Örümcek ağı - Spider silk




örümcek ağı bir protein lif tarafından bükülmüş örümcekler. Örümcekler kendi ipek yapmak ağlar ya da diğer hayvanları yakalamak için yapışkan ağlar olarak ya da yavrularını korumak ya da avını sarmak için yuva ya da koza işlevi gören diğer yapılar. Ayrıca ipeklerini kendilerini askıya almak için de kullanabilirler. havada süzülmek veya yırtıcılardan uzaklaşmak için. Çoğu örümcek, ipeklerinin kalınlığını ve yapışkanlığını farklı kullanımlar için değiştirir.
Bazı durumlarda, örümcekler ipeği yiyecek kaynağı olarak bile kullanabilirler.[1] Örümcekten zorla ipek toplamak için yöntemler geliştirilirken,[2] Birçok örümcekten ipek toplamak, örneğin ipek eğiren organizmalara kıyasla zordur. ipekböcekleri.
Bütün örümcekler üretir ipek ve hatta web olmayan binada örümcekler ipek, kur yapma ve çiftleşmeye çok yakından bağlıdır. Dişiler tarafından üretilen ipek, erkek titreşimli kur sinyalleri için bir iletim kanalı sağlarken, ağlar ve çekme halatları dişi cinsiyet feromonları için bir alt tabaka sağlar. Cinsel etkileşimler sırasında ipek üreten erkek örümceklerin gözlemleri, filogenetik olarak yaygın taksonlar arasında da yaygındır. Bununla birlikte, erkek tarafından üretilen ipeğin çiftleşmedeki işlevi çok az çalışılmıştır.[3]
Biyoçeşitlilik
Kullanımlar
Tüm örümcekler ipek üretir ve tek bir örümcek farklı kullanımlar için yedi farklı türde ipek üretebilir.[4] Bu, bir bireyin genellikle yalnızca bir tür ipek ürettiği böcek ipeklerinin tersidir.[5] Örümcek ipekleri, her biri ipeğin işlevine uygun özelliklere sahip birçok farklı ekolojik yolla kullanılabilir. Örümcekler geliştikçe, ipeklerinin karmaşıklığı ve çeşitli kullanımları da gelişir, örneğin 300-400 milyon yıl önceki ilkel tüp ağlarından 110 milyon yıl önceki karmaşık küre ağlarına.[6]
| Kullanım | Misal | Referans |
|---|---|---|
| Av yakalama | Tarafından üretilen küre ağları Araneidae (tipik orb-weavers); tüp ağları; dolaşan ağlar; tabaka ağları; dantel ağlar, kubbe ağlar; Bolas örümcekleri tarafından "balık tutmak" için kullanılan tek iplik. | [4][6] |
| Av immobilizasyonu | İpek, avı sarmak için "sarma bantları" olarak kullanılır. Genellikle bir zehir kullanarak hareketsiz kılan av ile birleştirilir. Türlerinde Scytodes ipek, zehirle birleştirilir ve Chelicerae. | [4] |
| Üreme | Erkek örümcekler sperm ağları üretebilir; örümcek yumurtaları ipek kozalarla kaplıdır. | [4][7] |
| Dağılım | "Balon" veya "uçurtma" küçük örümcekler tarafından havada süzülmek için, örneğin dağılma amacıyla kullanılır. | [8] |
| Yiyecek kaynağı | kleptoparazitik Argyrodes ev sahibi örümcek ağlarının ipeğini yemek. Bazı günlük geçici ağ dokumacıları da her gün kendi kullanılmayan ipeklerini yiyorlar, böylece ağır bir metabolik masrafı azaltıyor. | [1][9] |
| Yuva astarı ve yuva yapımı | Avrupa tüp ağ örümceği gibi "ilkel" örümcekler tarafından kullanılan tüp ağlar (Segestria florentina ). İplikler, dışarıya duyusal bir bağlantı sağlamak için yuvanın dışına yayılır. İpek, aile üyeleri gibi "tuzak kapı" kullanan örümceklerin kapaklarının bir bileşenidir. Ctenizidae ve "su" veya "dalış zili" örümceği Argyroneta aquatica ipek dalış çanını yapıyor. | [6] |
| Kılavuz çizgiler | Barınaktan kaçan bazı örümcekler, eve dönüş yolunu tekrar bulmak için bir ipek izi bırakacaktır. | [9] |
| Düşme çizgileri ve bağlantı çizgileri | Gibi birçok örümcek Salticidae sığınaktan çıkıp ipek izi bırakan, ters veya dikey yüzeylerden düşme durumunda acil durum hattı olarak kullanın. Diğerleri, hatta web sakinleri bile, alarma geçtiğinde bir ağdan kasıtlı olarak düşecek ve zamanı geldiğinde geri dönebilecekleri bir bırakma hattı olarak ipeksi bir ip kullanacaktır. Türler gibi bazıları Paramystaria, ayrıca besleme sırasında bir damla hattından sarkacaktır. | [9] |
| Alarm hatları | Gerçek tuzak ağlarını döndürmeyen bazı örümcekler, avlarının ayaklarının (karıncalar gibi) rahatsız edebileceği alarm ağları düzenler, örümceğin acele edip yemeği yeterince küçükse onu güvence altına almasını veya temastan kaçınmasını sağlar. davetsiz misafir çok zorlu görünüyor. | [9] |
| Feromonal yollar | Bazı gezgin örümcekler, karşı cinsin bir eş bulmak için izleyebileceği feromonlarla emprenye edilmiş büyük ölçüde kesintisiz bir ipek izi bırakacaktır. | [9] |
Türler

Tüm bu ekolojik kullanımlar için şartnamenin karşılanması, bir elyaf, bir elyaf yapısı veya bir ipek-globül gibi farklı geniş özelliklere uygun farklı ipek türleri gerektirir. Bu türler arasında yapıştırıcılar ve lifler bulunur. Bazı lif türleri yapısal destek için kullanılırken, diğerleri koruyucu yapılar inşa etmek için kullanılır. Bazıları enerjiyi etkili bir şekilde emebilir, bazıları ise titreşimi verimli bir şekilde iletebilir. Bir örümcekte bu ipek türleri farklı bezlerde üretilir; böylece belirli bir salgı bezinden alınan ipek, örümcek tarafından kullanımına bağlanabilir.
| Bez | İpek Kullanımı |
|---|---|
| Ampullate (majör) | Dragline ipeği - ağın dış kenarı ve parmaklıkları için, ayrıca cankurtaran halatı ve balonlama için kullanılır. |
| Ampullate (minör) | Web yapımı sırasında geçici iskele yapımında kullanılır. |
| Flagelliform | Yakalama sarmal ipek - ağın hatlarını yakalamak için kullanılır. |
| Tubuliform | Yumurta koza ipeği - koruyucu yumurta keseleri için kullanılır. |
| Asinform | Yeni yakalanan avı sarmak ve sabitlemek için kullanılır; erkek sperm ağlarında kullanılır; stabilimenta'da kullanılır. |
| Agrega | Yapışkan küreciklerin ipek tutkalı. |
| Piriform | Bağlantı noktaları için ayrı dişler arasında bağ oluşturmak için kullanılır. |
Özellikleri
Mekanik özellikler
Her örümcek ve her ipek türü, biyolojik işlevleri için optimize edilmiş bir dizi mekanik özelliğe sahiptir.
Çoğu ipek, özellikle de çekme halatlı ipek, olağanüstü mekanik özelliklere sahiptir. Eşsiz bir yüksek kombinasyon sergilerler. gerilme direnci ve genişletilebilirlik (süneklik ). Bu, bir ipek lifinin kırılmadan önce büyük miktarda enerji emmesini sağlar (sertlik, altındaki alan bir gerilme-uzama eğrisi).
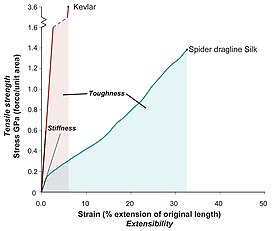
Ana akım medyada sık sık yapılan bir hata, ipeği diğer malzemelerle karşılaştırırken güç ve tokluğu karıştırmaktır.[kaynak belirtilmeli ] Ağırlık için ağırlık, ipek çelikten daha güçlüdür, ancak Çelik yelek. İpek her ikisinden de daha serttir.
Örümcek ipeği liflerinin mekanik özelliklerinin değişkenliği önemli olabilir ve moleküler hizalanma dereceleri ile ilgilidir.[10] Mekanik özellikler, büyük ölçüde ortam koşullarına, yani nem ve sıcaklığa bağlıdır.[11]
Gücü
Bir dragline ipek gerilme direnci yüksek dereceli alaşım ile karşılaştırılabilir çelik (450−2000 MPa),[12][13] ve yaklaşık yarısı kadar güçlü aramid filamentler, örneğin Twaron veya Kevlar (3000 MPa).[14]
Yoğunluk
Ağırlıklı olarak proteinden oluşan ipekler, çeliğin yoğunluğunun yaklaşık altıda biri kadardır (1,3 g / cm3). Sonuç olarak, Dünya'yı çevreleyecek kadar uzun bir telin ağırlığı 500 gramdan (18 ons) az olacaktır. (Spider dragline ipeğinin gerilme mukavemeti kabaca 1.3'tür.GPa. Çelik için listelenen gerilme mukavemeti biraz daha yüksek olabilir - ör. 1.65 GPa,[15][16] ancak örümcek ipeği çok daha az yoğun bir malzemedir, bu nedenle belirli bir örümcek ipeği ağırlığı, aynı ağırlıktaki çelikten beş kat daha güçlüdür.)
Enerji yoğunluğu
enerji yoğunluğu dragline örümcek ipeği kabaca 1.2×108 J / m3.[17]
Genişletilebilirlik
İpekler de son derece sünek Bazıları gevşemiş uzunluklarının beş katına kadar kopmadan uzanabiliyor.
Tokluk
Mukavemet ve süneklik kombinasyonu, dragline ipeklerine çok yüksek sertlik (veya kırılmaya çalış), ki bu "ticari poliaramid (aromatik naylon ) modern polimer elyaf teknolojisinin mihenk taşları olan filamentler ".[18][19]
Sıcaklık
Doğası gereği alakalı olma ihtimali düşük olsa da, çekme halatlı iplikler güçlerini −40 ° C ((40 ° F) altında ve 220 ° C (428 ° F) 'ye kadar tutabilir.[20] Birçok malzemede olduğu gibi, örümcek ipeği lifleri bir cam geçiş. Cam geçiş sıcaklığı neme bağlıdır, çünkü su bir yumuşatıcı ipek için.[11]
Süper kontraksiyon
Suya maruz kaldığında, çekme halat ipleri süper büzülmeye uğrar,% 50'ye kadar küçülür ve gerilim altında zayıf bir kauçuk gibi davranır.[11] Doğada kullanımına dair birçok hipotez öne sürülmüştür; en popüler olanı, sabah çiyini kullanarak gece inşa edilen ağları otomatik olarak germektir.[kaynak belirtilmeli ]
En yüksek performans
Bilinen en sert örümcek ipeği türler tarafından üretilir Darwin'in havlayan örümcek (Caerostris darwini): "Zorla ipe dizilen liflerin dayanıklılığı ortalama 350 MJ / m3 bazı örnekler 520 MJ / m'ye ulaşıyor3. Böylece, C. darwini ipek, daha önce tarif edilen ipeğin iki katından daha sert ve Kevlar'dan 10 kat daha serttir ".[21]
Yapıştırıcı özellikleri
İpek elyaf iki bileşenli bir piriform en az ipek alt tabaka kullanarak ipek iplikleri çeşitli yüzeylere yapıştırmak için kullanılan desenler ("bağlantı diskleri" olarak adlandırılır) şeklinde bükülür.[22] Piriform iplikler polimerize etmek ortam koşullarında hemen işlevsel hale gelir ve süresiz olarak kullanılabilir, biyolojik olarak parçalanabilir, çok yönlü ve çevrede bulunan diğer birçok malzeme ile uyumlu kalır.[22] Bağlantı diskinin yapışkan ve dayanıklılık özellikleri, düzelerdeki işlevlerle kontrol edilir.[23] İpeğin bazı yapışkan özellikleri benzer tutkal oluşan mikrofibriller ve lipit muhafazalar.[22]
İpek türleri
Birçok örümcek türünün farklı bezler konut dahil farklı amaçlar için farklı özelliklere sahip ipek üretmek, ağ inşaat, savunma, yakalama ve alıkoyma Av, yumurta koruması ve hareketlilik (ince "gossamer" iplik balonlaşma veya örümceğin ipek sıkılırken aşağı düşmesine izin veren bir iplik için). Farklı kullanımlara uygun özelliklere sahip farklı özel ipekler gelişmiştir. Örneğin, Argiope argentata her biri farklı bir amaç için kullanılan beş farklı ipek türü vardır:[24][25]
| İpek | Kullanım |
|---|---|
| büyük ampullate (dragline) ipek | Ağın dış kenarı ve parmaklıkları için ve ayrıca cankurtaran halatı için kullanılır. Birim ağırlık başına çelik kadar güçlü olabilir, ancak çok daha serttir. |
| yakalama sarmal (flagelliform) ipek | Ağın yakalama hatları için kullanılır. Yapışkan, son derece esnek ve sağlam. Yakalama spirali, spirale yerleştirilen agrega damlacıkları (örümcek tutkalı) nedeniyle yapışkandır. Flagelliformun esnekliği, agregatın ağa uçan hava avına yapışması için yeterli zaman sağlar. |
| tubiliform (a.k.a. cylindriform) ipek | Koruyucu yumurta keseleri için kullanılır. En sert ipek. |
| asiniform ipek | Yeni yakalanan avı sarmak ve sabitlemek için kullanılır. Dragline dahil diğer ipeklerden iki ila üç kat daha sert. |
| küçük ampülat ipek | Web yapımı sırasında geçici iskele yapımında kullanılır. |
| Piriform (piriform) | Piriform, çekme ipi için bağlantı diski görevi görür. Piriform, sabit bir ağ oluşturmak için örümcek ipeklerini birbirine yapıştırmada kullanılır. |
Yapısal
Protein hiyerarşisine kadar makroskopik yapı
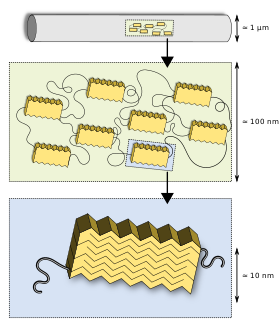
Diğer birçok biyomalzeme gibi ipekler de hiyerarşik bir yapıya sahiptir. Birincil yapı ... amino asit proteinlerinin dizisi (Spidroin ), esas olarak yüksek oranda tekrarlayan glisin ve alanin bloklarından oluşan,[26][27] bu nedenle ipekler genellikle bir blok ko-polimer olarak anılır. İkincil bir yapı seviyesinde, kısa yan zincirli alanin esas olarak kristalin alanlarda bulunur (beta sayfaları ), glisin çoğunlukla sarmal ve beta dönüş yapılarından oluşan sözde amorf matriste bulunur.[27][28] Örümcek ipeğine olağanüstü özelliklerini veren, sert kristalin segmentler ile gergin elastik yarı amorf bölgeler arasındaki etkileşimdir.[29][30] Lifin özelliklerini geliştirmek için protein dışında çeşitli bileşikler kullanılır. Pirolidin higroskopik özelliklere sahiptir ve ipeği nemli tutar ve aynı zamanda karınca istilasını engeller. Özellikle tutkal iplerinde yüksek konsantrasyonda oluşur. Potasyum hidrojen fosfat Salıverme hidrojen iyonları sulu çözelti içinde pH yaklaşık 4, ipek yapmak asidik ve böylece onu mantarlar ve bakteri aksi takdirde proteini sindirecektir. Potasyum nitrat proteinin asidik ortamda denatüre olmasını engellediğine inanılmaktadır.[31]
Bu ilk çok temel ipek modeli 1994 yılında Termonia tarafından tanıtıldı.[32] hidrojen bağları ile birbirine bağlanmış amorf bir matris içine gömülü kristalitleri önerdi. Bu model yıllar içinde geliştirildi: yarı kristal bölgeler bulundu[27] örümcek ipeği için önerilen bir fibriler deri çekirdek modelinin yanı sıra,[33] tarafından görselleştirildi AFM ve TEM.[34] Nanofibriler yapının boyutları ve kristal ve yarı kristal bölgelerin boyutları nötron saçılması.[35]
Liflerin mikroyapısal bilgileri ve makroskopik mekanik özellikleri ile ilişkilendirmek mümkün olmuştur.[36] Sonuçlar, sıralı bölgelerin (i) esas olarak düşük gerilmiş lifler için deformasyonla yeniden yönlendirildiğini ve (ii) sıralı bölgelerin fraksiyonunun, liflerin daha yüksek gerilmesi için giderek arttığını göstermektedir.

Örümceğin küre ağının, yapısal modüllerinin ve örümcek ağı yapısının şeması.[37] Solda bir küre ağının şematik bir çizimi gösterilmektedir. Kırmızı çizgiler, çekme çizgisini, radyal çizgiyi ve çerçeve çizgilerini temsil eder, mavi çizgiler spiral çizgiyi temsil eder ve küre ağının merkezi "göbek" olarak adlandırılır. Agrega bezinden salgılanan viskoz malzeme ile mavi renkle çizilmiş yapışkan toplar eşit aralıklarla spiral hat üzerinde yapılır. Piriform bezden salgılanan ataşman çimentosu, farklı hatları bağlamak ve sabitlemek için kullanılır. Mikroskobik olarak, örümcek ipeği ikincil yapısı spidroin'den oluşur ve sağ tarafta gösterilen yapıya sahip olduğu söylenir. Çekme çizgisi ve radyal çizgide, kristal bir-levha ve amorf bir sarmal yapı birbirine dokunmuştur. Büyük miktarda β-spiral yapısı, küre ağının yakalama kısmına elastik özellikler kazandırır. Yapısal modüller diyagramında, üst orta kısımda gösterildiği gibi, esas olarak MaSp1 ve MaSp2'nin iki proteininden oluşan çekme kancası ve radyal çizgilerin mikroskobik yapısı gösterilmektedir. Spiral çizgide kristalin β-yaprak bölgesi yoktur.

Protein içermeyen bileşim
Örümcek ipeklerinde topaklaşma davranışını etkileyebilecek ve nihai lifte bir koruma tabakası görevi görebilecek şekerler, lipitler, iyonlar ve pigmentler gibi protein dışında çeşitli bileşikler bulunur.[17]
Biyosentez ve lif eğirme
Örümcek ipeği de dahil olmak üzere ipek üretimi, diğer birçok lifli biyolojik materyalin üretiminden önemli bir yönden farklıdır: saçta keratin, bitkilerin hücre duvarlarında selüloz veya hatta sıkıştırılmış liflerden oluşan lifler olarak sürekli büyümekten ziyade böceklerin dışkı maddesi;[17] özel salgı bezlerinden sıvı ipek öncüsünden talep üzerine "eğrilir".[38]
Eğirme işlemi, örümceğin vücudundan bir lifin, örümceğin bacaklarından, örümceğin kendi ağırlığının altına düşmesi veya insanlar tarafından çekilmesi de dahil olmak üzere başka herhangi bir yöntemle çekilmesiyle gerçekleşir. "Eğirme" terimi yanıltıcıdır çünkü herhangi bir bileşenin dönüşü meydana gelmez, aksine tekstile benzetmeden gelir. Dönen tekerlekler. İpek üretimi bir pultrüzyon,[39] ekstrüzyona benzer şekilde, kuvvetin bir hazneden sıkıştırılmak yerine bitmiş elyafı çekerek indüklendiği incelikle. Eğrilmemiş ipek lifi, ipekten çekilir bezler herhangi bir örümcek türünde hem çok sayıda kopya hem de farklı bez türleri olabilir.[38]
İpek bezi

Bezin görünür veya harici kısmı, düze. Türlerin karmaşıklığına bağlı olarak, örümcekler genellikle çiftler halinde iki ila sekiz memeciye sahip olacaktır. Farklı örümceklerde, bir ucunda açıklığı olan bir keseden, karmaşık, çok bölümlü büyük ampülat bezlerine kadar değişen, oldukça farklı özelleşmiş bezler vardır. altın ipek orb-weavers.[53]
Örümceğin yüzeyinde görülebilen her memecinin arkasında, genel bir formu sağdaki şekilde "Genelleştirilmiş bir bezin şeması" gösterilen bir bez bulunur.
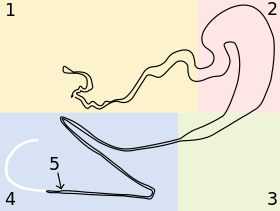
- Bez özellikleri
- Şekil 1'de 1 olarak etiketlenen bezin ilk bölümü, bezin salgı veya kuyruk bölümüdür. Bu bölümün duvarları, bu örümceğin dragline'sinin ana bileşenleri olan Spidroin I ve Spidroin II proteinlerini salgılayan hücrelerle kaplıdır. Bu proteinler, lifin çatlak oluşumunu ve hatta kendi kendini iyileştirmesini önlemeye yardımcı olduğu hipoteziyle, nihai lifin uzunluğu boyunca uzun kanallar oluşturmak için kademeli olarak uzayan damlacıklar biçiminde bulunur.[56]
- İkinci bölüm saklama kesesidir. Bu, örümcek tarafından talep edilene kadar jel benzeri bükülmemiş ipek uyuşturucu maddesini depolar ve korur. Eğrilmemiş ipek jeli depolamaya ek olarak, son lifin yüzeyini kaplayan proteinleri salgılar.[18]
- Huni, saklama kesesinin geniş çapını hızla daralan kanalın küçük çapına indirir.
- Nihai uzunluk, lif oluşumunun çoğunun yeri olan daralan kanaldır. Bu, etrafında birkaç sıkı dönüşe sahip sivrilen bir tüpten, neredeyse uçta (aşağıda 5 numaralı noktada ayrıntılı olarak bahsedilmiştir) sert ipek elyafın çıktığı bir tıkaçla biten bir valften oluşur. Buradaki tüp hiperbolik olarak incelir, bu nedenle bükülmemiş ipek, elyaf oluşumunda önemli bir faktör olan sabit uzama kayma gerilimi altındadır. Kanalın bu bölümü iyon değiştiren, dop pH'ını nötrden asidik hale düşüren ve fiberden suyu uzaklaştıran hücrelerle kaplıdır.[57] Toplu olarak, kayma gerilimi ve iyon ve pH değişiklikleri, sıvı ipek ilacının bir faz geçişine girmesine ve yüksek moleküler organizasyona sahip katı bir protein elyafına yoğunlaşmasına neden olur. Uçtaki tıkaç, elyafın etrafında kenetlenen, elyaf çapını kontrol eden ve suyu daha fazla tutan dudaklara sahiptir.
- Neredeyse sivrilen kanalın sonunda, şekil 1'de yaklaşık konum "5" olarak işaretlenmiş bir valf vardır. Bir süre önce keşfedilmesine rağmen, bu valfin kesin amacı hala tartışılmaktadır. Kırık liflerin yeniden başlatılmasına ve birleştirilmesine yardımcı olduğuna inanılıyor,[58] bir şekilde davranmak sarmal pompa elyafın kalınlığını düzenleyen,[39] ve / veya üzerine bir örümcek düşerken elyafın sıkıştırılması.[58][59] İpek böceğinin ipek presinin benzerliği ve bu kapakçıkların her birinin bu iki organizmada ipek üretiminde oynadığı rol hakkında bazı tartışmalar var.
Süreç boyunca bükülmemiş ipek nematik bir dokuya sahip gibi görünür,[60] benzer şekilde likit kristal, kısmen yüksek protein konsantrasyonundan (hacim başına ağırlık olarak yaklaşık% 30) kaynaklanmaktadır.[61] Bu, bükülmemiş ipeğin kanaldan sıvı olarak akmasına izin verir, ancak moleküler bir düzen sağlar.
Karmaşık bir eğirme alanına örnek olarak, bir yetişkinin düze aparatı Araneus diadematus (bahçe çapraz örümceği) aşağıda gösterilen bezlerden oluşur.[31] Kara dul örümceğinde de benzer çoklu salgı bezi mimarisi mevcuttur.[62]
- Bağlantı noktaları için 500 piriform bez
- Web çerçevesi için 4 ampullate rakor
- Yumurta keselerinin dış kaplaması için ve avı tuzağa düşürmek için yaklaşık 300 asiniform bez
- Yumurta kesesi ipeği için 4 adet tubuliform bez
- Yapıştırıcı fonksiyonlar için 4 agrega rakoru
- Yapışma çizgilerinin dişleri için 2 koronat bez
Yapay sentez

Örümcek ipeğini yapay olarak liflere sentezlemek için örtülmesi gereken iki geniş alan vardır. Bunlar, besleme stoğunun (örümceklerdeki bükülmemiş ipek macunu) sentezi ve eğirme koşullarının (huni, valf, konik kanal ve tıkaç) sentezidir. Birkaç farklı yaklaşım vardı, ancak bu yöntemlerden çok azı liflere verimli bir şekilde sentezlenebilen ipek üretti.
Hammadde
Eğrilmemiş ipeğin moleküler yapısı hem karmaşık hem de son derece uzundur. Bu, ipek liflerine istenen özellikleri kazandırsa da, aynı zamanda lifin kopyalanmasını zorlaştırır. İlgili proteinlerin bir kısmını veya tamamını veya bir kısmını veya tamamını kopyalama girişimlerinin temeli olarak çeşitli organizmalar kullanılmıştır. Bu proteinler daha sonra özütlenmeli, saflaştırılmalı ve özelliklerinin test edilebilmesi için döndürülmelidir.
| Organizma | Detaylar | Ortalama Maksimum kırılma gerilimi (MPa) | Ortalama Gerginlik (%) | Referans |
|---|---|---|---|---|
| Darwin'in havlayan örümceği (Caerostris darwini) | Madagaskar örümcek, nehirler boyunca 25 m uzunluğa kadar ipliklere sahip ağlar yapmakla ünlüdür. "C. darwini ipek, daha önce tanımlanan herhangi bir ipeğin iki katından daha serttir " | 1850 ±350 | 33 ±0.08 | [21] |
| Nephila klavipleri | Tipik altın küre dokuma örümceği | 710–1200 | 18–27 | [63][64] |
| Bombyx mori İpekböcekleri | İpekböcekleri, ölçülen örümcek proteinlerini ve liflerini ifade etmek için genetik olarak değiştirildi.[65] | 660 | 18.5 | [66] |
| E. coli | Büyük ve tekrarlayan bir molekülün sentezlenmesi (~ 300 kDa ) karmaşıktır, ancak en güçlü ipek için gereklidir. Buraya E. coli 556 kDa protein üretmek üzere tasarlanmıştır. Bu sentetik spidroinlerden eğrilen lifler, doğal örümcek ipeğinin mekanik performansını tüm genel ölçütlerle tam olarak kopyalayan ilk liflerdir. | 1030 ±110 | 18 ±6 | [67] |
| Keçi | Keçi sütlerinde ipek proteinleri salgılamak için genetik olarak modifiye edildi ve bunlar daha sonra saflaştırılabilir. | 285–250 | 30–40 | [68] |
| Tütün ve patates bitkileri | Tütün ve Patates bitkiler, ipek proteinleri üretmek için genetik olarak modifiye edildi. Patentler verildi,[69] ancak literatürde henüz hiçbir lif tanımlanmamıştır. | n / a | n / a | [70] |
Geometri
Nispeten basit moleküler yapıya sahip örümcek ipekleri, etkili bir elyafı eğirebilmek için karmaşık kanallara ihtiyaç duyar. Elyaf üretmek için kullanılan, ana türleri aşağıda kısaca tartışılan bir dizi yöntem vardır.
Şırınga ve iğne
Besleme stoğu, bir şırınga kullanılarak içi boş bir iğneden geçirilir. Bu yöntemin lifleri birçok durumda başarılı bir şekilde ürettiği gösterilmiştir.[71][72]
Çok ucuz ve üretilmesi kolay olmasına rağmen, bezin şekli ve koşulları çok gevşek bir şekilde tahmin edilmektedir. Bu yöntem kullanılarak oluşturulan liflerin, çevresel olarak istenmeyen kimyasallar gibi kimyasallarla liften suyu uzaklaştırarak sıvıdan katıya geçmesi için teşvik edilmesi gerekebilir. metanol[73] veya aseton,[72] ve ayrıca istenen özelliklere sahip liflere ulaşmak için lifin sonradan gerilmesini gerektirebilir.[74][71]
Mikroakışkanlar
Alanı olarak mikroakışkanlar olgunlaştıkça, mikroakışkanlar kullanılarak lifleri döndürmek için daha fazla girişimde bulunulması muhtemeldir. Bunlar, çok kontrol edilebilir olma ve çok küçük hacimlerde eğrilmemiş elyafı test etme avantajına sahiptir.[75][76] ancak kurulum ve geliştirme maliyetleri muhtemelen yüksek olacaktır. Bu alanda, doğada bulunan süreci taklit eden bir yöntemde liflerin eğrilmesi için bir patent verilmiştir ve lifler, ticari bir şirket tarafından başarıyla sürekli olarak eğrilmektedir.[77]
Elektrospinning
Elektrospinning bir sıvının kapiler hareket yoluyla dışarı akabilecek şekilde bir kap içinde tutulduğu çok eski bir tekniktir. İletken bir substrat aşağıda konumlandırılır ve sıvı ile substrat arasına elektrik potansiyelinde büyük bir fark uygulanır. Sıvı alt tabakaya çekilir ve küçük lifler neredeyse anında emisyon noktalarından atlar. Taylor koni, alt tabakaya, seyahat ederken kurur. Bu yöntemin hem organizmalardan kesilmiş ipekten hem de organizmalardan nano ölçekli lifler oluşturduğu gösterilmiştir. rejenere ipek fibroin.
İpekten oluşan diğer yapay şekiller
İpek, ilaç dağıtımı için küresel kapsüller, hücre iskeleleri ve yara iyileşmesi, tekstiller, kozmetikler, kaplamalar ve diğerleri gibi diğer şekil ve boyutlarda oluşturulabilir.[78][79] Örümcek ipeği proteinleri ayrıca süperhidrofobik yüzeylerde kendi kendine birleşerek nanoteller ve mikron boyutlu dairesel tabakalar oluşturabilir.[79] Son zamanlarda, rekombinant örümcek ipek proteinlerinin, hücre proliferasyonunu destekleyen protein geçirgen, süper güçlü ve süper esnek membranlar oluşturmak için ayakta duran bir çözeltinin sıvı hava arayüzünde kendi kendine birleşebildiği gösterilmiştir. Önerilen uygulamalar arasında cilt nakilleri ve çip üzerindeki organlarda destekleyici membranlar yer alır.[80]
Araştırma kilometre taşları
Örümcek ipeğinin uzun ve zengin bir geçmişe sahip bilimsel bir araştırma alanı olması nedeniyle, araştırmacıların daha önce yayınlanmış bulguları bağımsız olarak yeniden keşfeden talihsiz olayları olabilir. Aşağıda, bilimsel kabul metriği, alıntılar kullanılarak bilimsel topluluk tarafından ilgili ve önemli olarak kabul edilen, kurucu alanların her birinde yapılan keşiflerin bir tablosu yer almaktadır. Bu nedenle, yalnızca 50 veya daha fazla alıntı içeren makaleler dahil edilmiştir.
| Katkı alanı | Yıl | Başlıca araştırmacılar [Ref] | Makalenin başlığı | Sahaya katkı |
|---|---|---|---|---|
| Kimyasal Temel | 1960 | Fischer, F. & Brander, J.[81] | "Eine Analyze der Gespinste der Kreuzspinne" (örümcek ipeğinin amino asit bileşimi analizi) | |
| 1960 | Lucas, F. & vd.[82][83] | "Eklembacaklı İpek Liflerinin Bileşimi; Fibroinlerle ilgili karşılaştırmalı çalışmalar" | ||
| Gen Dizisi | 1990 | Xu, M. ve Lewis, R.V.[84] | "Bir Protein Süper Elyafının Yapısı - Örümcek Dragline İpek" | |
| Mekanik özellikler | 1964 | Lucas, F.[85] | "Örümcekler ve ipekleri" | Örümcek ipeğinin mekanik özelliklerini ilk kez bilimsel bir makalede diğer malzemelerle karşılaştırdı. |
| 1989 | Vollrath, F. & Edmonds, D. T.[86] | "Örümcek İpeğinin Mekanik Özelliklerinin Su ile Kaplama Yapılarak Modülasyonu" | İpliğin özelliklerini modüle eden örümcek ipeği fibroin ile su etkileşimini öneren ilk önemli makale. | |
| 2001 | Vollrath, F. & Shao, Z.Z.[87] | "Eğirme koşullarının bir örümceğin halatlı halat ipeğinin mekaniği üzerindeki etkisi" | ||
| 2006 | Plaza, G.R., Gine, G.V., Pérez-Rigueiro, J. & Elices, M.[11] | "Dragline ipeğinin termo-higro-mekanik davranışı: Camsı ve lastiksi haller" | Nem ve sıcaklığın mekanik özellikler üzerindeki birleşik etkisi. Cam geçiş sıcaklığının neme bağımlılığı. | |
| Yapısal Karakterizasyon | 1992 | Hinman, M.B. & Lewis, R. V[26] | "İkinci bir dragline ipek fibroin kodlayan bir klonun izolasyonu. Nephila clavipes dragline silk iki proteinli bir fiberdir" | |
| 1994 | Simmons, A. & vd.[88] | "Katı Hal C-13 Nmr of Nephila-Clavipes Dragline Silk, Kristal Bölgelerin Yapısını ve Kimliğini Kurar" | Örümcek ipeğinin ilk NMR çalışması. | |
| 1999 | Shao, Z., Vollrath, F. & vd.[89] | "Örümcek ipeğinin doğal ve süper sıkıştırılmış durumlarda Raman spektroskopisi kullanılarak analizi" | Örümcek ipeğinin ilk Raman çalışması. | |
| 1999 | Riekel, C., Muller, M. & vd.[90] | "Tek örümcek liflerinde X ışını kırınımının özellikleri" | Tek örümcek ipeği liflerinde ilk röntgen. | |
| 2000 | Knight, D.P., Vollrath, F. & vd.[91] | "Dragline örümcek ipliğinin eğrilmesinde beta geçişi ve stres kaynaklı faz ayrımı" | Eğirme sırasında ikincil yapısal geçiş onayı. | |
| 2001 | Riekel, C. ve Vollrath, F.[92] | "Örümcek ipeği lifi ekstrüzyonu: geniş ve küçük açılı X ışını mikro kırınım deneyleri" | Örümcek ipeği esansında ilk röntgen. | |
| 2002 | Van Beek, J. D. & vd.[28] | "Dragline ipeğinin moleküler yapısı: Protein omurgasının katlanması ve yönü" | ||
| Yapı-Mülk İlişkisi | 1986 | Gosline, G.M. & et al.[93] | "Örümcek ipeğinin yapısı ve özellikleri" | Yapıyı örümcek ipeğinin özellikleriyle ilişkilendirmeye ilk girişimi |
| 1994 | Termonia, Y[32] | "Örümcek İpeği Esnekliğinin Moleküler Modellemesi" | Bu yazıda sunulan X-ışını kanıtı; amorf bölgelere gömülü kristalitlerin basit modeli. | |
| 1996 | Simmons, A. & vd.[27] | "Dragline örümcek ağının kristal fraksiyonunun moleküler oryantasyonu ve iki bileşenli yapısı" | Alanin açısından zengin iki tür kristal bölge tanımlandı. | |
| 2006 | Vollrath, F. ve Porter, D.[94] | "Bir arketip protein elastomeri olarak örümcek ipeği" | Grup Etkileşimi Modellemesine dayalı örümcek ipeğine yeni anlayış ve model. | |
| Yerli Eğirme | 1991 | Kerkam, K., Kaplan, D. & vd.[95] | "Doğal İpek Salgılarının Sıvı Kristalliği" | |
| 1999 | Knight, D.P. & Vollrath, F.[96] | "Bir örümceğin ipek üretim hattında sıvı kristaller ve akış uzaması" | ||
| 2001 | Vollrath, F. & Knight, D.P.[17] | "Örümcek ipeğinin sıvı kristal iplikçiliği" | Örümcek ipeği üzerine en çok alıntı yapılan kağıt | |
| 2005 | Gine, G.V., Elices, M., Pérez-Rigueiro, J. & Plaza, G.R.[10] | "Süper sıkıştırılmış liflerin gerilmesi: eğirme ve örümcek ipeğinin değişkenliği arasında bir bağlantı" | Mekanik özelliklerin değişkenliğinin açıklaması. | |
| Sulandırılmış / Sentetik Örümcek İpeği ve Yapay İplik | 1995 | Prince, J. T., Kaplan, D. L. & vd.[97] | "Örümcek Halat İpeği İpeğini Kodlayan Sentetik Genlerin Oluşturulması, Klonlanması ve İfadesi" | Örümcek ipeğinin ilk başarılı sentezi E. coli. |
| 1998 | Arcidiacono, S., Kaplan, D.L. & et al.[98] | "Escherichia coli'de ifade edilen rekombinant örümcek ipeğinin saflaştırılması ve karakterizasyonu" | ||
| 1998 | Seidel, A., Jelinski, L.W. & et al.[99] | "Örümcek İpeğinin Yapay İpliği" | Yeniden yapılandırılmış örümcek ipeğinin ilk kontrollü ıslak eğirme işlemi. |
İnsan kullanır

Güneydeki köylüler Karpat Dağları tarafından inşa edilen tüpleri kesmek için kullanılır Atypus ve yaraları iç astarla kapatın. Bildirildiğine göre iyileşmeyi kolaylaştırdı ve hatta cilt ile bağlantılıydı. Bunun örümcek ipeğinin antiseptik özelliklerinden kaynaklandığına inanılıyor.[101] ve ipek zengindir çünkü K vitamini, kanın pıhtılaşmasında etkili olabilir.[102][Doğrulayın ] Önemli miktarlarda örümcek ipeğinin çıkarılması ve işlenmesindeki zorluklar nedeniyle, bilinen en büyük parça olan kumaş örümcek ipeğinden yapılmış, 11 x 4 fitlik (3.4 x 1.2 m) bir tekstildir. altın yapılan renk Madagaskar 2009 yılında.[103] Seksen iki kişi bir milyondan fazla toplamak için dört yıl çalıştı altın küre örümcekler ve onlardan ipek çıkar.[104]
İpek Nephila klavipleri ilgili araştırmada kullanıldı memeli nöronal yenilenme.[105]
Örümcek ipeği, nişangah teleskoplar, mikroskoplar gibi optik aletlerde,[106] ve teleskopik tüfek nişangahları.[107] 2011 yılında, örümcek ipeği lifleri optik alanında çok ince kırınım desenleri oluşturmak için kullanıldı. N-yarık interferometrik sinyaller optik iletişimde kullanılır.[108] 2012'de, bir dizi keman teli oluşturmak için örümcek ipeği lifleri kullanıldı.[109]
Yöntemlerin geliştirilmesi seri üretim örümcek ipeği, askeri, tıbbi ve tüketim mallarının imalatına yol açmıştır. balistik zırh atletik ayakkabılar, kişisel Bakım Ürün:% s, göğüs implantı ve kateter kaplamalar, mekanik insülin pompalar, moda kıyafetleri ve dış giyim.[110]
Örümcek ipeği askıya almak için kullanılır eylemsizlik hapsi füzyonu önemli ölçüde elastik kaldığından ve 10–20 K kadar düşük sıcaklıklarda kırılabilecek yüksek enerjiye sahip olduğundan, lazer ateşleme sırasında hedefler. Ayrıca, yayılmayan "hafif" atom numaralı elementlerden yapılmıştır. röntgen ışınlama sırasında, füzyon için gerekli basınç farkına ulaşılmayacak şekilde hedefi önceden ısıtabilir.[111]
Örümcek ipeği, insan vücudunun iç kısmının yüksek çözünürlüklü görüntülerini oluşturmak için lazerlerle birlikte kullanılabilecek biyolleri oluşturmak için kullanılmıştır.[1]
Sentetik örümcek ipeği üretme girişimleri
Örümcek ipeğine benzer lifleri üretmek için gereken karmaşık koşulları kopyalamak, araştırma ve erken aşamadaki imalatta zor olmuştur. Vasıtasıyla genetik mühendisliği, Escherichia coli bakteriler, mayalar, bitkiler, ipekböcekleri ve hayvanlar, bir örümceğinkilerden farklı, daha basit özelliklere sahip örümcek ipeği proteinleri üretmek için kullanılmıştır.[110] Sulu bir ortamda protein liflerinin ekstrüzyonu "ıslak eğirme" olarak bilinir. Bu işlem şimdiye kadar, doğal örümcek ipeği için 2,5–4 μm çaplarına kıyasla 10 ila 60 μm arasında değişen çaplarda ipek lifleri üretmiştir. Yapay örümcek ipekleri, doğal halat kazığı ipeğinden daha az ve daha basit proteine sahiptir ve sonuç olarak, doğal çekme ipi ipeğinin yarı çapı, gücü ve esnekliğidir.[110]
- Mart 2010'da, Kore İleri Bilim ve Teknoloji Enstitüsü doğrudan bakterileri kullanarak örümcek ipeği yapmayı başardı E. coli, örümceğin belirli genleri ile değiştirilmiş Nephila klavipleri. Bu yaklaşım, örümcek sağma ihtiyacını ortadan kaldırır ve örümcek ipeğinin daha uygun maliyetli bir şekilde üretilmesine izin verir.[112]
- A 556 kDa spider silk protein was manufactured from 192 repeat motifs of the Nephila klavipleri dragline spidroin, having similar mechanical characteristics as their natural counterparts, i.e., gerilme direnci (1.03 ± 0.11 GPa), modül (13.7 ± 3.0 GPa), extensibility (18 ± 6%), and toughness (114 ± 51 MJ/m3).[67]
- Şirket AMSilk gelişmiş Spidroin using bacteria, making it into an artificial spider silk.[110][113]
- Şirket Bolt Threads produces a recombinant spidroin using yeast, for use in apparel fibers and personal care. They produced the first commercial apparel products made of recombinant spider silk, trademarked Microsilk™, demonstrated in ties and beanies. They have also partnered with vegan activist and luxury designer Stella McCartney Hem de Adidas to produce Microsilk™ garments.[114][115]
- Şirket Kraig Biocraft Laboratories used research from the Universities of Wyoming ve Notre Dame to create silkworms that were genetically altered to produce spider silk.[116][117]
- The now defunct Canadian biyoteknoloji company Nexia successfully produced spider silk protein in transgenik keçiler that carried the gene for it; the milk produced by the goats contained significant quantities of the protein, 1–2 grams of silk proteins per litre of milk. Attempts to spin the protein into a fibre similar to natural spider silk resulted in fibres with tenacities of 2–3 grams per inkarcı.[118] Nexia used wet spinning and squeezed the silk protein solution through small extrusion holes in order to simulate the behavior of the spinneret, but this procedure was not sufficient to replicate the stronger properties of native spider silk.[119]
- The company Spiber has produced a synthetic spider silk that they are calling Q/QMONOS. In partnership with Goldwin, a ski parka made from this synthetic spider silk is currently in testing and is to be in mass production soon for less than $120,000 YEN.[120][121]
Referanslar
- ^ a b Miyashita, Tadashi; Maezono, Yasunori; Shimazaki, Aya (2004). "Silk feeding as an alternative foraging tactic in a kleptoparasitic spider under seasonally changing environments" (PDF). Zooloji Dergisi. 262 (3): 225–229. CiteSeerX 10.1.1.536.9091. doi:10.1017/S0952836903004540.
- ^ Work, Robert W.; Emerson, Paul D. (1982). "An Apparatus and Technique for the Forcible Silking of Spiders". Arachnology Dergisi. 10 (1): 1–10. JSTOR 3705113.
- ^ Scott, Catherine E.; Anderson, Alissa G.; Andrade, Maydianne C. B. (August 2018). "A review of the mechanisms and functional roles of male silk use in spider courtship and mating". Arachnology Dergisi. 46 (2): 173–206. doi:10.1636/JoA-S-17-093.1. ISSN 0161-8202. S2CID 53322197.
- ^ a b c d Foelix, R. F. (1996). Örümcek Biyolojisi. Oxford; New York: Oxford University Press. s.330.
- ^ Sutherland, TD; Young, JH; Weisman, S; Hayashi, CY; Merritt, DJ (2010). "Insect silk: One name, many materials". Yıllık Entomoloji İncelemesi. 55: 171–188. doi:10.1146 / annurev-ento-112408-085401. PMID 19728833.
- ^ a b c Hillyard, P. (2007). The Private Life of Spiders. London: New Holland. s. 160. ISBN 978-1-84537-690-1.
- ^ Nentwig, W. & Heimer, S. (1987). Wolfgang Nentwig (ed.). Ecological aspects of spider webs. Springer-Verlag. s. 211.
- ^ Flying spiders over Texas! Bir uçtan diğer uca. Chad B., Texas State University Undergrad Arşivlendi 26 Kasım 2011 Wayback Makinesi Describes the mechanical kiting of Spider "ballooning".
- ^ a b c d e Holm, Erik, Dippenaar-Schoeman, Ansie; Goggo Kılavuzu; LAPA publishers (URL: WWW.LAPA.co.za). 2010[sayfa gerekli ]
- ^ a b Guinea, G.V.; Elices, M.; Pérez-Rigueiro, J. & Plaza, G.R. (2005). "Stretching of supercontracted fibers: a link between spinning and the variability of spider silk". Deneysel Biyoloji Dergisi. 208 (1): 25–30. doi:10.1242/jeb.01344. PMID 15601874.
- ^ a b c d Plaza, Gustavo R.; Guinea, Gustavo V.; Pérez-Rigueiro, José; Elices, Manuel (2006). "Thermo-hygro-mechanical behavior of spider dragline silk: Glassy and rubbery states". Journal of Polymer Science Part B: Polymer Physics. 44 (6): 994–999. Bibcode:2006JPoSB..44..994P. doi:10.1002/polb.20751.
- ^ Griffiths, J. R.; Salanitri, V. R. (1980). "The strength of spider silk". Malzeme Bilimi Dergisi. 15 (2): 491–496. Bibcode:1980JMatS..15..491G. doi:10.1007/BF00551703. S2CID 135628690.
- ^ "Overview of materials for AISI 4000 Series Steel". www.matweb.com. Alındı 18 Ağustos 2010.
- ^ "DuPont Kevlar 49 Aramid Fiber". www.matweb.com. Alındı 18 Ağustos 2010.
- ^ Ganio Mego, Paolo (c. 2002). "Material Tensile Strength Comparison". Arşivlenen orijinal 26 Ekim 2009. Alındı 3 Ocak 2012.
- ^ Shao, Zhengzhong; Vollrath, F (2002). "Materials: Surprising strength of silkworm silk". Doğa. 418 (6899): 741. Bibcode:2002Natur.418..741S. doi:10.1038/418741a. PMID 12181556. S2CID 4304912.
- ^ a b c d Porter, D .; Vollrath, F .; Shao, Z. (2005). "Predicting the mechanical properties of spider silk as a model nanostructured polymer". Avrupa Fiziksel Dergisi E. 16 (2): 199–206. Bibcode:2005EPJE...16..199P. doi:10.1140/epje/e2005-00021-2. PMID 15729511. S2CID 32385814.
- ^ a b Vollrath, F. & Knight, D. P. (2001). "Liquid crystalline spinning of spider silk". Doğa. 410 (6828): 541–548. Bibcode:2001Natur.410..541V. doi:10.1038/35069000. PMID 11279484. S2CID 205015549.
- ^ "Spider Silk". www.chm.bris.ac.uk. Alındı 18 Ağustos 2010.
- ^ Yang, Y .; Chen, X .; Shao, Z.; Zhou, P .; Porter, D .; Knight, D. P.; Vollrath, F. (2005). "Toughness of Spider Silk at High and Low Temperatures". Gelişmiş Malzemeler. 17: 84–88. doi:10.1002/adma.200400344.
- ^ a b Agnarsson, Ingi; Kuntner, Matjaž; Blackledge, Todd A. (2010). Lalueza-Fox, Carles (ed.). "Bioprospecting Finds the Toughest Biological Material: Extraordinary Silk from a Giant Riverine Orb Spider". PLOS ONE. 5 (9): 11234. Bibcode:2010PLoSO...511234A. doi:10.1371/journal.pone.0011234. PMC 2939878. PMID 20856804.

- ^ a b c Wolff, J. O.; Grawe, I; Wirth, M; Karstedt, A; Gorb, S. N. (2015). "Spider's super-glue: Thread anchors are composite adhesives with synergistic hierarchical organization". Yumuşak Madde. 11 (12): 2394–2403. Bibcode:2015SMat...11.2394W. doi:10.1039/c4sm02130d. PMID 25672841.
- ^ Sahni, V; Harris, J; Blackledge, T. A.; Dhinojwala, A (2012). "Cobweb-weaving spiders produce different attachment discs for locomotion and prey capture". Doğa İletişimi. 3: 1106. Bibcode:2012NatCo...3.1106S. doi:10.1038/ncomms2099. PMID 23033082.
- ^ Cunningham, Aimee (2009). "Taken for a spin: Scientists look to spiders for the goods on silk". Bilim Haberleri. 171 (15): 231–234. doi:10.1002/scin.2007.5591711509.
- ^ Blackledge, TA; Hayashi, CY (2006). "Silken toolkits: Biomechanics of silk fibers spun by the orb web spider Argiope argentata (Fabricius 1775)". Deneysel Biyoloji Dergisi. 209 (Pt 13): 2452–2461. doi:10.1242/jeb.02275. PMID 16788028.
- ^ a b Hinman, M. B. & Lewis, R. V. (1992). "Isolation of a clone encoding a second dragline silk fibroin. Nephila clavipes dragline silk is a two-protein fiber". J. Biol. Kimya. 267 (27): 19320–19324. PMID 1527052.
- ^ a b c d Simmons, A. H.; Michal, C. A. & Jelinski, L. W. (1996). "Molecular orientation and two-component nature of the crystalline fraction of spider dragline silk". Bilim. 271 (5245): 84–87. Bibcode:1996Sci...271...84S. doi:10.1126/science.271.5245.84. PMID 8539605. S2CID 40043335.
- ^ a b van Beek, J. D.; Hess, S.; Vollrath, F. & Meier, B. H. (2002). "The molecular structure of spider dragline silk: Folding and orientation of the protein backbone". Proc. Natl. Acad. Sci. AMERİKA BİRLEŞİK DEVLETLERİ. 99 (16): 10266–10271. Bibcode:2002PNAS...9910266V. doi:10.1073/pnas.152162299. PMC 124902. PMID 12149440.
- ^ Liu, Y .; Sponner, A.; Porter, D .; Vollrath, F. (2008). "Proline and Processing of Spider Silks". Biyomakromoleküller. 9 (1): 116–121. doi:10.1021/bm700877g. PMID 18052126.
- ^ Papadopoulos, P.; Ene, R.; Weidner, I.; Kremer, F. (2009). "Similarities in the Structural Organization of Major and Minor Ampullate Spider Silk". Macromol. Rapid Commun. 30 (9–10): 851–857. doi:10.1002/marc.200900018. PMID 21706668.
- ^ a b Heimer, S. (1988). Wunderbare Welt der Spinnen. Urania. s. 12
- ^ a b Termonia, Y. (1994). "Molecular Modeling of Spider Silk Elasticity". Makro moleküller. 27 (25): 7378–7381. Bibcode:1994MaMol..27.7378T. doi:10.1021/ma00103a018.
- ^ Vollrath, F .; Holtet, T.; Thogersen, H. C. & Frische, S. (1996). "Structural organization of spider silk". Royal Society B Tutanakları. 263 (1367): 147–151. Bibcode:1996RSPSB.263..147V. doi:10.1098/rspb.1996.0023. S2CID 136879037.
- ^ Sponner, A.; Vater, Wolfram, Wolfram; Monajembashi, Shamci, Shamci; Unger, Eberhard, Eberhard; Grosse, Frank, Frank; Weisshart, Klaus, Klaus (2007). Scheibel, Thomas (ed.). "Composition and hierarchical organization of a spider silk". PLOS ONE. 2 (10): e998. Bibcode:2007PLoSO...2..998S. doi:10.1371/journal.pone.0000998. PMC 1994588. PMID 17912375.
- ^ Sapede, D.; Seydel, T.; Forsyth, V. T.; Koza, M. M.; Schweins, R.; Vollrath, F .; Riekel, C. (2005). "Nanofibrillar structure and molecular mobility in spider dragline silk". Makro moleküller. 34 (20): 623. Bibcode:2005MaMol..38.8447S. doi:10.1021/ma0507995.
- ^ Plaza, G.R.; Pérez-Rigueiro, J.; Riekel, C.; Perea, G.B.; Agulló-Rueda, F.; Burghammer, M .; Guinea, G.V.; Elices, M. (2012). "Relationship between microstructure and mechanical properties in spider silk fibers: identification of two regimes in the microstructural changes". Yumuşak Madde. 8 (22): 6015–6026. Bibcode:2012SMat....8.6015P. doi:10.1039/C2SM25446H.
- ^ Zhao, Yue; Hien, Khuat Thi Thu; Mizutani, Goro; Rutt, Harvey N. (June 2017). "Second-order nonlinear optical microscopy of spider silk". Uygulamalı Fizik B. 123 (6): 188. arXiv:1706.03186. Bibcode:2017ApPhB.123..188Z. doi:10.1007/s00340-017-6766-z. S2CID 51684427.
- ^ a b Andersson, M; Johansson, J; Rising, A (2016). "Silk Spinning in Silkworms and Spiders". Uluslararası Moleküler Bilimler Dergisi. 17 (8): 1290. doi:10.3390/ijms17081290. PMC 5000687. PMID 27517908.
- ^ a b Wilson, R. S. (1969). "control of drag-line spinning in certain spiders". Am. Zool. 9: 103–. doi:10.1093/icb/9.1.103.
- ^ Zhao, Yue; Li, Yanrong; Hien, K. T. T.; Mizutani, Goro; Rutt, Harvey N. (2019). "Observation of Spider Silk by Femtosecond Pulse Laser Second Harmonic Generation Microscopy". Sörf. Interface Anal. 51 (1): 50–56. arXiv:1812.10390. doi:10.1002/sia.6545. S2CID 104921418.
- ^ a b Rising, A.; Johansson, J. (2015). "Toward spinning artificial spider silk". Nat. Chem. Biol. 11 (5): 309–315. doi:10.1038/nchembio.1789. PMID 25885958.
- ^ a b c Eisoldt, L.; Thamm, C.; Scheibel, T. (2012). "The role of terminal domains during storage and assembly of spider silk proteins". Biyopolimerler. 97 (6): 355–361. doi:10.1002/bip.22006. PMID 22057429. S2CID 46685716.
- ^ Eisoldt, L.; Smith, A .; Scheibel, T. (2011). "Decoding the secrets of spider silk". Mater. Bugün. 14 (3): 80–86. doi:10.1016/S1369-7021(11)70057-8.
- ^ Tokareva, O.; Jacobsen, M.; Buehler, M .; Wong, J .; Kaplan, D. L. (2014). "Structure–function–property–design interplay in biopolymers: Spider silk". Acta Biomater. 10 (4): 1612–1626. doi:10.1016/j.actbio.2013.08.020. PMC 3926901. PMID 23962644.
- ^ a b Vollrath, F .; Knight, D. P. (2001). "Liquid crystalline spinning of spider silk". Doğa. 410 (6828): 541–548. Bibcode:2001Natur.410..541V. doi:10.1038/35069000. PMID 11279484. S2CID 205015549.
- ^ a b Kluge, J. A.; Rabotyagova, O.; Leisk, G. G.; Kaplan, D. L. (2008). "Spider silks and their applications". Trendler Biotechnol. 26 (5): 244–251. doi:10.1016/j.tibtech.2008.02.006. PMID 18367277.
- ^ Hijirida, D. H.; Do, K. G.; Michal, C.; Wong, S .; Zax, D.; Jelinski, L. W. (1996). "13C NMR of Nephila clavipes major ampullate silk gland". Biophys. J. 71 (6): 3442–3447. Bibcode:1996BpJ....71.3442H. doi:10.1016/S0006-3495(96)79539-5. PMC 1233831. PMID 8968613.
- ^ Lefvre, T.; Boudreault, S.; Cloutier, C.; Pezolet, M. (2008). "Conformational and orientational transformation of silk proteins in the major ampullate gland of Nephila clavipes spiders". Biyomakromoleküller. 9 (9): 2399–2407. doi:10.1021/bm800390j. PMID 18702545.
- ^ Lewis, R. V. (2006). "Spider silk: Ancient ideas for new biomaterials". Chem. Rev. 106 (9): 3762–3774. doi:10.1021/cr010194g. PMID 16967919.
- ^ Andersson, M .; et al. (2014). "Carbonic anhydrase generates CO2 and H+ that drive spider silk formation via opposite effects on the terminal domains". PLOS Biol. 12 (8): e1001921. doi:10.1371/journal.pbio.1001921. PMC 4122339. PMID 25093327.
- ^ Kronqvist, N.; et al. (2014). "Sequential pH-driven dimerization and stabilization of the N-terminal domain enables rapid spider silk formation". Nat. Commun. 5: 3254. Bibcode:2014NatCo...5.3254K. doi:10.1038/ncomms4254. PMID 24510122.
- ^ Knight, D. P.; Vollrath, F. (1999). "Liquid crystals and flow elongation in a spider's silk production line". Proc. R. Soc. B. 266 (1418): 519–523. doi:10.1098/rspb.1999.0667. PMC 1689793.
- ^ Dicko, C.; Porter, D .; Bond, J .; Kenney, J. M. & Vollratht, F. (2008). "Structural disorder in silk proteins reveals the emergence of elastomericity". Biyomakromoleküller. 9 (1): 216–221. doi:10.1021/bm701069y. PMID 18078324.
- ^ Lefèvre, T.; Boudreault, S.; Cloutier, C. & Pézolet, M. (2008). "Conformational and orientational transformation of silk proteins in the major ampullate gland of Nephila clavipes spiders". Biyomakromoleküller. 9 (9): 2399–2407. doi:10.1021/bm800390j. PMID 18702545.
- ^ Heim, M .; Keerl, D. & Scheibel, T. (2009). "Spider Silk: From Soluble Protein to Extraordinary Fiber". Angewandte Chemie Uluslararası Sürümü. 48 (20): 3584–3596. doi:10.1002/anie.200803341. PMID 19212993.
- ^ Heinhorst, S.; Cannon, G. (2002). "Nature: Self-Healing Polymers and Other Improved Materials". J. Chem. Educ. 79 (1): 10. Bibcode:2002JChEd..79...10H. doi:10.1021/ed079p10.
- ^ Knight, D. P.; Vollrath, F. (1 April 2001). "Changes in element composition along the spinning duct in a Nephila spider". Die Naturwissenschaften. 88 (4): 179–182. Bibcode:2001NW.....88..179K. doi:10.1007/s001140100220. ISSN 0028-1042. PMID 11480706. S2CID 26097179.
- ^ a b Vollrath, F. & Knight, D. P. (1998). "Structure and function of the silk production pathway in spider Nephila edulis". Int J Biol Macromol. 24 (2–3): 243–249. doi:10.1016/S0141-8130(98)00095-6. PMID 10342771.
- ^ Wilson, R. S. (1962). "The Control of Dragline Spinning in the Garden Spider". Üç Aylık Mikroskopik Bilim Dergisi. 103: 557–571.
- ^ Magoshi, J.; Magoshi, Y. & Nakamura, S. (1985). "Physical properties and structure of silk: 9. Liquid crystal formation of silk fibroin". Polym. Commun. 26: 60–61.
- ^ Chen, Xin; Knight, David P.; Vollrath, Fritz (1 July 2002). "Rheological characterization of nephila spidroin solution". Biyomakromoleküller. 3 (4): 644–648. doi:10.1021/bm0156126. ISSN 1525-7797. PMID 12099805.
- ^ Jeffery, F; La Mattina, C; Tuton-Blasingame, T; Hsia, Y; Gnesa, E; Zhao, L; Franz, A; Vierra, C (2011). "Microdissection of Black Widow Spider Silk-producing Glands". Görselleştirilmiş Deneyler Dergisi (47): 2382. doi:10.3791/2382. PMC 3341101. PMID 21248709.
- ^ Elices, M.; Plaza, G.R.; Arnedo, M.A .; Perez-Rigueiro, J.; Torres, F.G. & Guinea, G. (2009). "Mechanical Behavior of Silk During the Evolution of Orb-Web Spinning Spiders". Biyomakromoleküller. 10 (7): 1904–1910. doi:10.1021/bm900312c. PMID 19505138.
- ^ Swanson, B. O.; Blackledge, T. A.; Summers, A. P. & Hayashi, C. Y. (2006). "Spider dragline silk: Correlated and mosaic evolution in high-performance biological materials" (PDF). Evrim. 60 (12): 2539–2551. doi:10.1554/06-267.1. PMID 17263115. S2CID 14862626.
- ^ Shao, Z. Z. & Vollrath, F. (2002). "Materials: Surprising strength of silkworm silk". Doğa. 418 (6899): 741. Bibcode:2002Natur.418..741S. doi:10.1038/418741a. PMID 12181556. S2CID 4304912.
- ^ Wen, H. X.; et al. (2010). "Transgenic silkworms (Bombyx mori) produce recombinant spider dragline silk in cocoons". Moleküler Biyoloji Raporları. 37 (4): 1815–1821. doi:10.1007/s11033-009-9615-2. PMID 19633923. S2CID 12924107.
- ^ a b Bowen, C.H. (2018). "Recombinant Spidroins Fully Replicate Primary Mechanical Properties of Natural Spider Silk]". Biyomakromoleküller. 19 (9): 3853–3860. doi:10.1021/acs.biomac.8b00980. hdl:2060/20180007385. PMID 30080972.
- ^ Elices, M.; Guinea, G. V.; Plaza, G. R.; Karatzas, C.; Riekel, C.; Agulló-Rueda, F.; Daza, R .; Pérez-Rigueiro, J. (2011). "Bioinspired Fibers Follow the Track of Natural Spider Silk". Makro moleküller. 44 (5): 1166–1176. Bibcode:2011MaMol..44.1166E. doi:10.1021/ma102291m.
- ^ US patent 2008109923, Lewis, R. V., "Expression of spider silk proteins", published 2010-05-25, assigned to University of Wyoming
- ^ Scheller, J. & Conrad, U. (2005). "Plant-based material, protein and biodegradable plastic". Bitki Biyolojisinde Güncel Görüş. 8 (2): 188–196. doi:10.1016/j.pbi.2005.01.010. PMID 15753000.
- ^ a b Lazaris, A.; Arcidiacono, S, S; Huang, Y, Y; Zhou, J. F., JF; Duguay, F, F; Chretien, N, N; Welsh, E. A., EA; Soares, J. W., JW; Karatzas, C. N., CN (2002). "Spider silk fibers spun from soluble recombinant silk produced in mammalian cells". Bilim. 295 (5554): 472–476. Bibcode:2002Sci...295..472L. doi:10.1126/science.1065780. PMID 11799236. S2CID 9260156.
- ^ a b Seidel, A.; Liivak, Oskar; Calve, Sarah; Adaska, Jason; Ji, Gending; Yang, Zhitong; Grubb, David; Zax, David B.; Jelinski, Lynn W. (2000). "Regenerated spider silk: Processing, properties, and structure". Makro moleküller. 33 (3): 775–780. Bibcode:2000MaMol..33..775S. doi:10.1021/ma990893j.
- ^ Arcidiacono, S.; Mello, Charlene M.; Butler, Michelle; Welsh, Elizabeth; Soares, Jason W.; Allen, Alfred; Ziegler, David; Laue, Thomas; Chase, Susan (2002). "Aqueous processing and fiber spinning of recombinant spider silks". Makro moleküller. 35 (4): 1262–1266. Bibcode:2002MaMol..35.1262A. doi:10.1021/ma011471o.
- ^ Xia, X. X.; et al. (2010). "Native-sized recombinant spider silk protein produced in metabolically engineered Escherichia coli results in a strong fiber". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 107 (32): 14, 059–14, 063. Bibcode:2010PNAS..10714059X. doi:10.1073/pnas.1003366107. PMC 2922564. PMID 20660779.
- ^ Kinahan, M. E.; et al. (2011). "Tunable Silk: Using Microfluidics to Fabricate Silk Fibers with Controllable Properties". Biyomakromoleküller. 12 (5): 1504–1511. doi:10.1021/bm1014624. PMC 3305786. PMID 21438624.
- ^ Rammensee, S.; Slotta, U.; Scheibel, T. & Bausch, A. R. (2008). "Assembly mechanism of recombinant spider silk proteins (microfluidic)". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 105 (18): 6590–6595. Bibcode:2008PNAS..105.6590R. doi:10.1073/pnas.0709246105. PMC 2373321. PMID 18445655.
- ^ Spintec Engineering GmbH (Almanca'da)
- ^ Eisoldt, L.; Smith, A. & Scheibel, T. (2011). "Decoding the secrets of spider silk". Mater. Bugün. 14 (3): 80–86. doi:10.1016/s1369-7021(11)70057-8.
- ^ a b Gustafsson, L.; Jansson, R.; Hedhammar, M. & van der Wijngaart, W. (2018). "Structuring of Functional Spider Silk Wires, Coatings, and Sheets by Self-Assembly on Superhydrophobic Pillar Surfaces". Adv. Mater. 30 (3): 1704325. doi:10.1002/adma.201704325. PMID 29205540.
- ^ Gustafsson, Linnea; Panagiotis Tasiopoulos, Christos; Jansson, Ronnie; Kvick, Mathias; Duursma, Thijs; Gasser, Thomas Christian; van der Wijngaart, Wouter; Hedhammar, My (16 August 2020). "Recombinant Spider Silk Forms Tough and Elastic Nanomembranes that are Protein‐Permeable and Support Cell Attachment and Growth". Gelişmiş Fonksiyonel Malzemeler. 30 (40): 2002982. doi:10.1002/adfm.202002982.
- ^ Fischer, F. & Brander, J. (1960). "Eine Analyse der Gespinste der Kreuzspinne". Hoppe-Seyler'in Zeitschrift für Physiologische Chemie. 320: 92–102. doi:10.1515/bchm2.1960.320.1.92. PMID 13699837.
- ^ Lucas, F.; Shaw, J. T. B. & Smith, S. G. (1960). "The Composition of Arthropod Silk Fibrons". Insect Chemistry. Symp. 3: 208–214.
- ^ Lucas, F.; Shaw, J. T. B. & Smith, S. G. (1960). "Comparative studies of fibroins.I. The amino acid composition of various fibroins and its significance in relation to their crystal structure and taxonomy". Moleküler Biyoloji Dergisi. 2 (6): 339–349. doi:10.1016/S0022-2836(60)80045-9. PMID 13763962.
- ^ Xu, M. & Lewis, R. V. (1990). "Structure of a Protein Superfiber - Spider Dragline Silk". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 87 (18): 7120–7124. Bibcode:1990PNAS...87.7120X. doi:10.1073/pnas.87.18.7120. PMC 54695. PMID 2402494.
- ^ Lucas, F. (1964). "Spiders and their silks". Keşif. 25: 20–26.
- ^ Vollrath, F. & Edmonds, D. T. (1989). "Modulation Of The Mechanical-Properties Of Spider Silk By Coating With Water". Doğa. 340 (6231): 305–307. Bibcode:1989Natur.340..305V. doi:10.1038/340305a0. S2CID 4355740.
- ^ Vollrath, F .; Madsen, B. & Shao, Z. Z. (2001). "The effect of spinning conditions on the mechanics of a spider's dragline silk". Royal Society B Tutanakları. 268 (1483): 2339–2346. doi:10.1098/rspb.2001.1590. PMC 1088885. PMID 11703874.
- ^ Simmons, A.; Ray, E. & Jelinski, L. W. (1994). "Solid-State C-13 NMR of Nephila-Clavipes Dragline Silk Establishes Structure and Identity of Crystalline Regions". Makro moleküller. 27 (18): 5235–5237. Bibcode:1994MaMol..27.5235S. doi:10.1021/ma00096a060.
- ^ Shao, Z.; Vollrath, F .; Sirichaisit, J. & Young, R. J. (1999). "Analysis of spider silk in native and supercontracted states using Raman spectroscopy". Polimer. 40 (10): 2493–2500. doi:10.1016/S0032-3861(98)00475-3.
- ^ Riekel, C.; Bränden, C; Craig, C; Ferrero, C; Heidelbach, F; Müller, M (1999). "Aspects of X-ray diffraction on single spider fibers". Int. J. Biol. Macromol. 24 (2–3): 179–186. doi:10.1016/S0141-8130(98)00084-1. PMID 10342763.
- ^ Knight, D. P.; Knight, M. M. & Vollrath, F. (2000). "Beta transition and stress-induced phase separation in the spinning of spider dragline silk". Int. J. Biol. Macromol. 27 (3): 205–210. doi:10.1016/S0141-8130(00)00124-0. PMID 10828366.
- ^ Riekel, C. & Vollrath, F. (2001). "Spider silk fibre extrusion: combined wide- and small-angle X- ray microdiffraction experiments". Int. J. Biol. Macromol. 29 (3): 203–210. doi:10.1016/S0141-8130(01)00166-0. PMID 11589973.
- ^ Gosline, J. M.; DeMont, M. E. & Denny, M. W. (1986). "The structure and properties of spider silk". Gayret. 10: 37–43. doi:10.1016/0160-9327(86)90049-9.
- ^ Vollrath, F. & Porter, D. (2006). "Spider silk as an archetypal protein elastomer". Yumuşak Madde. 2 (5): 377–385. Bibcode:2006SMat....2..377V. doi:10.1039/b600098n. PMID 32680251. S2CID 97234857.
- ^ Kerkam, K.; Viney, C.; Kaplan, D. & Lombardi, S. (1991). "Liquid Crystallinity of Natural Silk Secretions". Doğa. 349 (6310): 596–598. Bibcode:1991Natur.349..596K. doi:10.1038/349596a0. S2CID 4348041.
- ^ Knight, D. P. & Vollrath, F. (1999). "Liquid crystals and flow elongation in a spider's silk production line". Royal Society B Tutanakları. 266 (1418): 519–523. doi:10.1098/rspb.1999.0667. PMC 1689793.
- ^ Prince, J. T.; McGrath, K. P.; Digirolamo, C. M. & Kaplan, D. L. (1995). "Construction, Cloning, and Expression of Synthetic Genes Encoding Spider Dragline Silk". Biyokimya. 34 (34): 10879–10885. doi:10.1021/bi00034a022. PMID 7662669.
- ^ Arcidiacono, S.; Mello, C.; Kaplan, D.; Cheley, S. & Bayley, H. (1998). "Purification and characterization of recombinant spider silk expressed in Escherichia coli". Uygulamalı Mikrobiyoloji ve Biyoteknoloji. 49 (1): 31–38. doi:10.1007/s002530051133. PMID 9487707. S2CID 35267049.
- ^ Seidel, A.; Liivak, O. & Jelinski, L. W. (1998). "Artificial Spinning of Spider Silk". Makro moleküller. 31 (19): 6733–6736. Bibcode:1998MaMol..31.6733S. doi:10.1021/ma9808880.
- ^ Maev Kennedy (24 January 2012). "Spider silk cape goes on show at V&A". gardiyan.
- ^ Heimer, S. (1988). Wunderbare Welt der Spinnen. Urania. s. 14
- ^ Jackson, Robert R. (1974). "Effects of D-Amphetamine Sulfate and Diazepam on Thread Connection Fine Structure in a Spider's Web". Arachnology Dergisi. 2 (1): 37–41. JSTOR 3704994.
- ^ [http://www.vam.ac.uk/content/articles/g/golden-spider-silk/ Golden spider silk] Victoria & Albert Museum
- ^ Leggett, Hadley (23 September 2009). "1 Million Spiders Make Golden Silk for Rare Cloth". Kablolu.
- ^ Allmeling, Christina; Jokuszies, Andreas; Reimers, Kerstin; Kall, Susanne; Vogt, Peter M. (2006). "Use of spider silk fibres as an innovative material in a biocompatible artificial nerve conduit". Hücresel ve Moleküler Tıp Dergisi. 10 (3): 770–777. doi:10.1111/j.1582-4934.2006.tb00436.x. PMC 3933158. PMID 16989736.
- ^ Berenbaum, Mayıs R., Field Notes – Spin ControlThe Sciences, The New York Academy of Sciences, Eylül / Ekim 1995
- ^ Example of use of spider silk for telescopic rifle sights. Bonnier Corporation. 1955. Alındı 24 Ağustos 2011.
- ^ Duarte F. J.; Taylor, T S; Siyah, A M; Davenport, Batı E; Varmette, P G (2011). "Güvenli serbest alan optik iletişimleri için N-yarıklı interferometre: 527 m intra interferometrik yol uzunluğu". Optik Dergisi. 13 (3): 5710. Bibcode:2011JOpt...13c5710D. doi:10.1088/2040-8978/13/3/035710.
- ^ Osaki, Shigeyoshi (2012). "Spider Silk Violin Strings with a Unique Packing Structure Generate a Soft and Profound Timbre". Fiziksel İnceleme Mektupları. 108 (15): 154301. Bibcode:2012PhRvL.108o4301O. doi:10.1103/PhysRevLett.108.154301. PMID 22587257.
- ^ a b c d Service, Robert F. (18 October 2017). "Spinning spider silk into startup gold". Science Magazine, American Association for the Advancement of Science. Alındı 26 Kasım 2017.
- ^ http://www.lle.rochester.edu/media/publications/documents/theses/Bonino.pdf
- ^ Xia, Xiao-Xia; Qian, Zhi-Gang; Ki, Chang Seok; Park, Young Hwan; Kaplan, David L .; Lee, Sang Yup (2010). "Native-sized recombinant spider silk protein produced in metabolically engineered Escherichia coli results in a strong fiber". Ulusal Bilimler Akademisi Bildiriler Kitabı. 107 (32): 14059–14063. Bibcode:2010PNAS..10714059X. doi:10.1073/pnas.1003366107. JSTOR 25708855. PMC 2922564. PMID 20660779.
- ^ "Draadkracht: spindoctors maken supersterk nepweb" [Wire strength: spin doctors make super strong fake cobweb] (in Dutch). KIJK. 21 Nisan 2012. Alındı 15 Ekim 2014.
- ^ https://boltthreads.com/technology/microsilk
- ^ https://boltthreads.com/technology/silk-protein
- ^ "University of Notre Dame and Kraig Biocraft Laboratories Create Artificial Spider Silk Breakthrough" (Basın bülteni). Kraig Biocraft Laboratories. 29 Eylül 2010. Alındı 3 Ocak 2012.
- ^ "Fraser Research Publicly Announced at Press Conference" (Basın bülteni). Notre Dame Üniversitesi. 1 Ekim 2010. Arşivlenen orijinal 10 Ekim 2010'da. Alındı 3 Ocak 2012.
- ^ Kluge, Jonathan A.; Rabotyagova, Olena; Leisk, Gary G.; Kaplan, David L. (May 2008). "Spider silks and their applications". Biyoteknolojideki Eğilimler. 26 (5): 244–251. doi:10.1016/j.tibtech.2008.02.006. PMID 18367277.
- ^ Scheibel, Thomas (November 2004). "Spider silks: recombinant synthesis, assembly, spinning, and engineering of synthetic proteins". Mikrobiyal Hücre Fabrikaları. 3 (1): 14. doi:10.1186/1475-2859-3-14. PMC 534800. PMID 15546497.
- ^ https://www.goldwin-sports.com/us/feature/goldwinskijacket/
- ^ https://qz.com/708298/synthetic-spider-silk-could-be-the-biggest-technological-advance-in-clothing-since-nylon/
Dış bağlantılar
- "The Silk Spinners", a BBC program about silk-producing animals
- Meadows, Robin (5 August 2014). "How Spiders Spin Silk". PLOS Biyoloji. 12 (8): e1001922. doi:10.1371/journal.pbio.1001922. PMC 4122354. PMID 25093404.
- Rejcek, Peter (11 April 2019). "The Tangled Web of Turning Spider Silk Into a Super Material". Tekillik Merkezi. Alındı 24 Nisan 2019.
- Victoria and Albert Museum (29 July 2019). "How was it made? Golden spider silk". Youtube. Alındı 8 Ağustos 2020.
| Araknoloji |  | |
|---|---|---|
| Taksonomi |
| |
| Anatomi | ||
| İnsan etkileşimi | ||
| Web'ler | ||
| ||