birkaç hafta boyunca hızlı olmasına rağmen, muhtemelen acemi Bu küçük tür, muhtemelen birkaç yıl gibi uzun bir süre yetişkin boyutuna ulaşmadı.[48] Diğer çalışmalar, erken dönem kuşlarda olduğu gibi yetişkin boyutuna büyümenin yavaş olduğu görüşünü desteklemiştir ( altricial hızlı bir şekilde yetişkin boyutuna ulaştığı bilinen kuşlar).[33] Çeşitli enantiornitelilerde kemik büyümesi hızına ilişkin araştırmalar, daha küçük türlerin daha büyük olanlardan daha hızlı büyüme eğiliminde olduğunu göstermiştir; bu, daha ilkel türlerde görülen modelin tersidir. Jeholornis ve avialan olmayan dinozorlarda.[49] Bazı analizler kemik histolojisini, enantiornitheans'ın tam olarak kuşa sahip olamayabileceğini endotermi bunun yerine bir ara metabolizma hızı.[50]
Enantiornitheans'ta sömürge yuvalanmasının kanıtı, Geç Kretase (Maastrihtiyen ) nın-nin Romanya.[51] Yuvalama alanlarından elde edilen kanıtlar, enantiornithe'ların yumurtalarını modern gibi gömdüklerini gösteriyor. megapodlar ki bu onların çıkarılmış süperprecocial uyarlamalarıyla tutarlıdır.[52]
Enantiornithean genç tüyler üzerine yapılan bir 2020 çalışması, modern megapodlarla ontolojik benzerlikleri daha da vurgulamaktadır, ancak megapodların karasal yaşam tarzının aksine çoğu enantiyornitenin ağaçta yaşayan doğası gibi birkaç farklılığa dikkat çekmektedir.[53]
Uçuş
Pek çok enantiornithealı, karmaşık kuyruklara sahip olmadıkları ve modern kuşlara kıyasla radikal olarak farklı kanat anatomisine sahip oldukları için, uçuş yeteneklerini test eden birkaç çalışmanın konusu oldular.
Geleneksel olarak, omuz kuşağı anatomisinin daha ilkel olduğu ve yere dayalı bir fırlatma mekanizmasını destekleyemediği varsayıldığından, aşağı uçanlar olarak kabul edilmişlerdir.[54] yanı sıra yokluğu nedeniyle dikdörtgenler birçok türde.[27][29][55]
Bununla birlikte, birkaç çalışma, modern kuşlar gibi, benzer şekilde karmaşık bir sinir sistemine ve kanat tüyü bağlarına sahip, verimli uçucular olduklarını göstermiştir. Ek olarak, karmaşık bir kuyruğun olmaması, kuş uçuşu bir bütün olarak - bazı soyu tükenmiş kuşlar gibi Lithornidler ayrıca karmaşık kuyruk tüylerinden yoksundu ama iyi uçuculardı.[56] ve yere dayalı bir fırlatma yapabilecek gibi görünüyorlar.[57]
Göğüs ve omuz kuşağı anatomisindeki farklılık nedeniyle, birçok enantiornithe, modern kuş türlerinden farklı olarak bir uçuş stili kullandı.[açıklama gerekli ]ancak daha tipik uçuş stilleri de mevcuttu.[58]
En azından Elsornis ikincil olarak olmuş gibi görünüyor uçamayan.[59]
Sınıflandırma
Bazı araştırmacılar sınıftaki enantiornitheans'ı gerçek kuşlarla birlikte sınıflandırır. Aves. Diğerleri daha kısıtlayıcı kullanır taç grubu Aves'in tanımı (yalnızca aşağıdakileri içerir: Neornithes, anatomik olarak modern kuşlar) ve enantiornitheans'ı daha kapsayıcı gruba yerleştirin Avialae. Enantiornitheans daha ileriydi Archæopteryx, Konfüçyüs, ve Sapeornis ama birkaç bakımdan modern kuşlardan daha ilkeldi, belki de ara bir evrimsel yol izliyorlardı.
Bilimsel analizlerin fikir birliği, Enantiornithes'in daha büyük grup içindeki iki ana gruptan biri olduğunu gösteriyor. Ornitotoras. Diğer ornitotorasin grubu Euornithes veya Ornithuromorpha, tüm canlı kuşları bir alt küme olarak içerir. Bu, enantiornitheans'ın kuş evriminin başarılı bir dalı olduğu, ancak modern kuşlara giden soydan tamamen ayrı bir şekilde çeşitlendiği anlamına gelir.[3] Ancak bir çalışma, paylaşılan göğüs anatomisinin bağımsız olarak elde edildiğini ve böyle bir ilişkinin yeniden incelenmesi gerektiğini bulmuştur.[60]
Enantiornithean sınıflandırması ve taksonomisi, tarihsel olarak bir dizi faktör nedeniyle karmaşık hale gelmiştir. 2010 yılında, paleontologlar Jingmai O'Connor ve Gareth Dyke, birçok örneği diğerlerinin kapsamlı bir şekilde değerlendirebileceği kadar ayrıntılı olarak tanımlayamayan bilim adamlarının yaygın uygulamalarına karşı bir dizi eleştiriyi özetledi. Bazı türler, özel koleksiyonlarda tutulan örneklere dayanılarak tanımlanmıştır, bu da önceki bulguların daha fazla incelenmesini veya incelenmesini imkansız hale getirir. Enantiornithes'in dünya çapındaki dağılımı göz önüne alındığında, diğer bilim insanlarının her örneği şahsen incelemesi çoğu zaman mümkün olmadığından ve muhtemelen önemli örnekler üzerinde yayınlanan birçok bilgilendirici olmayan açıklama nedeniyle, bu örneklerin çoğu "işlevsel hale gelir. nomina dubia ".[61] Dahası, pek çok tür, yeterince tanımlanmış olsalar bile bilimsel olarak çok bilgilendirici olmayacak şekilde, son derece parçalı örneklere dayanılarak adlandırılmıştır. Adı geçen tüm enantiornithean türlerin üçte birinden fazlası, yalnızca tek bir kemiğin bir parçasına dayanmaktadır. O'Connor ve Dyke, bu örneklerin Enantiornithes'in zaman aralığı veya coğrafi aralığı hakkındaki bilginin genişletilmesine yardımcı olabilmesine rağmen, onları tanımlamanın önemli olduğunu, bu tür örneklerin isimlendirilmesinin "haksız" olduğunu savundu.[61]
İlişkiler
Enantiornithes kardeş gruptur Euornithes ve birlikte bir clade aranan Ornitotoras (yukarıya bakın). Çoğu filogenetik Çalışmalar, Enantiornithes'i modern kuşlardan ve onların en yakın akrabalarından farklı bir monofiletik grup olarak ortaya çıkardı. Clarke ve Norell tarafından yapılan 2002 filogenetik analizi, enantiornitecilerin sayısını azalttı. otapomorfiler sadece dörde.[62]
Enantiornithe sistematiği, son derece geçicidir ve enantiornitheans'ın aşırı olma eğiliminde olması nedeniyle, herkesin bildiği gibi incelenmesi zordur. homoplastik veya ortak atalardan ziyade yakınsak evrim nedeniyle iskelet özelliklerinin çoğunda birbirine çok benzer.[30] Şimdiye kadar oldukça kesin görünen şey, enantiornitheans içinde, muhtemelen daha gelişmiş Euenantiornithes'e ek olarak bazı küçük bazal soyları içeren alt bölümler olduğudur. Avisauridae'nin bir örnek olarak geçerli bir grup oluşturması muhtemel görünse de, tüm bu soyların karşılıklı ilişkisinin ayrıntıları, aslında çoğunun geçerliliği tartışmalıdır. Filogenetik taksonomistler şimdiye kadar enantiornithe türlerinin sınırlandırmalarını önerme konusunda çok isteksizdi.[63]
Böyle bir tanımlama EuenantiornithesChiappe (2002) tarafından yakın çevredeki tüm türleri içerdiği şeklinde tanımlanmıştır. Sinornis daha Iberomesornis. Çünkü Iberomesornis genellikle en ilkel veya temel enantiornithe olarak bulunur, Euenantiornithes son derece kapsayıcı bir grup olabilir, hariç tüm Enantiornithes'lerden oluşur. Iberomesornis kendisi. Uyumlu olmasına rağmen filogenetik isimlendirme Euenantiornithes'in bu tanımı, bazı araştırmacılar tarafından ciddi şekilde eleştirildi. Paul Sereno, buna "kötü tanımlanmış clade [...] filogenetik bir tanımda kötü bir seçimin iyi bir örneği ".[63]
kladogram aşağıda Wang tarafından yapılan bir analizle bulundu et al. 2015'te Jingmai O'Connor tarafından oluşturulan önceki bir veri kümesinden güncellendi.[22]
| Ornitotoras |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cinslerin listesi
Incertae sedis
Enantiyornit taksonomisinin değerlendirilmesi zordur ve sonuç olarak, grup içindeki birkaç sınıf, filogenetik analizlerle tutarlı bir şekilde bulunur. Enantiornitheans'ların çoğu belirli bir aileye dahil değildir ve bu nedenle burada listelenmiştir. Bunların çoğu euenantiornitheans olarak kabul edildi, ancak bu ismin arkasındaki tartışma, enantiornitheans çalışmalarında tutarlı bir şekilde kullanılmadığı anlamına geliyor.
| İsim | Yıl | Oluşumu | yer | Notlar | Görüntüler |
|---|---|---|---|---|---|
| Abavornis | 1998 | Bissekty Oluşumu (Geç Kretase, Turoniyen -e Konyasiyen ) | Parçalanmış birçok Bissekty enantiornitheans'ından biri, yalnızca korakoidler | ||
| Alethoalaornis | 2007 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Kötü bilinen | ||
| Alexornis | 1976 | La Bocana Roja Formasyonu (Geç Kretase, Kampaniyen ) | Bilinen ilk enantiornitanlardan biri. Bir zamanlar eski bir akraba olduğu düşünülüyordu silindirler ve ağaçkakanlar | ||
| Avimaia | 2019 | Xiagou Formasyonu (Geç Kretase, Aptiyen ) | Bu cinsten bir örnek vücudunda açılmış bir yumurta ile öldü |  | |
| Bauxitornis | 2010 | Csehbánya Formasyonu (Geç Kretase, Santoniyen ) | Parçalı ancak tarsometatarsus yapısında benzersiz |  | |
| Katenolimus | 1998 | Bissekty Oluşumu (Geç Kretase, Turoniyen -e Konyasiyen ) | Parçalanmış birçok Bissekty enantiornitheans'ından biri, yalnızca bir korakoid | ||
| Cathayornis | 1992 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Tanımlanan ilk Jehol biota enantiornitheans'larından biri. Birçok türden bilinmesine rağmen, bazıları artık kendi cinslerine yerleştirilmiştir. Benzer bir görünüşe ve yaşam tarzına sahip olabilir Pitta |  | |
| Concornis | 1992 | Las Hoyas (Erken Kretase, Barremiyen ) | En eksiksiz Las Hoyas enantiornitheans'larından biri |  | |
| Cratoavis[64] | 2015 | Santana Oluşumu (Erken Kretase, Aptiyen ) | Şerit benzeri kuyruk tüyleriyle tamamlanmış, grubun çok iyi korunmuş bir Güney Amerikalı üyesi | ||
| Cruralispennia[32] | 2017 | Huajiying Formasyonu (Erken Kretase, Hauterivian ) | Alışılmadık bir ornitoorf benzeri pygostyle ve fırça benzeri uyluk tüyleri vardı. En eski enantiornitheans'lardan biri |  | |
| Cuspirostrisornis | 1997 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Başlangıçta yanlışlıkla sivri bir gagaya sahip olduğuna inanılıyor | ||
| Dalingheornis | 2006 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | Nedeniyle tırmanışa iyi adapte edilmiştir. heterodaktil ayaklar gibi Trogon | ||
| Dunhuangia[65] | 2015 | Xiagou Formasyonu (Erken Kretase, Aptiyen ) | Ornitoromorfların alışılmadık şekilde hakim olduğu bir bölge olan Changma havzasından bir enantiornithean | ||
| Elbretornis | 2009 | Lecho Oluşumu (Geç Kretase, Maastrihtiyen ) | Sadece kanat kemiklerinden bilinir. Diğer Lecho oluşumu enantiornitheans ile eşanlamlı olabilir | ||
| Elektorornis | 2019 | Burma Kehribar (Geç Kretase, Senomaniyen ) | Uzun bir orta parmak ile amber içinde korunmuş bir ayaktan bilinir | ||
| Elsornis | 2007 | Djadochta Oluşumu (Geç Kretase, Kampaniyen ) | Eksik olmasına rağmen iskeleti üç boyutlu korumaya sahiptir. Kanat oranları nedeniyle muhtemelen uçamaz | ||
| Enantiornis | 1981 | Lecho Oluşumu (Geç Kretase, Maastrihtiyen ) | Sadece birkaç kemikten bilinmesine rağmen, bu cins Enantiornithes'in adaşıdır. Aynı zamanda nesli tükenmeden önce grubun en büyük ve son temsilcilerinden biriydi. | ||
| Eoalulavis | 1996 | Las Hoyas (Erken Kretase, Barremiyen ) | Dahil olmak üzere tüyleri korur. Alula, kanat üzerindeki hava akışını kontrol eden özel bir tüy türü | ||
| Eocathayornis | 2002 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Bir zamanlar bazal yakın akraba olarak kabul edilir Cathayornis şimdi daha uzaktan ilişkili olduğu düşünülse de | ||
| Eoenantiornis | 1999 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | İyi korunmuş ancak filogenetik yerleştirmede tutarsız |  | |
| Evgenavis | 2014 | Ilek Formasyonu (Erken Kretase, Barremiyen ) | Sadece enantiornitheans ile bazı özellikleri paylaşan bir tarsometatarsustan bilinir | ||
| Explorornis | 1998 | Bissekty Oluşumu (Geç Kretase, Turoniyen -e Konyasiyen ) | Parçalanmış Bissekty enantiornitheans'larından sadece biri korakoidler | ||
| Falcatakely | 2020 | Maevarano Formasyonu (Geç Kretase, Maastrihtiyen ) | Modern kuşların aksine "ilkel" bir kafatası düzenlemesine sahip olmasına rağmen, yalnızca tek bir dişe sahip büyük bir burun geliştirdi | ||
| Feitianius[31] | 2015 | Xiagou Formasyonu (Erken Kretase, Aptiyen ) | Çoğu enantiyornitenin çift şerit benzeri tüylerinden farklı olarak ayrıntılı bir kuyruk tüylerine sahipti | ||
| Flexomornis | 2010 | Woodbine Oluşumu (Geç Kretase, Senomaniyen ) | Bulunan en eski Kuzey Amerika kuşçularından biri, ancak parça parça kalıntılardan biliniyor. | ||
| Fortunguavis[66] | 2014 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Ağaçlara tırmanmak için uyarlanmış olabilecek ayaklar ve pençeler dahil olmak üzere sağlam kemiklere sahipti | ||
| Grabauornis[67] | 2015 | Yixian Formasyonu (Erken Kretase, Barremiyen ) | Bu enantiornitenin kanatlarının oranları ve bir alulanın varlığı, bunun iyi bir uçuş olduğunu düşündürür. | ||
| Gracilornis | 2011 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Olası bir akraba Cathayornis karakteristik olarak ince kemiklerle | ||
| Gurilynia | 1999 | Nemegt Oluşumu (Geç Kretase, Maastrihtiyen ) | Az bilinen bir enantiornithean, ama belli ki grubun büyük ve geç hayatta kalan bir üyesi | ||
| Hollanda luceria[68] | 2010 | Barun Goyot Formasyonu (Geç Kretase, Kampaniyen ) | Başlangıçta bir ornitoromorf olarak tanımlanmış, ancak o zamandan beri yakından ilişkili bir enantiornithean olarak yeniden yorumlanmıştır. Lectavis.[69] | ||
| Holbotia[70] | 2015 | Andaikhudag Formasyonu (Erken Kretase, Aptiyen ) | Küçük olarak kabul edildi pterosaur 1977'deki keşfinden bu yana 2015'te resmi bir tanım alana kadar. Eşsiz boyun omurlarına ve ilkel bir damak tadına sahipti. | ||
| Houornis | 1997 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Bir zamanlar şüpheli veya bir tür Cathayornis, 2015 yılında yapılan bir çalışmada bunun geçerli bir cins olduğunu düşünmesine rağmen[71] | ||
| Huoshanornis | 2010 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Elinin ve göğüs kemiğinin yapısı nedeniyle çok manevra kabiliyeti olan bir pilot olabilir. | ||
| Iberomesornis | 1992 | Las Hoyas (Erken Kretase, Barremiyen ) | İyi kalıntılardan bilinen ilk enantiornitanlardan biri. Ayrıca grubun en eski ve en ilkel üyelerinden biri |  | |
| Incolornis | 1998 | Bissekty Oluşumu (Geç Kretase, Turoniyen -e Konyasiyen ) | Parçalanmış Bissekty enantiornitheans'larından sadece biri korakoidler. Bir türün bir zamanlar ait olduğu düşünülüyordu Enantiornis | ||
| Junornis[72] | 2017 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | O kadar iyi korunmuş ki, uçuş düzeni tüylerinin ve kanatlarının oranları kullanılarak yeniden oluşturulabilir. |  | |
| Kizylkumavis | 1984 | Bissekty Oluşumu (Geç Kretase, Turoniyen -e Konyasiyen ) | Parçalanmış birçok Bissekty enantiornitheans'ından biri, yalnızca bir humerus parça | ||
| Largirostrornis | 1997 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Muhtemelen ilgili Cuspirostrisornis veya eşanlamlısı Cathayornis | ||
| Lectavis | 1993 | Lecho Oluşumu (Geç Kretase, Maastrihtiyen ) | Grubun büyük ve uzun bacaklı bir üyesi, orantılı olarak modern kıyı kuşlarına benziyor | ||
| Lenesornis | 1996 | Bissekty Oluşumu (Geç Kretase, Turoniyen -e Konyasiyen ) | Parçalanmış birçok Bissekty enantiornitheans'ından biri, yalnızca bir synsacrum parça. Başlangıçta ait olduğu düşünülen İhtiyornis | ||
| Liaoningornis | 1996 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | Başlangıçta bir ornitozan olduğuna inanılıyordu, ancak şimdi Eoalulavis | ||
| Longchengornis | 1997 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Eşanlamlı olabilir Cathayornis | ||
| Martinavis | 2007 | Grès à Sürüngen Oluşumu, Lecho Oluşumu (Geç Kretase, Maastrihtiyen ) | Sadece humeri'den bilinmesine rağmen, bu cins büyüktü ve geniş bir yelpazede yaşıyordu. | ||
| Microenantiornis | 2017 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Grubun diğer enantiyornitelere kıyasla birkaç ilkel ve türetilmiş özelliğe sahip küçük bir üyesi | ||
| Monoenantiornis[73] | 2016 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | Enantiornitelerde yaşlandıkça çeşitli özelliklerin nasıl geliştiğini gösteren genç bir örnekten bilinmektedir. | ||
| Nanantius | 1986 | Toolebuc Oluşumu (Erken Kretase, Albiyen ) | Parçalı, ancak bir deniz kuşu olabilir, çünkü bu cinsten kalıntılar şu şekilde bulundu: Ichthyosaur bağırsak içeriği | ||
| Noguerornis | 1989 | El Montsec (Erken Kretase, Barremiyen ) | İzlenimlerini korur propatagyum omuzda bir kanadın bir parçasını oluşturan bir deri kanadı | ||
| Orienantius | 2018 | Huajiying Formasyonu (Erken Kretase, Hauterivian ) | Bu cinsten örneklerin birçok yumuşak doku ayrıntısı UV ışığı ile ortaya çıkarıldı. | ||
| Otogornis | 1993 | Yijinholuo Formasyonu (Erken Kretase ) | Kötü bilinen | ||
| Paraprotopteryx | 2007 | Qiaotou üyesi Huajiying Formasyonu (Erken Kretase, Aptiyen ?) | Görünüşe göre, çoğu enantiyornitede olduğu gibi sadece iki yerine dört şerit benzeri kuyruk tüyü vardı | ||
| Parvavis[74] | 2014 | Jiangdihe Oluşumu (Geç Kretase, Turoniyen -e Santoniyen ) | Küçük ama öldüğünde tamamen olgundur. Geç Kretase'ye tarihlenen tek tanımlanmış Çin enantiornithean | ||
| Piscivorenantiornis[75] | 2017 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Son öğün olan, balık kemiklerinden oluşan bir mide içeriğinin üzerinde korunmuş, eklemleri kırılmış bir iskeletten bilinmektedir. | ||
| Protopteryx | 2000 | Huajiying Formasyonu (Erken Kretase, Hauterivian ) | Grubun en eski ve en ilkel üyelerinden biri |  | |
| Pterygornis[22] | 2016 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Bu cinsten bir dezartiküle edilmiş iskelet, kuadratojugal dahil, kafatasının iyi korunmuş kemiklerine sahiptir. | ||
| Qiliania | 2011 | Xiagou Formasyonu (Erken Kretase, Aptiyen ) | Bu cinsin kalıntılarından bazıları iyi korunmuş arka bacakları içerir. tür isimleri, Q. graffini, Adını almıştır Greg Graffin gruptan Kötü din | ||
| Sazavis | 1989 | Bissekty Oluşumu (Geç Kretase, Turoniyen -e Konyasiyen ) | Parçalanmış birçok Bissekty enantiornitheans'ından biri, yalnızca bir tibiotarsus (incik kemiği) | ||
| Shangyang | 2019 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Alışılmadık bir şekilde, bu cinsin premaksillaları kaynaştı | ||
| Sinornis | 1992 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Tanımlanan ilk Jehol biota enantiornitheans'larından biri. Benzer Cathayornis ama genellikle farklı kabul edilir |  | |
| Xiangornis | 2012 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Bu cinsin eli, muhtemelen ornitoflarınkine benziyordu. yakınsak evrim. Grubun büyük bir üyesi | ||
| Yuanjiawaornis[76] | 2015 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | İyi kalıntılardan bilinen enantiyornitanlardan biri | ||
| Yungavolucris | 1993 | Lecho Oluşumu (Geç Kretase, Maastrihtiyen ) | Geniş ve alışılmadık derecede genişti tarsometatarsal (Ayak bileği kemiği) | 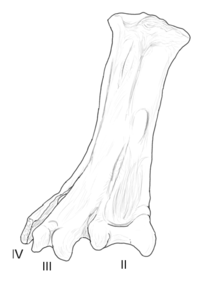 |
Longipterygidae
Longipterygidae dişleri sadece burun uçlarında olan, uzun burunlu erken Kretase enantiornitelerinden oluşan bir aileydi. Genellikle grubun oldukça temel üyeleri olarak kabul edilirler.[28]
| İsim | Yıl | Oluşumu | yer | Notlar | Görüntüler |
|---|---|---|---|---|---|
| Boluochia | 1995 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Başlangıçta yanlışlıkla kancalı bir gagaya sahip olduğuna inanılıyor | ||
| Camptodontornis | 2010 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Başlangıçta aradı Camptodontus, bu cins adı bir böcek | ||
| Dapingfangornis | 2006 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Alnında dikene benzer bir yapı olabilir | ||
| Longipteryx | 2001 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Ailenin en yaygın ve tanınmış üyesi |  | |
| Longirostravis | 2004 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | Diğer longipterygids gibi, çamur veya kabuktaki omurgasızları araştırmak için kullanılmış olabilecek ince bir burnu vardı. | ||
| Rapaxavis | 2009 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Ayaklarının yapısından dolayı tüneme konusunda uzman | ||
| Shanweiniao | 2009 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | Modern kuşlarda olduğu gibi kaldırma oluşturabilen birden fazla kuyruk tüyü elde etti |  | |
| Shengjingornis | 2012 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Ailenin büyük bir üyesi |
Pengornithidae
Pengornithidae büyük erken enantiornitheans ailesiydi. Çok sayıda küçük dişleri ve diğer enantiornitelerin çoğunda kaybolan çok sayıda ilkel özellikleri vardı.[2] Bazı araştırmalar, bunların hiç de enantiornitheans olmadıklarını, daha ziyade modern kuşlara daha yakın ornioromorflar olabileceğini iddia ediyor.
| İsim | Yıl | Oluşumu | yer | Notlar | Görüntüler |
|---|---|---|---|---|---|
| Chiappeavis[30] | 2015 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Birçok tüyden oluşan yelpaze şeklinde bir kuyruğa sahipti | ||
| Eopengornis | 2014 | Huajiying Formasyonu (Erken Kretase, Hauterivian ) | Ailenin bilinen en eski üyesi ve bilinen en eski enantiyanlardandır. Son derece iyi korunmuş kuyruk şeritlerine sahip | ||
| Parapengornis[77] | 2015 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Ayak ve kuyruk özellikleri nedeniyle ağaçkakan benzeri bir yaşam tarzına sahip olması önerildi |  | |
| Pengornis | 2008 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Keşfedilen ilk pengornithid ve aynı zamanda iyi kalıntılardan bilinen en büyük enantiyornitlerden biri |
"Bohaiornithidae"
"Bohaiornithids "büyük ama jeolojik olarak kısa ömürlü, uzun, çengelli pençeleri ve kıvrımlı uçları olan sağlam dişleri olan erken enantiyornitlilerdi. Bu yorum çok tartışmaya açık olsa da, yırtıcı kuşlara eşdeğer olabilirler.[1] monofil bu grup şüphelidir ve aslında bir evrimsel derece.[78]
| İsim | Yıl | Oluşumu | yer | Notlar | Görüntüler |
|---|---|---|---|---|---|
| Bohaiornis | 2011 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Başlangıçta ile korunduğu düşünülen mide taşı, daha sonra bunların mineral betonları olduğu bulundu | ||
| Gretcheniao | 2019 | Yixian Formasyonu (Erken Kretase, Barremiyen ) | Uçmak yerine kanat çırpmak için uyarlanmıştır. "Bohaiornithidae" nin parafili veya polifilini önerebilir | ||
| Linyiornis[79] | 2016 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Ailenin, yumurta geliştirdiğine inanılan yapılarla tamamlanmış, iyi korunmuş bir iskeletten bilinen olası bir üyesi | ||
| Longusunguis | 2014 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Ailenin oldukça tipik bir üyesi |  | |
| Parabohaiornis | 2014 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | Yakın bir akraba Bohaiornis | ||
| Shenqiornis | 2010 | Qiaotou üyesi Huajiying Formasyonu (Erken Kretase, Aptiyen ?) | Ailenin bilinen ilk üyesi, yakın bir akraba olarak görülmese de Bohaiornis birkaç yıl sonrasına kadar. Büyük bir postorbital kemiği korur | ||
| Sulcavis | 2013 | Yixian Formasyonu (Erken Kretase, Aptiyen ) | Yakın bir akraba Shenqiornis oluklu emaye dişlerinde fosil kuşlar arasında eşsiz |  | |
| Zhouornis | 2013 | Jiufotang Formasyonu (Erken Kretase, Aptiyen ) | İyi korunmuş bir beyin kasasına sahip büyük bir aile üyesi |
Gobipterygidae
Bu aile olabilir tek tip (yalnızca bir cins veya türden oluşur), grubun bazı üyeleri belirsiz veya yetersiz tanımlanmış ve tür türleriyle eşanlamlı olabilir, Gobipteryx minuta.
| İsim | Yıl | Oluşumu | yer | Notlar | Görüntüler |
|---|---|---|---|---|---|
| Gobipteryx | 1974 | Barun Goyot Formasyonu (Geç Kretase, Kampaniyen ) | Modern kuşlarınkilerle yakınsayan bir şekilde evrimleşen sağlam bir gagaya sahip, dişsiz, gelişmiş bir enantiornithean | ||
| Jibeinia | 1997 | Qiaotou üyesi Huajiying Formasyonu (Erken Kretase, Aptiyen ?) | Kötü bir şekilde biliniyor ve şu anda kaybolan bir iskeletten tarif ediliyor. İle eşanlamlı olabilir Vescornis | ||
| Vescornis | 2004 | Qiaotou üyesi Huajiying Formasyonu (Erken Kretase, Aptiyen ?) | Küçük ve kısa burunlu bir enantiornithean ile eşanlamlı olabilir Jibeinia |
Avisauridae
Avisauridae farklı kapsayıcılığın iki farklı tanımına tabidir. Cau ve Arduini'yi (2008) izleyen daha kapsamlı tanım burada kullanılmaktadır. Avisauridler, uzun ömürlü ve yaygın enantiornitelerdi; bunlar, esas olarak tarsometatarsallarının (ayak bileği kemikleri) belirli özellikleriyle ayırt edilirler. Grubun en büyük ve en gelişmiş üyeleri, Kuzey ve Güney Amerika'da Kretase'nin sonuna kadar hayatta kaldı, ancak bazı eski taksonlara kıyasla çok parçalı.
| İsim | Yıl | Oluşumu | yer | Notlar | Görüntüler |
|---|---|---|---|---|---|
| Avisaurus | 1985 | Hell Creek Oluşumu (Geç Kretase, Maastrihtiyen ) | İsimsiz avisaurid ve ailenin en büyük üyelerinden biri. Başlangıçta avialan olmayan bir dinozor olarak kabul edildi |  | |
| Enantiophoenix | 2008 | Ouadi al Gabour Formasyonu (Geç Kretase, Senomaniyen ) | Korunduğu için ağaç özüyle beslenmiş olabilir. kehribar boncuklar | ||
| Gettyia | 2018 | İki İlaç Oluşumu (Geç Kretase, Kampaniyen ) | İçin yeni bir cins Avisaurus gloriae | ||
| Halimornis | 2002 | Mooreville Tebeşir Oluşumu (Geç Kretase, Kampaniyen ) | Kıyı ortamında yaşardı | ||
| Intiornis | 2010 | Las Curtiembres Formasyonu (Geç Kretase, Kampaniyen ) | En büyük avisauridlerin bazılarıyla yakından ilişkili olmasına rağmen, bu cinsin üyeleri çok küçük kuşlardı. |  | |
| Mirarce | 2018 | Kaiparowits Oluşumu (Geç Kretase, Kampaniyen ) | En eksiksiz bilinen Kuzey Amerika avisaurid | ||
| Mystiornis | 2011 | Ilek Formasyonu (Erken Kretase, Barremiyen -e Aptiyen ) | Çeşitli gruplardan sayısız özelliğe sahiptir. Paraves, en çok örneklenen gruplar arasında avisauridlere benzese de | ||
| Neuquenornis | 1994 | Bajo de la Carpa Oluşumu (Geç Kretase, Santoniyen ) | İyi uçuş ve tüneme yeteneklerini gösteren uzun kanatlara ve ters halluksa sahip | ||
| Soroavisaurus | 1993 | Lecho Oluşumu (Geç Kretase, Maastrihtiyen ) | Çok yakın bir akraba Avisaurus |
Şüpheli cinsler ve dikkate değer isimsiz örnekler
- Gobipipus reshetovi: 2013 yılında yumurta kabuklarındaki embriyo örneklerinden tanımlanmıştır. Barun Goyot Formasyonu nın-nin Moğolistan. Bu örnekler embriyoniklere çok benziyordu. Gobipteryx örnekler, tanımlayıcıları olmasına rağmen Gobipipus (dahil bir dizi tartışmalı paleontolog Evgeny Kurochkin ve Sankar Chatterjee ) farklı düşünün.[80]
- Hebeiornis Fengningensis: Eşanlamlısı Vescornis aynı örnekten tarif edildiği için. 1999'da tanımlanmış olmasına rağmen, tanımından 5 yıl önce Vescornis, açıklama, açıklamasına kıyasla çok zayıftı Vescornis ikinci adın çoğu yazar tarafından öncelikli olduğu düşünülmektedir. Sonuç olarak, isim Hebeiornis kabul edilir nomen çıplak ("çıplak isim").
- ‘’Proornis ’’, Kuzey Kore'den şüpheli bir kuştur.
- "Liaoxiornis delicatus ": 1999'da enantiyoritik bir örnekten tanımlanmıştır. Yixian Formasyonu. Bu örnek başlangıçta küçük bir yetişkin olarak kabul edildi, ancak daha sonra bir yavru olarak bulundu. Other specimens have henceforth been assigned to the genus. Due to a lack of distinguishing feature, many paleontologists have considered this genus an undiagnostic nomen dubium.
- LP-4450: A juvenile of an indeterminate enantiornithean from the El Montsec Oluşum ispanya. Its 2006 description studied the histoloji of the skeleton, while later studies reported a squamosal bone present in the specimen but unknown in other enantiornitheans.
- IVPP V 13939: Briefly described in 2004, this Yixian enantiornithean had advanced pennaceous feathers on its legs, similar to (albeit shorter than) those of other paravians such as Microraptor ve Anchiornis.[26]
- DIP-V-15100 ve DIP-V-15101: Two different wings from hatchling specimens which were described in 2015. They attracted a significant amount of media attention upon their description. They were preserved in exceptional details due to having been trapped within Burmese amber for approximately 99 million years.[16]
- HPG-15-1: A partial corpse of a hatchling enantiornithean also preserved in Burmese amber. Although indeterminate, it attracted even more media attention than the two wings upon its description in 2017.[17]
- CUGB P1202: An indeterminate juvenile bohaiornithid -den Jiufotang Formasyonu. A 2016 analysis of its feathering found elongated putative melanosomes, suggesting that a large portion of its feathering was yanardöner.
- DIP-V-15102: Another corpse of an indeterminate hatchling preserved in Burmese amber. Described in early 2018.[18]
- MPCM-LH-26189 a/b: A partial skeleton of a hatchling from Las Hoyas in Spain, including both slab and counter-slab components. Its 2018 description revealed how various features developed in enantiornitheans as they aged. Such features include the kemikleşme of göğüs kemiği from various smaller bones, and the fusion of tail vertebrae into a Pygostyle.
- YLSNHM01001: A foot and tail preserved in Burmese amber.[81]
Referanslar
- ^ a b c Wang, Min; Zhou, Zhong-He; O'Connor, Jingmai K .; Zelenkov, Nikita V. (2014). "A new diverse enantiornithine family (Bohaiornithidae fam. nov.) from the Lower Cretaceous of China with information from two new species" (PDF). Vertebrata PalAsiatica. 52 (1): 31–76.
- ^ a b Wang, X .; O'Connor, J. K .; Zheng, X .; Wang, M .; Hu, H.; Zhou, Z. (2014). "Insights into the evolution of rachis dominated tail feathers from a new basal enantiornithine (Aves: Ornithothoraces)". Linnean Society Biyolojik Dergisi. 113 (3): 805–819. doi:10.1111/bij.12313.
- ^ a b c Chiappe, Luis M .; Walker, Cyril A. (2002). "Skeletal Morphology and Systematics of the Cretaceous Euenantiornithes (Ornithothoraces: Enantiornithes)". In Chiappe, Luis M.; Witmer, Lawrence M. (eds.). Mezozoik Kuşlar: Dinozorların Başlarının Üzerinde. California Üniversitesi Yayınları. pp. 240–67. ISBN 978-0-520-20094-4.
- ^ Chiappe, Luis M. (2007). Glorified Dinozorlar: Kuşların Kökeni ve Erken Evrimi. Hoboken, New Jersey: John Wiley and Sons. ISBN 978-0-471-24723-4.[sayfa gerekli ]
- ^ O'Connor, Jingmai K .; Chiappe, Luis M .; Gao, Chunling; Zhao, Bo (2011). "Anatomy of the Early Cretaceous enantiornithine bird Rapaxavis pani" (PDF). Acta Palaeontologica Polonica. 56 (3): 463–75. doi:10.4202/app.2010.0047.
- ^ Elzanowski, Andrzej (1974). "Preliminary note on the palaeognathous bird from the Upper Cretaceous of Mongolia" (PDF). Paleontoloji Polonica. 29: 103–9.
- ^ a b Walker, C.A. (1981). "New subclass of birds from the Cretaceous of South America". Doğa. 292 (5818): 51–3. Bibcode:1981Natur.292...51W. doi:10.1038/292051a0.
- ^ Hope, Sylvia (2002). "The Mesozoic Radiation of Neornithes". In Chiappe, Luis M.; Witmer, Lawrence M. (eds.). Mezozoik Kuşlar: Dinozorların Başlarının Üzerinde. California Üniversitesi Yayınları. pp. 339–88. ISBN 978-0-520-20094-4.
- ^ Feduccia, Alan (1996). Kuşların Kökeni ve Evrimi. New Haven: Yale Üniversitesi Yayınları. ISBN 978-0-300-06460-5.[sayfa gerekli ]
- ^ Sen, Hai-lu; Lamanna, Matthew C .; Harris, Jerald D .; Chiappe, Luis M .; O'Connor, Jingmai; Ji, Shu-an; Lü, Jun-chang; Yuan, Chong-xi; Li, Da-qing; Zhang, Xing; Lacovara, Kenneth J .; Dodson, Peter; Ji, Qiang (16 June 2006). "A Nearly Modern Amphibious Bird from the Early Cretaceous of Northwestern China". Bilim. 312 (5780): 1640–1643. Bibcode:2006Sci...312.1640Y. doi:10.1126/science.1126377. PMID 16778053.
- ^ a b Zhang, Zihui; Chiappe, Luis M .; Han, Gang; Chinsamy, Anusuya (2013). "A large bird from the Early Cretaceous of China: new information on the skull of enantiornithines". Omurgalı Paleontoloji Dergisi. 33 (5): 1176–89. doi:10.1080/02724634.2013.762708.
- ^ a b Zhou, Zhonghe; Clarke, Julia; Zhang, Fucheng (May 2008). "Insight into diversity, body size and morphological evolution from the largest Early Cretaceous enantiornithine bird". Anatomi Dergisi. 212 (5): 565–77. doi:10.1111/j.1469-7580.2008.00880.x. PMC 2409080. PMID 18397240.
- ^ Hu, Dongyu; Xu, Xing; Hou, Lianhai; Sullivan, Corwin (2012). "A New Enantiornithine Bird from the Lower Cretaceous of Western Liaoning, China, and Its Implications for Early Avian Evolution". Omurgalı Paleontoloji Dergisi. 32 (3): 639–45. doi:10.1080/02724634.2012.652321.
- ^ Martin, Anthony J.; Vickers-Rich, Patricia; Rich, Thomas H.; Hall, Michael; Angielczyk, Kenneth (January 2014). "Oldest known avian footprints from Australia: Eumeralla Formation (Albian), Dinosaur Cove, Victoria". Paleontoloji. 57 (1): 7–19. doi:10.1111/pala.12082.
- ^ Atterholt, Jessie; Hutchison, J. Howard; O’Connor, Jingmai K. (13 November 2018). "Kuzey Amerika'dan en eksiksiz enantiornitin ve Avisauridae'nin filogenetik analizi". PeerJ. 6: e5910. doi:10.7717 / peerj.5910. PMC 6238772. PMID 30479894.
- ^ a b c Xing, Lida; McKellar, Ryan C.; Wang, Min; Bai, Ming; O’Connor, Jingmai K .; Benton, Michael J .; Zhang, Jianping; Wang, Yan; Tseng, Kuowei; Lockley, Martin G .; Li, Gang; Zhang, Weiwei; Xu, Xing (28 June 2016). "Mummified precocial bird wings in mid-Cretaceous Burmese amber". Doğa İletişimi. 7 (1): 12089. Bibcode:2016NatCo...712089X. doi:10.1038/ncomms12089. PMC 4931330. PMID 27352215.
- ^ a b Xing, Lida; O'Connor, Jingmai K .; McKellar, Ryan C.; Chiappe, Luis M .; Tseng, Kuowei; Li, Gang; Bai, Ming (September 2017). "A mid-Cretaceous enantiornithine (Aves) hatchling preserved in Burmese amber with unusual plumage". Gondwana Araştırması. 49: 264–277. Bibcode:2017GondR..49..264X. doi:10.1016/j.gr.2017.06.001.
- ^ a b Xing, Lida; O'Connor, Jingmai K .; McKellar, Ryan C.; Chiappe, Luis M .; Bai, Ming; Tseng, Kuowei; Zhang, Jie; Yang, Haidong; Fang, Jun; Li, Gang (February 2018). "A flattened enantiornithine in mid-Cretaceous Burmese amber: morphology and preservation". Bilim Bülteni. 63 (4): 235–243. doi:10.1016/j.scib.2018.01.019.
- ^ Xing, Lida; McKellar, Ryan C.; O’Connor, Jingmai K .; Bai, Ming; Tseng, Kuowei; Chiappe, Luis M. (30 January 2019). "Orta Kretase Burma kehribarında korunmuş tamamen tüylü bir enantiornitin ayak ve kanat parçası". Bilimsel Raporlar. 9 (1): 927. Bibcode:2019NatSR ... 9..927X. doi:10.1038 / s41598-018-37427-4. PMC 6353931. PMID 30700773.
- ^ Xing, Lida; McKellar, Ryan C.; O'Connor, Jingmai K. (June 2020). "An unusually large bird wing in mid-Cretaceous Burmese amber". Kretase Araştırmaları. 110: 104412. doi:10.1016/j.cretres.2020.104412.
- ^ a b O’Connor, Jingmai K .; Chiappe, Luis M. (28 February 2011). "A revision of enantiornithine (Aves: Ornithothoraces) skull morphology". Sistematik Paleontoloji Dergisi. 9 (1): 135–157. doi:10.1080/14772019.2010.526639.
- ^ a b c Wang, Min; Hu, Han; Li, Zhiheng (21 August 2015). "A new small enantiornithine bird from the Jehol Biota, with implications for early evolution of avian skull morphology". Sistematik Paleontoloji Dergisi. 14 (6): 481–497. doi:10.1080/14772019.2015.1073801.
- ^ Wang, Min; Hu, Han (Ocak 2017). "Erken Kuşlar ve Dinosauri Akrabalarında Jugal ve Quadratojugal'ın Karşılaştırmalı Morfolojik İncelemesi". Anatomik Kayıt. 300 (1): 62–75. doi:10.1002 / ar.23446. PMID 28000410.
- ^ Chiappe, Luis M. (2009). "Downsized Dinosaurs: The Evolutionary Transition to Modern Birds". Evrim: Eğitim ve Sosyal Yardım. 2 (2): 248–56. doi:10.1007/s12052-009-0133-4.
- ^ Becker, Rachel (28 June 2016). "Bird wings trapped in amber are a fossil first from the age of dinosaurs". Doğa. doi:10.1038/nature.2016.20162.
- ^ a b Zhang, Fucheng; Zhou, Zhonghe (Ekim 2004). "Palaeontology: Leg feathers in an Early Cretaceous bird". Doğa. 431 (7011): 925. Bibcode:2004Natur.431..925Z. doi:10.1038/431925a. PMID 15496911.
- ^ a b Clarke, Julia A .; Zhou, Zhonghe; Zhang, Fucheng (March 2006). "Çin'den gelen yeni bir Erken Kretase ornitoürin sınıfından kuş uçuşunun evrimine ve Yixianornis grabaui". Anatomi Dergisi. 208 (3): 287–308. doi:10.1111 / j.1469-7580.2006.00534.x. PMC 2100246. PMID 16533313.
- ^ a b O'Connor, Jingmai K .; Wang, Xuri; Chiappe, Luis M .; Gao, Chunling; Meng, Qingjin; Cheng, Xiaodong; Liu, Jinyuan (12 March 2009). "Phylogenetic support for a specialized clade of Cretaceous enantiornithine birds with information from a new species". Omurgalı Paleontoloji Dergisi. 29 (1): 188–204. doi:10.1080/02724634.2009.10010371.
- ^ a b Chiappe, Luis M .; Bo, Zhao; O'Connor, Jingmai K .; Chunling, Gao; Xuri, Wang; Habib, Michael; Marugan-Lobon, Jesus; Qingjin, Meng; Xiaodong, Cheng (2014). "Erken Kretase kuşunun yeni bir örneği Hongshanornis longicresta: insights into the aerodynamics and diet of a basal ornithuromorph". PeerJ. 2: e234. doi:10.7717 / peerj.234. PMC 3898307. PMID 24482756.
- ^ a b c d O’Connor, Jingmai K .; Wang, Xiaoli; Zheng, Xiaoting; Hu, Han; Zhang, Xiaomei; Zhou, Zhonghe (January 2016). "An Enantiornithine with a Fan-Shaped Tail, and the Evolution of the Rectricial Complex in Early Birds". Güncel Biyoloji. 26 (1): 114–119. doi:10.1016/j.cub.2015.11.036. PMID 26748849.
- ^ a b O’Connor, Jingmai K .; Li, Da-Qing; Lamanna, Matthew C .; Wang, Min; Harris, Jerald D .; Atterholt, Jessie; You, Hai-Lu (30 December 2015). "A new Early Cretaceous enantiornithine (Aves, Ornithothoraces) from northwestern China with elaborate tail ornamentation". Omurgalı Paleontoloji Dergisi. 36 (1): e1054035. doi:10.1080/02724634.2015.1054035.
- ^ a b Wang, Min; O’Connor, Jingmai K .; Pan, Yanhong; Zhou, Zhonghe (2017/01/31). "Eşsiz krural tüyleri ve ornitoflu pulluk şeklinde bir pygostyle ile tuhaf bir Erken Kretase enantiornitin kuşu". Doğa İletişimi. 8: 14141. Bibcode:2017NatCo...814141W. doi:10.1038 / ncomms14141. PMC 5290326. PMID 28139644.
- ^ a b Sanz, José L .; Chiappe, Luis M .; Buscalioni, Angela D. (1995). "The Osteology of Concornis lacustris (Aves: Enantiornithes) from the Lower Cretaceous of Spain and a Reexamination of its Phylogenetic Relationships". Amerikan Müzesi Novitates (3133): 1–23. hdl:2246/3667.
- ^ Dalla Vecchia, Fabio M .; Chiappe, Luis M. (2003). "Kuzey Gondwana'nın Mesozoik'inden ilk kuş iskeleti". Omurgalı Paleontoloji Dergisi. 22 (4): 856–60. doi:10.1671 / 0272-4634 (2002) 022 [0856: FASFTM] 2.0.CO; 2. JSTOR 4524284.
- ^ O'Connor, Jingmai K. (1 January 2019). "Erkenci kuşların trofik alışkanlıkları". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 513: 178–195. Bibcode:2019PPP ... 513..178O. doi:10.1016 / j.palaeo.2018.03.006.
- ^ O'Connor, Jingmai K .; Zhou, Zhonghe; Smith, Andrew (6 November 2019). "Modern kuş sindirim sisteminin evrimi: Yanliao ve Jehol biyotalarından gelen paravian fosillerinden bilgiler". Paleontoloji. 63 (1): 13–27. doi:10.1111 / pala.12453.
- ^ Houston, David C.; Copsey, J. A. (1994). "Bone digestion and intestinal morphology of the Bearded Vulture". Raptor Araştırmaları Dergisi. 28 (2): 73–78.
- ^ Sanz, José L .; Chiappe, Luis M .; Fernádez-Jalvo, Yolanda; Ortega, Francisco; Sánchez-Chillón, Begoña; Poyato-Ariza1, Francisco J.; Pérez-Moreno, Bernardino P. (February 2001). "An early Cretaceous pellet". Doğa. 409 (6823): 998–1000. Bibcode:2001Natur.409..998S. doi:10.1038/35059172. PMID 11234054.
- ^ Mikhailov, Konstantin E. (1991). "Amniyotik omurgalıların fosil yumurta kabuklarının sınıflandırılması" (PDF). Acta Palaeontologica Polonica. 36 (2): 193–238.
- ^ Mikhailov, Konstantin E. (1996). "New Genera of Fossil Eggs from the Upper Cretaceous of Mongolia". Paleontological Journal. 30 (2): 246–8.
- ^ Elżanowski, Andrzej (1981). "Embryonic bird skeletons from the late Cretaceous of Mongolia" (PDF). Paleontoloji Polonica. 42: 147–79.
- ^ Sanz, José L .; Chiappe, Luis M .; Pérez-Moreno, Bernardino P .; Moratalla, José J.; Hernández-Carrasquilla, Francisco; Buscalioni, Angela D.; Ortega, Francisco; Poyato-Ariza, Francisco J.; Rasskin-Gutman, Diego; Martı́nez-Delclòs, Xavier (June 6, 1997). "A Nestling Bird from the Lower Cretaceous of Spain: Implications for Avian Skull and Neck Evolution". Bilim. 276 (5318): 1543–6. doi:10.1126/science.276.5318.1543.
- ^ Zhou, Zhonghe; Zhang, Fucheng (22 Ekim 2004). "Çin'in Alt Kretase'sinden Bir Precocial Avian Embriyo". Bilim. 306 (5696): 653. doi:10.1126 / bilim.1100000. PMID 15499011.
- ^ a b c Chiappe, Luis M .; Shu'an, Ji; Qiang, Ji (2007). "Juvenile Birds from the Early Cretaceous of China: Implications for Enantiornithine Ontogeny". Amerikan Müzesi Novitates. 3594: 1–46. doi:10.1206/0003-0082(2007)3594[1:JBFTEC]2.0.CO;2. hdl:2246/5890.
- ^ Elżanowski, Andrzej (1995). "Cretaceous birds and avian phylogeny". Courier Forschungsinstitut Senckenberg. 181: 37–53.
- ^ Kurochkin, E.N .; Chatterjee, S .; Mikhailov, K.E. (2013). "An embryonic enantiornithine bird and associated eggs from the cretaceous of Mongolia". Paleontological Journal. 47 (11): 1252–69. doi:10.1134 / S0031030113110087.
- ^ Kurochkin, E. N.; Chatterjee, S .; Mikhailov, K. E. (19 December 2013). "An embryonic enantiornithine bird and associated eggs from the cretaceous of Mongolia". Paleontological Journal. 47 (11): 1252–1269. doi:10.1134 / S0031030113110087.
- ^ Cambra-Moo, Oscar; Buscalioni, Ángela Delgado; Cubo, Jorge; Castanet, Jacques; Loth, Marie-Madeleine; de Margerie, Emmanuel; de Ricqlès, Armand (2006). "Histological observations of Enantiornithine bone (Saurischia, Aves) from the Lower Cretaceous of Las Hoyas (Spain)". Rendus Palevol Comptes. 5 (5): 685–91. doi:10.1016/j.crpv.2005.12.018.
- ^ O'Connor, Jingmai K .; Wang, Min; Zheng, Xiao-Ting; Wang, Xiao-Li; Zhou, Zhong-He (2014). "The histology of two female Early Cretaceous birds" (PDF). Vertebrata PalAsiatica. 52 (1): 112–28.
- ^ Chiappe, L.M. (1995). "The phylogenetic position of the Cretaceous birds of Argentina: Enantiornithes and Patagopteryx deferrariisi". Courier Forschungsinstitut Senckenberg. 181: 55–63.
- ^ Dyke, Gareth; Vremir, Mátyás; Kaiser, Gary; Naish, Darren (June 2012). "A drowned Mesozoic bird breeding colony from the Late Cretaceous of Transylvania". Die Naturwissenschaften. 99 (6): 435–42. Bibcode:2012NW.....99..435D. CiteSeerX 10.1.1.394.9006. doi:10.1007/s00114-012-0917-1. PMID 22575918.
- ^ Fernández, Mariela S .; Garcia, Rodolfo A .; Fiorelli, Lucas; Scolaro, Alejandro; Salvador, Rodrigo B .; Cotaro, Carlos N .; Kaiser, Gary W.; Dyke, Gareth J .; Farke, Andrew A. (17 April 2013). "A Large Accumulation of Avian Eggs from the Late Cretaceous of Patagonia (Argentina) Reveals a Novel Nesting Strategy in Mesozoic Birds". PLOS ONE. 8 (4): e61030. Bibcode:2013PLoSO...861030F. doi:10.1371/journal.pone.0061030. PMC 3629076. PMID 23613776.
- ^ O'Connor, Jingmai K .; Falk, Amanda; Wang, Min; Zheng, Xiao-Ting (2020). "First report of immature feathers in juvenile enantiornithines from the Early Cretaceous Jehol avifauna". Vertebrata PalAsiatica. 58: 24–44. doi:10.19615 / j.cnki.1000-3118.190823.
- ^ Padian, Kevin; Chiappe, Luis M. (11 January 2007). "Kuşların kökeni ve erken evrimi". Biyolojik İncelemeler. 73 (1): 1–42. doi:10.1111 / j.1469-185x.1997.tb00024.x.
- ^ Zhou, Shuang; Zhou, Zhong-He; O'Connor, Jingmai K. (2012). "A new basal beaked ornithurine bird from the Lower Cretaceous of Western Liaoning, China" (PDF). Vertebrata PalAsiatica. 50 (1): 9–24. Lay özeti – Phys.org (13 Şubat 2012).
- ^ Houde, Peter W. (1988). "Kuzey Yarımküre'nin Erken Tersiyerinden Paleognat Kuşlar". Publications of the Nuttall Ornithological Club (Cambridge Massachusetts, USA: Nuttall Ornithological Club) 22
- ^ Navalón, Guillermo; Marugán-Lobón, Jesús; Chiappe, Luis M .; Luis Sanz, José; Buscalioni, Ángela D. (6 October 2015). "Soft-tissue and dermal arrangement in the wing of an Early Cretaceous bird: Implications for the evolution of avian flight". Bilimsel Raporlar. 5 (1): 14864. Bibcode:2015NatSR...514864N. doi:10.1038/srep14864. PMC 4594305. PMID 26440221.
- ^ Wang, Xia; McGowan, Alistair J.; Dyke, Gareth J .; Turvey, Samuel T. (7 December 2011). "Avian Wing Proportions and Flight Styles: First Step towards Predicting the Flight Modes of Mesozoic Birds". PLOS ONE. 6 (12): e28672. Bibcode:2011PLoSO...628672W. doi:10.1371/journal.pone.0028672. PMC 3233598. PMID 22163324.
- ^ Chiappe, Luis M .; Suzuki, Shigeru; Dyke, Gareth J .; Watabe, Mahito; Tsogtbaatar, K .; Barsbold, Rinchen (January 2007). "Gobi çölünün Geç Kretase döneminden yeni bir Enantiornithine kuşu". Sistematik Paleontoloji Dergisi. 5 (2): 193–208. doi:10.1017/S1477201906001969.
- ^ Zheng, Xiaoting; Wang, Xiaoli; O'Connor, Jingmai; Zhou, Zhonghe (9 October 2012). "Insight into the early evolution of the avian sternum from juvenile enantiornithines". Doğa İletişimi. 3 (1): 1116. Bibcode:2012NatCo...3.1116Z. doi:10.1038/ncomms2104. PMID 23047674.
- ^ a b O'Connor, Jingmai; Dyke, Gareth (2010). "A Reassessment of Sinornis santensis ve Cathayornis yandica (Aves: Enantiornithes)". Avustralya Müzesi Kayıtları. 62: 7–20. doi:10.3853/j.0067-1975.62.2010.1540.
- ^ Clarke, Julia A .; Norell, Mark A. (2002). "The Morphology and Phylogenetic Position of Apsaravis ukhaana from the Late Cretaceous of Mongolia". Amerikan Müzesi Novitates. 3387: 1–46. CiteSeerX 10.1.1.693.8475. doi:10.1206 / 0003-0082 (2002) 387 <0001: TMAPPO> 2.0.CO; 2.
- ^ a b Sereno, P.C. (2005) TaxonSearch: Stem Archosauria Arşivlendi 2007-02-19 Wayback Makinesi. Version 1.0, 2005-NOV- 7. Retrieved 2006-OCT-02.
- ^ Carvalho; Novas; Agnolín; Isasi; Freitas; Andrade (2015). "A new genus and species of enantiornithine bird from the Early Cretaceous of Brazil". Brezilya Jeoloji Dergisi. 45 (2): 161–171. doi:10.1590/23174889201500020001.
- ^ Wang, Li; O'Connor, Zhou; You (2015). "Second species of enantiornithine bird from the Lower Cretaceous Changma Basin, northwestern China with implications for the taxonomic diversity of the Changma avifauna". Kretase Araştırmaları. 55: 56–65. doi:10.1016/j.cretres.2015.01.008.
- ^ Wang, M .; O'Connor, J. K .; Zhou, Z. (2014). "A new robust enantiornithine bird from the Lower Cretaceous of China with scansorial adaptations". Omurgalı Paleontoloji Dergisi. 34 (3): 657–671. doi:10.1080/02724634.2013.812101.
- ^ Dalsätt, J .; Ericson, P. G.; Zhou, Z. (2015). "A New Enantiornithes (Aves) from the Early Cretaceous of China". Acta Geologica Sinica. 86 (2): 801–807. doi:10.1111/1755-6724.12270.
- ^ Bell, Alyssa K .; Chiappe, Luis M .; Erickson, Gregory M .; Suzuki, Shigeru; Watabe, Mahito; Barsbold, Rinchen; Tsogtbaatar, K. (February 2010). "Gobi Çölü'nden (Moğolistan) bir Geç Kretase kuşu olan Hollanda luceria'nın tanımı ve ekolojik analizi". Kretase Araştırmaları. 31 (1): 16–26. doi:10.1016/j.cretres.2009.09.001.
- ^ Hartman, Scott; Mortimer, Mickey; Wahl, William R .; Lomax, Dean R .; Lippincott, Jessica; Lovelace, David M. (10 Temmuz 2019). "Kuzey Amerika'nın Geç Jura döneminden yeni bir paravya dinozoru, kuş uçuşunun geç kazanılmasını destekliyor". PeerJ. 7: e7247. doi:10.7717 / peerj.7247. PMC 6626525. PMID 31333906.
- ^ Zelenkov, Nikita V.; Averianov, Alexander O. (13 June 2015). "A historical specimen of enantiornithine bird from the Early Cretaceous of Mongolia representing a new taxon with a specialized neck morphology". Sistematik Paleontoloji Dergisi. 14 (4): 319–338. doi:10.1080/14772019.2015.1051146.
- ^ Wang, M .; Liu, D. (2015). "Taxonomical reappraisal of Cathayornithidae (Aves: Enantiornithes)". Sistematik Paleontoloji Dergisi. 14: 1–19. doi:10.1080/14772019.2014.994087.
- ^ Liu, Di; Chiappe, Luis M .; Serrano, Francisco; Habib, Michael; Zhang, Yuguang; Meng, Qinjing; Shawkey, Matthew (11 October 2017). "Flight aerodynamics in enantiornithines: Information from a new Chinese Early Cretaceous bird". PLOS One. 12 (10): e0184637. Bibcode:2017PLoSO..1284637L. doi:10.1371/journal.pone.0184637. PMC 5636078. PMID 29020077.
- ^ Hu, Han; O'Connor, Jingmai K. (14 November 2016). "First species of Enantiornithes from Sihedang elucidates skeletal development in Early Cretaceous enantiornithines". Sistematik Paleontoloji Dergisi. 15 (11): 909–926. doi:10.1080/14772019.2016.1246111.
- ^ Wang, Min; Zhou, Zhonghe; Xu, Guanghui (7 January 2014). "The first enantiornithine bird from the Upper Cretaceous of China". Omurgalı Paleontoloji Dergisi. 34 (1): 135–145. doi:10.1080/02724634.2013.794814.
- ^ Wang, Min; Zhou, Zhonghe (12 April 2017). "A morphological study of the first known piscivorous enantiornithine bird from the Early Cretaceous of China". Omurgalı Paleontoloji Dergisi. 37 (2): e1278702. doi:10.1080/02724634.2017.1278702.
- ^ Hu, Dongyu; Liu, Ying; Li, Jinhua; Xu, Xing; Hou, Lianhai (July 2015). "Yuanjiawaornis viriosus, gen. et sp. nov., a large enantiornithine bird from the Lower Cretaceous of western Liaoning, China". Kretase Araştırmaları. 55: 210–219. doi:10.1016/j.cretres.2015.02.013.
- ^ Hu, Han; O’Connor, Jingmai K .; Zhou, Zhonghe; Farke, Andrew A. (3 June 2015). "A New Species of Pengornithidae (Aves: Enantiornithes) from the Lower Cretaceous of China Suggests a Specialized Scansorial Habitat Previously Unknown in Early Birds". PLOS One. 10 (6): e0126791. Bibcode:2015PLoSO..1026791H. doi:10.1371/journal.pone.0126791. PMC 4454694. PMID 26039693.
- ^ Chiappe, Luis M .; Qingjin, Meng; Serrano, Francisco; Sigurdsen, Trond; Min, Wang; Bell, Alyssa; Di, Liu (25 October 2019). "Yeni BohaiornisÇin'in Erken Kretase'sinden bir kuş: enantiyornitin ilişkileri ve uçuş performansı ". PeerJ. 7: e7846. doi:10.7717 / peerj.7846. PMC 6816414. PMID 31667014.
- ^ Wang, Yan; Wang, Min; O'Connor, Jingmai K .; Wang, Xiaoli; Zheng, Xiaoting; Zhang, Xiaomei (11 January 2016). "A new Jehol enantiornithine bird with three-dimensional preservation and ovarian follicles". Omurgalı Paleontoloji Dergisi. 36 (2): e1054496. doi:10.1080/02724634.2015.1054496.
- ^ Kurochkin, E. N.; Chatterjee, S .; Mikhailov, K. E. (19 December 2013). "An embryonic enantiornithine bird and associated eggs from the cretaceous of Mongolia". Paleontological Journal. 47 (11): 1252–1269. doi:10.1134/s0031030113110087.
- ^ Xing, Lida; McKellar, Ryan C.; O’Connor, Jingmai K .; Niu, Kecheng; Mai, Huijuan (29 October 2019). "Burma kehribarında korunmuş orta Kretase enantiornitin ayak ve kuyruk tüyü". Bilimsel Raporlar. 9 (1): 15513. Bibcode:2019NatSR ... 915513X. doi:10.1038 / s41598-019-51929-9. PMC 6820775. PMID 31664115.
Dış bağlantılar
![]() İle ilgili medya Enantiornithes Wikimedia Commons'ta
İle ilgili medya Enantiornithes Wikimedia Commons'ta

















| Anatomi | |||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Davranış | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Evrim | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Fosil kuşlar | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| İnsan etkileşim | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Listeler | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Neornithes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||