Camarasaurus - Camarasaurus
| Camarasaurus | |
|---|---|
 | |
| Atlı iskelet döküm Asturias Jurassic Müzesi | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Alttakım: | †Sauropodomorpha |
| Clade: | †Sauropoda |
| Clade: | †Camarasauromorpha |
| Clade: | †Camarasauridae |
| Alt aile: | †Camarasaurinae Cope, 1878 |
| Cins: | †Camarasaurus Cope, 1877a |
| Türler | |
| †Camarasaurus supremus Cope, 1877a | |
| Türler | |
| |
| Eş anlamlı | |
| |
Camarasaurus (/ˌkæmərəˈsɔːrəs/ KAM-ər-ə-SAWR-əs ) bir cins nın-nin dört ayaklı, otçul dinozorlar. Devlerin en yaygın olanıydı Sauropodlar içinde bulunmak Kuzey Amerika. Fosil kalıntıları, Morrison Formasyonu nın-nin Colorado ve Utah ile çıkmak Geç Jura epoch (Kimmeridciyen -e Tithoniyen aşamalar), 155 ila 145 milyon yıl önce.
Camarasaurus Künt bir burun ve oldukça kare olan kemerli bir kafatasının ayırt edici bir kafatası profilini sundu. Muhtemelen sürülerde veya en azından aile gruplarında seyahat etti.
Adı, içindeki oyuk odalara atıfta bulunan "odacıklı kertenkele" anlamına gelir. omur (Yunan καμαρα (Kamara) "tonozlu oda" anlamına gelen veya kemerli kapaklı herhangi bir şey ve σαυρος (Sauros) "kertenkele" anlamına gelir).
Açıklama
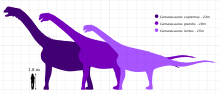
Camarasaurus ortaya çıkarılan en yaygın ve sıklıkla iyi korunmuş sauropod dinozorlarından biridir. En yaygın türlerin maksimum boyutu, C. lentus, yaklaşık 15 m (49 ft) uzunluğundaydı. En büyük türler, C. supremus, maksimum 23 m (75 ft) uzunluğa ve tahmini maksimum 47 ağırlığa ulaştıton (51.8 ton ).[1][2]
Kemerli kafatası nın-nin Camarasaurus dikkat çekici bir şekilde kare idi ve keskin olmayan burun birçok Fenestrae sağlam olmasına ve paleontologlar tarafından sık sık iyi durumda bulunmasına rağmen. 19 cm uzunluğunda (7,5 inç) diş şeklindeydi keskiler (spatula) ve çene boyunca eşit olarak düzenlenmiştir. Dişlerin gücü şunu gösterir: Camarasaurus Muhtemelen ince dişliye göre daha iri bitki materyali yedi diplodositler.[kaynak belirtilmeli ]
Her ön ekstremite beş parmağa sahipti ve iç ayak parmağı büyük, keskin pençe. Çoğu sauropod gibi, ön uzuvlar arka ayaklardan daha kısaydı, ancak yüksek pozisyonu omuzlar arkada küçük bir eğim olduğu anlamına geliyordu.
Diğer sauropodlarda görüldüğü gibi ağırlık tasarrufu amacına hizmet eden omurların birçoğu oyuk veya "pnömatik" idi; yani omurlar, akciğerlere bağlı karmaşık bir hava kesesi sistemi için geçitler ve boşluklarla dolu. Bu özellik o zamanlar çok az anlaşılmıştı Camarasaurus keşfedildi, ancak yapısı yaratığın "odacıklı kertenkele" anlamına gelen ismine ilham kaynağı oldu. Boyun ve dengeleme kuyruğu, bu büyüklükteki bir sauropod için normalden daha kısaydı. Camarasaurus, diğer bazı sauropodlar gibi, omurilik kalçalara yakın. Paleontologlar[DSÖ? ] başlangıçta bunun bir saniye olduğuna inanıyordu beyin, belki de böylesine büyük bir yaratığı koordine etmek için gerekli. Gerçekte, yoğun bir sinir sistemi -muhtemelen refleks veya otomatik- faaliyet alanı olsa da, bir beyin değildi; bu tür genişlemeler, omurgalı hayvanlarda bir dereceye kadar sıklıkla bulunur.[kaynak belirtilmeli ]
Camarasaurus Grandis diğer sauropodlardan daha sağlam bir yarıçapa sahipti Venenosaurus.[3]
Bir örnek Camarasaurus SMA 0002 olarak adlandırılır (aynı zamanda CathetosaurusWyoming'in Howe-Stephens Quarry'den "E.T." olarak anılan), yumuşak doku kanıtı gösteriyor. Çene hattı boyunca, hayvanın diş etleri gibi görünen kemikleşmiş kalıntıları geri kazanıldı, bu da diş etleriyle kaplı derin dişlere sahip olduğunu ve sadece kronların uçlarının çıktığını gösteriyor. Dişler, öldükten sonra, diş etleri çekilirken, kururken ve çürümeyle sıkılırken yuvalarından daha da dışarı itildi. Numunenin muayeneleri, kesin olarak bilinmemekle birlikte, dişlerin sert dış pullarla ve muhtemelen bir çeşit gagayla kaplandığını da göstermektedir.[4]
Keşif tarihi


İlk kaydı Camarasaurus 1877'den gelir, birkaçının dağılması omur bulundu Colorado, yazan Oramel W. Lucas. Uzun süredir devam eden ve sert rekabetini sürdürmek ( Kemik Savaşları ) ile Othniel Charles Marsh, paleontolog Edward Drinker Cope kemikler için para ödedi ve hızlı hareket etti, aynı yıl isimlerini verdi. Marsh daha sonra sauropod bulgularından bazılarını adlandırdı. Morosaurus grandis, ancak bugün çoğu paleontolog onları bir tür Camarasaurus Re: "Morosaurus" agilis. Bu tür adlandırma çatışmaları, iki rakip dinozor avcısı arasında yaygındı.
Tam bir iskelet Camarasaurus tarafından tarif edilmedi Charles W. Gilmore, 1925'e kadar. Çünkü bir gençten kalma iskelet CamarasaurusBununla birlikte, zamanın birçok örneği dinozorun şu anda bilindiğinden çok daha küçük olduğunu gösteriyor.
Türler

türler nın-nin Camarasaurus Cope'nin orijinal türü, C. supremus ("en büyük odacıklı kertenkele" anlamına gelir) adı 1877'de verilmiştir. Keşfedildiğinden beri diğer türler arasında C. grandis ("büyük odacıklı kertenkele") 1877'de, C. lentus 1889'da ve C. lewisi (başlangıçta yeni bir cins olarak tanımlanmıştır Cathetosaurus1988'de.
C. supremus, adından da anlaşılacağı gibi, bilinen en büyük türdür. Camarasaurus ve Jurassic'in son döneminden bilinen en büyük sauropodlardan biri Morrison Formasyonu. Büyük boyutu dışında, neredeyse ayırt edilemezdi C. lentus. C. supremus bir bütün olarak cinsin tipik bir örneği değildi ve yalnızca oluşumun en son, en üst kısımlarından biliniyordu. Her ikisi de C. grandis ve C. lentus daha küçüktü ve Morrison'un ilk aşamalarında meydana geldi.
Stratigrafik kanıtlar, kronolojik dizinin üç tür arasındaki fiziksel farklılıklarla uyumlu olduğunu ve Morrison Formasyonu içinde evrimsel bir ilerlemeyi tanımladığını göstermektedir. C. grandis en eski türdür ve Morrison'un en alt kaya katmanlarında meydana gelmiştir. C. lentus daha sonra ortaya çıktı, birlikte varoldu C. grandis Muhtemelen iki türün omurga anatomisindeki farklılıkların önerdiği gibi farklı ekolojik nişler nedeniyle birkaç milyon yıldır. Daha sonraki bir aşamada, C. grandis rock rekorundan kayboldu, geriye sadece C. lentus. Sonra C. lentusda kayboldu; aynı zamanda, C. supremus en üst katmanlarda ortaya çıktı. Türlerin bu ani ardıllığı ve ikisi arasındaki çok yakın benzerlik şunu gösteriyor: C. supremus doğrudan doğruya evrimleşmiş olabilir C. lentus, daha büyük, daha sonra hayatta kalan hayvan popülasyonunu temsil eder.[5]
C. lewisi türler başlangıçta olarak adlandırıldı Cathetosaurus Lewis ve daha sonra eşanlamlı Camarasaurus. Bir 2013 analizi, iki cinsi tekrar böldü.[6]
Sınıflandırma

bilimsel sınıflandırma nın-nin Camarasaurus, kullanmak Linnaean sistemi, sağ üstteki kutuda verilmiştir, ancak paleontologlar arasında, dinozorların bu taksonomik sınıflandırma yöntemi, kladistik -den ilham alan filogenetik taksonomi.
D'Emic (2012) sonrası Macronaria'nın basitleştirilmiş bir cladogramı aşağıda gösterilmiştir:[7]
| Macronaria |
| |||||||||||||||||||||||||||
Camarasaurus olarak kabul edilir baz alınan macronaryan, tüm makronaryanların ortak atasıyla daha yakından ilgili türetilmiş gibi formlar Brachiosaurus.
Paleobiyoloji

Herding
Bir fosil kaydı iki yetişkin ve yaklaşık 150 milyon yıl önce (kuzeydoğuda) Geç Jura döneminde birlikte ölen 12,2 m uzunluğunda (40 ft) bir genç Wyoming, Amerika Birleşik Devletleri).[not 1] Vücutlarının, alüvyon çamurundaki son dinlenme yerlerine kadar akan (sel) bir nehir tarafından yıkandığı varsayıldı. Senaryo şunu öneriyor: Camarasaurus seyahat etti sürüler veya en azından aile gruplarında. Diğer sitelerde fosil camarasaur yumurtalar Düzgün bir şekilde düzenlenmiş değil, çizgiler halinde bulunmuş yuvalar diğer bazı dinozorlarda olduğu gibi, bu da çoğu sauropod gibi Camarasaurus gençliğine bakmadı.
Besleme
Daha önce, bilim adamları şunu önerdiler: Camarasaurus ve diğer sauropodlar yutmuş olabilir mide taşı (taşlar) midede yiyeceğin öğütülmesine, çok pürüzsüz hale geldiklerinde kusmalarına veya geçmelerine yardımcı olmak için. Bununla birlikte, mide taşlarına ilişkin kanıtların daha yeni analizi, durumun böyle olmadığını gösteriyor. Güçlü, sağlam dişleri Camarasaurus sauropodların çoğundan daha gelişmiştir ve ortalama olarak her 62 günde bir değiştirilmiştir (M. D'Emic ve ark.). Camarasaurus olabilir çiğnenmiş yutmadan önce bir dereceye kadar ağzına yiyecek.[8] Diğer bulgular gösteriyor ki Camarasaurus spp. diğer sauropodlardan farklı olarak tercih edilen bitki örtüsü, rekabet etmeden aynı ortamı paylaşmalarına izin verir.[9]
Büyüme
Uzun kemik histoloji araştırmacıların belirli bir bireyin ulaştığı yaşı tahmin etmesini sağlar. Griebeler ve arkadaşları tarafından yapılan bir çalışma. (2013) uzun kemik histolojik verilerini inceledi ve şu sonuca varmıştır: Camarasaurus sp. CM 36664 14.247 kilogram (15.7 kısa ton), 20 yaşında cinsel olgunluğa ulaştı ve 26 yaşında öldü.[10]
Metabolizma
Kartal et al. gerçekleştirilen kümelenmiş izotop termometresi çeşitli Jurassic sauropodların dişlerini kaplayan minede Camarasaurus. 32.4–36.9 ° C (90.3–98.4 ° F) sıcaklıklar elde edildi, bu da modern memelilerinkiyle karşılaştırılabilir.[11]
Paleopatoloji

Bir Camarasaurus pelvis iyileşti Dinozor Ulusal Anıtı Utah'da, Allosaurus.[12]
1992'de kısmi C. grandis iskelet keşfedildi Bryan Küçük Stegosaurus Ocağı Morrison Formasyonunun Canon City, Colorado.[13] Bu örnek, DMNH 2908 olarak kataloglanan kısmi bir sağ humerusu ve arka ve kuyruktan ilişkili omurları korudu.[13] 2001'de Lorie McWhinney, Kenneth Carpenter ve Bruce Rothschild humerusta gözlemlenen bir patolojinin tanımını yayınladı.[14] Not ettiler jukstakortikal lezyon 25 x 18 cm genişliğinde, dokuma liflere benzeyen kemikten yapılmıştır.[15] olmasına rağmen dokuma kemik aksesuar diş kemiğinde oluşur, diğer yerlerde yaralanma veya hastalık belirtisidir.[15] Dokuma kemiğin "dalgalı lifli demetleri", m. Brachialis.[15] Lezyon füzyon ve eksikliği gözeneklilik yakın ve uzak uçlarında periostitisin inaktif olduğunu veya iyileştiğini gösterir.[15] McWhinney ve diğer araştırmacılar, bu yaralanmanın hayvan için sürekli bir zorluk kaynağı olacağını savundu.[16] Kaslara baskı uygulardı.[14] Bu basınç, kasların kan damarlarını ve sinirlerini sıkıştırarak her iki uzvun hareket aralığını azaltır. fleksör ve ekstansör kaslar.[14] Bu etki, mM. Brachialis, m. Brachoradialis ve daha az ölçüde m. biceps brachii lezyonun humerus üzerindeki konumuna.[14] Araştırmacılar, kasların iltihaplanmasının ve periost ön ekstremitenin alt bölgesinde de ek komplikasyonlara neden olabilirdi.[17] Lezyon ayrıca uzun vadede fasiit ve miyozist.[14] Bu patolojik süreçlerin kümülatif etkisi, uzvun hareket etme yeteneği üzerinde orta ila şiddetli etkilere sahip olacak ve "yiyecek aramak ve avcılardan kaçmak gibi günlük aktiviteleri gerçekleştirmeyi zorlaştırdı".[16]
Patolojinin nedenini belirlemeye yardımcı olmak için McWhinney ve diğer araştırmacılar, CT tarama 3 mm'lik artışlarla.[18] CT taraması, kütlenin tutarlı bir radyodansite ve kemik korteksinden bir radyoaktif hat.[19] Stres kırığı veya osteomiyelit veya enfeksiyöz periostitis gibi bulaşıcı süreçlere dair hiçbir kanıt bulunamadı.[18] Ayrıca dışladılar osteokondrom mahmuzun ekseni humerusun dikey eksenine göre 25 ° olduğundan, humerus eksenine 90 ° 'de bir osteokondrom oluşmuş olacaktır.[15] Bilim adamları tarafından mahmuz taşıyan lezyonun kökeni için belirlenen diğer adaylar şunları içeriyordu:
- Hipertrofik osteoartropati - bu mahmuz benzeri sürecin varlığıyla dışlanmış olsa da[20]
- Osteoid osteoma - ama bu mahmuzu açıklamaz veya osteoblastik reaksiyon[21]
- Shin atelleri veya tibial stres sendromu - birçok semptom ortak tutulacağı için olası bir köken, ancak incik atelleri çıkıntıyı açıklamayacaktır.[22]
- Miyozit ossificans traumatica (sirkumscripta) - Mümkün, ancak olası olmayan kaynak.[23]
- Avülsiyon yaralanması - McWhinney ve diğer araştırmacılar, "kasların tekrarlayan aşırı yorulmasının" neden olduğu bir avülsiyon hasarının humerustaki lezyon için en olası kaynak olduğunu düşündüler.[14] Araştırmacılar, lezyonun avülsiyondan kaynaklandığına inanıyorlardı. m. Brachialis "aşağı doğru eğimli bir eliptik kütle" oluşumuna neden olur.[16] Kemik çıkıntısına, tabanında bir yırtılmayı takiben osteoblastik bir tepki neden olmuştur. m. Brachioradialis fleksör hareketinden kaynaklanır.[16]
Paleoekoloji
Yetişme ortamı

Morrison Formasyonu, nehrin doğu kanadı boyunca yer almaktadır. kayalık Dağlar, fosil açısından zengin bir bölgeye ev sahipliği yapmaktadır. Geç Jura Kaya. Akrabaları da dahil olmak üzere çok sayıda dinozor türü burada bulunabilir. Camarasaurus gibi Diplodocus, Apatozorlar, ve Brachiosaurus ancak camarasaurlar, oluşumdaki dinozorların en bol olanlarıdır.[24] Bir dizi tam iskelet bulunmuştur. Colorado, Yeni Meksika, Utah, ve Wyoming genellikle 2-6 stratigrafik bölgelerde bulunur.[25] Göre radyometrik tarihleme, Morrison tortul katmanları tabanda 156,3 milyon yıl önce (Mya), en üstte 146,8 Mya arasında değişiyor ve bu da onu geç yerleştiriyor. Oksfordiyen, Kimmeridciyen ve erken Tithoniyen aşamalar Geç Jura dönemine ait.[26][27] Çevresi olarak yorumlanır yarı kurak farklı ile ıslak ve kuru mevsimler.
Dinozor ve fosillerin izini sürmek Özellikle New Mexico'dan Alberta ve Saskatchewan'a kadar uzanan ve Rocky Dağları'nın Ön Sırası'nın öncüleri batıya doğru ilerlemeye başladığında oluşan Morrison Havzasında bulunur. Doğu cephelerinden aşınmış malzeme drenaj havzaları akarsular tarafından taşındı ve nehirler ve yatırıldı bataklık ovalar, göller, nehir kanalları ve taşkın yatakları.[28] Oluşum yaş olarak benzerdir Solnhofen Kireçtaşı Almanya'da oluşumu ve Tendaguru Oluşumu içinde Tanzanya. 1877'de, Kemik Savaşları, erken dönem paleontologlar arasında fosil toplama rekabeti Othniel Charles Marsh ve Edward Drinker Cope.
Paleofauna

Morrison Formasyonu, aşağıdakiler gibi devasa sauropod dinozorların hakim olduğu bir ortamı ve zamanı kaydeder. Barosaurus, Diplodocus, Apatozorlar, Brontosaurus, ve Brachiosaurus. Yan yana yaşayan dinozorlar Camarasaurus otçullar dahil ornithischians Camptosaurus, Gargoyleosaurus, Dryosaurus, Stegosaurus, ve Othnielosaurus. Bu paleo ortamdaki yırtıcılar, theropodlar Saurophaganax, Torvosaurus, Ceratosaurus, Marshosaurus, Stokesosaurus, Ornitoestler,[29] ve Allosaurus theropod örneklerinin% 75'ini oluşturan ve en üst sırada yer alan tropik seviye Morrison'ın besin ağı.[30][31] Camarasaurus yaygın olarak aynı sitelerde bulunur Allosaurus, Apatozorlar, Stegosaurus, ve Diplodocus.[32]
Bu bölgedeki diğer organizmalar dahil çift kabuklular, Salyangozlar, ışın yüzgeçli balıklar, kurbağalar, semenderler, kaplumbağalar, sfenodontlar, kertenkele karasal ve suda yaşayan Crocodylomorphans ve birkaç tür pterozorlar gibi Harpactognathus ve Mesadactylus. Mevcut erken memeliler docodonts (gibi Docodon ), çok tüberküloz simetrodontlar ve Triconodonts. Dönemin bitki örtüsü fosilleri ile ortaya çıkarılmıştır. yeşil alg, mantarlar, yosunlar, at kuyruğu, sikadlar, ginkgoes ve birkaç aile iğne yapraklılar. Bitki örtüsü, nehir kenarındaki ormanlardan farklıdır. ağaç eğrelti otları, ve eğrelti otları (Galeri ormanları ), eğrelti otuna savanalar gibi ara sıra ağaçlarla Araucaria kozalaklı ağaç Brachyphyllum.[33]
Notlar
- ^ Omurgalı Paleontolojisi Bölümü tarafından kazılmıştır. Kansas Üniversitesi Doğa Tarihi Müzesi ve Biyoçeşitlilik Merkezi, 1997 ve 1998 tarla sezonlarında.
Referanslar
- ^ Benton, Michael J. (2012). Tarihöncesi Yaşam. Edinburgh, İskoçya: Dorling Kindersley. s. 270–271. ISBN 978-0-7566-9910-9.
- ^ Foster, John (2007). Jurassic West: Morrison Formasyonunun Dinozorları ve Dünyaları. Indiana University Press. s. 201, 248. ISBN 978-0-253-34870-8.
- ^ "Önelimb", Tidwell, Carpenter ve Meyer (2001). Sayfa 148.
- ^ "Ağızda Camarasaurus'a Bakmak".
- ^ "Camarasaurus grandis, "Foster (2007). Sayfa 204.
- ^ Mateus, O. ve Tschopp E. (2013). Cathetosaurus geçerli bir sauropod cinsi olarak ve ile karşılaştırmalar Camarasaurus. Omurgalı Paleontoloji Dergisi, Program ve Özetler, 2013. 173.
- ^ D'Emic, M.D. (2012). "Titanosauriform sauropod dinozorlarının erken evrimi" (PDF). Linnean Society'nin Zooloji Dergisi. 166 (3): 624–671. doi:10.1111 / j.1096-3642.2012.00853.x.
- ^ Kanatlar ve Sander (2006).
- ^ "En büyük dinozorlardaki yüksek diş değiştirme oranları, evrimsel başarılarına katkıda bulundu".
- ^ Griebeler, EM; Klein, N; Sander, PM (2013). "Uzun Kemik Histolojik Verileri Kullanılarak Büyüme Eğrilerinden Çıkarılan Sauropodomorf Dinozorların Yaşlanması, Olgunlaşması ve Büyümesi: Metodolojik Kısıtlamalar ve Çözümlerin Değerlendirilmesi". PLOS ONE. 8 (6): e67012. Bibcode:2013PLoSO ... 867012G. doi:10.1371 / journal.pone.0067012. PMC 3686781. PMID 23840575.
- ^ Eagle, R.A .; Tütken, T .; Martin, T.S .; Tripati, A.K .; Fricke, H.C .; Connely, M .; Cifelli, R.L .; Eiler, J.M. (2011-07-22). "Fosil Biyominerallerde İzotopik (13C-18O) Sıralamasından Belirlenen Dinozor Vücut Sıcaklıkları". Bilim. 333 (6041): 443–445. Bibcode:2011Sci ... 333..443E. doi:10.1126 / science.1206196. PMID 21700837. S2CID 206534244.
- ^ "Camarasaurus", Dodson, vd. 56.Sayfa
- ^ a b "Giriş", McWhinney, Carpenter ve Rothschild (2001); sayfa 365.
- ^ a b c d e f "Soyut", McWhinney, Carpenter ve Rothschild (2001); sayfa 364.
- ^ a b c d e "Açıklama", McWhinney, Carpenter ve Rothschild (2001); sayfa 367.
- ^ a b c d "Sonuçlar", McWhinney, Carpenter ve Rothschild (2001); sayfa 376.
- ^ "Açıklama", McWhinney, Carpenter ve Rothschild (2001); sayfa 369.
- ^ a b "Açıklama", McWhinney, Carpenter ve Rothschild (2001); sayfa 370.
- ^ "Açıklama", McWhinney, Carpenter ve Rothschild (2001); sayfalar 370-371.
- ^ "Tartışma", McWhinney, Carpenter ve Rothschild (2001); sayfalar 373-373.
- ^ "Tartışma", McWhinney, Carpenter ve Rothschild (2001); sayfalar 373-374.
- ^ "Tartışma", McWhinney, Carpenter ve Rothschild (2001); sayfa. 374.
- ^ "Tartışma", McWhinney, Carpenter ve Rothschild (2001); sayfalar 374-375.
- ^ "Camarasaurus supremus, "Foster (2007). Sayfa 201." Bolluklar ve Çeşitlilikler "ibid. Sayfa 248.
- ^ "Ek", Foster (2007). Sayfa 328.
- ^ Trujillo, K.C .; Chamberlain, K.R .; Strickland, A. (2006). "Güneydoğu Wyoming'in Üst Jura Morrison Formasyonu için SHRIMP analizinden Oxfordian U / Pb yaşları, biyostratigrafik korelasyonlar için çıkarımlar". Amerika Jeoloji Topluluğu Programlı Bildiri Özetleri. 38 (6): 7.
- ^ Bilbey, S.A. (1998). "Cleveland-Lloyd Dinozor Ocağı - yaş, stratigrafi ve çökelme ortamları". Carpenter, K .; Chure, D .; Kirkland, J.I. (eds.). Morrison Formasyonu: Disiplinlerarası Bir Çalışma. Modern Jeoloji. 22. Taylor ve Francis Group. s. 87–120. ISSN 0026-7775.
- ^ Russell, Dale A. (1989). Zamanda Bir Odyssey: Kuzey Amerika Dinozorları. Minocqua, Wisconsin: NorthWord Press. sayfa 64–70. ISBN 978-1-55971-038-1.
- ^ Foster, J. (2007). "Ek." Jurassic West: Morrison Formasyonunun Dinozorları ve Dünyaları. Indiana University Press. s. 327-329.
- ^ Foster, John (2007). "Allosaurus fragilis". Jurassic West: Morrison Formasyonunun Dinozorları ve Dünyaları. Bloomington, Indiana: Indiana University Press. s. 170–176. ISBN 978-0-253-34870-8. OCLC 77830875.
- ^ Foster, John R. (2003). Rocky Mountain Bölgesi, Morrison Formasyonunun (Üst Jura) Omurgalı Faunasının Paleoekolojik Analizi, ABD. New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni, 23. Albuquerque, New Mexico: New Mexico Doğa Tarihi ve Bilim Müzesi. s. 29.
- ^ Dodson, Peter; Behrensmeyer, A.K .; Bakker, Robert T .; McIntosh, John S. (1980). "Jurassic Morrison Formasyonunun dinozor yataklarının tafonomisi ve paleoekolojisi". Paleobiyoloji. 6 (2): 208–232. doi:10.1017 / S0094837300025768.
- ^ Carpenter, Kenneth (2006). "Büyüklerin en büyüğü: mega-sauropodun kritik bir yeniden değerlendirilmesi Amphicoelias fragillimus". Foster, John R .; Lucas, Spencer G. (editörler). Üst Jurassic Morrison Formasyonunun Paleontolojisi ve Jeolojisi. New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. 36. Albuquerque, New Mexico: New Mexico Doğa Tarihi ve Bilim Müzesi. s. 131–138.
Kaynaklar
- Cope, E.D. (1877a). "Colorado'nun Dakota eopoc'undan devasa bir Suudi’de." Paleontolojik Bülten (25): 5–10.
- Cope, E.D. (1877b). "On Reptilian, Colorado'nun Dakota Yatakları'ndan kalmıştır". Paleontoloji Bülteni (26): 193–197.
- Cope, E.D. (1877c). "On Amphicoelias, Colorado'nun Dakota döneminden bir Saurian cinsi". American Philosophical Society'nin Bildirileri (17): 242–246.
- Cope, E.D. (1878). "Colorado'nun Dakota Devri Omurgalıları Üzerine". Paleontoloji Bülteni. 17 (28): 233–247. JSTOR 982303.
- Cope, E.D. (1879). "Yeni Jurassic Dinosauria". Amerikan doğa bilimci. 13: 401–404.
- Marsh, O.C (1877). "Jurassic Formasyonundan Yeni Dinosauri Sürüngenlerinin Bildirimi". American Journal of Science. 3. seri. 14 (84): 514–516. Bibcode:1877AmJS ... 14..514M. doi:10.2475 / ajs.s3-14.84.514. S2CID 130488291.
- Marsh, O. C. (1878a). "Yeni Dinosauri Sürüngenlerinin Bildirimi". American Journal of Science. 3. seri. 15 (87): 241–244. Bibcode:1878AmJS ... 15..241M. doi:10.2475 / ajs.s3-15.87.241. S2CID 131371457.
- Marsh, O. C. (1878b). "Amerikan Jura Dinozorlarının Başlıca Karakterleri, Bölüm I". American Journal of Science. 3. seri. 16 (95): 411–416. doi:10.2475 / ajs.s3-16.95.411. hdl:2027 / hvd.32044107172876. S2CID 219245525.
- Marsh, O.C (1889). "Avrupa ve Amerika Dinosauria'nın temel formlarının karşılaştırılması". American Journal of Science. 3. seri. 37 (220): 323–330. doi:10.2475 / ajs.s3-37.220.323. S2CID 130946518.
- Holland, W.J. (1919). "Bölüm V. Paleontoloji". 31 Mart 1919'da sona eren yıla ait müdürün yıllık raporu (Rapor). Carnegie Müzesi. s. 38. hdl:2027 / osu.32435027945948.
- Holland, W.J. (1924). "Uintasaurus douglassi HOLLAND Türünün Açıklaması". Carnegie Müzesi Yıllıkları. 15 (2–3): 119–138.
- Osborn, H. F .; Mook, C.C. (1921). "Camarasaurus, Amphicoelias ve Cope'un diğer sauropodları". Amerika Jeoloji Derneği Bülteni. 30 (1): 379–388. Bibcode:1919GSAB ... 30..379O. doi:10.1130 / GSAB-30-379. hdl:2027 / mdp.39015042532476.
- Elinger, T.U.H. (1950). "Camarosaurus annae-yeni bir Amerikan sauropod Dinozoru". Amerikan Doğa Uzmanı. 84 (816): 225–228. doi:10.1086/281626.
- Jensen, J.A. (1988). "Colorado Platosu'nun Üst Jura'ından dördüncü yeni bir sauropod dinozoru ve sauropod iki ayaklılığı". Great Basin Naturalist. 48 (2): 121–145. JSTOR 41712420.
- McIntosh, J. S .; Miller, W. E .; Stadtman, K. L .; Gillette, D. D. (1996). "Camarasaurus lewisi'nin Osteolojisi (Jensen, 1988)". Brigham Young Üniversitesi Jeoloji Çalışmaları. 41: 73–95.
- McIntosh, J. S .; Miles, C. A .; Cloward, K. C .; Parker, J.R. (1996). "Camarasaurus'un Yeni Neredeyse Tamamlanmış İskeleti". Gunma Doğa Tarihi Müzesi Bülteni (1): 1–87.
- Dodson, Peter; Britt, Brooks; Carpenter, Kenneth; Forster, Catherine A .; Gillette, David D .; Norell, Mark A .; Olshevsky, George; Parrish, J. Michael; Weishampel, David B. (1993-01-01). Dinozorlar Çağı. Yayınlar International, LTD. ISBN 978-0-7853-0443-2.
- Foster, J. (2007). Jurassic West: Morrison Formasyonunun Dinozorları ve Dünyaları. Indiana University Press. ISBN 978-0-253-34870-8.
- Tidwell, V .; Carpenter, K .; Meyer, S. (2001). Tanke, D. H .; Carpenter, K. (editörler). "Utah, Cedar Dağı Formasyonunun Zehirli Şeridi Üyesinden (Alt Kretase) Yeni Titanosauriform (Sauropoda)". Mesozoik Omurgalı Yaşamı: 139–165.
- Wings, Oliver; Sander, P. Martin (2007). "Sauropod dinozorlarında mide değirmeni yok: devekuşlarında mide taşı kütlesi ve işlevinin analizinden yeni kanıtlar" (PDF). Royal Society B Tutanakları. 274 (1610): 635–640. doi:10.1098 / rspb.2006.3763. PMC 2197205. PMID 17254987. Arşivlenen orijinal (PDF) 2011-07-13 tarihinde. Alındı 2009-07-24.
- McWhinney, Lorrie; Carpenter, Kenneth; Rothschild, Bruce (2001), "25. Dinosaurian Humeral Periostitis: Fosil Kayıtlarında Bir Juxtacortical Lezyon Olgusu", Mesozoik Omurgalı YaşamıIndiana University Press, s. 364–377
Dış bağlantılar
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||












