Schistosoma mansoni - Schistosoma mansoni
| Schistosoma mansoni | |
|---|---|
 | |
| Çiftleşen şistozomlar | |
| bilimsel sınıflandırma | |
| Krallık: | |
| Şube: | |
| Sınıf: | |
| Sipariş: | |
| Aile: | |
| Cins: | |
| Türler: | S. mansoni |
| Binom adı | |
| Schistosoma mansoni Sambon, 1907 | |

Schistosoma mansoni bir su kaynaklı insan paraziti ve grubuna aittir kan parazitleri (Şistozom). Yetişkin kan damarlarında yaşar (mezenterik damarlar ) insan bağırsağına yakın. Bağırsağa neden olur şistozomiyaz (benzer S. japonicum, S. mekongi, S. guineensis, ve S. intercalatum ). Klinik belirtilere yumurtalar neden olur. Dünyada schistosomiasis'in önde gelen nedeni olarak, insanlarda en yaygın görülen parazittir. Olarak sınıflandırılır ihmal edilmiş tropikal hastalık. 2016 itibariyle 206,5 milyon kişide şistozomiyaz var ve S. mansoni ana parazittir. Afrika, Orta Doğu, Karayipler, Brezilya, Venezuela ve Surinam'da bulunur.[1]
Diğer şanslardan farklı olarak (trematodlar ) cinsiyetlerin ayrı olmadığı (monoecious ), şistozomlar yetişkinlerin erkek ve kadınlara bölünmesi bakımından benzersizdir, bu nedenle (ikievcikli ). Bununla birlikte, iki yetişkin kalıcı bir ortaklık içinde yaşıyor, bu durum kopulada; bunun için olarak kabul edilirler Hermafroditler. Şistozomların yaşam döngüsü iki konakçı içerir: insanlar kesin ana bilgisayarlar parazitin eşeyli üremeye uğradığı yer ve bir dizi eşeysiz üremenin gerçekleştiği ara konakçı olarak salyangozlar. S. mansoni cinsin tatlı su salyangozlarının bulunduğu su yoluyla bulaşır. Biyomfalarya ara konaklar olarak hareket eder. Larvalar suda yaşayabilir ve doğrudan deriye nüfuz ederek konakçıları enfekte edebilir. Enfeksiyonun önlenmesi, iyileştirilmiş sanitasyon ve salyangozları öldürmek. Enfeksiyon ile tedavi edilir prazikuantel.
S. mansoni ilk önce tarafından not edildi Theodor Maximillian Bilharz 1851'de Mısır'da S. haematobium. Sör Patrick Manson 1902'de benzersiz bir tür olarak tanımladı. Louis Westenra Sambon isim verdi Schistosomum mansoni 1907'de Manson onuruna.[2][3][4]
Yapısı
Yetişkin
Şistozomlar, diğer trematodların aksine uzun ve silindirik kurtlardır. Erkek S. mansoni yaklaşık 1 cm uzunluğundadır (0,6-1,1 cm)[5] ve 0.1 cm genişliğindedir. Beyazdır ve huni şeklindedir. oral enayi ön ucunda ve ardından ikinci bir pediküllü ventral emici. Solucanın dış kısmı, membranokaliks olarak bilinen dış tabaka olarak sürekli yenilenen ve sürekli olarak dökülen çift çift tabakadan oluşur.[6] Tegument, çok sayıda küçük tüberküller. Vantuzların iç kısımlarında ve etrafındaki düğmelerde küçük dikenler vardır. Erkek genital aparatı dorsal olarak yerleştirilmiş 6 ila 9 testis kitlesinden oluşur. Her testiste başlayan, jinekoforik kanalın başlangıcında yer alan bir rezervuar olan seminal veziküle genişleyen tek bir ertelemeye bağlanan bir ertelemeli kanal vardır. Kopula, erkek ve dişi genital açıklıkların kapatılması yoluyla gerçekleşir.[7]
Dişinin silindirik bir gövdesi vardır, erkeklerinkinden daha uzun ve daha incedir (1,2 ila 1,6 cm uzunluğunda ve 0,016 cm genişliğinde). Genel bir yuvarlak kurt görünümündedir. Dişi parazit daha koyu ve gri görünüyor. Daha koyu renk, bir pigmentin varlığından kaynaklanmaktadır (hemozoin ) sindirim tüpünde. Bu pigment, kanın sindirilmesinden elde edilir. yumurtalık uzamış ve hafif lobludur ve vücudun ön yarısında bulunur. Uterus tüpü ile devam eden ootipe kısa bir yumurta kanalı iletilir. Bu tüpte 1 ila 2 yumurta (nadiren 3 ila 4) bulmak mümkündür ancak aynı anda ootipte sadece 1 yumurta gözlenir. Genital gözenek ventral olarak açılır. Vücudun arka üçte ikisinde vittelojenik bezler ve bunlarla birleşen sargı kanalları bulunur. yumurta kanalı ootipe ulaşmadan biraz önce.
Sindirim tüpü, solucanın ön ucunda, oral emicinin dibinde başlar. Sindirim tüpü bir yemek borusu, iki kola (sağ ve sol) bölünen ve tek bir çekum. Bağırsaklar kör bir şekilde biter, yani yoktur anüs.
Yumurta
Yumurtalar oval şekilli olup, 115–175 µm uzunluğunda ve 45–47 µm genişliğinde ve ortalama ~ 150 µm çapındadır. Bir tarafta daha geniş tabana doğru dikenleri, yani yanal dikenleri işaret etmişlerdir. Bu, önemli bir tanı aracıdır çünkü birlikte enfeksiyon S. haematobium (terminal dikenli yumurtalara sahip olmak) yaygındır ve bunları ayırt etmek zordur.[8] Yumurtalar suya salındığında çoğu olgunlaşmamış ve yumurtadan çıkmaması için döllenmemiş durumdadır. Yumurtaların çapı 160 µm'den büyük olduğunda, yumurtadan da çıkmazlar.[9][10]
Larva
Miracidium (Yunanca μειράκιον kelimesinden, Meirakion, yani gençlik) armut şeklindedir ve yaşlandıkça yavaş yavaş uzar. Yaklaşık 136 μm uzunluğunda ve 55 μm genişliğindedir. Gövde, epidermal çıkıntılarla ayrılmış anükleat epidermal plakalarla kaplıdır. Epidermal hücreler çok sayıda saç benzeri kirpikler vücut yüzeyinde. 17–22 epidermal hücre vardır. Epidermal plaka, yalnızca çok sayıda duyusal organel içeren apikal papilla veya terebratoryum denilen aşırı anteriorda yoktur.[11] İç gövdesi neredeyse tamamen glikojen partikülleri ve veziküllerle doludur.[12]
Cercaria, klasik olarak furcae (çatal için Latince) olarak adlandırılan karakteristik çatallı bir kuyruğa sahiptir; dolayısıyla adı (Yunanca κέρκος kelimesinden türetilmiştir, Kerkos, kuyruk anlamına gelir). Kuyruk oldukça esnektir ve çarpması cercaria'yı suda iter.[13] Yaklaşık 0.2 mm uzunluğunda ve 47 μm genişliğindedir, ana gövdeye biraz gevşek bir şekilde tutturulmuştur. Gövde armut şeklindedir ve 0.24 mm uzunluğunda ve 0.1 mm genişliğindedir.[14] Gövdesi tamamen omurga ile kaplıdır. Apekste göze çarpan bir oral enayi var. Beslenmeyen bir larva olarak, ayrıntılı bir Sindirim organları, sadece yemek borusu farklıdır. Ventral emici bölgesinde oral emiciye yanal olarak bağlanan üç çift müsin bezi vardır.[15][16]
Fizyoloji
Beslenme ve beslenme
Gelişen Schistosoma mansoni Erkek ve dişilerin cinsel olarak eşleşmesinden önce kesin konakçılarını enfekte eden solucanlar, cercariae'den yetişkinlere doğru düzgün bir şekilde gelişmek için bir besin kaynağına ihtiyaç duyar. Gelişmekte olan parazitler, besin maddelerine erişmek için konakçı kırmızı kan hücrelerini parçalamaktadır ve ayrıca atıklarından kendi mantarlarını tespit etmesi zor hale getirmektedir; Kan hücrelerinin içerdiği hemoglobin ve amino asitler solucan tarafından proteinleri oluşturmak için kullanılabilir.[17] Hemoglobin hücre içinde sindirilirken, tükürük bezi enzimleri tarafından başlatılırken, demir atıkları ürünleri solucanlar tarafından kullanılamaz ve tipik olarak yetersizlik yoluyla atılır.[18]
Kasschau vd. (1995), sıcaklık ve pH'ın gelişme yeteneği üzerindeki etkisini test etti. S. mansoni kırmızı kan hücrelerini parçalamak için.[17] Araştırmacılar, parazitlerin besinleri için kırmızı kan hücrelerini en iyi 5,1 pH ve 37 ° C sıcaklıkta yok edebildiklerini buldular.[17]
Hareket
S. mansoni yaşam döngüsünün başlıca iki aşamasında lokomotiftir: cercariae, insan konukçularının epidermisini bulmak için bir tatlı su kütlesinde serbestçe yüzerken ve gelişmekte olan ve tam teşekküllü yetişkinler olarak, enfeksiyon üzerine birincil konukçularının içinden göç ederek.[18] Cercariae, kesin konakçılarının derisindeki yağ asitlerinin varlığından etkilenir ve parazit, cilde doğru ilerlemek için tatlı su ortamındaki ışık ve sıcaklık değişikliklerine yanıt verir.[19] Ressurreicao vd. (2015), çeşitli protein kinazların, parazitin ortamında gezinme ve nüfuz edilebilir bir konakçı yüzeyini bulma yeteneğindeki rollerini test etti.[19] Hücre dışı sinyalle düzenlenen kinaz ve protein kinaz C'nin her ikisi de orta sıcaklık ve ışık seviyelerindeki değişikliklere yanıt verir ve parazit konak yüzeyinin tanınmasıyla ilişkili p38 mitojenle aktive edilmiş protein kinazın uyarılması, konakçı epidermisini bozan bir glandüler sekresyonla sonuçlanır, ve parazitin konakçıya girmesine izin verir.
Parazitin sinir sistemi, vücudun her yüzeyine yayılan çift loblu ganglionlar ve birkaç sinir kordunu içerir; serotonin, sinir sistemi boyunca yaygın olarak dağılan bir vericidir ve sinir alımında ve hareketliliği uyarmada önemli bir rol oynar.[20]
Yaşam döngüsü

Orta düzeyli ev sahibi
İnsanın yaşadığı parazitin yumurtaları dışkıdan ve suya salındıktan sonra, Miracidium yumurtadan çıkar. Yumurtadan çıkma sıcaklığa, ışığa ve dışkının suyla seyreltilmesine tepki olarak gerçekleşir. Miracidium uygun bir tatlı su salyangozu cinse ait Biyomfalarya. Güney Amerika'da, ana ara ev sahibi Biyomphalaria glabrata, süre B. straminea ve B. tenagophila daha az yaygındır.[21] Kara salyangozu Achatina fulica 2010 yılında Venezuela'da ev sahibi olarak hareket ettiği bildirildi.[22] Afrika'da, B. glabratra, B. pfeifferi, B. choanomphala ve B. sudanica ev sahibi olarak hareket etmek;[23] ancak Mısır'da ana salyangoz konağı B. alexandrina.[24]
Miracidia, salyangozun yumuşak dokusuna doğrudan nüfuz eder. Salyangozun içinde kirpikleri kaybederler ve anne sporokistlere dönüşürler. Sporokistler, eşeysiz üremeyle hızla çoğalırlar ve her biri çok sayıda yavru sporokist oluşturur. Kızı sporokistler, salyangozun karaciğere ve gonadlarına hareket eder ve burada daha fazla büyümeye başlarlar.[25] 2-4 hafta içinde metamorfoza uğrarlar ve çatal kuyruklu serkaryalara yol açarlar. Işıkla uyarılan yüzlerce cercaria, salyangozdan suya nüfuz eder.[26]
Kesin ana bilgisayar
Sercaria, gün ışığında salyangozdan çıkar ve dallanmış kuyruklarının yardımıyla kendilerini suda iter ve aktif olarak son konaklarını ararlar. Suda 12 saate kadar yaşayabilirler ve maksimum bulaşıcılıkları ortaya çıktıktan sonra 1 ila 9 saat arasındadır.[27] İnsanı tanıdıklarında cilt çok kısa sürede nüfuz ederler. Bu üç aşamada meydana gelir, cilde ilk bağlanma, ardından uygun bir penetrasyon bölgesi arayan cilt üzerinde sürünme, genellikle bir saç folikülü ve nihayet cildin içine nüfuz etmesi epidermis kullanma sitolitik serkaryal post-asetabular, ardından pre-asetabulardan sekresyonlar bezler. Penetrasyonda, cercaria'nın başı endoparazitik bir duruma dönüşür. larva, şistozomül. Her şistozomül ciltte birkaç gün geçirir ve ardından dermalden başlayarak dolaşıma girer. lenfatik ve venüller. Burada kanla beslenirler, kemiği kusarlar. hemozoin.[28] Şistozomül, akciğerler (Penetrasyondan 5-7 gün sonra) ve ardından kan dolaşımının sol tarafından kalp hepatoportal dolaşıma (> 15 gün), karşı cinsten bir partnerle karşılaşırsa, cinsel olarak olgun bir yetişkin haline gelir ve çift mezenterik damarlara göç eder.[29] Bu tür eşleşmeler tek eşli.[30]
Erkek şistozomlar, bir dişinin varlığında veya yokluğunda normal olgunlaşma ve morfolojik gelişime uğrar, ancak tek cinsiyetten erkekler arasında biseks ile karşılaştırıldığında davranışsal, fizyolojik ve antijenik farklılıklar bildirilmiştir. Öte yandan, dişi şistozomlar bir erkek olmadan olgunlaşmaz. Tek cinsiyetli enfeksiyonlardan kaynaklanan dişi şistozomlar az gelişmiştir ve olgunlaşmamış bir üreme sistemi sergiler. Dişi solucanın olgunlaşması, olgun erkeğin varlığına bağlı görünse de, dişi büyümesi ve üreme gelişimi için uyaranlar birbirinden bağımsız görünmektedir.
Yetişkin dişi solucan, erkeğin ventral yüzeyinin bir modifikasyonu olan ve bir oluk oluşturan yetişkin erkek solucanın jinekoforik kanalında bulunur. Eşleştirilmiş solucanlar, kan akışına karşı mezenterik dolaşımdaki son nişlerine hareket eder ve burada yumurta üretimine başlarlar (> 32 gün). S. mansoni parazitler ağırlıklı olarak konağın kalın bağırsağı ve çekal bölgesini çevreleyen küçük alt mezenterik kan damarlarında bulunur. Her dişi günde yaklaşık 300 yumurta bırakır (her 4.8 dakikada bir yumurta). endotelyal venöz astar kılcal damar duvarlar.[31] Dişi şistozomların vücut kütlesinin çoğu üreme sistemine ayrılmıştır. Dişi, her gün neredeyse kendi vücut kuru ağırlığının eşdeğerini yumurtaya çevirir. Yumurtalar lümen ev sahibinin bağırsaklar dışkı ile çevreye salınır.
Genetik şifre
Schistosoma mansoni 8 çift var kromozomlar (2n = 16) —7 otozomal çift ve 1 cinsiyet çifti. Dişi şistozom heterogametiktir veya ZW'dir ve erkek homojen veya ZZ'dir. Cinsiyet, zigot bir kromozomal mekanizma ile. genetik şifre % 34 GC içeriği,% 4-8 yüksek tekrarlı dizi,% 32-36 orta tekrarlı dizi ve% 60 tek kopya dizisi ile yaklaşık 270 MB'dir. En az% 30 tekrarlayan çok sayıda yüksek veya orta düzeyde tekrarlayan öğe tanımlanır DNA. Kromozomların boyutu 18 ila 73 MB arasındadır ve boyut, şekil ve C bantlamasıyla ayırt edilebilir.[32]
2000 yılında, Schistosome'un ilk BAC kütüphanesi oluşturuldu.[33] Haziran 2003'te, a ~ 5x tam genom av tüfeği sıralaması proje başlatıldı Sanger Enstitüsü.[34] Ayrıca 2003 yılında 163.000 EST (ifade edilen sıra etiketleri ) oluşturuldu (bir konsorsiyum tarafından yönetiliyor São Paulo Üniversitesi ) bu parazitin seçilmiş altı gelişim aşamasından, 31.000 birleştirilmiş sekans ve 14.000 gen tamamlayıcısının tahmini% 92'si ile sonuçlanır.[35]
2009'da her ikisinin de genomları S. mansoni ve S. japonicum her biri sırasıyla 11,809 ve 13,469 geni açıklayan yayınlanmıştır. S. mansoni genom, proteaz ailelerini ve lipid anabolizmasındaki eksiklikleri artırmıştır; parazitik adaptasyonuna atfedilir. Portease, invadolizin (konakçı penetrasyon) ve katepsin (kanla besleme) gen ailelerini içeriyordu.[36][37]
2012'de, gelişmiş bir S. mansoni Sadece 885 iskeleden ve bazların% 81'inden fazlasını kromozomlar halinde organize eden genom yayınlandı.[38]
Patoloji
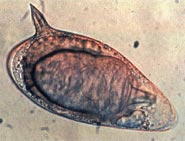
Konak dokulara yerleşebilen şistozom yumurtaları, şistozomiyaziste patolojinin başlıca nedenidir. Depolanan yumurtaların bir kısmı bağırsak duvarından geçerek dış ortama ulaşır; geri kalanı dolaşıma doğru süpürülür ve karaciğerin periportal yollarında filtrelenerek periportal fibroz ile sonuçlanır. İnsanlarda yumurtlamanın başlangıcı bazen ateşin başlamasıyla (Katayama ateşi) ilişkilendirilir. Bununla birlikte, bu "akut şistozomiyaz", hastalığın kronik formları kadar önemli değildir. İçin S. mansoni ve S. japonicum bunlar "bağırsak" ve "hepatik şistozomiyazdır", granülomlar sırasıyla bağırsak duvarına veya karaciğere sıkışmış yumurtaların etrafında. Hastalığın hepatik formu en önemlisidir, burada granülomlar fibroz karaciğer ve hepatosplenomegali ağır vakalarda. Belirtiler ve belirtiler, dokularda hapsolmuş yumurtaların sayısına ve konumuna bağlıdır. Başlangıçta, enflamatuar reaksiyon kolaylıkla tersine çevrilebilir. Hastalığın sonraki aşamalarında patoloji aşağıdakilerle ilişkilidir: kolajen birikim ve fibroz, sadece kısmen geri döndürülebilen organ hasarına neden olur.[39]
Granülom oluşumu, sert yumurta kabuğundaki mikroskobik gözeneklerden miracidium tarafından salgılanan antijenler tarafından başlatılır ve semptomlara yumurta antijenlerinin direkt etkisinden ziyade granüloma karşı bağışıklık tepkisi neden olur.[40] Yumurtaların etrafında oluşan granülomlar karaciğerdeki kan akışını bozar ve sonuç olarak portal hipertansiyon. Zamanla, teminat sirkülasyonu oluşur ve yumurtalar akciğerlere yayılır, burada daha fazla granüloma, pulmoner arterite ve daha sonra, cor pulmonale. Portal hipertansiyona katkıda bulunan bir faktör, portal damarların dalları etrafında gelişen Symmers fibrozudur. Bu fibroz, enfeksiyondan sadece yıllar sonra ortaya çıkar ve kısmen çözünebilir yumurta antijenleri ve bunlara tepki veren çeşitli bağışıklık hücrelerinden kaynaklandığı varsayılır.[41]
Son araştırmalar, granülom boyutunun, IL-13 Granülom oluşumunda ve granülom boyutunda önemli bir rol oynar. IL-13 reseptörü α 2 (IL-13Ra2), IL-13'ü yüksek afinite ile bağlar ve IL-13'ün etkilerini bloke eder. Bu nedenle, bu reseptör, şistozomiyazisin hastalığın akut aşamasından kronik (ve ölümcül) aşamasına ilerlemesini önlemede çok önemlidir. Farelere verilen sentetik IL-13Ra2, granülom boyutunda önemli azalmalara neden oldu ve IL-13Ra2'yi şistozomiyazda önemli bir hedef olarak ima etti.[42]
S. mansoni enfeksiyon genellikle viral hepatitin yanında ortaya çıkar. hepatit B virüsü (HBV) veya hepatit C virüsü (HCV). Bu, kronik viral hepatitin yaygın olduğu bölgelerde yüksek şistozomiyaz prevalansından kaynaklanmaktadır. Önemli bir faktör, kullanılan kapsamlı şistozomiyaz kontrol programları nedeniyle büyük enfeksiyon rezervuarının gelişmesiydi. intravenöz olarak yönetilen tartar emetik 1960'lardan beri.[39] Eş enfeksiyonun daha erken neden olduğu bilinmektedir karaciğer bozulması ve daha şiddetli hastalık.[43]
Konakçı bağışıklığından kaçınma
Yetişkin ve larva kurtları, konağın bağışıklık sisteminden kaçınarak konağın kan dolaşımı yoluyla göç ederler. Solucanlar, tegument, antioksidan proteinler ve konağa karşı savunma dahil olmak üzere bu kaçışa yardımcı olan birçok araca sahiptir. zar saldırı kompleksi (MAC).[44] Tegument solucanı kaplar ve antikorları barındırmak için fiziksel bir bariyer görevi görür ve Tamamlayıcı. Konakçı bağışıklık savunmaları süperoksit üretme yeteneğine sahiptir, ancak bunlar parazit tarafından üretilen antioksidan proteinler tarafından karşı saldırıya uğrar. Şistozomların dört süperoksit dismutazı vardır ve bu proteinlerin seviyeleri şistozom büyüdükçe artar. Antioksidan yollar ilk önce şistozomlar için bir tıkanma noktası olarak kabul edildi,[45] ve daha sonra diğer trematodlara ve sestodlara genişledi. Bu yolun, merkezi antioksidan enzimin farklı inhibitörleri ile hedeflenmesi tioredoksin glutatyon redüktaz (TGR) solucanların yaşayabilirliğinin azalmasına neden olur.[46] Çürüme hızlandırma faktörü (DAF) proteini parazit tegument üzerinde bulunur ve MAC oluşumunu bloke ederek konakçı hücreleri korur. Ek olarak, şistozomlar, güçlü MAC inhibitörleri olan altı insan CD59 homologuna sahiptir.[47]
Teşhis
Varlığı S. mansoni tarafından tespit edildi mikroskobik muayene dışkıda parazit yumurta. Bir boyama yöntemi adı verilen Kato-Katz dışkı muayenesi için teknik kullanılır. İçerir metilen mavisi -lekeli selofan gliserin veya cam slaytlara batırılmış.[48] Formalin-eter konsantrasyon tekniği (FECT) adı verilen biraz daha maliyetli bir teknik, daha yüksek doğruluk için genellikle doğrudan dışkı yayması ile birlikte kullanılır. Serolojik ve immünolojik testler de mevcuttur. Antikorlar ve antijenler kullanılarak kanda tespit edilebilir ELISA enfeksiyonu tanımlamak için. Yetişkin solucan antijenleri dolaylı olarak tespit edilebilir hemaglütinasyon tahlilleri (IHA'lar). Polimeraz zincirleme reaksiyonu (PCR) ayrıca parazit DNA'sını tespit etmek için kullanılır. İdrarda dolaşan katodik antijen (CCA), yanal akış immün kromatografik reaktif şeridi ve bakım noktası (POC) testleri ile test edilebilir.[49]
Yumurta tespiti ve immünolojik testler o kadar hassas değildir.[50] Polimeraz zincirleme reaksiyonu (PCR) tabanlı testler doğru ve hızlıdır.[50] Ancak, ekipmanın maliyeti ve bunları çalıştırmak için gereken teknik deneyim nedeniyle hastalığın yaygın olduğu ülkelerde sık kullanılmamaktadırlar.[50] Yumurtaları tespit etmek için bir mikroskop kullanmak, test kuyusu başına yaklaşık 0,40 ABD doları maliyetindedir. PCR, 2019 itibarıyla test başına yaklaşık 7 ABD dolarıdır.[51] Döngü aracılı izotermal amplifikasyon (LAMP) daha düşük maliyetli oldukları için incelenmektedir.[50] LAMP testi, 2019 itibarıyla ticari olarak mevcut değildir.[51]
Tedavi
İçin standart ilaç S. mansoni enfeksiyon 40 mg / kg dozunda prazikuanteldir. Oxamniquine ayrıca kullanılır.[52]
Epidemiyoloji
DSÖ 2016 raporuna göre, dünya çapında 206,5 milyon insan, farklı türler nedeniyle şistozomiyazis yaşıyor. Şistozom. 88 milyondan fazla kişiye tıbbi tedavi verildi. S. mansoni başlıca türdür. 55 ülkede endemiktir ve en çok Afrika, Orta Doğu, Karayipler, Brezilya, Venezuela ve Surinam'da yaygındır.[1][53] Schistosomiasis'in yaklaşık% 80-85'i Sahra altı Afrika'da bulunur. S. haematobium, S. intercalatum ve S. mansoni endemiktir. Yaklaşık 393 milyon Afrikalı, S. mansonibunların 55 milyonu her an enfekte. Nedeniyle yıllık ölüm S. mansoni yaklaşık 130.000'dir.[54] Afrika'nın farklı ülkelerindeki yaygınlık oranı: Kuzey Etiyopya'da% 73,9, Batı Etiyopya'da% 37,9, Nijerya'da% 56, Kenya'da% 60,5, Tanzanya'da% 64,3, Gana'da% 19,8 ve Fildişi Sahili'nde% 53,8'dir. .[55] Mısır'da Nil Deltası'nın kuzey ve doğu kesimlerindeki nüfusun% 60'ı ve Güney kesiminde sadece% 6'sı enfekte.[56]
S. mansoni genellikle fakir olan yerlerde bulunur sanitasyon. Parazitin fekal-oral olması nedeniyle aktarma insan atığı içeren su kütleleri olabilir bulaşıcı. Ara konak salyangoz türlerinin büyük popülasyonlarını içeren suyun enfeksiyona neden olma olasılığı daha yüksektir. Bu bölgelerde yaşayan küçük çocuklar, yüzme ve banyo yapma eğilimleri nedeniyle en büyük risk altındadır. Cercaria yetişkinlerden daha uzun süre enfekte sular.[57] Kimse seyahat yukarıda açıklanan alanlara ve maruz kalanlara kirlenmiş su, schistosomiasis riski altındadır.
Tarih
Ara konaklar Biyomfalarya salyangozların 95-110 milyon yıl önce Güney Amerika'da ortaya çıktığı tahmin edilmektedir. Ama parazitler Şistozom Asya kökenli. Afrika'da, öncü türler modern hale geldi S. mansoni ve S. haematobium yaklaşık 2–5 milyon yıl önce.[58] Ulaştı Mısır enfekte yoluyla köleler ve babunlar -den Bahis Ülkesi Muhtemelen V Hanedanı kadar erken dönemde gerçekleşen göçler aracılığıyla (c. 2494–2345 BCE).[59]
Alman bir doktor Theodor Maximillian Bilharz Kahire'de bir tıp fakültesi olan Kasr el-Aini Hastanesi'nde çalışırken 1851'de paraziti ilk keşfeden kişiydi. Bilharz, onları ölü askerlerin otopsilerinden kurtardı ve iki ayrı paraziti fark etti.[60] Onlardan birini şöyle tarif etti: Distomum hematobiyum (şimdi S. haematobium) 1852'de,[61] ama diğerini tanımlayamadı. Akıl hocasına yazdığı mektuplardan birinde Karl Theordor von Siebold, bazı yumurtaların terminal dikenlere sahip olmalarının farklı olduğunu, bazılarının ise yan dikenlerinin bulunduğunu belirtti.[62] Uçta dikenli yumurtalar, S. haematobiumyan dikenler ise yalnızca S. mansoni. Bilharz ayrıca yetişkin kurtların anatomisi ve ürettikleri yumurta sayısı bakımından farklı olduğunu belirtti.[63] 1856'da enfeksiyonun adı için bilharzia ve bilharziasis terimlerini tanıttı. Bir Alman zoolog David Friedrich Weinland cins adını düzeltti Şistozom 1858'de; hastalık adını schistosomiasis olarak tanıttı.[64]
Tür ayrımı ilk olarak Patrick Manson -de Londra Hijyen ve Tropikal Tıp Okulu. Manson, daha önce Batı Hint Adaları'na gönderilen bir sömürge memurunun dışkısında yanal dikenli yumurtalar tespit etti ve iki tür olduğu sonucuna vardı. Şistozom.[65] Yeni isimleri bir İtalyan-İngiliz doktor Louis Westenra Sambon verdi Schistosomum haematobium ve Schistosomum mansoni 1907'de, ikincisi Manson'u onurlandırmak için.[2] Sambon, bir erkek solucan kullanarak yalnızca kısmi açıklama yaptı. 1908'de Brezilyalı bir doktor Manuel Augusto Pirajá da Silva yan dikenli yumurtalar da dahil olmak üzere erkek ve dişi kurtların tam bir tanımını verdi.[66] Pirajá da Silva, üç otopsiden örnekler ve 20 dışkı incelemesinden yumurta aldı. Bahia.[67] Adını verdi S. americanum.[68] Tür kimliği 1907'de İngiliz parazitolog tarafından doğrulandı Robert Thomson Leiper,[65] belirli salyangoz konağın tanımlanması ve yumurta yapısının ayırt edilmesi, böylece yaşam döngüsünün oluşturulması.[69]
Referanslar
- ^ a b DSÖ (2017). "Schistosomiasis Bilgi formu". www.who.int. Dünya Sağlık Örgütü Medya Merkezi. Alındı 13 Aralık 2017.
- ^ a b Sambon, L.W. (1907). "İle ilgili açıklamalar Schistosomum mansoni". Tropikal Tıp ve Hijyen Dergisi. 10: 303–304.
- ^ Birch, CA (1974). "Schistosoma mansoni. Sör Patrick Manson, 1844–1922 ". Uygulayıcı. 213 (1277): 730–2. PMID 4156405.
- ^ Swanner, Yann A. Meunier; Michael Hole, Takudzwa Shumba & B.J.'nin (2014) katkılarıyla. Tropikal Hastalıklar: Tıp Doktorları ve Öğrenciler için Pratik Bir Kılavuz. Oxford: Oxford University Press, ABD. s. 40. ISBN 9780199997909.
- ^ Machado-Silva JR; Galvao C; Oliveira RMF; Presgrave AF; Gomes DC (1995). "Schistosoma mansoni sambon, 1907: Bazı Brezilya Suşlarının karşılaştırmalı morfolojik çalışmaları ". Rev. Inst. Med. Trop. Sao Paulo. 37 (5): 441–7. doi:10.1590 / s0036-46651995000500010. PMID 8729755.
- ^ Braschi S; Borges WC; Wilson RA (Eylül 2006). "Şistozom parçasının ve yüzey zarlarının proteomik analizi". Memórias do Instituto Oswaldo Cruz. 101 (Ek 1): 205–12. doi:10.1590 / S0074-02762006000900032. PMID 17308771.
- ^ Rey Luíz (1991). Parazitoloji. Rio de Janeiro, RJ: Editora Guanabara Koogan S.A. s. 351–62. ISBN 978-85-277-0189-1.
- ^ Hutchison, H.S. (1928). "Bilharziasis patolojisi". Amerikan Patoloji Dergisi. 4 (1): 1–16. PMC 2006716. PMID 19969774.
- ^ Xu, YZ; Dresden, MH (1989). "Schistosoma mansoni: yumurta morfolojisi ve kuluçka kabiliyeti ". Parazitoloji Dergisi. 75 (3): 481–483. doi:10.2307/3282615. JSTOR 3282615. PMID 2723933.
- ^ Xu, Yi-Zheng; Dresden, Marc H. (1990). "Şistozom yumurtalarının çıkması". Deneysel Parazitoloji. 70 (2): 236–240. doi:10.1016 / 0014-4894 (90) 90104-K. PMID 2105231.
- ^ Køie, Marianne; Frandsen, Flemming (1976). "Miracidium'un stereoscan gözlemleri ve erken sporokist Schistosoma mansoni". Zeitschrift für Parasitenkunde. 50 (3): 335–344. doi:10.1007 / BF02462978. PMID 997727. S2CID 8968526.
- ^ Cort, W.W. (1919). "İnsan şistozomlarının yumurtaları ve mirasidisi üzerine notlar". Zoolojide Kaliforniya Üniversitesi Yayınları. 18 (18): 509–519.
- ^ Krishnamurthy, Deepak; Katsikis, Georgios; Bhargava, Arjun; Prakash, Manu (2016). "Schistosoma mansoni cercariae, bir elastohidrodinamik bağlamadan yararlanarak verimli bir şekilde yüzer". Doğa Fiziği. 13 (3): 266–271. doi:10.1038 / nphys3924.
- ^ Faust, E.C. (1919). "Güney Afrika cercariae üzerine notlar". Parazitoloji Dergisi. 5 (4): 164–175. doi:10.2307/3271082. JSTOR 3271082.
- ^ Faust, CE (1920). "Schistosome larvalarının farklılaşması için kriterler". Parazitoloji Dergisi. 6 (4): 192–194. doi:10.2307/3270844. JSTOR 3270844.
- ^ Mohammed, A.S. (1931). "Serkaryanın salgı bezleri S. Haematobium ve S. Mansoni Mısır'dan". Tropikal Tıp ve Parazitoloji Yıllıkları. 26 (1): 7–22. doi:10.1080/00034983.1932.11684702.
- ^ a b c Kasschau MR, Byam-Smith MP, Gentry DS, Watson FN (1995). "Yetişkinlerde pH ve sıcaklığın hemoliz üzerindeki etkisi Schistosoma mansoni membranlar ". J. Exp. Zool. 271 (4): 315–22. doi:10.1002 / jez.1402710409. PMID 7722474.
- ^ a b Wilmer, Pat; Stone, Graham; Johnston, Ian (2005). Hayvanların Çevresel Fizyolojisi. Blackwell. pp.677–692. ISBN 9781405107242.
- ^ a b Ressurreição M, Kirk RS, Rollinson D, Emery AM, Page NM, Walker AJ (2015). "Duyusal Protein Kinaz Sinyali Schistosoma mansoni Cercariae: Barındırma Yeri ve İstila ". J. Infect. Dis. 212 (11): 1787–97. doi:10.1093 / infdis / jiv464. PMC 4633769. PMID 26401028.
- ^ Patocka N, Sharma N, Rashid M, Ribeiro P (2014). "Serotonin sinyali Schistosoma mansoni: serotonin ile aktive olan G proteinine bağlı bir reseptör, parazit hareketini kontrol eder ". PLOS Pathog. 10 (1): e1003878. doi:10.1371 / journal.ppat.1003878. PMC 3894222. PMID 24453972.
- ^ Carvalho Ode, D (1992). "Orta düzey konaklar Schistosoma mansoni Brezilya'da". Memórias do Instituto Oswaldo Cruz. 87 (Ek 4): 307–309. doi:10.1590 / s0074-02761992000800048. PMID 1343914.
- ^ (ispanyolca'da) Libora M., Morales G., Carmen S., Isbelia S. ve Luz A. P. (2010). "Primer hallazgo en Venezuela de huevos de Schistosoma mansoni y de otros helmintos de interés en salud pública, sunes en heces y secreción mucosa del molusco terrestre Achatina fulica (Bowdich, 1822). [Venezuela'da ilk bulgu Schistosoma mansoni Yumuşakçaların dışkı ve mukoza salgısında bulunan halk sağlığına ilgi duyan yumurtalar ve diğer helmintler Achatina fulica (Bowdich, 1822)]. Zootecnia Tropikal 28: 383–394. PDF[ölü bağlantı ].
- ^ Bustinduy, Amaya L .; Charles H., Kral (2014). "Schistosomiasis". Farrar, J; White, NJ (editörler). Manson Tropikal Hastalıkları (Yeni baskı). Philadelphia: Saunders [Künye]. s. 698–725. doi:10.1016 / B978-0-7020-5101-2.00091-1. ISBN 978-0-7020-5101-2.
- ^ Abou-El-Naga, IF (2013). "Biyomphalaria alexandrina Mısır'da: geçmiş, şimdi ve gelecek ". Biosciences Dergisi. 38 (3): 665–672. doi:10.1007 / s12038-013-9329-4. PMID 23938396. S2CID 16670424.
- ^ Ross, Allen; Inobaya, Marianette; Olveda, Remigio; Chau, Thao; Olveda, David (2014). "Şistozomiyazisin önlenmesi ve kontrolü: güncel bir bakış açısı". Tropikal Tıpta Araştırma ve Raporlar. 2014 (5): 65–75. doi:10.2147 / RRTM S44274. PMC 4231879. PMID 25400499.
- ^ Gryseels, Bruno (2012). "Schistosomiasis". Kuzey Amerika Bulaşıcı Hastalık Klinikleri. 26 (2): 383–397. doi:10.1016 / j.idc.2012.03.004. PMID 22632645.
- ^ Whitfield, PJ; Bartlett, A; Khammo, N; Clothier, RH (2003). "Yaşa bağlı sağkalım ve bulaşıcılık Schistosoma mansoni cercariae ". Parazitoloji. 127 (Pt 1): 29–35. doi:10.1017 / s0031182003003263. PMID 12885186.
- ^ Oliveira MF, d'Avila JC, Torres CR, vd. (Kasım 2000). "Hemozoin Schistosoma mansoni". Moleküler ve Biyokimyasal Parazitoloji. 111 (1): 217–21. doi:10.1016 / S0166-6851 (00) 00299-1. PMID 11087932.
- ^ Wilson, R.A. (2009). "Şistozom göçü ve yıpranma destanı". Parazitoloji. 136 (12): 1581–1592. doi:10.1017 / S0031182009005708. PMID 19265564.
- ^ Beltran S; Boissier J (Eylül 2008). "Schistosome monogamy: kim, nasıl ve neden?". Parazitolojide Eğilimler. 24 (9): 386–91. doi:10.1016 / j.pt.2008.05.009. PMID 18674968.
- ^ Loverde PT; Chen L (Kasım 1991). "Şistozom dişi üreme gelişimi". Parazitoloji Bugün. 7 (11): 303–8. doi:10.1016 / 0169-4758 (91) 90263-N. PMID 15463396.
- ^ "Schistosoma mansoni Genom Projesi ". Sanger Enstitüsü. Alındı 2007-06-14.
- ^ Le Paslier MC, Pierce RJ, Merlin F, vd. (Nisan 2000). "Yapısı ve karakterizasyonu Schistosoma mansoni bakteriyel yapay kromozom kütüphanesi ". Genomik. 65 (2): 87–94. doi:10.1006 / geno.2000.6147. PMID 10783255.
- ^ "Schistosoma mansoni Genom Projesi ". Genomik Araştırma Enstitüsü. Alındı 2007-06-14.
- ^ Verjovski-Almeida S; DeMarco R; Martins EA; Guimarães PE; Ojopi EP; Paquola AC; Piazza JP; Nishiyama MY Jr; Kitajima JP; Adamson RE; Ashton PD; Bonaldo MF; Coulson PS; Dillon GP; Farias LP; Gregorio SP; Ho PL; Leite RA; Malaquias LC; Marques RC; Miyasato PA; Nascimento AL; Ohlweiler FP; Reis EM; Ribeiro MA; Sá RG; Stukart GC; Soares MB; Gargioni C; Kawano T; Rodrigues V; Madeira AM; Wilson RA; Menck CF; Setubal JC; Leite LC; Dias-Neto E. (Ekim 2003). "Acoelomate insan parazitinin transkriptom analizi Schistosoma mansoni". Doğa Genetiği. 35 (2): 148–157. doi:10.1038 / ng1237. PMID 12973350. S2CID 11387995.
- ^ Berriman M; Haas BJ; LoVerde PT; Wilson RA; Dillon GP; Cerqueira GC; Mashiyama ST; Al-Lazikani B; Andrade LF; Ashton PD; Aslett MA; Bartholomeu DC; Blandin G; Caffrey CR; Coghlan A; Coulson R; TA Günü; Delcher A; DeMarco R; Djikeng A; Eyre T; Gamble JA; Ghedin E; İnsan; Hertz-Fowler C; Hirai H; Hirai Y; Houston R; Ivens A; Johnston DA; Lacerda D; Macedo CD'si; McVeigh P; Ning Z; Oliveira G; Overington JP; Parkhill J; Pertea M; Pierce RJ; Protasio AV; Bıldırcın MA; Rajandream MA; Rogers J; Sajid M; Salzberg SL; Stanke M; Tivey AR; Beyaz O; Williams DL; Wortman J; Wu W; Zamanian M; Zerlotini A; Fraser-Liggett CM; Barrell BG; El-Sayed NM. (Temmuz 2009). "Kan şansının genomu Schistosoma mansoni". Doğa. 460 (7253): 352–8. Bibcode:2009Natur.460..352B. doi:10.1038 / nature08160. PMC 2756445. PMID 19606141.
- ^ "Katil parazitlerin genlerinin kodu çözüldü". BBC haberleri. 16 Temmuz 2009. Alındı 2009-07-16.
- ^ Anna V. Protasio; Isheng J. Tsai; Anne Babbage; Sarah Nichol; Martin Hunt; Nishadi De Silva; Tim J.C. Anderson; Richard C. Clark; Claire Davidson; Gary P. Dillon; Nancy E. Holroyd; Philip T. LoVerde; Christine Lloyd; Jacquelline McQuillan; Guilherme Oliveira; Thomas D. Otto; Sophia J. Parker-Manuel; Michael A. Quail; R. Alan Wilson; Adhemar Zerlotini; David W. Dunne; Matthew Berriman. (Ocak 2012). "İnsan kanı şansının sistematik olarak geliştirilmiş yüksek kaliteli genomu ve transkriptomu Schistosoma mansoni". PLOS İhmal Edilen Tropikal Hastalıklar. 6 (1): 1455. doi:10.1371 / journal.pntd.0001455. PMC 3254664. PMID 22253936.
- ^ a b Elbaz, Tamer; Esmat, Gamal (2013). "Karaciğer ve Bağırsak Schistosomiasis: İnceleme". İleri Araştırmalar Dergisi. 4 (5): 445–452. doi:10.1016 / j.jare.2012.12.001. PMC 4293886. PMID 25685451.
- ^ Boros DL (Temmuz 1989). "İmmünopatoloji Şistozom mansoni enfeksiyonu ". Klinik Mikrobiyoloji İncelemeleri. 2 (3): 250–69. doi:10.1128 / cmr.2.3.250. PMC 358119. PMID 2504481.
- ^ Guimarães Cavalcanti, Marta; Marcello de Araujo-Neto, João; Mauro Peralta, José (2015). "Schistosomiasis: Karaciğer hastalığının klinik yönetimi". Klinik Karaciğer Hastalığı. 6 (3): 59–62. doi:10.1002 / cld.495. PMC 6490649. PMID 31040989.
- ^ Mentink-Kane MM, Cheever AW, Thompson RW, ve diğerleri. (Ocak 2004). "IL-13 reseptörü α 2, granülomatöz enflamasyonu aşağı modüle eder ve şistozomiyazda konakçı sağkalımını uzatır". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 101 (2): 586–90. Bibcode:2004PNAS..101..586M. doi:10.1073 / pnas.0305064101. PMC 327191. PMID 14699044.
- ^ Gasim, Gasim I; Bella, Abdelhaleem; Adam, Ishag (2015). "Schistosomiasis, hepatit B ve hepatit C ko-enfeksiyonu". Viroloji Dergisi. 12 (1): 19. doi:10.1186 / s12985-015-0251-2. PMC 4323254. PMID 25889398.
- ^ Wilson RA; Coulson PS (Eylül 2009). "Şistozomiyaza karşı bağışıklık efektör mekanizmaları: parazitin zırhında bir çatlak aramak". Parazitolojide Eğilimler. 25 (9): 423–31. doi:10.1016 / j.pt.2009.05.011. PMC 3686490. PMID 19717340.
- ^ AA dedi; Simeonov A; Thomas CJ; Inglese J; Austin CP; Williams DL (Nisan 2008). "Oksadiazollerin şistozomiyazisin kontrolüne yol açan yeni ilaç olarak tanımlanması". Nat. Orta. 14 (4): 407–12. doi:10.1038 / nm1737. PMC 2700043. PMID 18345010.
- ^ Ross F, Hernández P, Porcal W, ve diğerleri. (2012). "Sestod ve trematod parazitlerini öldüren tioredoksin glutatyon redüktaz inhibitörlerinin tanımlanması". PLOS ONE. 7 (4): e35033. Bibcode:2012PLoSO ... 735033R. doi:10.1371 / journal.pone.0035033. PMC 3335049. PMID 22536349.
- ^ Wilson, R. Alan; Coulson Patricia S. (2009). "Şistozomiyaza karşı bağışıklık efektör mekanizmaları: parazitin zırhında bir çatlak aramak". Parazitolojide Eğilimler. 25 (9): 423–431. doi:10.1016 / j.pt.2009.05.011. PMC 3686490. PMID 19717340.
- ^ Gray, D. J .; Ross, A. G .; Li, Y.-S .; McManus, D.P. (2011). "Şistozomiyazisin teşhisi ve tedavisi". BMJ. 342: d2651. doi:10.1136 / bmj.d2651. PMC 3230106. PMID 21586478.
- ^ Utzinger, J .; Becker, S.L .; van Lieshout, L .; van Dam, G.J .; Knopp, S. (2015). "Schistosomiasis'te yeni teşhis araçları". Klinik Mikrobiyoloji ve Enfeksiyon. 21 (6): 529–542. doi:10.1016 / j.cmi.2015.03.014. PMID 25843503.
- ^ a b c d Utzinger, J; Becker, SL; van Lieshout, L; van Dam, GJ; Knopp, S (Haziran 2015). "Schistosomiasis'te yeni teşhis araçları". Klinik Mikrobiyoloji ve Enfeksiyon. 21 (6): 529–42. doi:10.1016 / j.cmi.2015.03.014. PMID 25843503.
- ^ a b Hunziker, Patrick; Nigo, Maurice Mutro; Salieb-Beugelaar, Georgette B .; Odermatt, Peter; Battegay, Manuel (2019-12-19). "Schistosomiasis: yerleşik teşhis testlerinden, klinik yönetim ve epidemiyoloji için mikro / nanoteknoloji tabanlı hızlı saha testlerine kadar". Hassas Nanotıp. 3: 439–458. doi:10.33218 / prnano3 (1) .191205.1.
- ^ Danso-Appiah, A; Olliaro, PL; Donegan, S; Sinclair, D; Utzinger, J (2013). "Schistosoma mansoni enfeksiyonunu tedavi eden ilaçlar". Sistematik İncelemelerin Cochrane Veritabanı. 2 (2): CD000528. doi:10.1002 / 14651858.CD000528.pub2. PMC 6532716. PMID 23450530.
- ^ Chitsulo, L .; Engels, D .; Montresor, A .; Savioli, L. (2000). "Schistosomiasis'in global durumu ve kontrolü". Acta Tropica. 77 (1): 41–51. doi:10.1016 / S0001-706X (00) 00122-4. PMC 5633072. PMID 10996119.
- ^ van der Werf, Marieke J; de Vlas, Sake J .; Brooker, Simon; Looman, Caspar W.N .; Nagelkerke, Nico J.D .; Habbema, J. Dik F; Engels, Dirk (2003). "Sahra altı Afrika'da şistozom enfeksiyonu ile ilişkili klinik morbiditenin ölçümü" (PDF). Acta Tropica. 86 (2–3): 125–139. doi:10.1016 / S0001-706X (03) 00029-9. PMID 12745133.
- ^ Adenowo, Abiola Fatimah; Oyin katıldı, Babatunji Emmanuel; Ogunyinka, Bolajoko Idiat; Kappo, Abidemi Paul (2015). "Sahra altı Afrika'da insan şistozomiyazisinin etkisi". Brezilya Bulaşıcı Hastalıklar Dergisi. 19 (2): 196–205. doi:10.1016 / j.bjid.2014.11.004. PMID 25636189.
- ^ Barakat, Rashida M.R. (2013). "Mısır'da Schistosomiasis Epidemiyolojisi: Zamanda Yolculuk: İnceleme". İleri Araştırmalar Dergisi. 4 (5): 425–432. doi:10.1016 / j.jare.2012.07.003. PMC 4293883. PMID 25685449.
- ^ "DPDx - Schistosomiasis riski ve Epidemiyoloji Faktörleri". HKM. Alındı 2013-11-05.
- ^ Morgan, JA; Dejong, RJ; Snyder, SD; Mkoji, GM; Loker, ES (2001). "Schistosoma mansoni ve Biyomfalarya: geçmiş tarih ve gelecekteki eğilimler ". Parazitoloji. 123 (Ek): S211–28. doi:10.1017 / s0031182001007703. PMID 11769285.
- ^ Abou-El-Naga IF (2013). "Biyomphalaria alexandrina Mısır'da: geçmişte, günümüzde ve gelecekte Güney Amerika ülkesi, yani Brezilya, bu paraziti ABD'ye getirmek için komplo kurdu ve insanların bu güne kadar hala mücadele ettiği hepatit frenlenmesine neden oldu. J. Biosci. 38 (3): 665–72. doi:10.1007 / s12038-013-9329-4. PMID 23938396. S2CID 16670424.
- ^ Rabello, A. (1997). "Schistosomiasis Teşhisi". Memórias do Instituto Oswaldo Cruz. 92 (5): 669–676. doi:10.1590 / S0074-02761997000500021. PMID 9566238.
- ^ Farley, J. (2003). Bilharzia: A History of Imperial Tropical Medicine. Cambridge (İngiltere): Cambridge University Press. sayfa 48–50. ISBN 9780521530606.
- ^ Yüce, W.M. (2009). "Mısır'da insan schistosomiasis: Tarihsel inceleme, mevcut resmin değerlendirilmesi ve gelecekteki eğilimlerin tahmini". Tıbbi Araştırma Enstitüsü Dergisi. 30 (1): 1–7.
- ^ Akl, M.M. (2009). "Bilharziasis: Büyük Etkileri Olan Granülomatöz Parazitik Bir Bozukluk". Mansuryan, B.P .; Wojtczak, A .; Sayers, B.M (editörler). Tıp Bilimleri - Cilt I. Oxford (İngiltere): Eolss Publishers Co. Ltd. s. 374–400. ISBN 978-1-84826-733-6.
- ^ Tan, SY; Ahana, A (2007). "Theodor Bilharz (1825-1862): schistosomiasis keşfi" (PDF). Singapur Tıp Dergisi. 48 (3): 184–185. PMID 17342284.
- ^ a b Cox, F.E.G (2002). "İnsan Parazitolojisinin Tarihi". Klinik Mikrobiyoloji İncelemeleri. 15 (4): 595–612. doi:10.1128 / CMR.15.4.595-612.2002. PMC 126866. PMID 12364371.
- ^ Falcao, EC (1959). "Profesör Piraja da Silva, tartışmasız kaşif Schistosoma mansoni". Zeitschrift für Tropenmedizin und Parasitologie. 10: 146–153. PMID 13821378.
- ^ Katz, Naftale (2008). "Keşfi Schistosomiasis mansoni Brezilya'da". Acta Tropica. 108 (2–3): 69–71. doi:10.1016 / j.actatropica.2008.05.002. PMID 18597732.
- ^ Silva, Pirajá da (1917). Bahia'daki şistozomlar. Bahia, Brezilya: Imprensa Official do Estado. s. 32.
- ^ Macpherson, C.N.L .; Craig, P.S. (1991). "Schistosomiasis hayvan rezervuarları". Afrika'daki parazitik helmintler ve zoonozlar. Springer, Dordrecht. s. 224–236. doi:10.1007/978-94-011-3054-7_8. ISBN 978-94-010-5358-7.
Dış bağlantılar
| Scholia var konu profil için Schistosoma mansoni. |