Kuş seslendirme - Bird vocalization

Kuş seslendirme ikisini de içerir kuş sesleri ve kuş şarkıları. Teknik olmayan kullanımda, kuş şarkılar insan kulağına melodik olan kuş sesleridir. İçinde ornitoloji ve kuş gözlemciliği şarkılar (nispeten karmaşık seslendirmeler), işlevle çağrılardan (nispeten basit seslendirmeler) ayırt edilir.
Tanım
Şarkılar ve çağrılar arasındaki ayrım karmaşıklığa, uzunluğa ve bağlama dayanır. Şarkılar daha uzun ve daha karmaşıktır ve bölge[3] ve kur yapma ve çiftleşme Çağrılar şu işlevlere hizmet etme eğilimindeyken alarmlar veya bir üye tutmak sürü temas halinde.[4] Howell ve Webb (1995) gibi diğer otoriteler, ayrımı işleve dayalı olarak yaparlar, böylece güvercinler gibi kısa seslendirmeler ve hatta davul çalma gibi vokal olmayan sesler ağaçkakanlar ve "Winnowing " nın-nin Keskinlikler 'gösteri uçuşundaki kanatlar, şarkı olarak kabul edilir.[5] Yine de diğerleri, şarkının hece çeşitliliğine ve zamansal düzenliliğe sahip olmasını gerektirir, benzer şekilde tanımlayan tekrarlayan ve dönüştürücü kalıplara benzer. müzik. Kuş gözlemciliği ve ornitolojide genellikle hangi seslerin şarkı ve hangilerinin çağrı olduğu üzerinde anlaşılır ve iyi bir alan rehberi ikisi arasında ayrım yapacaktır.
Kuş şarkısı en iyi şekilde sipariş Passeriformes. Bazı gruplar neredeyse sessizdir ve yalnızca vurmalı ve ritmik gibi sesler Leylekler, faturalarını dağıtan. Bazı manakinlerde (Pipridae ), erkekler mekanik ses üretimi için çeşitli mekanizmalar geliştirmişlerdir. stridülasyon bazı böceklerde bulunanlardan farklı değil.[6]
Bazı türler uçarken şarkı söylese de, şarkı genellikle belirgin tüneklerden verilir. Seslerin kullanımının aksine mekanik yollarla üretilmesi Syrinx çeşitli olarak adlandırılmıştır enstrümantal müzik tarafından Charles Darwin, mekanik sesler[7] ve daha yakın zamanda seslenme.[8] Dönem sonat gaga, kanatlar, kuyruk, ayaklar ve vücut tüyleri gibi şırıngalı olmayan yapılar kullanılarak üretilen, kasıtlı olarak modüle edilmiş iletişim sinyalleri olan vokal olmayan sesler üretme eylemi olarak tanımlanmıştır.[8]
Tropikal dışı olarak Avrasya ve Amerika neredeyse tüm şarkılar erkek kuşlar tarafından üretilir; ancak tropik bölgelerde ve daha büyük ölçüde çöl kemerleri Avustralya ve Afrika dişilerin erkekler kadar şarkı söylemesi daha tipiktir. Bu farklılıklar uzun zamandır biliniyor[9][10] ve genellikle Avustralya ve Afrika'nın kurak bölgelerinin çok daha az düzenli ve mevsimlik iklimine atfedilirler, çünkü gıda tedariki hiçbir zaman minimum seviyenin üzerine çıkmadığı için uzun yıllar üreyemezlerse de, koşullar uygun olduğunda kuşların üremesini gerektirir.[9] Mevsimsel düzensiz yetiştirme ile, her iki cinsiyetin de üreme durumuna getirilmesi gerekir ve seslendirme, özellikle düet bu amaca hizmet eder. Tropik bölgelerde, Avustralya ve Güney Afrika'da kadın seslendirmelerinin yüksek sıklığı, çok daha güçlü çift bağları ve bölgesellik üreten çok düşük ölüm oranlarıyla da ilgili olabilir.[11]
Anatomi ve psikoloji
Kuş ses organına, Syrinx;[12] alt kısmında kemikli bir yapıdır. trakea (aksine gırtlak tepesinde memeli trakea). Sirenks ve bazen çevreleyen hava kesesi, kuşun havayı zorladığı zarların oluşturduğu ses dalgalarına rezonansa girer. Kuş, zarlardaki gerilimi değiştirerek perdeyi kontrol eder ve ekshalasyon kuvvetini değiştirerek hem perdeyi hem de hacmi kontrol eder. Trakeanın iki tarafını bağımsız olarak kontrol edebilir, bu şekilde bazı türler aynı anda iki nota çıkarabilir.
Fonksiyon

Kuş ötüşünün iki ana işlevinden biri, eş çekiciliğidir.[14] Bilim adamları, kuş şarkısının evrildiğini varsayıyorlar. cinsel seçim ve deneyler, kuş şarkısının kalitesinin uygunluğun iyi bir göstergesi olabileceğini düşündürmektedir.[15] Deneyler ayrıca parazitlerin ve hastalıkların şarkı hızı gibi şarkı özelliklerini doğrudan etkileyebileceğini ve bu nedenle güvenilir sağlık göstergeleri olarak işlev görebileceğini göstermektedir.[16][17] Şarkı repertuarının da bazı türlerde uygunluğa işaret ettiği görülüyor.[18][19] Erkek kuşların tutunma ve reklam verme yeteneği bölgeler şarkı kullanmak aynı zamanda kondisyonlarını da gösterir. Bu nedenle dişi bir kuş, şarkılarının kalitesine ve şarkı repertuarlarının büyüklüğüne göre erkekleri seçebilir.
Kuş ötüşünün ikinci temel işlevi bölge savunmasıdır.[14] Bölgesel kuşlar, bölge sınırlarını aşmak için şarkı kullanarak birbirleriyle etkileşime girecekler. Şarkı, güvenilir bir kalite göstergesi olabileceğinden, bireyler rakiplerinin kalitesini fark edebilir ve enerjik olarak maliyetli bir kavgayı önleyebilir.[15] Şarkı repertuarına sahip kuşlarda, bireyler aynı şarkı tipini paylaşabilir ve bu şarkı türlerini daha karmaşık iletişim için kullanabilir.[20] Bazı kuşlar, bir şarkı tipi eşleşmesi ile (yani aynı şarkı tipiyle) paylaşılan bir şarkı tipine cevap verecektir.[21] Bu agresif bir sinyal olabilir; ancak sonuçlar karışıktır.[20] Kuşlar, repertuar eşleşmeleri kullanarak da etkileşime girebilir; burada bir kuş, rakibinin repertuarında bulunan ancak şu anda söylediği şarkı olmayan bir şarkı türüyle yanıt verir.[22] Bu, şarkı tipi eşleştirmeden daha az agresif bir hareket olabilir.[22] Şarkı karmaşıklığı, erkeklerin bölgesel savunmasıyla da bağlantılıdır ve daha karmaşık şarkılar daha büyük bir bölgesel tehdit olarak algılanır.[23]
Kuş çağrıları yoluyla iletişim, aynı türden bireyler arasında veya hatta türler arasında olabilir. Kuşlar, tehdide özgü seslendirmeler ve hareketler yoluyla alarm verirler ve kuş alarmları, belirli bir tehdidi belirlemek ve ona karşı korumak için diğer kuşlar da dahil olmak üzere diğer hayvan türleri tarafından anlaşılabilir.[24] Mobbing Çağrılar, bir baykuş veya başka bir yırtıcı hayvanın bulunabileceği bir bölgedeki kişileri işe almak için kullanılır. Bu çağrılar, geniş frekans spektrumları, keskin başlangıç ve sonlanma ve türler arasında yaygın olan ve kolayca bulunarak diğer potansiyel "gangsterlere" yardımcı olduğuna inanılan tekrarlama ile karakterize edilir. Öte yandan, çoğu türün alarm çağrıları karakteristik olarak tizdir ve arayanın yerini bulmasını zorlaştırır.[25]
Tek tek kuşlar, çağrılarıyla birbirlerini tanımlayacak kadar hassas olabilir. Kolonilerde yuva yapan pek çok kuş, çağrılarını kullanarak civcivlerini bulabilir.[26] Aramalar, ekolojik çalışmalarda insan araştırmacılar tarafından bile bireysel tanımlama için bazen yeterince farklıdır.[27]
Birçok kuş düet çağrısına katılır. Bazı durumlarda, düetler neredeyse tek bir çağrı gibi görünecek kadar mükemmel zamanlanmıştır. Bu tür bir çağrıya antifonal düet denir.[28] Bu tür düet, bıldırcınlar da dahil olmak üzere çok çeşitli ailelerde belirtilmiştir.[29] çalılar,[30] yedikardeşler benzeri Scimitar babblers ve bazı baykuşlar[31] ve papağanlar.[32] Bölgesel ötücü kuşlarda kuşların karşı bölgelerine saldırı simülasyonu ile uyandırıldıklarında.[33] Bu, tür içi agresif rekabette bir rol anlamına gelir.
Bazen, üreme sonrası sezonda seslendirilen şarkılar, Türdeş kulak misafiri olanlar.[34] İçinde siyah boğazlı mavi ötleğenler Yetiştirilen ve çoğalan erkekler, ses gelişimlerini etkilemek için yavrularına şarkı söylerken, üremekte başarısız olan erkekler genellikle yuvaları terk eder ve sessiz kalır. Üreme sonrası şarkı, bu nedenle, üreme başarısı olasılığı daha yüksek olan belirli habitatların başarısız erkeklerini yanlışlıkla bilgilendirir. sosyal iletişim Seslendirme ile yüksek kaliteli habitatların bulunması için bir kısayol sağlar ve çeşitli bitki örtüsü yapılarını doğrudan değerlendirme zahmetinden kurtarır.

Bazı kuşlar mükemmel ses çıkarır taklit. Bazı tropikal türlerde, drongos oluşumunda rolü olabilir karma tür yiyecek arayan sürüler.[35] Vokal taklit, benzerleri, diğer türleri ve hatta insan yapımı sesleri içerebilir. Ses taklitçiliğinin işlevleri üzerine, uygunluğun bir göstergesi olarak hareket ederek, kuluçka parazitlerine yardım ederek veya avlanmaya karşı koruma sağlayarak cinsel seçime dahil olabilecekleri önerileri dahil olmak üzere birçok hipotez yapılmıştır, ancak herhangi bir işlev için güçlü destek eksiktir.[36] Birçok kuşun, özellikle boşluklarda yuva yapanların, yakın mesafeden yırtıcıları caydırmaya yardımcı olabilecek yılan benzeri bir tıslama sesi çıkardığı bilinmektedir.[37]
Bazı mağaralarda yaşayan türler, yağ kuşu[38] ve swiftlet'ler (Kolokalya ve Aerodramus Türler),[39] işitilebilir ses kullanın (ses konumunun çoğu 2 ile 5 kHz arasında meydana gelir)[40]) için ekolokasyon mağaraların karanlığında. Yararlandığı bilinen tek kuş infrasound (yaklaşık 20 Hz'de) Batı capercaillie.[41]
Kuşların işitme aralığı 50 Hz'nin altındadır (infrasound ) 1 ile 5 kHz arasında maksimum hassasiyetle yaklaşık 12 kHz.[19][42] siyah jacobin yaklaşık 11,8 kHz'de ses üretmede olağanüstüdür. Bu sesleri duyup duymadıkları bilinmemektedir.[43]
Bir çevrede kuşların çağırdığı frekans aralığı, habitatın kalitesine ve ortam seslerine göre değişir. Akustik adaptasyon hipotezi, dar bant genişlikleri, düşük frekanslar, uzun elementler ve elementler arası aralıkların karmaşık bitki örtüsü yapılarına sahip habitatlarda (ki bu sesleri emer ve boğar) bulunurken, yüksek frekanslar, geniş bant genişliği, yüksek frekans modülasyonları ( tril) ve kısa elementler ve ara elementler, engelleyici bitki örtüsü olmayan açık habitatlarda beklenebilir.[44][45][46]
Düşük frekanslı şarkılar, engelli, yoğun bitki örtüsüne sahip habitatlar için idealdir çünkü düşük frekanslı, yavaş modüle edilmiş şarkı unsurları, ses yansıtan bitki örtüsünden yankılanmalar yoluyla sinyal bozulmasına daha az duyarlıdır. Hızlı modülasyonlara sahip yüksek frekanslı çağrılar, açık alanlarda daha az bozundukları için açık habitatlar için idealdir.[47][48] Akustik adaptasyon hipotezi ayrıca şarkı özelliklerinin çevrenin yararlı akustik özelliklerinden yararlanabileceğini belirtir. Dar frekanslı bant genişliği notaları, yoğun bitki örtüsüne sahip habitatlarda yankılanmalarla hacim ve uzunluk olarak artar.[49]
Mevcut frekans aralığının bölündüğü ve kuşlar, farklı türler arasındaki örtüşmenin frekans ve zaman açısından azaltılması için çağrı yaptığı varsayılmıştır. Bu fikir "akustik niş" olarak adlandırıldı.[50] Kuşlar, ortamın düşük frekanslı gürültünün olduğu kentsel alanlarda daha yüksek sesle ve daha yüksek perdede şarkı söyler.[51][52] Trafik gürültüsünün üreme başarısını düşürdüğü bulunmuştur. büyük baştankara (Parus majör) akustik frekanstaki örtüşme nedeniyle.[53] Şarkı hacmindeki artış, daha yüksek frekanslı şarkılarda olduğu gibi, kentsel alanlarda kuşlara uygunluğu geri kazandırdı.[54]
Kuşların şarkı karmaşıklığında enlemesine farklılıklar gösterdiği öne sürülmüştür, ancak şarkı karmaşıklığının enlem veya göç davranışıyla arttığına dair güçlü bir kanıt yoktur.[55]
2019 yılında yayınlanan bir araştırmaya göre, beyaz çan kuşu 125'e ulaşarak kuşlar için şimdiye kadar kaydedilmiş en yüksek sesli aramayı yapardB.[56][57] Kayıt daha önce çığlık atan piha 116 dB ile.[58]
Nöroanatomi

Kuş şarkılarının edinilmesi ve öğrenilmesi, iki bağlantı yolunda hizalanmış bir grup farklı beyin alanını içerir:[59]
- Ön ön beyin yolu (vokal öğrenme ): Memeli bazal gangliyonlarına bir homolog olan Alan X'den oluşur; yan kısmı magnoselüler aynı zamanda kuş bazal ganglionlarının bir parçası olarak kabul edilen anterior nidopallium (LMAN) çekirdeği; ve medial talamusun (DLM) dorso-lateral bölümü.
- Posterior alçalan yol (ses üretimi): HVC'den oluşur (uygun ad, ancak bazen yüksek ses merkezi ); arcopallium'un (RA) sağlam çekirdeği; ve trakeosiringeal kısmı hipoglossal çekirdek (nXIIts).[60][61]
Posterior alçalan yol (PDP), bir kuşun yaşamı boyunca normal şarkı üretimi için gerekliyken, anterior ön beyin yolu (AFP) şarkı öğrenme, esneklik ve bakım için gereklidir, ancak yetişkin şarkı üretimi için gerekli değildir.[62]
Şarkı sistemindeki her iki sinir yolu da HVC, bilgileri hem RA'ya (premotor çekirdek) hem de ön ön beynin Alan X'e yansıtır. Posterior alçalan yolla ilgili bilgiler (ayrıca ses üretimi veya motor yol olarak da adlandırılır), HVC RA'ya ve daha sonra RA'dan trakeosiringeal kısmına hipoglossal sinir (nXIIts), daha sonra sirenksin kas kasılmalarını kontrol eder.[59][63]
Ön ön beyin yolundaki bilgiler, HVC Alan X'e (bazal gangliya), ardından Alan X'ten DLM'ye (talamus) ve DLM'den LMAN'a, daha sonra vokal öğrenme ve RA'ya geri dönen bağlantılar yoluyla vokal prodüksiyon yolları. Bazı araştırmacılar, LMAN ve RA arasındaki bağlantının, şarkı çıkışı için motor programını uyarlamalı olarak değiştiren (kuşun kendi şarkısını ezberlenmiş şarkı şablonuyla karşılaştırarak) işitsel geri bildirim değerlendirmesine dayanan öğretici bir sinyal taşıdığı bir model öne sürdüler.[62][64] Bu eğitici sinyalin üretilmesi, Alan X ve LMAN'daki işitsel nöronlar tarafından kolaylaştırılabilir ve kuşun kendi şarkısının (BOS) zamansal nitelikleri için seçicilik gösterir ve öğretmen şarkısı, BOS ile ezberlenmiş öğretmen şarkısını karşılaştırmak için bir platform sağlar.[64][65]
AFP ile PDP arasındaki gerçek zamanlı hata düzeltme etkileşimleriyle ilgili modeller gelecekte dikkate alınacaktır. Diğer güncel araştırmalar, altında yatan hücresel mekanizmaları keşfetmeye başladı. HVC şarkı yapısının zamansal kalıplarının kontrolü ve hece üretiminin RA kontrolü.[66]Her iki yola da dahil olan beyin yapıları gösteriyor cinsel dimorfizm birçok kuş türünde, genellikle erkeklerin ve dişilerin farklı şekilde ötmesine neden olur. Beyindeki bilinen bazı dimorfizm türleri arasında çekirdek boyutu, mevcut nöron sayısı ve bir çekirdeği diğerine bağlayan nöron sayısı bulunur.[67]
Son derece dimorfik zebra ispinozlarında (Taeniopygia guttata), sadece erkeklerin tipik olarak şarkı söylediği bir tür, HVC ve RA'nın boyutu erkeklerde kadınlara göre yaklaşık üç ila altı kat daha büyüktür ve Alan X, kadınlarda tanınabilir görünmemektedir.[68] Araştırmalar, erken gelişim sırasında seks steroidlerine maruz kalmanın beyindeki bu farklılıklardan kısmen sorumlu olduğunu göstermektedir. Dişi zebra ispinozları yumurtadan çıktıktan sonra estradiol ile tedavi edilmiş, ardından testosteron veya dihidrotestosteron Yetişkinlikte (DHT) tedavisi, boyut olarak erkeklere benzer bir RA ve HVC geliştirecek ve ayrıca erkek benzeri şarkı söyleme davranışı gösterecektir.[69]
Tek başına hormon tedavisi, beyin yapıları veya tam olarak erkekler gibi davranışlara sahip dişi ispinozlar üretmiyor gibi görünüyor. Dahası, diğer araştırmalar, memeli cinsel farklılaşması hakkındaki mevcut bilgilerimize dayanarak beklenilenle çelişen sonuçlar göstermiştir. Örneğin erkek zebra ispinozları, yumurtadan çıkan yavrular hala normal erkeksi şarkı söyleme davranışı geliştirdikçe kısırlaştırılmış veya seks steroid inhibitörleri verilmişlerdir.[67] Bu, z kromozomundaki genlerin aktivasyonu gibi diğer faktörlerin de normal erkek şarkı gelişiminde rol oynayabileceğini düşündürmektedir.[70]
Hormonların ayrıca yetişkin kuşlarda şarkı söyleme ve şarkı çekirdekleri üzerinde aktivasyon etkisi vardır. Kanaryalarda (Serinus canaria), dişiler normalde erkeklerden daha az sıklıkta ve daha az karmaşıklıkla şarkı söyler. Bununla birlikte, yetişkin dişilere androjen enjeksiyonları yapıldığında, şarkı söylemeleri neredeyse erkek benzeri bir frekansa yükselecektir.[71] Ayrıca, androjen enjekte edilen yetişkin dişiler de HVC ve RA bölgelerinde artan bir boyut sergilemektedir.[72] Melatonin Pek çok ötücü kuş, şarkı çekirdeklerinin nöronlarında melatonin reseptörleri gösterdiğinden, yetişkinlerde şarkı davranışını etkilediğine inanılan başka bir hormondur.[73]
İkisi de Avrupa sığırcık (Sturnus vulgaris) ve ev kuşu (Yoldan geçen kişi), karanlığa ve melatonin salgılarına farklı maruz kalmalarla ilişkili şarkı çekirdeklerindeki değişiklikleri göstermiştir.[74][75] Bu, melatoninin, gün ışığı miktarının yıl boyunca önemli ölçüde değiştiği bölgelerde yaşayan ötücü kuşlarda şarkı söyleme davranışının mevsimsel değişikliklerinde rol oynayabileceğini göstermektedir. Diğer bazı çalışmalar, şarkı sistemindeki beyin yapılarının morfolojisindeki mevsimsel değişiklikleri incelemiş ve bu değişikliklerin (yetişkin nörojenezi, gen ifadesi) fotoperiyot, hormonal değişiklikler ve davranış tarafından belirlendiğini bulmuştur.[76][77]
Gen FOXP2 İnsanlarda hem konuşma üretimini hem de dili anlamayı etkileyen kusurları, hem genç zebra ispinozlarında hem de yetişkin kanaryalarda ses esnekliği dönemlerinde X Bölgesi'nde yüksek oranda ifade edilir.[78]
Öğrenme

Farklı kuş türlerinin şarkıları çeşitlidir ve genellikle türlere özgüdür. Türler, şarkılarının karmaşıklığı ve söyledikleri farklı şarkı türlerinin sayısı açısından büyük farklılıklar gösterir (şarkılarda 3000'e kadar). kahverengi atıcı ); bazı türlerdeki bireyler de aynı şekilde farklılık gösterir. Gibi birkaç türde lir kuşları ve alaycı kuşlar şarkılar, bireyin yaşamı boyunca öğrenilen keyfi unsurları, bir tür taklitçiliği (kuş başka bir türe geçmediği için belki daha iyi "sahiplenme" olarak adlandırılsa da (Ehrlich ve diğerleri) içine yerleştirdi). 1773 gibi erken bir tarihte, kuşların çağrıları öğrendikleri ve çapraz besleme deneyler linnet yapımında başarılı oldu Acanthis kenevir bir paraşütçünün şarkısını öğren, Alauda arvensis.[80] Pek çok türde, temel şarkı türün tüm üyeleri için aynı olmasına rağmen, genç kuşların şarkılarının bazı ayrıntılarını babalarından öğrendikleri ve bu varyasyonların nesiller boyunca oluştuğu görülmektedir. lehçeler.[81]
Yavru kuşlarda şarkı öğrenimi iki aşamada gerçekleşir: gencin babayı veya diğer akraba kuşu dinlemesini ve şarkının (şarkı şablonu) spektral ve zamansal niteliklerini ezberlemesini içeren duyusal öğrenme ve yavru kuş üretimini içeren duyusal öğrenme kendi seslendirmelerini yapar ve ezberlenmiş şarkı şablonuyla doğru bir şekilde eşleşene kadar şarkısını pratik eder.[82]
Sensorimotor öğrenme aşamasında şarkı üretimi, benzer şekilde "alt şarkı" adı verilen oldukça değişken alt seslendirmelerle başlar. gevezelik insan bebeklerde. Kısa bir süre sonra, çocuk şarkısı taklit edilen yetişkin şarkısının bazı tanınabilir özelliklerini gösterir, ancak yine de kristalize şarkının klişesinden yoksundur - buna "plastik şarkı" denir.[59]
İki veya üç aylık şarkı öğrenme ve provadan (türe bağlı olarak) sonra, genç, spektral ve zamansal stereotipi (hece üretimi ve hece sıralamasında çok düşük değişkenlik) ile karakterize edilen kristalize bir şarkı üretir.[83] Gibi bazı kuşlar zebra ispinozları Kuş ötüşü araştırmaları için en popüler tür olan, üst üste binen duyusal ve duyusal-motor öğrenme aşamalarına sahiptir.[79]
Araştırmalar, kuşların ötüşü edinmelerinin bir tür motor öğrenme bölgelerini içeren Bazal ganglion. Ayrıca, PDP (bkz. Nöroanatomi aşağıda) kabul edildi homolog bir memeli motor yoluna beyin zarı ve alçalırken beyin sapı AFP'nin bazal gangliya ve talamustan geçen memeli kortikal yoluna homolog olduğu düşünülmektedir.[59] Kuş şarkısı modelleri motor öğrenme insanların nasıl öğrendiğine ilişkin modeller geliştirmede yararlı olabilir konuşma.[84] Zebra ispinozları gibi bazı türlerde şarkı öğrenimi ilk yılla sınırlıdır; bunlara "yaş sınırlı" veya "sınırlı" öğrenciler denir. Kanaryalar gibi diğer türler, cinsel açıdan olgun yetişkinler gibi yeni şarkılar geliştirebilirler; bunlara "açık uçlu" öğrenciler denir.[85][86]
Araştırmacılar, öğrenilen şarkıların kültürel etkileşim yoluyla daha karmaşık şarkıların geliştirilmesine izin verdiğini, böylece kuşların akrabalarını tanımlamalarına ve şarkılarını farklı akustik ortamlara uyarlamalarına yardımcı olan türler arası lehçelere izin verdiğini varsaydılar.[87]
Kuş sesleri öğrenmede işitsel geribildirim
Thorpe'un 1954'teki ilk deneyleri, bir kuşun bir öğretmenin şarkısını duyabilmesinin önemini gösterdi. Kuşlar, müstehcen erkeklerin etkisinden uzakta, izole bir şekilde yetiştirildiklerinde, yine de şarkı söylerler. Yaptıkları şarkı "izole şarkı" olarak adlandırılır, yabani bir kuşun şarkısını anımsatırken, yabani şarkıdan belirgin şekilde farklı özellikler gösterir ve karmaşıklığından yoksundur.[88][89] Sensörimotor döneminde kuşun kendi şarkılarını duyabilmesinin önemi daha sonra Konishi tarafından keşfedildi. Şarkı kristalleşme döneminden önce kulakları sağır olan kuşlar, vahşi tip ve izole şarkılardan belirgin şekilde farklı şarkılar üretmeye devam etti.[90][91] Bu bulguların ortaya çıkmasından bu yana, araştırmacılar duyusal / duyusal-motor öğrenmeyi kolaylaştıran ve kuşun kendi şarkısının ezberlenmiş şarkı şablonuyla eşleşmesine aracılık eden nöral yolları araştırıyorlar.
Son yıllarda yapılan birkaç çalışma, kuşların ötüşü öğrenmesinin altında yatan sinir mekanizmalarına, şarkının üretimi veya sürdürülmesinde yer alan ilgili beyin yapılarına lezyonlar uygulayarak ya da kuşların şarkı kristalizasyonundan önce ve / veya sonra sağırlaşması yoluyla baktı. Bir başka yeni deneysel yaklaşım, kuşun şarkısını kaydetmek ve ardından kuş şarkı söylerken onu tekrar çalmaktı, bu da rahatsız edici işitsel geribildirime neden olmaktı (kuş kendi şarkısının üst üste gelmesini ve önceki bir şarkı hecesinin parçalanmış bir bölümünü duyar).[83] Nordeen ve Nordeen'den sonra[92] Leonardo & Konishi (1999), yetişkin kuşlarda şarkının kristalize edilmiş şarkıyla sürdürülmesi için işitsel geri bildirimin gerekli olduğunu gösterdikçe dönüm noktası niteliğinde bir keşif yaptılar, Leonardo ve Konishi (1999), yetişkin şarkı bakımında işitsel geri bildirimin rolünü daha da keşfetmek için bir işitsel geribildirim pertürbasyon protokolü tasarladı. , yetişkin şarkılarının, rahatsız edici işitsel geribildirime uzun süre maruz kaldıktan sonra nasıl kötüleştiğini araştırmak ve yetişkin kuşların, rahatsız edici geribildirim maruziyetinden çıkarıldıktan sonra zaman içinde kristalize olmuş şarkıyı ne ölçüde kurtarabileceğini incelemek. Bu çalışma, yetişkin şarkı stabilitesini korumada işitsel geri bildirimin rolü için daha fazla destek sundu ve kristalize kuş şarkılarının yetişkin bakımının statik değil dinamik olduğunu gösterdi.
Brainard ve Doupe (2000), LMAN'ın (ön ön beyin), kuş tarafından üretilen şarkı ile ezberlenmiş şarkı şablonu arasındaki farklılıkları tespit ettiği ve ardından eğitici bir hata sinyali gönderdiği için hata düzeltmede birincil rol oynadığı bir model önermektedir. şarkı üretimi için motor programını düzeltmek veya değiştirmek için vokal üretim yolundaki yapılar. Brainard ve Doupe (2000), yaptıkları çalışmada, sağır edici yetişkin kuşların, değişen işitsel geribildirim ve motor programın adaptif olmayan modifikasyonu nedeniyle şarkı stereotipinin kaybına yol açarken, yetişkin kuşların ön ön beyin yolunda lezyona neden olduğunu göstermiştir. sağır olması şarkının stabilizasyonuna yol açmıştır (sağır kuşlarda LMAN lezyonları hece üretiminde ve şarkı yapısında daha fazla bozulmayı önlemiştir).
Şu anda, LMAN'ın eğitici bir hata sinyali üretme ve bunu motor üretim yoluna yansıtmadaki rolünü açıklığa kavuşturan iki rakip model var:
Kuşun kendi şarkısı (BOS) -tuned hata düzeltme modeli
- Şarkı söyleme sırasında, LMAN nöronlarının aktivasyonu, kuş tarafından üretilen şarkıdan gelen işitsel geri bildirim ile depolanan şarkı şablonu arasındaki eşleşmeye bağlı olacaktır. Bu doğruysa, LMAN nöronlarının ateşleme hızları işitsel geribildirimdeki değişikliklere duyarlı olacaktır.
Hata düzeltmenin etkin kopya modeli
- Bir efferans kopyası Şarkı üretimi için motor komutunun gerçek zamanlı hata düzeltme sinyalinin temelini oluşturur. Şarkı söyleme sırasında, LMAN nöronlarının aktivasyonu, şarkıyı oluşturmak için kullanılan motor sinyaline ve bu motor komutuna bağlı olarak beklenen işitsel geri bildirimin öğrenilen tahminine bağlı olacaktır. Bu modelde hata düzeltme daha hızlı gerçekleşir.
Leonardo [93] Bu modelleri, yetişkin zebra ispinozlarının tek LMAN nöronlarında normal ve tedirgin işitsel geribildirime sahip koşullarda şarkı söyleme sırasında artış oranlarını kaydederek doğrudan test etti. LMAN nöronlarının ateşleme hızları işitsel geribildirimdeki değişikliklerden etkilenmediğinden ve bu nedenle LMAN tarafından üretilen hata sinyali işitsel geribildirimle ilgisiz göründüğünden, sonuçları BOS ayarlı hata düzeltme modelini desteklemedi. Dahası, bu çalışmadan elde edilen sonuçlar, LMAN nöronlarının şarkı söyleme sırasında motor sinyalin efference kopyası (ve beklenen işitsel geri bildirim tahminlerinin) tarafından etkinleştirildiği ve nöronların daha kesin zamana sahip olmasını sağlayan efference kopya modelinin tahminlerini destekledi. - işitsel geribildirimdeki değişikliklere kilitlendi.
Ayna nöronları ve ses öğrenimi
Bir ayna nöron bir nöron hem kişi bir eylemi gerçekleştirdiğinde hem de aynı eylemin bir başkası tarafından yapıldığını algıladığında deşarj olur.[94] Bu nöronlar ilk olarak makak maymunlar, ancak son araştırmalar ayna nöron sistemlerinin insanlar dahil diğer hayvanlarda mevcut olabileceğini öne sürüyor.[95]

Ayna nöronlar aşağıdaki özelliklere sahiptir:[94]
- Yer alırlar motor öncesi korteks.
- Hem duyusal hem de motor özellikler sergilerler.
- Eyleme özgüdürler - ayna nöronları, yalnızca bir kişi belirli bir eylem türünü gerçekleştirirken veya gözlemlerken (örneğin, bir nesneyi kavrarken) etkindir.
Çünkü ayna nöronları her ikisini de sergiliyor duyusal ve motor Bazı araştırmacılar, ayna nöronların duyusal deneyimin motor yapılara haritalanmasına hizmet edebileceğini öne sürmüşlerdir.[97] Bunun kuş boyu öğrenme açısından etkileri vardır - birçok kuş şarkılarını almak ve sürdürmek için işitsel geri bildirime güvenir. Ayna nöronlar, kuşun duyduklarının, ezberlenmiş bir şarkı şablonuyla nasıl karşılaştırıldıklarının ve ne ürettiklerinin bu karşılaştırmasına aracılık ediyor olabilir.
Bu işitsel-motor nöronları araştıran Jonathan Prather ve Duke Üniversitesi'ndeki diğer araştırmacılar, tek nöronların aktivitesini HVC'ler nın-nin bataklık serçeleri.[96] HVC'den Alan X'e (HVC) yansıyan nöronlarınX nöronlar) kuş kendi şarkısını çaldığında oldukça duyarlıdır. Bu nöronlar, kuş aynı şarkıyı söylerken de benzer şekillerde ateşlenir. Bataklık serçeleri 3-5 farklı şarkı türü kullanır ve sinirsel aktivite, hangi şarkının duyulduğuna veya söylendiğine bağlı olarak değişir. HVCX nöronlar yalnızca birincil şarkı türü olan şarkılardan birinin sunumuna (veya şarkı söylemesine) yanıt olarak ateşlenir. Ayrıca, şarkı hecesinde belirli bir aşamada ateşlenerek zamansal olarak seçicidirler.
Prather, vd. , kuşun şarkı söylemesinden önceki ve sonraki kısa süre boyunca HVC'sininX nöronlar duyarsız hale gelir işitsel giriş. Başka bir deyişle, kuş kendi şarkısına "sağır" olur. Bu, bu nöronların bir doğal deşarj motor çıktısının ve işitsel girdinin doğrudan karşılaştırılmasına izin verir.[98] Bu, işitsel geri bildirim yoluyla öğrenmenin altında yatan mekanizma olabilir. Bu bulgular aynı zamanda Leonardo'nun (2004) kuş ötüşü öğrenme ve üretimde hata düzeltme efferans kopya modeli ile uyumludur.
Genel olarak, HVCX bataklık serçelerindeki işitsel motor nöronlar, keşfedilen görsel motor ayna nöronlarına çok benzer. primatlar. Ayna nöronları gibi, HVCX nöronlar:
- Bir ön motor beyin bölgesi
- Hem duyusal hem de motor özellikler sergileyin
- Eyleme özgüdür - bir yanıt yalnızca "birincil şarkı türü" tarafından tetiklenir
Ayna nöron sisteminin işlevi hala belirsizdir. Bazı bilim adamları, ayna nöronların diğerlerinin eylemlerini anlamada rol oynayabileceğini düşünüyor. taklit, akıl teorisi ve dil edinimi şu anda yetersiz olmasına rağmen nörofizyolojik bu teorileri destekleyen kanıtlar.[97] Özellikle kuşlarla ilgili olarak, ayna nöron sisteminin temelde yatan genel bir mekanizma olarak hizmet etmesi mümkündür. vokal öğrenme, ancak daha fazla araştırmaya ihtiyaç var. Şarkı öğrenmeye yönelik etkilere ek olarak, ayna nöron sistemi de bir rol oynayabilir. bölgesel davranışlar şarkı türü eşleştirme ve sayma gibi.[99][100]
Kültürel aktarım yoluyla öğrenme
Hayvanlarda kültür genellikle belirli popülasyonların karakteristiği olan sosyal olarak aktarılan davranış kalıplarından ("gelenekler") oluşacak şekilde tanımlanır.[101] Kuş şarkılarının öğrenilmiş doğası ve "diyalekt" benzeri yerel varyasyonların kanıtı, bazı araştırmacıları kuşların varlığını varsaymaya yöneltti. kuş kültürü.
Söylendiği gibi yukarıda, kuş şarkısının öğrenmeye olan bağımlılığı Thorpe tarafından incelendi. kavurucu Hayatlarının ilk haftasından ayrı olarak yetiştirilenler, diğer samanlara kıyasla oldukça anormal ve daha az karmaşık şarkılar üretir.[102] Bu, ötücü kuşlarda şarkı gelişiminin birçok yönünün aynı türün daha yaşlı üyelerinin ders vermesine bağlı olduğunu gösterdi. Daha sonraki çalışmalar, bir samanlığın şarkısında kanarya benzeri unsurlar gözlemledi. kanaryalar,[103] genç kuşlar tarafından şarkı öğrenmede öğretmenlerin güçlü rolünü kanıtlıyor.
Benzer chaffinch şarkı türlerinin (farklı unsurlarına ve sıralarına göre kategorize edilmiş) benzer coğrafi bölgelerde kümelendiği gözlemlendi,[104] ve bu keşif, kuş cıvıltılarında "lehçeler" hakkında hipotezlere yol açtı. O zamandan beri, bu şarkı türü varyasyonlarının lehçeler insan dilinde bulduklarımız gibi. Bunun nedeni, belirli bir coğrafi bölgenin tüm üyelerinin aynı şarkı tipine uymaması ve aynı zamanda onu diğer tüm türlerden ayıran bir şarkı türünün tekil bir özelliğinin olmamasıdır (belirli kelimelerin belirli lehçelere özgü olduğu insan lehçelerinin aksine) ).[101]
Bu öğrenme kanıtına ve yerelleştirilmiş şarkı türlerine dayanarak, araştırmacılar, bir tür kültürel aktarım olarak kuş sesinin sosyal öğrenimini araştırmaya başladı. Bu kültürü oluşturan davranış kalıpları şarkıların kendisidir ve şarkı türleri gelenek olarak kabul edilebilir.
Dopamin devreleri ve kültürel aktarım
Yakın zamanda yapılan bir araştırma, zebra ispinozlarındaki bir dopamin devresinin, öğretmenlerden kuş şarkılarının sosyal olarak öğrenilmesini teşvik edebileceğini göstermiştir.[105] Elde edilen veriler, genç zebra ispinozlarındaki bazı beyin bölgelerinin, zebra ispinoz şarkısı çalan hoparlörler tarafından değil, kendine özgü (yani aynı tür) öğretmenlerin şarkılarıyla heyecanlandığını gösteriyor. Ek olarak, HVC'ye salınan dopaminin şarkının kodlanmasına yardımcı olduğunu gösteriyorlar.
Kuş sesiyle öğrenmenin evrimsel olarak korunması
Kültürel tuzak hipotezi
20. yüzyılda kuş ötüşü üzerine önemli miktarda araştırma yapılmasına rağmen, hiçbiri kuş şarkılarının arkasındaki evrimsel "kullanımı", özellikle geniş vokal repertuarlarıyla açıklayamadı. Buna yanıt olarak Lachlan ve Slater, geniş çeşitlilikteki şarkıların kalıcılığını açıklamak için bir "kültürel tuzak" modeli önerdiler.[106] Bu model, bir "filtreler" kavramına dayanmaktadır, burada:
- bir erkek ötücü kuşun (yani şarkıcının) filtresi geliştirebileceği şarkıların aralığını içerir
- bir dişi ötücü kuşun (yani alıcının) filtresi, eş seçimi için kabul edilebilir bulduğu şarkı aralığını içerir
Olası bir durumda, popülasyon esas olarak geniş filtreli kuşlardan oluşur. Bu popülasyonda, geniş filtreye sahip bir erkek ötücü kuş, dar filtreli birkaç dişi tarafından nadiren seçilecektir (çünkü erkek şarkısının daha dar bir filtreye düşmesi olası değildir). Bu dişinin artık çiftleşmek için nispeten küçük bir erkek seçimi olacak ve bu nedenle dişinin dar filtresinin genetik temeli kalıcı olmayacak. Bir başka olası durum, çoğunlukla dar filtrelere sahip bir nüfusla ilgilidir. İkinci popülasyonda, geniş filtreli erkekler, daha yaşlı, dar filtreli erkeklerden öğrenerek eş seçiminin reddedilmesinden uygulanabilir bir şekilde kaçınabilirler. Bu nedenle, geniş filtreli kuşların ortalama üreme başarısı, öğrenme olasılığı ile artırılır ve ses öğrenme ve geniş şarkı repertuarları (yani geniş filtreler) el ele gider.[106][101]
Kültürel tuzak hipotezi, seçici baskıların genotipler ve bunların kültürel sonuçları arasındaki etkileşimden ortaya çıktığı gen-kültür birlikte evrimine bir örnektir.[106]
Possible correlation with cognitive ability
Various studies have shown that adult birds that underwent stress during critical developmental periods produce less complex songs and have smaller HVC brain regions.[107][108] These has led some researchers to hypothesize that sexual selection for more complex songs indirectly selects for stronger cognitive ability in males.[109] Further investigation showed that male şarkı serçeleri with larger vocal repertoires required less time to solve detour-reaching cognitive tasks.[110] Some have proposed that bird song (among other sexually selected traits such as flashy coloring, body symmetry, and elaborate courtship) allow female songbirds to quickly assess the cognitive skills and development of multiple males.
Identification and systematics

The specificity of bird calls has been used extensively for species identification. The calls of birds have been described using words or nonsense syllables or line diagrams.[111] Common terms in English include words such as şarlatan, cıvıldamak ve cıvıltı. These are subject to imagination and vary greatly; iyi bilinen bir örnek, beyaz boğazlı serçe 's song, given in Kanada gibi O sweet Canada Canada Canada ve Yeni ingiltere gibi Old Sam Peabody Peabody Peabody (Ayrıca Where are you Frederick Frederick Frederick?). In addition to nonsense words, grammatically correct phrases have been constructed as likenesses of the vocalizations of birds. Örneğin, çizgili baykuş produces a motif which some bird guides describe as Who cooks for you? Who cooks for you all? with the emphasis placed on sen.[112] The term "warblish" has been coined to explain this approach to bird call description.[113]

Kay Electric Company started by former Bell Labs engineers Harry Foster and Elmo Crump made a device that was marketed as the "Sona-Graph" in 1948. This was adopted by early researchers[114] including C.E.G.Bailey who demonstrated its use for studying bird song in 1950.[115] Kullanımı spectrograms to visualize bird song was then adopted by Donald J. Borror[116] and developed further by others including W. H. Thorpe.[117][118] These visual representations are also called sonograms or sonagrams. Beginning in 1983, some field guides for birds use sonograms to document the calls and songs of birds.[119] The sonogram is objective, unlike descriptive phrases, but proper interpretation requires experience. Sonograms can also be roughly converted back into sound.[120][121]
Bird song is an integral part of bird courtship and is a pre-zygotic isolation mechanism involved in the process of türleşme. Birçok alopatrik sub-species show differences in calls. These differences are sometimes minute, often detectable only in the sonograms. Song differences in addition to other taxonomic attributes have been used in the identification of new species.[122] The use of calls has led to proposals for splitting of species complexes such as those of the Mirafra bushlarks.[123]
Smartphone apps such as Bird Song Id USA[124] enables automatic recognition of birds using sounds.[125] Sevmek Shazam, those apps work by comparing spektrografik database for matches.
Kuş dili
kuşların dili has long been a topic for anecdote and speculation. That calls have meanings that are interpreted by their listeners has been well demonstrated. Yurtiçi tavuklar have distinctive alarm calls for aerial and ground predators, and they respond to these alarm calls appropriately.[126][127]
Ancak, bir dil has, in addition to words, dilbilgisi (that is, structures and rules). Studies to demonstrate the existence of language have been difficult due to the range of possible interpretations. For instance, some have argued that in order for a communication system to count as a language it must be "combinatorial",[128] having an open ended set of grammar-compliant sentences made from a finite vocabulary.
Birşey üzerine araştırma yapmak papağanlar tarafından Irene Pepperberg is claimed to demonstrate the innate ability for grammatical structures, including the existence of concepts such as nouns, adjectives and verbs.[129] Vahşi doğada Siyah şapkalı Chickadees innate vocalizations have been rigorously shown[128] to have combinatorial language. Üzerine çalışmalar sığırcık vocalizations have also suggested that they may have recursive structures.[130]
Dönem kuş dili may also more informally refer to patterns in bird vocalizations that communicate information to other birds or other animals in general.[131]
Some birds have two distinct "languages", one for internal communications and one for use in flocks. All birds have a separate type of communication for "songs" vs. communicating danger and other information. Konrad Lorenz demonstrated that Jackdaws have "names" identifying each individual in the flock and when beginning flight preparations each of them say one other bird's name creating a "chain". In his book King Solomon's Ring, he describes the name he was given by the birds and how he was recognized several years later in a far away location following WWII.
Studies in parakeets have shown a striking similarity between konuşan kuş 's verbal areas in the brain and the equivalent human brain areas, suggesting that mimicry has much to do with the construction of language and its structures and order.[132] Recent research showed birds construct sentence like communications with a syntax and grammar.[133]
Kültürde
Kayıt
Bu bölüm genişlemeye ihtiyacı var. Yardımcı olabilirsiniz ona eklemek. (Eylül 2011) |
The first known recording of birdsong was made in 1889 by Ludwig Koch,[134] who went on to become an eminent wildlife recordist and BBC natural history presenter.[134]
Other notable birdsong recordists include Eric Simms, Chris Watson and, in France, Jean-Claude Roché, François Charron, Fernand Deroussen.
Müziğin içinde
Müziğin içinde, Birdsong has influenced composers and musicians in several ways: they can be inspired by birdsong; they can intentionally imitate bird song in a composition, as Vivaldi ve Beethoven did, along with many later composers; they can incorporate recordings of birds into their works, as Ottorino Respighi first did; ya da beğen Beatrice Harrison ve David Rothenberg, they can duet with birds.[135][136][137][138] Authors including Rothenberg have claimed that birds sing on traditional scales as used in human music,[139][140][141] but at least one songbird does not choose notes in this way.[142]
Among birds which habitually borrow phrases or sounds from other species, the way they use variations of ritim, relationships of müzikal perde ve kombinasyonları notlar müziğe benzeyebilir.[143] Hollis Taylor's in-depth analysis of alaca kasap kuşu vocalizations provides a detailed rebuttal to objections of birdsong being judged as music.[144] The similar motor constraints on human and avian song may have driven these to have similar song structures, including "arch-shaped and descending melodic contours in musical phrases", long notes at the ends of phrases, and typically small differences in pitch between adjacent notes, at least in birds with a strong song structure like the Eurasian treecreeper Certhia familiaris.[145]
Şiirde
Bird song is a popular subject in şiir. Famous examples inspired by bird song include the 1177 Persian poem "Kuşların Konferansı ", in which the birds of the world assemble under the wisest bird, the ibibik, to decide who is to be their king.[146] In English poetry, John Keats 's 1819 "Bülbüle Ode " ve Percy Bysshe Shelley 's 1820 "Bir Skylark'a " are popular classics.[147][148] Ted Hughes 's 1970 collection of poems about a bird character, "Karga ", is considered one of his most important works.[149] Bird poems by Gerard Manley Hopkins include "Sea and Skylark" and "Windhover ".[150]
Ayrıca bakınız
Referanslar
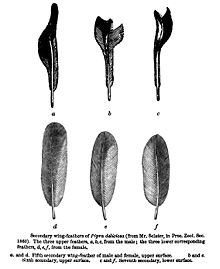
- ^ Sclater, P. L. (1860). "List of Birds collected by Mr. Fraser in Ecuador, at Nanegal, Calacali, Perucho, and Puellaro, with notes and descriptions of new species". Proc. Zool. Soc. Londra: 83–97.
- ^ Darwin, Charles (1871). The descent of man and selection in relation to sex. hacim 2. Londra: John Murray. s. 65–66. ISBN 978-1-108-00510-4.
- ^ Boswall, Jeffery. "Why do birds sing?". İngiliz Kütüphanesi. Arşivlenen orijinal 29 Ocak 2020.
- ^ Ehrlich, Paul R .; David S. Dobkin & Darryl Wheye. ""Bird Voices" and "Vocal Development" from Birds of Stanford essays". Alındı 9 Eylül 2008.
- ^ Howell, Steve N. G. & Sophie Webb (1995). Meksika ve Kuzey Orta Amerika Kuşları Rehberi. Oxford University Press. ISBN 978-0-19-854012-0.
- ^ Bostwick, Kimberly S. & Richard O. Prum (2005). "Courting Bird Sings with Stridulating Wing Feathers". Bilim. 309 (5735): 736. doi:10.1126/science.1111701. PMID 16051789. S2CID 22278735.
- ^ Manson-Barr, P. and Pye, J. D. (1985). Mechanical sounds. In A Dictionary of Birds (ed. B. Campbell and E. Lack), pp. 342-344. Staffordshire: Poyser.
- ^ a b Bostwick, Kimberly S. & Richard O. Prum (2003). "High-speed video analysis of wing-snapping in two manakin clades (Pipridae: Aves)". Deneysel Biyoloji Dergisi. 206 (Pt 20): 3693–3706. doi:10.1242/jeb.00598. PMID 12966061. S2CID 8614009.
- ^ a b Robinson, Angus; "The Biological Significance of Bird Song in Australia" içinde Emu; 48(4), pp. 291-315
- ^ Hartshorne, Charles; "Some Biological Principles Applicable to Song Behavior" in Wilson Bülteni Cilt 70, No. 1 (Mar., 1958), pp. 41-56
- ^ Slater Peter J. B., Mann Nigel I. (2004). "Why do the females of many bird species sing in the tropics?". Kuş Biyolojisi Dergisi. 35 (4): 289–294. doi:10.1111/j.0908-8857.2004.03392.x.
- ^ Attenborough, D. 1998. Kuşların Hayatı. BBC publication.0563-38792-0
- ^ Avustralya Müzesi Çevrimiçi. "Crows and Ravens". Arşivlendi from the original on 1 September 2007. Alındı 12 Ağustos 2007.
- ^ a b Catchpole, C.; Slater, P.J.B (2008). Bird Song: Biological Themes and Variation. Cambridge University Press.
- ^ a b Read, A. W. & D. M. Weary (1990). "Sexual selection and the evolution of bird song: A test of the Hamilton-Zuk hypothesis". Davranışsal Ekoloji ve Sosyobiyoloji. 26 (1): 47–56. doi:10.1007/BF00174024. S2CID 25177326.
- ^ Garamszegi, L. Z .; A. P. Møller; János Török; Gábor Michl; Péter Péczely; Murielle Richard (2004). "Immune challenge mediates vocal communication in a passerine bird: an experiment" (PDF). Davranışsal Ekoloji. 15 (1): 148–157. doi:10.1093/beheco/arg108.
- ^ Redpath, S. M.; Bridget M Appleby; Steve J Petty (2000). "Do male hoots betray parasite loads in Tawny Owls?". Kuş Biyolojisi Dergisi. 31 (4): 457–462. doi:10.1034/j.1600-048X.2000.310404.x.
- ^ Reid, J. M.; Peter Arcese; Alice L. E. V. Cassidy; Sara M. Hiebert; James N. M. Smith; Philip K. Stoddard; Amy B. Marr & Lukas F. Keller (2005). "Fitness Correlates of Song Repertoire Size in Free-Living Song Sparrows (Melospiza melodisi)" (PDF). Amerikan Doğa Uzmanı. 165 (3): 299–310. doi:10.1086/428299. PMID 15729661. S2CID 12547933.
- ^ a b Møller AP; J. Erritzøe; L. Z. Garamszegi (2005). "Covariation between brain size and immunity in birds: implications for brain size evolution" (PDF). Evrimsel Biyoloji Dergisi. 18 (1): 223–237. CiteSeerX 10.1.1.585.3938. doi:10.1111/j.1420-9101.2004.00805.x. PMID 15669979. S2CID 21763448.
- ^ a b Searcy, W. A.; Beecher, M.D. (2009). "Song as an aggressive signal in songbirds". Hayvan Davranışı. 78 (6): 1281–1292. doi:10.1016/j.anbehav.2009.08.011. S2CID 30360474.
- ^ Falls, J. B.; Krebs, J. R.; McGregor, P.K. (1982). "Song matching in the great tit (Parus major) the effect of similarity and familiarity". Hayvan Davranışı. 30 (4): 997–1009. doi:10.1016/S0003-3472(82)80188-7. S2CID 53189625.
- ^ a b Beecher, M.D .; Stoddard, P.K.; Cambell, E.S.; Horning, C.L. (1996). "Repertoire matching between neighbouring song sparrows". Hayvan Davranışı. 51 (4): 917–923. doi:10.1006/anbe.1996.0095. S2CID 26372750.
- ^ Hill, S. D.; Brunton, D. H.; Anderson, M. A .; Weihong, J. (2018). "Fighting talk: complex song elicits more aggressive responses in a vocally complex songbird". İbis. 160 (2): 257–268. doi:10.1111/ibi.12542.
- ^ "A Bird's World: Speaking in a Bird's Language". Museum of Science, Boston. 2008.
- ^ Marler, P. (1955). "Characteristics of some animal calls". Doğa. 176 (4470): 6–8. Bibcode:1955Natur.176....6M. doi:10.1038/176006a0. S2CID 4199385.
- ^ Lengagne, T.; J. Lauga & T. Aubin (2001). "Intra-syllabic acoustic signatures used by the King Penguin in parent-chick recognition: an experimental approach" (PDF). Deneysel Biyoloji Dergisi. 204 (Pt 4): 663–672. PMID 11171348.
- ^ Wayne Delport; Alan C Kemp; J. Willem H Ferguson (2002). "Vocal identification of individual African Wood Owls Strix woodfordii: a technique to monitor long-term adult turnover and residency". İbis. 144 (1): 30–39. doi:10.1046/j.0019-1019.2001.00019.x.
- ^ Thorpe, W. H. (23 February 1963). "Antiphonal Singing in Birds as Evidence for Avian Auditory Reaction Time". Doğa. 197 (4869): 774–776. Bibcode:1963Natur.197..774T. doi:10.1038/197774a0. S2CID 30542781.
- ^ Stokes, A. W.; H. W. Williams (1968). "Antiphonal calling in quail" (PDF). Auk. 85 (1): 83–89. doi:10.2307/4083626. JSTOR 4083626.
- ^ Harris, Tony; Franklin, Kim (2000). Shrikes ve Bush-Shrikes. Princeton University Press. s. 257–260. ISBN 978-0-691-07036-0.
- ^ Osmaston, B. B. (1941). ""Duetting" in birds". İbis. 5 (2): 310–311. doi:10.1111/j.1474-919X.1941.tb00620.x.
- ^ Power, D. M. (1966). "Antiphonal duetting and evidence for auditory reaction time in the Orange-chinned Parakeet". Auk. 83 (2): 314–319. doi:10.2307/4083033. JSTOR 4083033.
- ^ Hyman, Jeremy (2003). "Countersinging as a signal of aggression in a territorial songbird" (PDF). Hayvan Davranışı. 65 (6): 1179–1185. doi:10.1006/anbe.2003.2175. S2CID 38239656.
- ^ Betts, M.G .; Hadley, A.S .; Rodenhouse, N .; Nocera, J.J. (2008). "Göçmen Ötücü Kuşun Yetiştirme Yeri Seçiminde Toplumsal Bilgi Bitki Yapısının Önünde". Bildiriler: Biyolojik Bilimler. 1648. 275 (1648): 2257–2263. doi:10.1098 / rspb.2008.0217. PMC 2603235. PMID 18559326.
- ^ Goodale, E. & Kotagama, S. W. (2005). "Testing the roles of species in mixed-species bird flocks of a Sri Lankan rain forest". Tropikal Ekoloji Dergisi. 21 (6): 669–676. doi:10.1017/S0266467405002609.
- ^ Kelley, L. A .; Coe, R. L.; Madden, J. R.; Healy, S. D. (2008). "Vocal mimicry in songbirds". Hayvan Davranışı. 76 (3): 521–528. doi:10.1016/j.anbehav.2008.04.012. S2CID 53192695.
- ^ Marler, Peter; Hans Willem Slabbekoorn (2004). Nature's music: the science of birdsong. Akademik Basın. s. 145. ISBN 978-0-12-473070-0.
- ^ Suthers RA & Hector DH (1985). "The physiology of vocalization by the echolocating Oilbird, Steatornis caripensis". J. Comp. Physiol. 156 (2): 243–266. doi:10.1007/BF00610867. S2CID 1279919.
- ^ Suthers RA & Hector DH (1982). "Mechanism for the production of echolocating clicks by the Grey Swiftlet, Collocalia spodiopygia". J. Comp. Physiol. Bir. 148 (4): 457–470. doi:10.1007/BF00619784. S2CID 39111110.
- ^ Coles RB; Konishi M & Pettigrew JD (1987). "Hearing and echolocation in the Australian Grey Swiftlet, Collocalia spodiopygia". J. Exp. Biol. 129: 365–371.
- ^ Lieser M; P. Berthold1 & G. A. Manley (2005). "Infrasound in the capercaillie ( Tetrao urogallus )". Ornitoloji Dergisi. 146 (4): 395–398. doi:10.1007/s10336-005-0003-y. S2CID 22412727.
- ^ Dooling, R.J. (1982). Auditory perception in birds. Acoustic Communication in Birds, Vol. 1 (eds D.E. Kroodsma & E.H. Miller). s. 95–130.
- ^ Olson, Christopher R.; Fernández-Vargas, Marcela; Portfors, Christine V.; Mello, Claudio V. (2018). "Black Jacobin hummingbirds vocalize above the known hearing range of birds". Güncel Biyoloji. 28 (5): R204–R205. doi:10.1016/j.cub.2018.01.041. PMID 29510104. S2CID 3727714.
- ^ Derryberry, Elizabeth (July 2009). "Ecology Shapes Birdsong Evolution: Variation in Morphology and Habitat Explains Variation in White-Crowned Sparrow Song". Amerikan Doğa Uzmanı. 174 (1): 24–33. doi:10.1086/599298. PMID 19441960. S2CID 8606774.
- ^ Boncoraglio, G. & Nicola Saino (2007). "Habitat structure and the evolution of bird song: a meta-analysis of the evidence for the acoustic adaptation hypothesis". Fonksiyonel Ekoloji. 21: 134–142. doi:10.1111/j.1365-2435.2006.01207.x.
- ^ Morton, E.S. (1975). "Ecological sources of selection on avian sounds". Amerikan doğa bilimci. 109 (965): 17–34. doi:10.1086/282971. S2CID 55261842.
- ^ Ey, Elodie; Fischer, J. (13 April 2012). "The "acoustic adaptation hypothesis" - a review of the evidence from birds, anurans and mammals". Biyoakustik. 19 (1–2): 21–48. doi:10.1080/09524622.2009.9753613. S2CID 84971439.
- ^ Tubaro, Pablo L .; Segura, Enrique T. (November 1994). "Dialect Differences in the Song of Zonotrichia capensis in the Southern Pampas: A Test of the Acoustic Adaptation Hypothesis". Akbaba. 96 (4): 1084–1088. doi:10.2307/1369117. JSTOR 1369117.
- ^ Slabbekoorn, Hans; Ellers, Jacintha; Smith, Thomas B. (2002). "Birdsong and sound transmission: the benefits of reverberations" (PDF). Akbaba. 104 (3): 564–573. doi:10.1650/0010-5422(2002)104[0564:basttb]2.0.co;2.
- ^ Krause, Bernard L. (1993). "The Niche Hypothesis" (PDF). Soundscape Bülteni. 06. Arşivlenen orijinal (PDF) 2008-03-07 tarihinde.
- ^ Henrik Brumm (2004). "The impact of environmental noise on song amplitude in a territorial bird". Hayvan Ekolojisi Dergisi. 73 (3): 434–440. doi:10.1111/j.0021-8790.2004.00814.x. S2CID 73714706.
- ^ Slabbekoorn, H. & Peet, M. (2003). "Birds sing at a higher pitch in urban noise". Doğa. 424 (6946): 267. Bibcode:2003Natur.424..267S. doi:10.1038/424267a. PMID 12867967. S2CID 4348883.
- ^ Halfwerk, Wouter; Holleman, L.J.M.; Lessells, C.M.; Slabbekoorn, H. (February 2011). "Negative impact of traffic nosie on avian reproductive success". Uygulamalı Ekoloji Dergisi. 48 (1): 210–219. doi:10.1111/j.1365-2664.2010.01914.x. S2CID 83619284.
- ^ Luther, David A.; Derryberry, E.P. (Nisan 2012). "Birdsongs keep pace with city life: changes in song over time in an urban songbird affects communication". Hayvan Davranışı. 83 (4): 1059–1066. doi:10.1016/j.anbehav.2012.01.034. S2CID 31212627.
- ^ Najar, N.; Benedict, L. (2019). "The relationship between latitude, migration and the evolution of bird song complexity". İbis. 161 (1): 1–12. doi:10.1111 / ibi.12648.
- ^ Jeffrey Podos; Mario Cohn-Haft (21 October 2019). "Extremely loud mating songs at close range in white bellbirds". Güncel Biyoloji. 29 (20): R1068–R1069. doi:10.1016/j.cub.2019.09.028. PMID 31639347. S2CID 204823663.
- ^ "World's 'loudest bird': Meet the white bellbird". Newsbeat (video). BBC haberleri. 22 Ekim 2019.
- ^ NEMETH, ERWIN (2004-01-01). "Measuring the Sound Pressure Level of the Song of the Screaming Piha Lipaugus Vociferans: One of the Loudest Birds in the World?". Biyoakustik. 14 (3): 225–228. doi:10.1080/09524622.2004.9753527. ISSN 0952-4622. S2CID 84218370.
- ^ a b c d e Nottebohm, F. (2005). "Birdsong'un Sinir Temeli". PLOS Biol. 3 (5): 163. doi:10.1371 / journal.pbio.0030164. PMC 1110917. PMID 15884976.
- ^ Brainard, M. S. & Doupe, A. J. (2000). "Auditory feedback in learning and maintenance of vocal behavior". Doğa Yorumları Nörobilim. 1 (1): 31–40. doi:10.1038/35036205. PMID 11252766. S2CID 5133196.
- ^ Carew, Thomas J. (2000). Behavioral Neurobiology: The Cellular Organization of Natural Behavior. Sinauer Associates. ISBN 978-0-87893-092-0.
- ^ a b Kao, M.H.; Doupe, A.J.; Brainard, M.S. (2005). "Contributions of an avian basal ganglia-forebrain circuit to real=time modulation of song". Doğa. 433 (7026): 638–642. Bibcode:2005Natur.433..638K. doi:10.1038/nature03127. PMID 15703748. S2CID 4352436.
- ^ Suthers, R. (2004). "How birds sing and why it matters". In Marler, P.; Slabbekoorn, H. (eds.). Nature's music:The science of birdsong. Akademik Basın. pp. 272–295. ISBN 978-0-12-473070-0.
- ^ a b Brainard, M. S. & Doupe, A. J. (2000). "Interruption of a basal ganglia-forebrain circuit prevents plasticity of learned vocalizations". Doğa. 404 (6779): 762–766. Bibcode:2000Natur.404..762B. doi:10.1038/35008083. PMID 10783889. S2CID 4413588.
- ^ Kojima, S .; Doupe, A. (2008). "Neural encoding of auditory temporal context in a songbird basal ganglia nucleus, and its independence of birds' song experience". Avrupa Nörobilim Dergisi. 27 (5): 1231–1244. doi:10.1111/j.1460-9568.2008.06083.x. PMC 2408885. PMID 18364039.
- ^ Long, M.A.; Jin, D.Z.; Fee, M.S. (2010). "Support for a synaptic chain model of neuronal sequence generation". Doğa. 468 (7322): 394–399. Bibcode:2010Natur.468..394L. doi:10.1038/nature09514. PMC 2998755. PMID 20972420.
- ^ a b Balthazart, J.; Adkins-Regan (2002). Sexual differentiation of brain and behavior in birds. Hormonlar, Beyin ve Davranış. 4. pp. 223–301. doi:10.1016/b978-012532104-4/50068-8. ISBN 9780125321044. PMID 18406680.
- ^ Nottebohm, F. & Arnold, A.P. (1976). "Ötücü kuş beyninin ses kontrol bölgelerinde cinsel dimorfizm". Bilim. 194 (4261): 211–213. Bibcode:1976Sci ... 194..211N. doi:10.1126 / science.959852. PMID 959852.
- ^ Gurney, M.E. & Konishi, M. (1980). "Zebra ispinozlarında beyin ve davranışta hormon kaynaklı cinsel farklılaşma". Bilim. 208 (4450): 1380–1383. Bibcode:1980Sci...208.1380G. doi:10.1126 / science.208.4450.1380. PMID 17775725. S2CID 11669349.
- ^ Tomaszycki, M.L.; Peabody, C.; Replogle, K.; Clayton, D.F; Tempelman, R.J.; Wade, J. (2009). "Sexual differentiation of the zebra finch song system: potential roles for sex chromosome genes". BMC Neuroscience. 10: 24. doi:10.1186/1471-2202-10-24. PMC 2664819. PMID 19309515.
- ^ Leonard, S. L. (1 May 1939). "Induction of Singing in Female Canaries by Injections of Male Hormone". Deneysel Biyoloji ve Tıp. 41 (1): 229–230. doi:10.3181/00379727-41-10631. S2CID 87078020.
- ^ Nottebohm, F. (1980). "Testosterone triggers growth of brain vocal control nuclei in adult female canaries". Beyin Araştırması. 189 (2): 429–36. doi:10.1016/0006-8993(80)90102-X. PMID 7370785. S2CID 25845332.
- ^ Ball, G.F. & Balthazart, J. (2002). Neuroendocrine mechanisms regulating reproductive cycles and reproductive behavior in birds. Hormones, Brain, and Behavior. 2. pp. 649–798. doi:10.1016/b978-012532104-4/50034-2. ISBN 9780125321044.
- ^ Bentley, G.E.; Van’t Hof, T.J.; Ball, G.F. (1999). "Seasonal neuroplasticity in the songbird telencephalon: A role for melatonin". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 96 (8): 4674–4679. Bibcode:1999PNAS...96.4674B. doi:10.1073/pnas.96.8.4674. PMC 16391. PMID 10200321.
- ^ Cassone, V.M.; Bartell, P.A.; Earnest D.J. & Kumar, V. (2008). "Duration of melatonin regulates seasonal changes in song control nuclei of the house sparrow, Passer domesticus: Independence from gonads and circadian entrainment". Biyolojik Ritimler Dergisi. 23 (1): 49–58. doi:10.1177/0748730407311110. PMID 18258757. S2CID 206544790.
- ^ Ball, G.F.; Auger, C.J.; Bernard, D.J.; Charlier, T.D.; Sartor, J.J.; Riters, L.V.; Balthazart, J. (2004). "Seasonal plasticity in the song control system: Multiple brain sites of steroid hormone action and the importance of variation in song behavior". New York Bilimler Akademisi Yıllıkları. 1016 (1): 586–610. Bibcode:2004NYASA1016..586B. doi:10.1196/annals.1298.043. PMID 15313796. S2CID 42818488.
- ^ London, S.E.; Replogle, K.; Clayton, D.F. (2009). "Developmental shifts in gene expression in the auditory forebrain during the sensitive period for song learning". Developmental Neurobiology. 69 (7): 436–450. doi:10.1002/dneu.20719. PMC 2765821. PMID 19360720.
- ^ Scharff C; Haesler S (2005). "FoxP2'ye evrimsel bir bakış açısı: kesinlikle kuşlar için mi?". Nörobiyolojide Güncel Görüş. 15 (6): 694–703. doi:10.1016 / j.conb.2005.10.004. PMID 16266802. S2CID 11350165.
- ^ a b Brainard, M. S. & Doupe, A. J. (2002). "Ötücü kuşların bize öğrenmek hakkında ne öğrettiği". Doğa. 417 (6886): 351–358. Bibcode:2002Natur.417..351B. doi:10.1038 / 417351a. PMID 12015616. S2CID 4329603.
- ^ Barrington, D. (1773). "Experiments and observations on the singing of birds". Kraliyet Cemiyetinin Felsefi İşlemleri. 63: 249–291. doi:10.1098/rstl.1773.0031. S2CID 186207885.
- ^ Marler, P.; M. Tamura (1962). "Song dialects in three populations of the white-crowned sparrow". Condor. 64 (5): 368–377. doi:10.2307/1365545. JSTOR 1365545.
- ^ Konishi, M. (2010). "From central pattern generator to sensory template in the evolution of birdsong". Brain & Language. 115 (1): 18–20. doi:10.1016/j.bandl.2010.05.001. PMID 20955898. S2CID 205791930.
- ^ a b Leonardo, A.; Konishi, M. (1999). "Decrystallization of adult birdsong by perturbation of auditory feedback". Doğa. 399 (6735): 466–470. Bibcode:1999Natur.399..466L. doi:10.1038/20933. PMID 10365958. S2CID 4403659.
- ^ Teramitsu I; Kudo LC; London SE; Geschwind DH & White SA (2004). "Parallel FoxP1 and FoxP2 expression in songbird and human brain predicts functional interaction". J. Neurosci. 24 (13): 3152–63. doi:10.1523/JNEUROSCI.5589-03.2004. PMC 6730014. PMID 15056695.
- ^ Nottebohm, F. (2004). "The road we travelled: discovery, choreography, and significance of brain replaceable neurons". New York Bilimler Akademisi Yıllıkları. 1016 (1): 628–658. Bibcode:2004NYASA1016..628N. doi:10.1196/annals.1298.027. PMID 15313798. S2CID 11828091.
- ^ Brenowitz, Eliot A. & Michael D. Beecher (2005). "Song learning in birds: diversity and plasticity, opportunities and challenges" (PDF). Sinirbilimlerindeki Eğilimler. 28 (3): 127–132. doi:10.1016/j.tins.2005.01.004. PMID 15749165. S2CID 14586913.
- ^ Slater, P. J. B. (1989). "Bird song learning: causes and consequences". Ethol. Ecol. Evol. 1: 19–46. doi:10.1080/08927014.1989.9525529.
- ^ Thorpe, W. (1954). "The process of song-learning in the chaffinch as studied by means of the sound spectrograph". Doğa. 173 (4402): 465–469. Bibcode:1954Natur.173..465T. doi:10.1038/173465a0. S2CID 4177465.
- ^ Metzmacher, M. (2016). "Alauda: Chaffinch song learning : Thorpe conclusions revisited". Alauda. 84: 465–469. hdl:2268/204189.
- ^ Konishi, M. (1965). "The role of auditory feedback on the control of vocalization in the white-crowned sparrow". Tierpsychologie için Zeitschrift. 22 (7): 770–783. doi:10.1111/j.1439-0310.1965.tb01688.x (etkin olmayan 2020-11-09). PMID 5874921.CS1 Maint: DOI Kasım 2020 itibarıyla etkin değil (bağlantı)
- ^ Marler, P. (1970). "A comparative approach to vocal learning: Song development in the white-crowned sparrows". Karşılaştırmalı ve Fizyolojik Psikoloji Dergisi. 71 (2, Pt.2): 1–25. doi:10.1037/h0029144.
- ^ Nordeen, K.W.; Nordeen, E.J. (1994). "Auditory feedback is necessary for the maintenance of stereotyped song in adult zebra finches". Behavioral and Neural Biology. 71 (1): 58–66. doi:10.1016/0163-1047(92)90757-U. PMID 1567334.
- ^ Leonardo, A. (2004). "Experimental test of error-correction birdsong model". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 101 (48): 16935–16940. doi:10.1073/pnas.0407870101. PMC 534752. PMID 15557558.
- ^ a b Rizzolatti G.; Craighero L. (2004). "The mirror-neuron system". Annu. Rev. Neurosci. 27: 169–192. doi:10.1146/annurev.neuro.27.070203.144230. PMID 15217330. S2CID 1729870.
- ^ Oberman L. M.; Pineda J. A.; Ramachandran V. S. (2007). "The human mirror neuron system: A link between action observation and social skills". Sosyal Bilişsel ve Duyuşsal Sinirbilim. 2 (1): 62–66. doi:10.1093/scan/nsl022. PMC 2555434. PMID 18985120.
- ^ a b Prather J. F.; Peters S.; Nowicki S.; Mooney R. (2008). "Precise auditory-vocal mirroring in neurons for learned vocal communication". Doğa. 451 (7176): 305–310. Bibcode:2008Natur.451..305P. doi:10.1038/nature06492. PMID 18202651. S2CID 4344150.
- ^ a b Dinstein I.; Thomas C.; Behrmann M.; Heeger D.J. (2008). "A mirror up to nature". Güncel Biyoloji. 18 (1): R13–18. doi:10.1016/j.cub.2007.11.004. PMC 2517574. PMID 18177704.
- ^ Tchernichovski O.; Wallman J. (2008). "Behavioral neuroscience: Neurons of imitation". Doğa. 451 (7176): 249–250. Bibcode:2008Natur.451..249T. doi:10.1038/451249a. PMID 18202627. S2CID 205035217.
- ^ Miller G (2008). "Mirror neurons may help songbirds stay in tune". Bilim. 319 (5861): 269. doi:10.1126/science.319.5861.269a. PMID 18202262. S2CID 34367648.
- ^ Richard Mooney (5 June 2014). "Auditory–vocal mirroring in songbirds" (PDF). Philosophical Transactions of the Royal Society B: Biological Sciences Online. Alındı 16 Aralık 2015.[kalıcı ölü bağlantı ]
- ^ a b c Riebel, Katharina; Lachlan, Robert F.; Slater, Peter J. B. (2015-05-01), Naguib, Marc; Brockmann, H. Jane; Mitani, John C.; Simmons, Leigh W. (eds.), "Chapter Six - Learning and Cultural Transmission in Chaffinch Song", Advances in the Study of BehaviorAkademik Basın, 47, pp. 181–227, doi:10.1016/bs.asb.2015.01.001, alındı 2020-01-30
- ^ Thorpe, W. H. (2008-06-28). "The Learning of Song Patterns by Birds, with Especial Reference to the Song of the Chaffinch Fringilla Coelebs". İbis. 100 (4): 535–570. doi:10.1111/j.1474-919x.1958.tb07960.x. ISSN 0019-1019.
- ^ Slater, P. J. B. (1983-04-01). "Chaffinch Imitates Canary Song Elements and Aspects of Organization". Auk. 100 (2): 493–495. doi:10.1093/auk/100.2.493. ISSN 0004-8038.
- ^ Slater, P. J. B .; Ince, S. A. (1979). "Cultural Evolution in Chaffinch Song". Davranış. 71 (1/2): 146–166. doi:10.1163/156853979X00142. ISSN 0005-7959. JSTOR 4534000.
- ^ Tanaka, Masashi; Sun, Fangmiao; Li, Yulong; Mooney, Richard (2018). "A mesocortical dopamine circuit enables the cultural transmission of vocal behaviour". Doğa. 563 (7729): 117–120. Bibcode:2018Natur.563..117T. doi:10.1038/s41586-018-0636-7. ISSN 1476-4687. PMC 6219627. PMID 30333629.
- ^ a b c Lachlan, Robert F.; Slater, Peter J. B. (1999-04-07). "The maintenance of vocal learning by gene–culture interaction: the cultural trap hypothesis". Kraliyet Cemiyeti B Bildirileri: Biyolojik Bilimler. 266 (1420): 701–706. doi:10.1098/rspb.1999.0692. ISSN 0962-8452. PMC 1689831.
- ^ Schmidt, K. L.; MacDougall-Shackleton, E. A.; Kubli, S. P.; MacDougall-Shackleton, S. A. (2014-06-20). "Developmental Stress, Condition, and Birdsong: A Case Study in Song Sparrows". Bütünleştirici ve Karşılaştırmalı Biyoloji. 54 (4): 568–577. doi:10.1093/icb/icu090. ISSN 1540-7063. PMID 24951504.
- ^ S., Nowicki; W., Searcy; S., Peters (2002-12-01). "Brain development, song learning and mate choice in birds: a review and experimental test of the "nutritional stress hypothesis"". Karşılaştırmalı Fizyoloji Dergisi A. 188 (11–12): 1003–1014. doi:10.1007/s00359-002-0361-3. ISSN 0340-7594. PMID 12471497. S2CID 14298372.
- ^ Boogert, N. J.; Fawcett, T. W.; Lefebvre, L. (2011-04-18). "Mate choice for cognitive traits: a review of the evidence in nonhuman vertebrates". Davranışsal Ekoloji. 22 (3): 447–459. doi:10.1093/beheco/arq173. ISSN 1045-2249.
- ^ Boogert, Neeltje J .; Anderson, Rindy C.; Peters, Susan; Searcy, William A.; Nowicki, Stephen (2011). "Song repertoire size in male song sparrows correlates with detour reaching, but not with other cognitive measures". Hayvan Davranışı. 81 (6): 1209–1216. doi:10.1016/j.anbehav.2011.03.004. ISSN 0003-3472. S2CID 21724914.
- ^ Saunders, Aretas A (1951) Guide to Bird Songs. Doubleday ve Şirket
- ^ Sibley, David (2000). Sibley Kuş Rehberi. Knopf. ISBN 978-0-679-45122-8.
- ^ Sarvasy, Hannah (2016). "Warblish: Verbal Mimicry of Birdsong". Etnobiyoloji Dergisi. 36 (4): 765–782. doi:10.2993/0278-0771-36.4.765. hdl:1885/112092. ISSN 0278-0771. S2CID 89990174.
- ^ Baker, Myron C. (2001). "Bird Song Research: The Past 100 years" (PDF). Kuş Davranışı. 14: 3–50.
- ^ Bailey, C. E. G (2008). "Towards an Orthography of Bird Song". İbis. 92: 115–131. doi:10.1111/j.1474-919X.1950.tb01739.x.
- ^ Borror, Donald J.; Reese, Carl R. (1953). "The Analysis of Bird Songs by Means of a Vibralyzer" (PDF). Wilson Bülteni. 65 (4): 271–276.
- ^ Thorpe, W. H. (1958). "The learning of song patterns by birds, with especial reference to the song of the chaffinch Fringilla coelebs". İbis. 100 (4): 535–570. doi:10.1111/j.1474-919X.1958.tb07960.x.
- ^ Slater, P. J. B. (2003). "Fifty years of bird song research: a case study in animal behaviour". Hayvan Davranışı. 65 (4): 633–639. doi:10.1006/anbe.2003.2051. S2CID 53157104.
- ^ Robbins, Chandler S.; Bertel Bruun; Herbert S. Zim; Arthur Singer (1983). A Guide To Field Identification: Birds of North America. Golden Field Guides (Second ed.). Western Publishing Company. s.14. ISBN 978-0-307-33656-9.
- ^ Meijer, P.B.L. (1992). "An Experimental System for Auditory Image Representations". Biyomedikal Mühendisliğinde IEEE İşlemleri. 39 (2): 112–121. doi:10.1109/10.121642. PMID 1612614. S2CID 34811735.
- ^ "US Patent. 20030216649. Audible output sonogram analyzer". Freepatentsonline.com. 2003-11-20. Alındı 2014-06-03.
- ^ Alström, P .; Ranft, R. (2003). "The use of sounds in avian systematics, and the importance of bird sound archives". İngiliz Ornitologlar Kulübü Bülteni. 123A: 114–135.
- ^ Alström, P. (1998). "Taxonomy of the Mirafra assamica complex" (PDF). Çatal kuyruk. 13: 97–107. Arşivlenen orijinal (PDF) 2008-03-07 tarihinde.
- ^ "App - Bird Song Id USA: Nature Apps - Sunbird". sunbird.tv. Alındı 2016-06-08.
- ^ Munford, Monty (2015-03-28). "Don't know birdsong? There's a (great) app for that". Telegraph.co.uk. Alındı 2016-06-08.
- ^ Collias, N. E. (1987). "The vocal repertoire of the Red Junglefowl: A spectrographic classification and the code of communication". Akbaba. 89 (3): 510–524. doi:10.2307/1368641. JSTOR 1368641. S2CID 87662926.
- ^ Evans, C. S.; Makedonya, J. M .; Marler, P. (1993). "Effects of apparent size and speed on the response of chickens, Gallus gallus, to computer-generated simulations of aerial predators". Hayvan Davranışı. 46: 1–11. doi:10.1006/anbe.1993.1156. S2CID 53197810.
- ^ a b Hailman, Jack; Ficken, Millicent (1986). "Combinatorial animal communication with computable syntax: Chick-a-dee calling qualifies as "language" by structural linguistics". Hayvan Davranışı. 34 (6): 1899–1901. doi:10.1016/S0003-3472(86)80279-2. S2CID 53172611.
- ^ Pepperberg, I.M. (2000). The Alex Studies: Cognitive and Communicative Abilities of Grey Parrots. Harvard Üniversitesi Yayınları.
- ^ Marcus, Gary F. (2006-04-27). "Language: Startling starlings". Doğa. 440 (7088): 1117–1118. Bibcode:2006Natur.440.1117M. doi:10.1038/4401117a. PMID 16641976. S2CID 14191866.
- ^ Young, Jon (2008). "Bird Language: Exploring the Language of Nature with Jon Young". OWLink Media. Arşivlenen orijinal 2010-07-28 tarihinde.
- ^ Birds Can Tell Us a Lot about Human Language Adam Fishbein, February 2, 2018 (Bilimsel amerikalı )
- ^ Birds use language like humans joining calls together to form sentences, March 8, 2015 The Daily Telegraph (First published in Nature Communications)
- ^ a b "Archive Pioneers - Ludwig Koch and the Music of Nature". BBC Arşivleri. BBC. 2009-04-15. Alındı 2 Eylül 2011.
- ^ Matthew Head (1997). "Birdsong and the Origins of Music". Kraliyet Müzik Derneği Dergisi. 122 (1): 1–23. doi:10.1093/jrma/122.1.1.
- ^ Clark, Suzannah (2001). Music Theory and Natural Order from the Renaissance to the Early Twentieth Century. Cambridge University Press. ISBN 978-0-521-77191-7.
- ^ Reich, Ronni (15 October 2010). "NJIT professor finds nothing cuckoo in serenading our feathered friends". Yıldız Defteri. Alındı 19 Haziran 2011.
- ^ Taylor, Hollis (2011-03-21). "Composers' Appropriation of Pied Butcherbird Song: Henry Tate's "undersong of Australia" Comes of Age". Online Müzik Araştırmaları Dergisi. 2.
- ^ Rothenberg, David (2005). Why Birds Sing. Allen Lane.
- ^ Hareket, Andrew (10 Aralık 2005). "Tam uçuşta". Gardiyan. Alındı 24 Nisan 2016.
- ^ "Kuşlar Neden Şarkı Söylüyor". Britanya Yayın Şirketi (BBC Dört). 1 Kasım 2010. Alındı 24 Nisan 2016.
- ^ Underwood, Emily (15 Ağustos 2016). "Birdsong Müzik Değil, Sonuçta". Bilim. Alındı 24 Nisan 2016.
- ^ Baptista, Luis Felipe; Keister, Robin A. (2005). "Neden Birdsong Bazen Müzik Gibi". Biyoloji ve Tıp Alanında Bakış Açıları. 48 (3): 426–443. doi:10.1353 / pbm.2005.0066. PMID 16085998. S2CID 38108417.
- ^ Taylor, Hollis (2017). "Is Birdsong Music? Outback Encounters with an Australian Songbird". Indiana University Press. Alındı 2017-06-25.
- ^ Tierney, Adam T .; Russo, Frank A .; Patel, Aniruddh D. (2011). "İnsan ve kuş şarkı yapısının motor kökenleri". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 108 (37): 15510–15515. Bibcode:2011PNAS..10815510T. doi:10.1073 / pnas.1103882108. PMC 3174665. PMID 21876156.
- ^ Attar, Farid al-Din (1984). Darbandi, Afkham; Davis, Dick (eds.). Kuşların Konferansı. Penguen Klasikleri. ISBN 978-0-14-044434-6.
- ^ Brooks, Cleanth; Warren, Robert Penn (1968). Stillinger, Jack (ed.). Bülbüle Ode. Keats's Odes. Prentice-Hall. sayfa 44–47.
- ^ Sandy, Mark (2002). "To a Skylark". Edebiyat Ansiklopedisi. Alındı 22 Nisan 2016.
- ^ "Crow - The Ted Hughes Society Journal". The Ted Hughes Society. 2012. Arşivlenen orijinal 2 Temmuz 2015. Alındı 22 Nisan 2016.
- ^ Hopkins, Gerard Manley (1985). Şiirler ve Düzyazı. Penguin Books. ISBN 9780140420159.
Dış bağlantılar
- Avian Vocalizations Center Michigan Eyalet Üniversitesi
- The Nature Explorers Audio and video of Western North American birds.
- Bird Language: Exploring the Language of Nature with Jon Young A blog with stories and tips for learning the patterns in bird vocalizations.
- Large collection of audio bird calls collected in Arizona itibaren Ask A Biologist.
- xeno-canto: Community online database of downloadable bird sounds from around the globe ≈250,000 recordings of ≈9400 species as of Aug 2015. See also xeno-canto.
- British Library's archive of bird sounds representing more than 8,000 species.
- The Sound Approach A guide to understanding bird sound.
- Doğayı Dinle includes article "The Language of Birds"
- Bird language articles
- Bird songs in movies: an unnatural history Humor piece on soundtrack errors
- How do Birds Sing? The mechanics and anatomy of birdsong production
- Song Bird Science Shared resource for birdsong scientists
- Bioacoustic Research Program -de Cornell Ornitoloji Laboratuvarı distributes a number of different free birdsong synthesis & analysis programs.
- Macaulay Kütüphanesi -de Cornell Ornitoloji Laboratuvarı dünyanın en büyük hayvan sesleri ve ilgili video koleksiyonudur.
- Audio Pitch Tracer Accurate transcription of clean recordings of bird vocalizations to midi
| Aktiviteler |  | |
|---|---|---|
| Ürün:% s | ||
| Sanatta |
| |
| Türler | ||
| İnsanlar |
| |
| İlişkili |
| |