Spinophorosaurus - Spinophorosaurus
| Spinophorosaurus | |
|---|---|
 | |
| 3D baskılı yeniden inşası holotip iskelet Naturkundemuseum Braunschweig; yerdeki fosiller otantik | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Alttakım: | †Sauropodomorpha |
| Clade: | †Sauropoda |
| Clade: | †Gravisauria |
| Cins: | †Spinophorosaurus Remes et al., 2009 |
| Türler: | †S. nigerensis |
| Binom adı | |
| †Spinophorosaurus nigerensis Remes et al., 2009 | |
Spinophorosaurus bir cins nın-nin Sauropod Dinozor şimdi ne yaşadı Nijer esnasında Orta Jura dönem. İlk iki örnek 2000'li yıllarda Alman ve İspanyol ekipler tarafından zor şartlar altında kazıldı. İskeletler Avrupa'ya getirildi ve dijital olarak kopyalandı. Spinophorosaurus iskeletine sahip ilk sauropod 3D baskılı ve gelecekte Nijer'e iade edileceklerdi. İki örnek birlikte, cinsin iskeletinin çoğunu ve en çok bilinenlerden birini temsil ediyordu. baz alınan zamanının ve yerinin sauropodları. İlk iskelet, holotip örneği yeni cins ve türlerin Spinophorosaurus nigerensis 2009 yılında; genel ad ("omurga taşıyan kertenkele"), başlangıçta çivili olduğu düşünülen şeyi ifade eder osteodermler ve özel adı (Niger ve -ensis) bulunduğu yeri ifade eder. Aynı bölgeden bir yavru sauropod daha sonra cinse atandı.
Alt yetişkin holotip örneğinin yaklaşık 13 m (43 ft) uzunluğunda olduğu tahmin edilirken, paratip yaklaşık 14 m (46 ft) uzunluğundaydı. Bu bireylerin ulaştığı omuz yüksekliğinin yaklaşık 4 m (13 ft) ve ağırlığın yaklaşık 7 metrik ton (7.7 kısa ton) olduğu tahmin edildi. Braincase kısa, derin ve genişti ve nöroanatomi bazal ile bazı açılardan orta Sauropodomorflar ve daha fazlası türetilmiş neozoropodlar. Dişler spatulaydı (kaşık şeklinde) ve geniş aralıklıydı dişler tacın tepesinde, sauropodlarda atalardan kalma bir özellik. Boyun Spinophorosaurus 13 içeren sauropodlar arasında en tamamen bilinenlerden biridir omur. Sırt omurları, daha sonra daha fazla türetilmiş sauropodların tipik bir özelliği olan, çok sayıda küçük hava dolu iç odaya sahipti. Kuyruk güçlü kas sistemiyle güçlendirilmişti ve uzun ve üst üste binme nedeniyle oldukça sert olan bir arka bölüme sahipti. Chevron kemikleri. Kemiklerin başlangıçta kuyruk ucuna yerleştirilen sivri uçlu osteodermler olduğu düşünülen kemiklerin daha sonra köprücük kemiği.
Spinophorosaurus ya çok bazal bir sauropod ya da içeride sınıflandırılmıştır Eusauropoda, daha türetilmiş bir grup. Örneklerin anatomisi, yaşı ve konumu, sauropod evriminde önemli gelişmelerin Kuzey Afrika, muhtemelen tarafından kontrol edilir iklim bölgeleri ve bitki biyocoğrafya. Özellikleri vestibüler aparat görme ve koordineli göz, baş ve boyun hareketlerinin önemli olduğunu öne sürmek Spinophorosaurus. İskeletin 3 boyutlu modelleri, iskeletini test etmek için kullanıldı. hareket açıklığı. Bir çalışma, yüksek olabileceğini öne sürüyor tarayıcı ve bir başkası olası çiftleşme duruşlarını inceledi. Omurların merkezi ile sinir kemerleri arasındaki dikişler, ön kısımda daha karmaşıktı. gövde nın-nin Spinophorosaurus, çünkü bu bölgede muhtemelen en fazla stres yaşanıyordu. Spinophorosaurus dan bilinmektedir Irhazer Shale, bir jeolojik oluşum çağda Orta Jura olduğu düşünülüyordu. Büyük bir nehir-vadi sisteminde nehirlerden ve göllerden birikintilerle oluşmuştur.
Keşif

Zengin dinozor faunası Nijer 1960'larda ve 1970'lerde Fransız ve İtalyan kazıları ile bilimsel olarak dikkat çekmiştir. Bu, yeni cins itibaren Alt Kretase kayalar, en belirgin şekilde iguanodonti Ouranosaurus. Daha eski bir kaya dizisi, Tiourarén Formasyonu Amerikalı paleontolog tarafından araştırıldı Paul Sereno 1999 ve 2003 yılları arasında Nijer'de geniş çaplı bir kazı çalışması yürütmüş olan. Başlangıçta Alt Kretase'ye düştüğü düşünülse de, şu anda formasyonun çok daha eski olduğu düşünülmektedir. Orta Jura yaş. Sereno, yeni dinozorları adlandırdı. Sauropod Jobaria ve Theropod Afrovenator Tiourarén'den; buluntuların çoğu olarak bilinen bir uçurum boyunca keşfedildi Falaise de Tiguidit güneyde Agadez Bölgesi. İçinde Marendet, Sereno kısmi ayrıldı Jobaria bir turistik cazibe olarak sahada iskeletler.[2]:11–13[1] 2003 yılında başlayan PALDES projesi (Paleontología y Desarrollo - "Paleontoloji ve geliştirme") güney Agadez Bölgesi'nde kazılar yürütüyordu. İspanyol bilimi ve insani yardım kurumları arasında ortak bir çaba olan PALDES, paleontolojik araştırmaları bölge için bir gelişim programı ile birleştirmeyi amaçladı. Bu, Tadibene'de yeni bir paleontoloji müzesinin planlanan inşaatı da dahil olmak üzere altyapının iyileştirilmesi, eğitim yapıları ve turizmin teşvikini içeriyordu.[3]
2005 yılının başlarında, Alman kaşifler Ulrich Joger ve Edgar Sommer yerel bölgeden sonra Agadez'in güneyindeki yarı çölleri keşfettiler. Tuaregler Sommer'e bölgede büyük kemiklerin oluştuğunu bildirmişti. Sommer, Tuareg halkı için yerel eğitim sistemini iyileştirme konusunda uzmanlaşmış bir yardım kuruluşu olan CARGO'nun kurucusudur; Joger ise bir biyolog ve Devlet Doğa Tarihi Müzesi, Braunschweig, Almanya. Dönüş yollarında, bir grup Tuareg ile sohbet ettiler ve ardından onları küçük kemik parçalarıyla dolu yakınlardaki tepelik bir alana yönlendirdiler. Bu bölge (dinozorların daha önce kazılmadığı yer), Kuzey Denizi'nin yaklaşık 30 km (19 mil) kuzeyindedir. Falaise de Tiguidit ve kasabanın yakınında Aderbissinat Agadez Bölgesi'nde ve Irhazer Shale (veya Argiles de l'Irhazer), bir jeolojik oluşum Tiourarén'in altında (ve dolayısıyla biraz daha yaşlı). Bir saatlik bir aramadan sonra Joger, yüzeyden dışarıya çıkan yuvarlak bir kemik ucu keşfetti ve bu, daha fazla kazıdan sonra tam bir kazı olduğu ortaya çıktı. uyluk (uyluk kemiği) daha sonra ne olacağı holotip örneği nın-nin Spinophorosaurus. İlişkili kürek kemiği (kürek kemiği) ve bir omur kısa süre sonra keşfedildi. tortu yörede sert ama kırılgan silttaşı hafif çekiç darbeleri ile kemiklerden çıkarılabilir.[2]:17–27, 38[1][4]

Joger ve Sommer daha sonra destek için yerel Tuareg'leri işe aldı ve iki gün sonra, neredeyse eksiksiz, eklemli bir vertebral kolon ve birkaç uzuv ve pelvik kemik içeren numunenin çoğunu ortaya çıkardı. Omurga sütunu neredeyse tam bir daire oluşturdu, kuyruğun ucu kafatasının bekleneceği, ancak bulunamayacağı yere yerleştirildi. Ekipman ve kazı izni olmadığından, numuneyi enkazla kapladılar ve şimdi Braunschweig müzesi tarafından yürütülecek tam ölçekli bir bilimsel kazı planlayarak Almanya'ya döndüler. Müzeye 2006 yılında Nijer Cumhuriyeti tarafından resmi kazı izni vaat edildi; Bunun karşılığında müze, Injitane yerleşim yerinde yerel Tuareg çocukları için yeni bir okul inşa edip donatacaktı. 2006 sonbaharında, Sommer ve Joger, Braunschweig müzesinin diğer ortakları ile birlikte, kazı hazırlıkları için bölgeyi yeniden ziyaret ettiler ve test ekipmanı ve metodoloji için pelvik kemiklerden birini alçıya koydular. Ekip ayrıca siteden yaklaşık 1 km (0,6 mil) uzaklıkta theropod izlerini keşfetti. Hem okulu hem de kazıyı finanse edecek sponsorlar 2007'nin başlarında bulundu. "Projekt Dino" adlı resmi kampanya, 1 Mart 2007'de, ekipmanlı iki kamyonun Braunschweig'den Nijer'e doğru yola çıkıp İspanya, Fas, Moritanya üzerinden bir rotaya girmesiyle başladı. ve Mali (nehrin içinden geçen kısa rota Sahra terörist saldırı riski nedeniyle mümkün değildi). Ekibin on daimi üyeden oluşan diğer kısmı uçakla geldi.[2]:17–27, 38[1][5][6] Bu, neredeyse bir asırdır Afrika'ya ilk Alman dinozor keşif gezisiydi.[1]:29[7]
Bu arada, liderliğindeki İspanyol projesi PALDES ekibi Elche Paleontoloji Müzesi, bölgede çalışıyordu. 2007 yılının başlarında, Aderbissinat belediye başkanı Mohamed Echika, PALDES ekibinin daha önce Almanlar tarafından keşfedilen iskeleti kazmasına izin verdi; iskelet daha sonra İspanya'ya gönderildi. Bu faaliyetlerden habersiz, Alman ekibinin öncüsü, 16 Mart'ta vardıklarında boş bir kazı alanı buldu (profesyonel bir kazı izleri gösteren); kamyonlar 20 Mart'ta geldi. Alman ekibi hayal kırıklığına uğramasına rağmen ikinci bir Spinophorosaurus örnek, gelecek paratip, 17 Mart'ta, ilkinden 15 m (49 ft) uzakta, küçük kemik parçalarıyla dolu bir alanda keşif amaçlı bir açmada kısa süre sonra çene ve diş parçaları ortaya çıktı; ertesi gün kaburgalar, omurlar, a humerus (üst kol kemiği) ve bir kürek kemiği tanımlanabilir. 19 Mart'ta sekiz yerel kazı yardımcıları gruba katıldı. 20 Mart'ta, kamyonların gelmesinden önce, yerel yardımcıların önceki gece yıkamak için kullanması nedeniyle başlangıçta 200 l (53 ABD galonu) olan tatlı su rezervi tükendi. ekip üyeleri bayılacak. Kazı, sıcaklıklar 43–45 ° C'ye (109–113 ° F) ulaştığında genellikle 12:00 ile 15:00 arasında kesintiye uğradı. 25 Mart'ta, Alman ekip üyelerinin ikisi dışında hepsi hasta, acı çekiyordu. ishal ve dolaşım sorunları. Kazı boyunca ilerleme fotoğraflar ve saha notları ile belgelenmiştir.[2]:29–45[8]

27 Mart'a kadar, humerus, kürek kemiği ve gelecekteki paratipin çoğu kaburga kemiği çoktan koruyucu sıva ile sarılmış ve çıkarılmıştı. İskeletin altında başka kemik görünmese de ekip, tüm fosillerin toplandığından emin olmak için fazladan 60-80 cm (24-31 inç) tortu çıkardı.[2]:60 2 Nisan'da kazı tamamlandı ve fosiller limana taşınmak üzere paketlendi. Cotonou 3 Nisan tarihinde Echika, ekibe ilk iskeletin izniyle bir İspanyol grup tarafından kazıldığını açıkladı. Ekibi, Agadez'in yaklaşık 80 km (50 mil) güneyinde, Tiguidit uçurumunda bulunan başka bir fosil bölgesine telafi olarak götüreceğine söz verdi. Ekip orada olası bir oyunun arka kısmını buldu. Jobaria iskelet ancak önümüzdeki sezona kadar sahadaki en büyük bloğu terk etmek zorunda kaldı. Başkalarını bloğu toplamaktan caydırmak için, patlayıcı bir kukla üretildi ve fosile, İspanyolca bir uyarı etiketi ile yapıştırıldı.[2]:68–73[9] Alman takımı, 2008'de bir sonraki sezon bloğu aldı; PALDES ekibi, salgınların patlak vermesinin ardından o yıl için kazı planlarını iptal etmişti. Tuareg isyanı (2007–2009).[2]:100, 107

İki Spinophorosaurus örnekler geçici olarak İspanyol ve Alman müzelerinde barındırılıyordu. Nijer Cumhuriyeti ile sözleşme yaparak, gelecekte ülkeye geri gönderileceklerdi. Musée National d'Histoire Naturelle içinde Niamey yanı sıra daha küçük, yeni inşa edilmiş bir yerel müze.[2]:143 Gelecekteki paratip örneği 18 Mart 2007'de Almanya'ya geldi; Braunschweig müzesi, iki buçuk yıl süren hazırlığı için ayrı bir fabrika binası kiraladı. Buna paralel olarak, şu anda işbirliği yapan Alman ve İspanyol ekipleri tarafından ortak bir makale hazırlandı. Alman ekibi, lazer taramasını kullanarak her iki numunenin kemiklerini ve parçalarını 3D olarak dijital hale getirdi. Braunschweig'deki iskeletin yalnızca% 70'i tamamlandığından, İspanya'daki örnek eksik parçaları doldurmak için kullanıldı; Bu süreçte, İspanya'daki iskeletin, Almanların başlangıçta keşfettiği ve kaybettiği iskelet olduğu keşfedildi. 3D taramalar dijital olarak onarıldı ve deforme edilmedi, 3D olarak basılmıştır ve Braunschweig müzesinin sergisi için monte edilmiş bir iskelete monte edildi (3D baskı ile yeniden üretilen ilk sauropod iskeleti). Yaşam boyu bir yaşam modeli Spinophorosaurus"Namu" lakaplı (müzenin adından sonra) müzenin ana girişinin önüne dikildi.[2]:79–85[10][11] İspanyol ekip, holotip fotoğraflarından ayrı 3B modeller üretti. fotogrametri (haritalamak için bir nesnenin farklı açılardan fotoğraflarının çekildiği yerlerde);[12] 2018'de Elche müzesinde bir kuyruk omuru sergilendi.[13] Garcia-Martínez ve meslektaşları 2018 konferans özetinde, daha iyi korunmuş birinci ve beşinci arka omurlara dayalı olarak kötü korunan ikinci arka omurun morfolojisini yeniden yapılandırdıklarını açıkladılar. Bu kullanılarak yapıldı dönüm noktası tabanlı geometrik morfometri, karşılık gelen 3B koordinatların her bir omurdan toplandığı ve istatistiksel olarak analiz edildiği yerlerde.[14]
İlk iskelet (GCP-CV-4229 olarak kataloglandığı Elche müzesi ile NMB-1699-R olarak kataloglanan Braunschweig müzesi arasında bölünmüştür.[15]) yeni cins ve türlerin holotipi yapıldı Spinophorosaurus nigerensis Alman paleontolog Kristian Remes ve 2009 yılında Alman ve İspanyol ekiplerinden meslektaşları tarafından. Jenerik isim şunlardan oluşmaktadır: spina, Latince "başak"; PhoroYunanca "dayanmak"; ve Sauros, yani kertenkele (tam anlamıyla "omurga taşıyan kertenkele") anlamına gelir: ad, başlangıçta şu şekilde yorumlandı osteodermler daha sonra temsil ettiği öne sürülen sivri uçlu köprücük kemiği yerine. Spesifik ad, Nijer'e atıfta bulunur. takson keşfedildi.[16][1][17][18] Holotip örneği, bir Braincase, bir postorbital kemik, bir skuamozal, bir uydurmak, bir pterygoid, bir yuvarlak ve neredeyse tamamlandı postkraniyal iskelet eksik olan göğüs kemiği, antebrachium, manus ve pes. İkinci, paratip örneği (NMB-1698-R), kısmi bir kafatası ve tamamlanmamış bir postkraniyal iskeletten oluşur. Bu örnekte korunan ancak holotipte korunmayan öğeler, bir premaksilla, üst çene, gözyaşı, diş hekimi, açısal, sağ tarafın sırt kaburgaları, kol kemiği ve bir pedal falanksı. Aralarında çakışan iskelet elemanlarının aynı olması ve aynı stratigrafik tabakadaki iskeletlerin yakınlığı nedeniyle iki örnek aynı taksona ait kabul edilir.[1] Tarif edildiği zamanda, Spinophorosaurus Kuzey Afrika'dan en çok bilinen Orta Jura sauropod'uydu,[1] ve 2018 itibariyle en eksiksiz erken dönemlerden biri olarak kabul edildi (neosauropod ) sauropodlar.[19]
2012'de, PALDES ekibinden Adrián Páramo ve Francisco Ortega, ikisinden birkaç metre uzaklıkta, yerde keşfedilen küçük bir sauropod iskeleti (örnek GCP-CV-BB-15) bildirdi. Spinophorosaurus örnekler; tüm fosiller muhtemelen aynı yataklardandır. Küçük iskelet, tüm boyun omurları ve bazı sırt omurları dahil olmak üzere 14 omurdan (bazıları eklemli) oluşur. Omurgasının merkezi, ormandan% 20 daha küçüktür. Spinophorosaurus ve nöroentral sütür açık, bu da onun bir genç olduğunu gösteriyor. İskeletin birkaç farklı özelliği, Spinophorosaurusve bu cinsin bazı özellikleri mevcut olmasa da, büyük olasılıkla bir genç Spinophorosaurus (farklılıklar muhtemelen ontogeny, büyüme sırasındaki değişiklikler).[20][15]
Açıklama

Holotip numunesi, vertebral kolon boyunca ölçüldüğünde yaklaşık 13 metre (43 ft) uzunluğundayken, paratip yaklaşık yüzde 13 daha büyüktü ve yaklaşık 14 m (46 ft) ölçüldü.[1][2] Holotip iskeletinin 3 boyutlu fotogrametri modeli baştan kuyruğa 11,7 m (38 ft) boyutlarındadır, oranlar 2B iskelet rekonstrüksiyonlarına dayanan tahminlerden farklıdır.[12] Bu bireylerin ulaştığı omuz yüksekliğinin yaklaşık 4 m (13 ft) olduğu tahmin edilmektedir.[2] ve ağırlık yaklaşık 7 metrik ton (7.7 kısa ton).[1][21] Holotip numunesinin endokraniyal ve nöromerkezi dikişler sırasıyla kafatasında ve omurlarda kaynaşmamış, bu da onun bir alt yetişkin olduğunu gösterirken, paratip örneğinde tamamen kaynaşmış nöro-merkezden sütürler var.[1]
2020'de Vidal ve meslektaşları, Spinophorosaurus, dijital iskeletin montajına göre. Orijinal 2009 iskelet yeniden yapılandırması dinozoru yatay bir duruşla gösterirken, dijital rekonstrüksiyon uzun omuzlar ve yükselmiş bir boyun ile daha dikey bir duruş gösterdi.[22]
Kafatası

Ön kemikleri kafatası çatı kafatasının diğer dikişlerinden farklı olarak orta hatta kaynaşmıştır. Cepheler arasında küçük bir pineal foramenfrontal ve paryetal kemikler arasında sütürün önünde yaklaşık 10 mm (0,4 inç). Kafatasının arka kenarında açık bir postparietal çentik vardı, aksi halde yalnızca dikraeozorlar ve Abrosaurus. Yapışık ön kemiklerin önündeki orta çizgiyi takip eden bir çentik, her bir burun kemiğinden ön kısımların arasına uzanan bir sivri uç olabileceğini gösterir. Nigersaurus ancak sauropodlar arasında genel olarak alışılmadık bir durumdur. Kafatasının çatısındaki bir çift büyük açıklık olan üst temporal fenestra, uzun olandan dört kat daha genişti; bu kadar genişletilmiş fenestralar için tipikti Shunosaurus ve dahası türetilmiş (veya "gelişmiş") sauropodlar. oksipital kondil kafatasının arkasındaki gibi yanlarda içbükey Shunosaurus. bazal tubera (kafatasının alt tarafındaki kas bağlantıları görevi gören bir çift uzantı) genişletilmiş ve bilinen sauropodlar arasında benzersiz olan yanlara yönlendirilmişti. Kuadratın arka tarafında bir içbükeylik yoktu, atalara ait aksi takdirde sadece bilinen durum Tazoudasaurus sauropodlar arasında. Spatula (kaşık şeklindeki) dişler, genişletilmiş, aralıklı olarak benzersizdi. dişler kronun ön kenarında daha yüksek diş sayısı ile kronun üstünün etrafında.[1][23]
Beyin kasası Spinophorosaurus önden arkaya geniş ve kısaydı, orta derecede derin ve genel olarak nispeten büyük boyuttaydı. Diğer Jurassic sauropodlardan önemli ölçüde farklıydı, muhtemelen hariç Atlasaurus. Paylaştılar basipterygoid süreci üzerinde kendi bazfenoidler güçlü bir şekilde geriye doğru yönlendirilmeleri benzerdi. DAVRANMAK endocast Holotipin beyin boşluğunun% 50'si, iyi işaretlenmiş pontin ve serebral fleksiyonlara, büyük ve dikdörtgen bir hipofiz fossasına sahip olması ve beynin yapısının nispeten kalın olan boşluklarla örtülmüş olması bakımından çoğu sauropoda benzediğini gösterdi. meninksler yanı sıra dural venöz sinüsler. labirent of İç kulak uzun ve ince olması karakteristikti yarım dairesel kanallar, aşağıdakilere benzer Massospondylus ve Giraffatitan. Onun nöroanatomi bazı yönlerden ortadaydı baz alınan (veya "ilkel") sauropodomorflar ve daha türetilmiş neosauropodlar.[23]
Omurga ve kaburgalar

Omurga sütunu neredeyse tamamen bilinmektedir ve holotip, tam bir boyun içeren birkaç sauropod örneğinden biridir.[24] Boyun 13'ten oluşuyordu boyun omurları. Bagajda 12 sırt ve dört sakral omur. Kuyruk 37'den fazla içeriyordu kaudal omur.[1] Karmaşık elemanlar, tek tek omurlar bir alt kısımdan oluşur, merkezve bir üst kısım, sinir kemeri. Sinir kemerinin önemli yer işaretleri, yukarı doğru projeksiyonu içerir. nöral omurga (dikenli süreç) ve yana doğru çıkıntı diyapofizler, bunlar birlikte omurlara önden ve arkadan görünümlerde T şekli verir. Çift eklem süreçleri komşu omurlarla bağlantı kurmak önden çıkıntı yapıyor (prezygapophyses) ve arka (postzygapophyses).[25]
Servikal omurlar, Jobaria ve Cetiosaurus. Merkezlerinin uzunluğu yaklaşık 3.1 kat daha genişti; bu nedenle genel olarak sauropodlara kıyasla orta derecede uzamışlardı, ancak genellikle diğer bazal formlardan daha uzundu. Boyun merkezinin yan taraflarında öne doğru derinleşen büyük kazılar vardı; böyle pleurocoels ayrıca geliştirildi Jobaria ve Patagosaurus. Aksine Jobariapleurocoeller, eğik bir kemikli sırt ile alt bölümlere ayrılmamıştı. Merkezin ön ucunun alt tarafında orta hat omurgası mevcuttu ve Cetiosaurus. Prezygapophyses uçları üçgen bir uzantıya sahipti ve bu da Jobaria, bu cinste daha derin olmasına rağmen. Postzigapofizlerin üstünde nispeten büyüktü epipofizler, kas bağlanması için kemik çıkıntıları. Servikal omurlar, Güney Amerika ve Hindistan'daki bazal sauropodlardan farklıydı. Diyapofizler (sinir kemerinin yana bakan süreçleri) hafifçe aşağı bakacak şekilde eğimliydi ve arka kenarlarında üçgen flanşlar vardı - bu güney formlarında görülmeyen özellikler. Dahası, sinir dikenleri arka ve ön yüzeylerinde buruşuktu (buruşuk) ve boyun tabanına yakın, yandan görünüşte daha geniş ve daha az yüksekti. Yandan bakıldığında, merkez ile sinirsel ark arasında U şeklinde bir çöküntü mevcuttu. otapomorfi (benzersiz özellik) Spinophorosaurus.[1]

Sırt omurları, devetüyü iç yapıya sahip olmaları bakımından alışılmadık bir durumdu (çok sayıda küçük hava dolu odacık içerir). Bu özellik, aksi takdirde çok daha sonra bilinir. Titanosauriforms yanı sıra Mamenchisauridler, nerede bağımsız olarak gelişti eski gruptan.[26] Ön dorsal omurlar, merkezlerinde derin pleurocoeller göstermesine rağmen, bu açıklıklar gövdenin arkasına doğru çok daha sığ hale geldi. En arkadaki dorsal omurlar da orantılı olarak kısaydı. İçinde Amygdalodon ve Patagosaurus tersine, arkaya doğru sırtlar daha uzundu ve pleurocoelleri belirgin hale getirdi. Sırt omurlarının sinir kanalı çok dar fakat yüksekti.[1] Hiposfan-hipantrum eklemleri (aksesuar eklem süreçleri) tüm sırtlarda mevcuttu ve bu da omurgayı daha sert hale getirdi.[1][27] Sinir dikenleri, diğer bazal sauropodlarda olduğu gibi ön ve arka taraflarında belirgin buruşukluklara sahipti. En öndeki kaudal omurlar, nöral dikenlerde, sırtlarda görüldüğü gibi aynı buruşukluklara sahipti, aksi takdirde yalnızca Omeisaurus. Kuyruğun arka kısmında, sinir dikenleri kuvvetli bir şekilde arkaya doğru eğimlidir ve bazı Doğu Asya sauropodlarına benzer şekilde, sonraki omurun ön kısmına uzanır. Barapasaurus, ve Jobaria.[1]
İkinci ila beşinci sırt omurlarının kaburgaları düzleştirilmiş ve geriye doğru yönlendirilmişken, altıncı ila on birinci sırtın kaburgaları enine kesitte daha dairesel ve daha dikey olarak yönlendirilmişti. Göğüs kafesi bu nedenle açıkça pektoral ve lomber bölüme ayrılabilir; böyle bir farklılaşma sadece tek bir başka sauropod olan dikraeosaurid'de tanımlanmıştır. Brachytrachelopan. Ayrıca, pektoral kaburgaların uçlarında, göğüs kemiğine bağlanan göğüs kemiği kaburgaları için bağlantı yerleri vardı. Kuyruğun ön kısmında köşeli çift ayraçlar (eşleştirilmiş vertebral merkezin altındaki kemikler), temel durum olan bıçak benzeri idi. Kuyruğun arka kısmında köşeli çift ayraçlar çubuk şeklindeydi ve sol ve sağ meslektaşları birbirinden ayrıldı. Bu çubuk benzeri köşeli çift ayraçlar, merkezin alt kenarlarına sıkıca tutturulmuş olurdu. Vertebral merkezin orta uzunluğunda önceki ve sonraki köşeli çift ayraçlar ile eklemlendi, böylece vertebral eklemi destekleyerek kuyruğun bükülmesini kısıtladılar.[1]
Kemerler, uzuvlar ve yanlış tanımlanmış kuyruk sivri uçları

Kürek kemiği, genişletilmiş, yelpaze şeklinde bir alt uç ile güçlü bir şekilde kavisli olan sauropodlar arasında benzersizdi. Eusauropodlar için bir özellik olan enine kesitte D şeklindeydi. Üst ucu genişletildi ve alt kenarda kemikli bir flanşa sahipti.[19] Bu açılardan Asya'daki mamenchisauridlere benziyordu, ancak Gondwanan formlarından farklıydı. Vulcanodon, Barapasaurus, ve Patagosaurusüst ucun sadece zayıf bir şekilde genişlediği ve arka flanşın olmadığı. korakoidkürek kemiğinin alt ucuna eklemlenen, otapomorf olarak kabul edilen belirgin bir böbrek şekli gösterdi. İçinde büyük bir pazı tüberkülü vardı. biceps brachii kas bağlı. Klavikulaya göre daha ince olmasına rağmen sağlamdı. Jobaria. Ön ayaklardan sadece humerus korunmuştur. Alt ucu asimetrikti ve genişletilmiş aksesuar kondilleri (kemiğin alt ön kenarındaki öne doğru çıkıntılar) vardı - aksi takdirde sadece mamenchisauridlerde görülen özellikler. Pelvisin pubis ve iskiumu sağlamdı, ikincisi sonunda genişledi. Üst femur (uyluk kemiği), bir küçük trokanter üst ucunda - arka ayağı öne ve içeri doğru çeken kaslar için bağlantı yeri görevi gören kemikli bir çıkıntı. dördüncü trokanter arka yüzeyden çıkıntı yapan ve arka ayağı geriye çeken tutturulmuş kaslar, özellikle Spinophorosaurus. Dördüncü trokanterin yakınında, diğer sauropodlarda bulunmayan büyük bir açıklık ve dolayısıyla bir otapomorfi vardı. Tibia (incik kemiği) diğer bazal sauropodlarınkine benziyordu ve fibula (baldır kemiği) sağlamdı. Ayak bileğinin üst tarafı astragalus Kemikli bir duvarla ayrılmayan tibia ve fibula ile artikülasyon için fasetlere sahipti ve sekize kadar besin foramina (kan damarlarının kemiğe girmesine izin veren açıklıklar).[1]

Başlangıçta sol ve sağ osteoderm (deride oluşan kemik) olarak yorumlanan elementler holotip iskelet ile bulundu. Bu kemikler, sivri uçlu bir çıkıntının çıktığı yuvarlak bir tabana sahipti; iç yüzeyler pürüzlü ve içbükeydi. Pelvik bölgede bulunmasına rağmen, Remes ve meslektaşları, canlı hayvanın kuyruğunun ucunda bulunduklarını ve bu cinsin ayırt edici bir özelliği olduğunu düşündüler. Bu pozisyon, sol ve sağ elementlerin birbirine yakın bulunmasına dayanıyordu, bu da onların vücudun orta hattından geldiklerini gösteriyordu. Ayrıca, kuyruk sopaları veya sivri uçlar gösteren diğer dinozorlarda da kuyruğun arka kısmının uzamış köşeli çift ayraçlarla sertleşmesi gözlemlenmiştir. Benzer dikenler, ilgili sauropoddaki kuyruk kulübünün parçasıydı. Shunosaurus; böyle bir kuyruk kulübü muhtemelen mevcut değildi Spinophorosaurus, en arkadaki kuyruk omurları çok küçük hale geldiğinden. Sağ taraftaki osteoderm, soldan biraz daha büyük ve şekli biraz farklıydı. Bu onların bir çift oluşturmadıklarını gösterir; bu durumda muhtemelen birbirlerinin tersine çevrilmiş karşılığı olurlar. Aksine, bu farklılıklar başlangıçta iki çift diken olduğunu gösterdi.[1]
2013 yılında, paleontologlar Emanuel Tschopp ve Octávio Mateus, varsayılan kuyruk sivri uçlarını yeniden incelediler ve diğer zırhlı dinozurlarda görülen osteodermlerin tipik engebeli yüzeyine veya kulübe benzer genişlemeye sahip olmadıklarını keşfettiler. Shunosauru. Kırık kenarları nedeniyle, bu elemanların başlangıçta önerildiği gibi farklı boyutlarda olup olmadığından da şüphe ettiler. Bu elementler kürek kemiğinin altında bulunduğundan, bunun yerine köprücük kemiğini temsil ettiklerini ve fosillerin bu açıdan yeniden değerlendirilmesi gerektiğini öne sürdüler.[18] Vidal ve meslektaşları 2015 ve 2018'de bu öneriyle hemfikirdi.[28][19]
Sınıflandırma


İlk Filogenetik analiz Remes ve meslektaşları tarafından sunulan önerdi Spinophorosaurus Bilinen en bazal sauropodlar arasında yer aldı, sadece biraz daha türetildi. Vulcanodon, Cetiosaurus, ve Tazoudasaurus. Bu pozisyonda, kardeş takson nın-nin Eusauropoda daha fazla türetilmiş sauropodları içeren bir grup. Yazarlar, bu çok bazal pozisyona verilen desteğin zayıf olduğunu kabul ettiler ve kuzey Afrika ve Laurasya'dan diğer sauropodlarla anatomik benzerlikleri açıklayabilecek özauropodlar içindeki birkaç alternatif yerleşimi tartıştılar.[1] Eusauropoda dışında benzer bir bazal konum, daha sonraki birkaç çalışma tarafından önerildi,[29][30][31] hangisi yerleştirildi Spinophorosaurus kardeş takson olarak Tazoudasaurus[29] veya Volkheimeria.[30] 2013 konferans özetinde, paleontolog Pedro Mocho ve meslektaşları, yeni hazırlanmış kemiklerden daha fazla bilgi ekleyerek, cinsin filogenetik ilişkilerini yeniden değerlendirdiler. Spinophorosaurus eusauropod'lar içinde yuvalanmıştı. Bu analize göre, cins, Shunosaurus ve Barapasaurus ve yakın Patagosaurus ve mamenchisaurids.[26] Eusauropoda'da çok daha türetilmiş sistematik bir konum, 2015 yılında yapılan bir çalışmada da önerildi. Spinophorosaurus kardeş takson olmak Nebulasaurus.[32]
Cladogram Nair ve Salisbury, 2012'ye göre, Spinophorosaurus:[29]
| Eusauropoda içinde daha türetilmiş bir pozisyonu destekleyen Xing ve arkadaşları, 2015'e dayanan Cladogram:[32]
|
Evrim
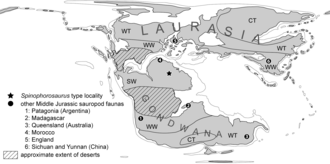
En çok bilinen bazal sauropodlardan biri olarak, Spinophorosaurus erken evrime ışık tutmaya yardımcı oldu ve paleobiyocoğrafya Grubun. Bu, özellikle Asya dışında, Erken ve Orta Jura kalıntılarının seyrekliği nedeniyle belirsizdi. Remes ve meslektaşları şunu buldu: Spinophorosaurus Orta Jura Doğu Asya sauropodları (özellikle boyun ve kuyruk omurları, kürek kemiği ve humerusta) ile özellikleri paylaşır, ancak Alt ve Orta Jura Güney Amerika ve Hint taksonlarından çok farklıdır (farklılıklar arasında omurga özelliklerinin şekli ve gelişimi ve kürek kemiğinin şekli bulunur ve humerus). Bunun Orta Jura sauropod faunaları ile açıklanabileceğini öne sürdüler. Laurasia (Kuzey süper kıta zamanın) ve Güney Gondvana (güney süper kıta) coğrafi engellerle ayrılmış durumda. Daha önce, sauropodların süper kıta boyunca dağıldığına inanılıyordu. Pangea (Laurasia ve Gondwana'dan oluşan) Erken ve Erken Orta Jura döneminde. Görece az çeşitliliğe sahipti. kıta dağıldı ve tek başına gelişen sauropod gruplarına yol açtı. vekalet Geç Orta ve Geç Jura döneminde. Spinophorosaurus Doğu Asya sauropodları arasında bir zamanlar türetildiği düşünülen özelliklerin plesiomorfik Özauropodlar (Laurasia'yı kolonileştiren öküropodlar) arasında (ataların özellikleri), aynı zamanda Spinophorosaurus). Güney Gondwana'yı kolonileştiren eusauropodlar, izolasyon sırasında söz konusu atasal özelliklerini kaybeden grubun özelleşmiş bir soyuydu. Remes ve meslektaşları, bu yorumları desteklemek için daha fazla kanıta ihtiyaç duyulduğunu ancak Kuzey Afrika, Avrupa ve Doğu Asya'nın Jura sauropodları arasında bir bağlantı olduğundan emin olduklarını belirtti.[1]
Anatomisinin gösterdiği gibi Spinophorosaurus ve Orta Jura sauropod dağılımının örüntüsü, sauropod evriminde önemli gelişmeler Kuzey Afrika'da meydana gelmiş olabilir. Yakındı ekvator Erken ve Orta Jura'da, yüksek bitki üretkenliği ve çeşitliliği ile yazın yağışlı bir iklime sahipti. Öte yandan, Güney Gondwana'nın sauropodları, Erken Jura'daki Orta Gondwanan Çölü tarafından ekvator bölgesinden izole edildi ve bu da Güney ve Kuzey Gondwanan floralarında farklılıklara yol açtı. Orta Jura'nın sonlarında çöl küçüldüğünde, neosauropodlar ortaya çıktı ve Orta Jura'nın tipik özelliği olan sauropod faunasının yerini aldı. Dan beri Jobaria Kuzey Afrika'nın Neosauropoda üssüne yakın olması nedeniyle diplodocoids Üst Jura, neosauropodlarda yaygın olarak dağılmışlardır ve bunların bazı alt grupları, sauropodların evriminde bir "sıcak nokta" olarak hareket etmiş olabilecek Jura Pangea'nın ekvator bölgesinden kaynaklanmış olabilir. Sauropodların Jurassic çeşitlendirmesi muhtemelen iklim bölgeleri ve sadece kıtasal farklılaşmadan ziyade bitki biyocoğrafyası.[1]
Paleobiyoloji

Spinophorosaurus ve diğer bazı sauropodomorflar azalmamış vestibüler cihazlar Bu, ağır, bitki yiyen dört ayaklılara yol açan bir soydan beklense de, iç kulakta denge ve yönelim için bir duyu sistemi. Neden bilinmiyor Spinophorosaurus bu özelliği korudu, ancak sauropodomorf labirentlerinin boyutu ve morfolojisi, örneğin boyun uzunluğu ve hareketliliğiyle ilişkili olabilir. Vestibüler aparatın genişlemesi, görme ve koordineli göz, baş ve boyun hareketlerinin öneminin bir göstergesidir, ancak sauropod vestibüler özelliklerinin yorumu hala belirsizdir.[23]
2018 konferansında Benjamin Jentgen-Ceschino ve meslektaşları, radyal olarak yönlendirilmiş kanallarla karakterize edilen bir kemik dokusu türü olan radyal fibrolameller kemik (RFB), korteks bir yetişkine ait humerusun (en dıştaki kemik tabakası) Spinophorosaurus örnek ve genç bir yetişkin Isanosaurus. Normalde, sauropodların uzun kemiklerindeki korteks bunun yerine fibrolameller kompleks gösterir. RFB, hızlı büyüyen kemik dokusu olarak kabul edilir ve daha önce sauropodlarda rapor edilmemiştir. En dış kortekste bulunması olağandışıdır, çünkü genellikle en içteki kortekste (juvenil büyümeye karşılık gelir) beklenir. RFB'si Spinophorosaurus dış kortekse gömüldü, ardından normal bir fibrolameller kompleksi izledi, bu da hayvanın hızlandırılmış büyümesinden sonra bir süre hayatta kaldığını gösteriyor ki bu muhtemelen patolojik yaralanma nedeniyle kemik büyümesi.[33]
Hareket

Dan beri Spinophorosaurus en çok bilinen bazal sauropodlardan biridir, için iyi bir modeldir. biyomekanik sauropod biyolojisini ve anatomik özelliklerin işlevini anlamaya yardımcı olan çalışmalar. 2015 yılında Vidal ve meslektaşları tarafından 3B modelin manipülasyonu, en öndeki 20 kuyruk omurunun her birinin 20 to'ye kadar ve yanlara 8-10 def'ye kadar sapabileceğini gösterdi. bükülme kemik morfolojisi ile sınırlandırılmadan önce. Canlı bir hayvandaki gerçek hareket aralığı daha küçük olurdu çünkü yumuşak doku ve büyük şerit kemikleri sınırlayıcı bir faktör olurdu. The first 20 tail vertebrae also became longer towards the end of the tail, as seen in diplodocids, and were powered by enlarged hypaxial musculature, as indicated by the large chevron bones and upwards-angled transverse processes in the first tail vertebrae. Movement in the rear part of the tail was limited as the chevrons were overlapping, as is also the case in dromaeosauridler ve ankilozoridler. Together, these features of the tail may have been part of a specialised function, which is unknown.[28] Joger and colleagues argued in 2009 that the supposed spikes formed a thagomiser at the end of the tail that would have been used for defence against predators.[2]
In 2017, John Fronimos and Jeffrey Wilson used Spinophorosaurus as a model to study how the complexity of neurocentral sutures (the rigid bağlantı connecting the neural arch of a vertebra to its centrum) in sauropods may have contributed to the strength of the spine. Sauropods reached their often enormous body sizes by both rapid and extended growth. As long as an individual was growing, the neurocentral sutures were composed of cartilage to permit this growth. Upon reaching full body size, this cartilage would turn into bone, closing the suture. Cartilage, however, is much less resistant to forces that could potentially dislocate the neural arch, such as bükme ve burulma; these become especially relevant at large body sizes. Sauropods and other archosauriforms counteract this structural weakness by increasing the complexity of the suture, meaning that the surfaces that connect the neural arch to its centrum had complex ridges and furrows that interlocked. İçinde Spinophorosaurus, suture complexity was most pronounced in the front part of the trunk, indicating that stresses were highest in this region, probably because of the weight of the long neck and rib cage. Complexity became weaker towards the skull and the sacrum. The orientation of the ridges allows for identifying the type of stress that affected the vertebra: In the neck vertebrae, ridges would mainly have prohibited dislocation of the neural arch in a front-to-back direction, while the ridges in the trunk vertebrae were more effective in impeding rotation.[34]
In a 2018 conference abstract, Vidal used the virtual Spinophorosaurus skeleton to test hypothetical mating postures that have been proposed for sauropods that would involve a "kloakal öpücük " (as is performed by most birds) rather than a male intromittent organ. Such hypotheses include "leg over back", the male mounting from behind (with the female standing or lying down), as well as "backwards mating". Postures that resulted in dislocation or osteological stops were not considered feasible. The "leg over back" posture was rejected, since it would be impossible for a standing male to reach its leg over the back and hip of a crouching female, even if it was half the size of the male (and it is unknown if sauropods were cinsel olarak dimorfik in size). The posture wherein the male mounted the female from behind while resting its front legs on the back of the female was possible (the tail was flexible enough to get out of the way), but even if the female was lying down, the cloacae would not approach each other close enough for a "cloacal kiss". The "cloacal kiss" would only be possible through "backwards mating", wherein the male and female approached each other backwards and joined their cloacae with their tails flexed sideways, and movement of the legs would allow individuals of different sizes to mate this way. "Backwards mating" is the only way sauropods could have mated without the need of a male intromittent organ. Such a pose is common in mating eklembacaklılar and in the "bağlı "-phase of mating etobur gibi memeliler Canis.[35]

Vidal and colleagues, in 2020, used 3D models of both the holotype and the juvenile skeleton to estimate the range of motion (flexibility) of the neck. Such estimates assume that the original distance between vertebrae can be reliably predicted, and that the articular processes stay in contact at all times. Vidal and colleagues demonstrated that both assumptions indeed hold true in modern zürafalar, increasing confidence in range of motion estimates in extinct animals in general. Gibi Spinophorosaurus grew, the range of motion of the neck would increase (making steeper neck postures possible); the gap between vertebrae become larger; and the neck become more inclined in neutral posture. Similar changes during growth are also observed in giraffes. The neck would have been as flexible as that of giraffes thanks to the higher number of vertebrae, even though the individual joints were much less flexible than in giraffes. Spinophorosaurus would possibly have been able to feed using the same postures as giraffes, and it could have been the most basal sauropod adapted for high browsing. High browsing is also suggested by anatomical features, including the narrow snout, broad teeth, and proportionally long humerus compared to the scapula. As in the giraffe, both the juvenile and grown Spinophorosaurus individual would not have been able to reach the ground just by lowering their necks, and possibly splayed their forelimbs for drinking. While sleeping, giraffes bend their necks sideways against the body. Although the vertebral articulations would have been flexible enough for such bending in Spinophorosaurus, it might have been prohibited by the elongated cervical ribs.[15]
Paleoçevre

Spinophorosaurus is known from the Irhazer Shale of Niger, which has been determined to represent the base of the Irhazer Group (based on the presence of kırmızı kil Bu nasıl interbedded sand beds with traces of subaerial exposure and dinozor ayak izleri ), hangisi stratigrafik olarak below the outcrops of the Tegama Grubu. The Irhazer Group was earlier considered Jurassic to earliest Cretaceous in age, as the overlying Tiourarén Formation was once considered to be of Lower Cretaceous age. Subsequent studies have found the latter formation to be Middle Jurassic to early Late Jurassic. The stratigraphical and phylogenetic position of Spinophorosaurus (compared with other basal sauropods from around the world) indicates it could be Middle Jurassic (Bajocian –Batoiyen ) yaş. Since the strata of the Irhazer Group cannot yet be directly dated, it might be as old as Lower Jurassic. The lower limit is defined by the underlying Agadez Group, hangisi Üst Triyas in age (based on the presence of Chirotherium fosillerin izini sürmek ).[1][36]
Spinophorosaurus skeletons were discovered in a massive to finely laminated red siltstone, whose matris contained some carbonate. The fossils were recovered from the upper half of this several metres-thick siltstone layer. The layers there are subhorizontal, bearing minor faults, and commonly contain palaeosoils and carbonate at the top of the unity (about a metre above the level with the skeletons).[1] Sediments of the Irhazer Group were deposited under conditions that were Fluvio -göl (associated with rivers and lakes), during a time when the Iullemmeden Havzası was part of a great river-valley system, which was connected with the Proto-Atlantik coast of Gondwana through adjacent basins.[36][37]

Four theropod teeth were found closely associated with the Spinophorosaurus holotype (by a vertebra, pubis, and in the acetabulum); three had similarities with Megalosauridae ve Allosauridae, while the fourth belongs to what may be one of the earliest known members of Spinosauridae. The fact that the teeth are poorly preserved while the Spinophorosaurus skeleton is well-preserved indicates that the teeth were transported before being buried with the sauropod.[38] Dinosaur tracks were found some hundred metres away from the Spinophorosaurus site, including a trackway of six footprints from a mid-sized sauropod as well as 120 well-preserved didactyl (two-toed) theropod footprints. The didactyl tracks were originally thought to have been left by paravyan theropods (such as dromaeosaurs or Troodontidler, which have two weight-bearing toes), but were subsequently interpreted as having been produced by swimming theropods (explaining why one toe did not leave a trace).[36][39]
Referanslar
- ^ a b c d e f g h ben j k l m n Ö p q r s t sen v w x y Remes, K.; Ortega, F .; Fierro, I.; Joger, U.; Kosma, R.; Marín Ferrer, J. M.; Ide, O. A.u; Maga, A .; Farke, A. A. (2009). "A new basal sauropod dinosaur from the Middle Jurassic of Niger and the early evolution of Sauropoda". PLOS ONE. 4 (9): e6924. Bibcode:2009PLoSO...4.6924R. doi:10.1371/journal.pone.0006924. PMC 2737122. PMID 19756139.
- ^ a b c d e f g h ben j k l Joger, U.; Kosma, R.; Krüger, F. J. (2009). Projekt Dino: Die Entdeckungsgeschichte neuer Dinosaurier in Niger, Afrika (Almanca'da). Braunschweig: Cargo Verlag. ISBN 978-3-938693-17-9.
- ^ Ortega, F .; Fierro, I.; Chiappe, L.; et al. (2009). Paldes project and the vertebrate paleontology heritage of Niger country. First International Congress on North African Vertebrate Palaeontology (NAVEP1). 25–27 May 2009, Marrakech – Morocco. s. 49–51.
- ^ Joger, U. "The place of discovery and the excavation of 2005". Google Arts & Culture. Arşivlendi 20 Nisan 2019 tarihinde orjinalinden. Alındı 20 Nisan 2019.
- ^ Joger, U. "A discovery site of tracks". Google Arts & Culture. Arşivlendi 20 Nisan 2019 tarihinde orjinalinden. Alındı 20 Nisan 2019.
- ^ Joger, U. "Excavation tour 2007 – Start of the journey". Google Arts & Culture. Arşivlendi 20 Nisan 2019 tarihinde orjinalinden. Alındı 20 Nisan 2019.
- ^ Joger, U. (2018). "BRAUNSCHWEIG: Staatliches Naturhistorisches Museum Braunschweig". In Beck, L. A. (ed.). Zoological Collections of Germany. Natural History Collections. Springer. pp. 183–196. doi:10.1007/978-3-319-44321-8_15. ISBN 978-3-319-44321-8.
- ^ Kosma, R. "Oh no, where's our dino?". Google Arts & Culture. Arşivlendi 20 Nisan 2019 tarihinde orjinalinden. Alındı 20 Nisan 2019.
- ^ Witzmann, F.; Hampe, O.; Rothschild, B. M .; Joger, U.; Kosma, R.; Schwarz, D.; Asbach, P. (2016). "Subchondral cysts at synovial vertebral joints as analogies of Schmorl's nodes in a sauropod dinosaur from Niger". Omurgalı Paleontoloji Dergisi. 36 (2): e1080719. doi:10.1080/02724634.2016.1080719. S2CID 86187185.
- ^ Kosma, R. (2018). "The Paleontological Collection of the State Museum of Natural History (SNHM) in Braunschweig, Germany". In Beck, L. A.; Joger, U. (eds.). Paleontological Collections of Germany, Austria and Switzerlan. Natural History Collections. Springer. sayfa 81–92. doi:10.1007/978-3-319-77401-5_9. ISBN 978-3-319-77401-5.
- ^ Ritter, A. "Reconstruction of the dinosaur skeleton". Google Arts & Culture. Arşivlendi 20 Nisan 2019 tarihinde orjinalinden. Alındı 20 Nisan 2019.
- ^ a b Vidal, D .; Aberasturi, A.; Mocho, P.; Ortega, F .; Sanz, J. L. (2016). "Assembling a virtual Spinophorosaurus skeleton: what can it teach us about the evolution of eusauropods?". VII Jornadas Internaciones sobre Paleontología de Dinosaurios y su Entorno. Salas de los Infantes, Burgos. Burgos. s. 147–148.
- ^ "El MUPE expone por primera vez una vértebra del cuello de Spinophorosaurus nigerensis". AQUÍ en Elche (ispanyolca'da). 2018. Alındı 15 Haziran 2019.
- ^ García-Martínez, D.; Vidal, D .; Ortega, F. (2018). Using 3D geometric morphometrics to estimate missing vertebrae in a Spinophorosaurus dorsal spine (Middle Jurassic, Niger). 16th European Association of Vertebrate Paleontologists Meeting, Caparica (Portugal). European Association of Vertebrate Palaeontologists. s. 75.
- ^ a b c Vidal, Daniel; Mocho, Pedro; Páramo, Adrián; Sanz, José Luis; Ortega, Francisco (2020-01-13). "Ontogenetic similarities between giraffe and sauropod neck osteological mobility". PLOS ONE. 15 (1): –0227537. Bibcode:2020PLoSO..1527537V. doi:10.1371/journal.pone.0227537. ISSN 1932-6203. PMC 6957182. PMID 31929581. Alındı 2020-01-13.
- ^ Timmons, J. (2018). "Fossil discoveries in Niger with Dr. Ralf Kosma". Mostly Mammoths, Mummies and Museums. Arşivlendi 2 Mart 2019 tarihinde orjinalinden. Alındı 24 Şubat 2019.
- ^ Holtz, T. R. (2011). "Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix" (PDF). Alındı 13 Ocak 2012.
- ^ a b Tschopp, E .; Mateus, O. (2013). "Clavicles, interclavicles, gastralia, and sternal ribs in sauropod dinosaurs: new reports from Diplodocidae and their morphological, functional and evolutionary implications". Anatomi Dergisi. 222 (3): 321–340. doi:10.1111/joa.12012. PMC 3582252. PMID 23190365.
- ^ a b c Mocho, P.; Vidal, D .; Aberasturi, A.; Kosma, R.; Ortega, F. (2018). New information about the appendicular skeleton of Spinophorosaurus nigerensis (Middle Jurassic, Niger). 16th European Association of Vertebrate Paleontologists Meeting, Caparica (Portugal). European Association of Vertebrate Palaeontologists. s. 124.
- ^ Blázquez, A. P.; Ortega, F. (2012). A probable juvenile Spinophorosaurus nigerensis (Sauropoda) from the Middle Jurassic of Niger. 10th European Association of Vertebrate Paleontologists Meeting, At Teruel (España).
- ^ Paul, G. S. (2016). Princeton Dinozorlar Saha Rehberi. Princeton University Press. s. 196. ISBN 978-0-691-16766-4.
- ^ Vidal, D .; Mocho, P.; Aberasturi, A.; Sanz, J. L.; Ortega, F. (2020). "High browsing skeletal adaptations in Spinophorosaurus reveal an evolutionary innovation in sauropod dinosaurs". Bilimsel Raporlar. 10 (1): 6638. Bibcode:2020NatSR..10.6638V. doi:10.1038/s41598-020-63439-0. PMID 32313018. S2CID 215819745.
- ^ a b c Knoll, F.; Witmer, L. M .; Ortega, F .; Ridgely, R. C.; Schwarz-Wings, D. (2012). Farke, A. A. (ed.). "The braincase of the basal sauropod dinosaur Spinophorosaurus and 3D reconstructions of the cranial endocast and inner ear". PLOS ONE. 7 (1): e30060. Bibcode:2012PLoSO...730060K. doi:10.1371/journal.pone.0030060. PMC 3260197. PMID 22272273.
- ^ Taylor, M. P. (2015). Almost all known sauropod necks are incomplete and distorted. PeerJ Hazır Baskılar. doi:10.7287/peerj.preprints.1418v1.
- ^ Brett-Surman, M.K.; Holtz, Jr., Thomas R.; Farlow, James O. (2012). "The Osteology of the Dinosaurs". The Complete Dinosaur, Second Edition. Indiana University Press. s. 135–149. ISBN 978-0-253-00849-7.
- ^ a b Mocho, P.; Ortega, F .; Aberasturi, A.; Escaso, F. (2013). "Spinophorosaurus (Sauropoda), a new look inside eusauropod evolution". Abstract Book of the VI International Symposium About Dinosaurs Paleontology and Their Environment. Salas de los Infantes Burgos. sayfa 89–90.
- ^ Apesteguía, S. (2005). "Evolution of the hyposphene-hypantrum complex within Sauropoda". In Tidwell, V.; Carpenter, K. (editörler). Thunder-Lizards: The Sauropodomorph Dinosaurs. Bloomington ve Indianapolis: Indiana University Press. pp.248 –267. ISBN 978-0-253-34542-4.
- ^ a b Vidal, D. C.; Ortega, F. (2015). The specialized tail of Spinophorosaurus nigerensis (Sauropoda. Middle Jurassic) and the osteological limits on its range of motion. 63rd SVPCA (Symposium for Vertebrate Palaeontology and Comparative Anatomy), At Southampton, England.
- ^ a b c Nair, J. P.; Salisbury, S. W. (2012). "New anatomical information on Rhoetosaurus brownei uzun adam, 1926, a gravisaurian sauropodomorph dinosaur from the Middle Jurassic of Queensland, Australia". Omurgalı Paleontoloji Dergisi. 32 (2): 369. doi:10.1080/02724634.2012.622324. S2CID 85317450.
- ^ a b Holwerda, F. M.; Pol, D. (2018). "Phylogenetic analysis of Gondwanan basal eusauropods from the Early-Middle Jurassic of Patagonia, Argentina". İspanyol Paleontoloji Dergisi. 33 (2): 289. doi:10.7203/sjp.33.2.13604.
- ^ Bronzati, M .; Benson, R. B. J .; Rauhut, O. W. M .; Mannion, P. (2018). "Rapid transformation in the braincase of sauropod dinosaurs: integrated evolution of the braincase and neck in early sauropods?". Paleontoloji. 61 (2): 289–302. doi:10.1111/pala.12344.
- ^ a b Xing, L .; Miyashita, T .; Currie, P. J .; You, H.; Zhang, J .; Dong, Z. (2015). "A new basal eusauropod from the Middle Jurassic of Yunnan, China, and faunal compositions and transitions of Asian sauropodomorph dinosaurs". Acta Palaeontologica Polonica. 60 (1): 145–155. doi:10.4202 / app.2012.0151. S2CID 59143277.
- ^ Jentgen-Ceschino, C.; Stein, K. (2018). Cases of pathological bone growth in Isanosaurus ve Spinophorosaurus (Sauropoda). 5th International Palaeontological Congress – Pierre and Marie Curie Campus of Sorbonne University; National Museum of Natural History, Paris, France. doi:10.13140/RG.2.2.10914.27840.
- ^ Fronimos, J. A.; Wilson, J. A. (2017). "Neurocentral suture complexity and stress distribution in the vertebral column of a sauropod dinosaur". Ameghiniana. 54 (1): 36–49. doi:10.5710/AMGH.05.09.2016.3009. S2CID 132983807.
- ^ Vidal, D. C. (2018). Could sauropods perform a 'cloacal kiss´? Evidence of mating capabilities capabilities from a virtual Spinophorosaurus. XVI EJIP, At Zarautz.
- ^ a b c Mudroch, A.; Richter, U.; Joger, U.; Kosma, R.; Idé, O.; Maga, A .; Farke, A. A. (2011). "Didactyl tracks of paravian theropods (Maniraptora) from the ?Middle Jurassic of Africa". PLOS ONE. 6 (2): e14642. Bibcode:2011PLoSO...614642M. doi:10.1371/journal.pone.0014642. PMC 3038851. PMID 21339816.
- ^ Moody, R. T. J. (1997). "The Iullemmeden basin". In Selley, R. C. (ed.). Sedimentary Basins of the World. 3. Elsevier Science. s. 89–103. doi:10.1016/S1874-5997(97)80008-1. ISBN 978-0444825711.
- ^ Serrano-Martínez, A .; Vidal, D .; Sciscio, L .; Ortega, F .; Knoll, F. (2015). "Isolated theropod teeth from the Middle Jurassic of Niger and the early dental evolution of Spinosauridae". Acta Palaeontologica Polonica: 403–415. doi:10.4202 / app.00101.2014. S2CID 53331040.
- ^ Lockley, M. G.; Harris, J. D .; Li, R .; Xing, L. D.; van der Lubbe, T. (2016). "Two-toed tracks through time: on the trail of "raptors" and their allies". In P. L. Falkingham; Da. Marty; A. Richter (eds.). Dinosaur Tracks – The Next Steps. Bloomington ve Indianapolis: Indiana University Press. pp. 183–200. ISBN 978-0-253-02102-1.
Dış bağlantılar
- The Dino Project – Google Arts & Culture photo series about the Braunschweig Museum excavations in Niger
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||












