San Francisco Haliç Ekolojisi - Ecology of the San Francisco Estuary
San Francisco Haliç ile birlikte Sacramento – San Joaquin Nehri Deltası oldukça değişmiş bir ekosistem. Bölge, su temini, nakliye, tarım ve son zamanlarda banliyö geliştirme ihtiyaçlarını karşılamak için yoğun bir şekilde yeniden tasarlandı. Bu ihtiyaçlar, suyun hareketinde ve peyzajın doğasında doğrudan değişikliklere neden olmuştur ve yerli olmayan türler. Yeni türler ülkenin mimarisini değiştirdi besin ağı kesinlikle setler Delta olarak bilinen karmaşık sistemi oluşturan adaların ve kanalların manzarasını değiştirdi.[1]
Bu makale özellikle düşüklerin ekolojisi ile ilgilidir. tuzluluk Haliç bölgesi (LSZ). Tarihi bir yeniden inşa etmek besin ağı LSZ için bir dizi nedenden dolayı zordur. Birincisi, tarihi olarak haliçleri işgal eden türlere dair net bir kayıt yok. İkincisi, San Francisco Haliç ve Deltası 10.000 yıllık tarihinin büyük bir kısmında jeolojik ve hidrolojik geçiş halindedir ve bu nedenle haliçin "doğal" durumunu tanımlamak "hareket eden bir hedefi vurmak" gibidir.[1] İklim değişikliği hidrolojik mühendislik, değişen su ihtiyaçları ve yeni Tanıtılan türler Haliçin besin ağı konfigürasyonunu değiştirmeye devam edecek. Bu model, gıda ağının konfigürasyonunu değiştiren son değişiklikler veya tür girişleri hakkında notlar ile mevcut durumun anlık görüntüsünü sağlar. Mevcut besin ağının dinamiklerini anlamak, haliçin işleyişini ve tür çeşitliliğini iyileştirmeye yönelik restorasyon çabalarında faydalı olabilir.
Fiziksel coğrafya
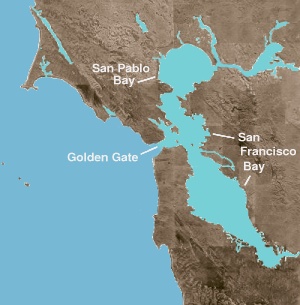
San francisco bay hem bir Defne ve bir Haliç. Önceki terim, açık okyanustan fiziksel bir sığınak sağlayan herhangi bir giriş veya koy anlamına gelir. Haliç, tatlı suyun bir okyanus veya denizle buluştuğu herhangi bir fizyografik özelliktir. Körfezin kuzey kısmı, hem deniz hem de tatlı su akılarının hakim olduğu bir dizi fiziksel setten oluşan acı bir haliçtir. Bu coğrafi varlıklar tuzlu sudan tazeye (veya batıdan doğuya) hareket ediyor: San Pablo Körfezi Merkez Körfez'in hemen kuzeyinde; Carquinez Boğazı yol açan dar, derin bir kanal Suisun Koyu; ve Delta of Sacramento ve San Joaquin nehirler.
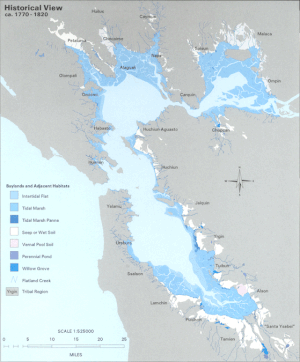
20. yüzyıla kadar, haliçin LSZ'si tule hakim tatlı su sulak alanlar. Bu tarihi sulak alanların% 80 ila 95'i, Körfez Bölgesi çevresindeki arazi kullanımını ve gelişmeyi kolaylaştırmak için doldurulmuştur.[2] Sınırlarında habitat kaybı pelajik bölge yerel kayıp yarattığı düşünülmektedir pelajik balık türleri, avlanmaya karşı savunmasızlığı artırarak.
gelgit arası ve Bentik Haliç şu anda hakim Çamurluklar bu büyük ölçüde aşağıdakilerden türetilen sedimantasyonun sonucudur altın madeni içinde Sierra Nevada 19. yüzyılın sonlarında. Yüksek tortu yüklerine yönelik eğilim, 1950'lerde, Central Valley Projesi çökeltinin çoğunu barajların arkasına hapsetmek ve haliçten yıllık net çökelti kaybına neden olmak.[3] Bu nedenle, bulanıklık son derece yüksek kalmasına rağmen, çamur tabakaları yavaş yavaş azalmakta gibi görünmektedir. Suyun yüksek bulanıklığı, yüksek besin bulunabilirliğinin yüksek fitoplankton üretimine yol açmadığı San Francisco Haliçinde var olan benzersiz durumdan sorumludur. Bunun yerine, çoğu yosun fotosentetik organizması ışıkla sınırlıdır.[4]
Delta da aynı şekilde ağır bir değişiklik yaşadı. 19. yüzyıldan başlayarak, tarım alanlarını düzenli sel baskınlarından korumak için, doğal olarak oluşan setler kalıcılık için güçlendirildi. Bu çiftliklerin çoğu, Delta su yollarının ortasında oluşan turba adalarında kuruldu. Yoğun tarım Toprağın yüksek karbon içeriğini oksitleyerek toprak kütlesinde önemli kayıplara neden oldu. Sonuç olarak, bu adalar deniz seviyesinin yaklaşık 6 metre altına indi veya battı.[5] Delta bugün, havzaları su yüzeyinin çok altında olan "yüzen kaseler" gibi görünen adalarla noktalanmış, oldukça yırtık su yollarından oluşmaktadır.[6] Bu adalar, deniz seviyesinin çökmesi nedeniyle yüksek sel riski altındadır. Tuzlulukta müteakip doğuya doğru kaymanın, San Francisco Halici'nin tüm LSZ'sinin ekolojisini dramatik bir şekilde değiştirmesi bekleniyor.[7]
Hidrodinamik

LSZ yaklaşık 2 psu (pratik tuzluluk birimleri tuzluluk ölçümü) ve yaklaşık 6 psu ila 0.5 psu arasında değişir. Haliçin birincil tatlı su girdileri, bölgesel yağış, Sacramento Nehri ve San Joaquin Nehri'nin akışından kaynaklanmaktadır.[8]
Nehir girişi büyük ölçüde yukarı havza rezervuar salımlarıyla kontrol edilir. Bu girişin önemli bir kısmı, federal tarafından Delta dışına ihraç edilmektedir. Central Valley Projesi ve Devlet Su Projesi güneye Kaliforniya tarımsal ve kentsel kullanım için. Bu değişiklikler, haliçten dışarıya akıştaki varyasyonun çoğunu ortadan kaldırmıştır. altın Kapı ), kışın daha düşük çıkış ve yazın haliçte tarihsel olarak bulunandan daha yüksek çıkış yaratır. Ortalama olarak, haliçteki tatlı su akışları, tarihi akışların% 50'sini oluşturmaktadır.[9] Fitoplankton, Zooplankton ve larva ve yetişkin balıklar ihraç pompalarına girerek bu organizmaların bolluğu üzerinde potansiyel olarak önemli ancak bilinmeyen bir etkiye neden olabilir. Bu özellikle nesli tükenmekte olan kişiler için geçerli olabilir delta kokusu küçük bir endemik balık; tarihsel anlatılarda muazzam bir şekilde bol olarak tanımlanmış olması dışında istisnai değildir.[10] Delta kokusunun, yaz başlarında Delta'da yukarı doğru göç edip yumurtladığına ve yumurtalarını ve larvalarını sürüklenme riskinin yüksek olmasına yol açtığına inanılıyor.[11] Kokunun yönetimi şu anda tartışmanın kaynağıdır çünkü ekolojisi Kaliforniya'daki koruma, geliştirme ve tarımın farklı su ihtiyaçlarını çatışma sürecine sokmaktadır.
Suyun haliç dışına hareketi karmaşıktır ve bir dizi faktöre bağlıdır. Gelgit döngüleri Suyun 24 saatlik bir süre içinde dört kez Altın Kapı'dan uzaklaşmasına neden olur.
Düşük Tuzluluk Bölgesi için bir işaret olarak 2 psu kullanılarak, dalgalanmaların yönü ve büyüklüğü Altın Kapı'dan veya X2'den kilometre cinsinden uzaklık olarak izlenebilir. X2'nin konumu, giriş, ihracat ve gelgitler dahil olmak üzere bir dizi fiziksel parametreye dayandığından, konumu günlük ve mevsimsel bir döngüde kilometrelerce değişir; bir yıl boyunca, yüksek akış dönemlerinde San Pablo Körfezi'nden yaz kuraklığı sırasında Deltaya kadar değişebilir. X2'nin konumu, ihracat talebi beklentisiyle yukarı akış rezervuarlarından su bırakılarak dikkatle izlenir ve korunur. Bu, Eyalet Su Kurulu Kararı 1641 tarafından zorunlu hale getirilmiştir ve X2, Şubat ayından Mayıs ayına kadar veya Collinsville'in doğusunda ( Ocak, Haziran, Temmuz ve Ağustos aylarında Golden Gate Köprüsü'nün 81 nehir kilometre yukarısında. (D-1641 pp 150)
Yerçekimsel sirkülasyon tabakalı yüksek tuzlu suyun karaya doğru akmasına neden olurken, üstten düşük tuzlu su denize doğru akar.[12] Yerçekimi sirkülasyonunun etkisi en çok tatlı su akışının yüksek olduğu dönemlerde ortaya çıkarak haliçteki pelajik organizmaların dağılımını ve tuz alanını korumak için olumsuz bir geri bildirim sağlar.
Karışım, yerçekimi sirkülasyonunun karaya bakan kenarında, genellikle su sütununun daha az tabakalaştığı X2 civarında önemlidir.[13] Carquinez Boğazı'nın doğu ucundaki "Benicia Tümseği" nde sabit bir karışma bölgesi oluşur ve burada derin kanal Suisun Körfezi'ne girerken önemli ölçüde sığlaşır.[14] Tuzluluğun muhafaza edilmesinde karıştırma kritiktir, öyle ki, X2'yi batıya kısa bir mesafe taşımak için çok büyük miktarda tatlı su gerekir. Karıştırma ayrıca pelajik organizmaların haliçteki konumunu korumasına yardımcı olur.[1] sistemin dışına birincil ve ikincil üretimin ilerlemesini yavaşlatmak.
Pelajik bölge
Pelajik organizmalar hayatlarının tamamını veya bir kısmını açık suda geçirirler. yetişme ortamı kenarlarla değil, tuzluluk ve sıcaklığa fizyolojik toleransla tanımlanır. Düşük Tuzluluk Bölgesi (LSZ) San Francisco Haliç Bu eşsiz birleşim noktasında hayatta kalmak için uzmanlaşmış bir organizma grubu için bir habitat oluşturur. karasal, temiz su, ve deniz etkiler. Farklı yaşam alanlarına sahip birçok habitat varken ekolojiler bu parçası Haliç (deniz, tatlı su, gelgit bataklığı ve bentik çamurluk sistemleri) her biri, tatlı su, besin maddeleri, karbon ve organizmaların ihracatı ve ithalatı yoluyla LSZ'ye bağlıdır.[1]
LSZ'deki organizmaların dağılımı ve bolluğu hem abiyotik hem de biyotik faktörlere bağlıdır. Abiyotik faktörler şunları içerir: fiziksel coğrafya ve hidroloji Haliç dahil besin girişler, tortu yük, bulanıklık çevresel stokastisite, iklim ve antropojenik etkiler.[8] Abiyotik faktörler, nehir ağzı ortamında üretimi yönlendirme eğilimindedir ve biyotik faktörlerin aracılık eder.
Biyotik faktörler arasında besin alımı ve birincil üretim, ikincil üretim nın-nin Zooplankton, besin ağı ve trofik dinamik enerjik transfer, sistem içinde ve dışında yayılma, hayatta kalma ve ölüm, yırtıcılık, ve rekabet itibaren Tanıtılan türler.
Besin ağı

Tarihsel olanı karakterize etmek zor besin ağı Geçtiğimiz yüzyılda coğrafya, hidroloji ve tür kompozisyonunda meydana gelen dramatik değişiklikler nedeniyle San Francisco Haliç'i. Ancak 1970'lerde başlayan izleme, gıda ağının tarihsel dinamikleri hakkında bazı bilgiler verir. 1980'lerden önce, LSZ'ye fitoplankton güdümlü bir gıda ağı hakimiyetindeydi, istikrarlı bir mezoplankton popülasyonu. kopepod Eurytemora affinis ve San Francisco Körfezi ile tipikleştirilmiş büyük makrozooplankton karides ve mysid karidesler.[15] Bunlar kuzey gibi yerel filtre besleyicilere besin ve enerji sağladı. hamsi (Engraulis mordax) ve planktivorlar gibi delta kokusu ve çocuk Somon.
Besin ağı değişikliği, birincil ve ikincil üretim ile ilgili bölümlerde anlatıldığı gibi, tarihsel olarak artan bulanıklık ve daha yakın zamanda ortaya çıkan türler tarafından yönlendirilmiştir.
Özellikle, tanıtılan yüksek temizlenme oranı Amur nehri midye Potamocorbula amurensis popülasyon, plankton yoğunluğunda on katlık bir düşüşe neden oldu ve bu da, Benthos ve atık kırıntı üretiminde varsayılan bir artış.[16] Bu atığın mikrobiyal döngüyü beslediği varsayılmaktadır, bu da kopepod gibi mikrozooplanktonda bir artışa neden olur. Limnoithona tetraspina, kullanan rotiferler ve siliatlar.
Bu değişiklikler, balık stoklarının azalmasının bir nedenidir. Örneğin kuzey hamsi, Engraulis mordax, 1980'lere kadar Düşük Tuzluluk Bölgesinde, haliçteki yayılış alanı Orta ve Güney Körfezlerle sınırlı olana kadar oldukça boldu.[17] Bunun nedeni muhtemelen Amur Nehri'nin girişinden sonraki davranışsal tepkidir (Potamocorbula amurensis) istiridye ve ardından plankton mevcudiyetindeki düşüş.
Daha yakın zamanlarda, genel bir pelajik organizma düşüşü (POD) tanımlandı ve bu, bilimsel, yönetimsel ve politik topluluklarda çok fazla endişe kaynağı oldu.[18]
Aşağıdakiler dahil birkaç anahtar tür: delta kokusu, longfin kokusu, çizgili levrek, ve threadfin shad 2001 yılında başlayan bolluktaki kademeli düşüş nedeniyle "ilgi çekici türler" ilan edilmiştir.[19] Buna ikincil üretkenlikteki benzer bir düşüş eşlik etti ve şu anda birçok araştırmanın kaynağı. Gıda ağındaki düşüş, Delta'dan su ihracatı ve kentsel, endüstriyel veya tarımsal kaynaklardan toksik maddeler dahil olmak üzere POD'u açıklamak için bir dizi hipotez öne sürüldü.
Yapımcılar
Birincil üretim ve besin alımı
Birincil üretim fitoplankton ile enerji ve temel besinleri biyolojik olarak mevcut bir forma (yani gıda) sabitler. fotosentez. Fitoplankton üretimi büyük ölçüde fiziksel parametrelerle yapılandırılır: besin bulunabilirliği, güneş ışığı, bulanıklık ve sıcaklık.
San Francisco Estuary, birincil üretim için kullanılabilen, büyük ölçüde atık su arıtma tesislerinden, tarımsal ve kentsel drenajdan ve okyanustan elde edilen çok sayıda besin kaynağına sahiptir.[20][21] Buna rağmen, haliç nispeten düşük bir birincil üretim oranına sahip olma eğiliminde olması bakımından benzersizdir.[22] Bu muhtemelen iki faktöre bağlıdır: şeklinde büyük nitrojen girdileri amonyum metabolik olarak daha ucuz olan NH3'ü tercih eden fitoplankton tarafından nitrat alımını baskılayan4+ ve fotosentez için gereken ışığı su kolonunun üst birkaç santimetresine kadar sınırlayan yüksek bulanıklık.[21] Bu bulanıklık, dünyadaki hidrolik altın madenciliğinin mirasıdır. Sierra Nevada 1850'lerde.[23]
Haliçte suyun yüksek kalış süresi, fitoplankton biyokütlesinin birikmesine izin vererek yoğunluğu artırırken, düşük kalma süresi fitoplanktonu haliçten uzaklaştırır.[1] İkincisi, yüzey sularının parçacıkları ve planktonu aşağıya doğru akıtmaya meyilli olduğu yüksek akış dönemlerinde haliçin ana kanalları için tipiktir.
Otçul ayrıca fitoplanktonu su kolonundan uzaklaştırır. Pelajik besin ağı fitoplankton üretimine dayanırken, bu üretimin çoğu, tanıtılan Amur Nehri midyesi tarafından avlanma yoluyla bentoslara yönlendirilir (Corbula amurensis ). Fitoplankton biyokütlesi seviyeleri, yaygın olarak kullanılmaya başlanmasından sonra bir derece azaldı. C. amurensis 1980'lerin ortalarında ve toparlanmadı.[24]
Fotosentetik üretim
Ana kaynağı fotosentetik olarak türetilmiş enerji fitoplankton. Genel konuşma, diyatomlar ve mikroflagellatlar, biyolojik olarak kullanılabilir karbon Haliçte.[25] Diğer türler, özellikle Dinoflagellatlar üretebilir zararlı alg çiçekleri veya kırmızı gelgitler, bunlar gıda ağına asimilasyon için daha az hazırdır.[26]
Birincil üretim fitoplanktondan iki farklı faktörün bir fonksiyonudur: büyüme oranları ve birikim (Şekil 1). LSZ, yüksek konsantrasyonlarda besinler kentsel ve tarımsal kaynaklardan fitoplankton üretim oranları oldukça düşüktür.[27] Nitrat fitoplankton tarafından büyüme için optimal olarak kullanılır, ancak amonyum (büyük ölçüde kanalizasyon deşarjlarından kaynaklanmaktadır) büyüme hızı üzerinde baskılayıcı bir etkiye sahiptir. Bu nedenle, besin değeri sınırlı olmamakla birlikte, fitoplankton, mevcut nitrojen türleri nedeniyle daha yavaş büyüme eğilimindedir.[28] Büyüme oranı üzerindeki bir başka baskılayıcı faktör de yüksek bulanıklık Haliçin kabiliyetini sınırlayan fotosentetik olarak aktif radyasyon (PAR) su sütununun üst birkaç santimetresinin ötesine nüfuz etmek için. Bu, fitoplankton fotosentezini nispeten sığ bir fotik bölge. Böylece su sütunu dır-dir tabakalı bulanıklık yüksektir ve amonyum mevcuttur, fitoplankton büyüme hızı tipik olarak baskılanır.[29][30]
Fitoplankton birikimi esas olarak aşağıdakilerin sonucudur: kalış süresi.[27] Kuzey Deltası ve Suisun Koyu Aşağı akış için bölge boyunca hareket eden yüksek su hacmi nedeniyle nispeten düşük kalma sürelerine sahiptir ve ihracat Güney Kaliforniya'ya. Sistemin bu kısmında su daha hızlı hareket ettiğinden, sistem dışına verimlilik ilerledikçe birikim oranı azalmaktadır. Aksine, güney Deltası'nın bazı kısımları, sistem boyunca hareket eden düşük su hacmi nedeniyle daha yüksek bir ikamet süresine sahiptir; aslında, San Joaquin Nehri'nden gelen akış eksikliği ve ihracat pompalama nedeniyle su zaman zaman geriye doğru akıyor. Yaz aylarında fitoplankton yoğunluğu, haliçin diğer bölgelerine göre burada daha büyük bir mertebede olabilir.[31]
Dinoflagellatların zararlı alg çoğalmaları (HAB'lar) veya siyanobakteriler onları birçok organizmaya zararlı hale getiren toksik metabolik yan ürünler üretir. Yüksek besin konsantrasyonları ve sıcaklıkların bir kombinasyonuyla beslenen HAB'lar, diatomları ve mikroflagellatları rekabetçi bir şekilde dışlayarak gıda ağı üzerinde iki kat negatif etkiye sahiptir ve biyolojik olarak kullanılabilir birincil üretimi daha da azaltır. Kesin olsa da omurgasızlar gibi çift kabuklular doğrudan etkilenmeyebilir, toksinleri gıda zincirine yayabilir, hastalanabilir veya öldürebilirler avcılar. Nasıl olduğu iyi anlaşılmadı kopepodlar etkilenir. İstilacı yosun Microcystis aeruginosa Delta'da artık yaz aylarında yaygındır ve kopepod üretkenliğini azaltabilir (potansiyel olarak kanserojen insanlar için).[32]
Detrital üretim
Muazzam miktarlarda tortu ve döküntü LSZ aracılığıyla akı. Bunların çoğu, çözünmüş ve parçacık halindeki organik kalıntılardır. organik madde (Sırasıyla DOM ve POM). Yukarı akış kaynaklarına ek olarak, organik madde yerel organizma ölümlerinden ve atık üretiminden de birikebilir.[33][34]
Detritivorlar potansiyel olarak büyük öneme sahip alternatif ve paralel bir besin ağı yaratarak bu enerji kaynağından yararlanın. Bunun nedeni ise karbon fiksasyonu kırıntılı besin ağına geçiş tabakalaşma, bulanıklık veya gün uzunluğu ile sınırlı değildir, bunların hepsi fotosentezi sınırlar. Detrital üretim, yalnızca Delta sisteminden gelen girdiler ve ilerlemeler ile sınırlı olarak sürekli gerçekleşir.[25][35]
Bakteriler, DOM ve POM'un biyolojik olarak kullanılabilir karbona dönüşümünün başlıca ajanlarıdır. mikrobiyal döngü. Bu mekanizma, sınırlı besin maddelerinde özellikle önemlidir deniz sistemler, nerede bakteri besinleri batan artıklardan serbest bırakarak fotik bölgeye geri dönüştürülmesini sağlar. San Francisco Halici'ndeki mikrobiyal döngünün işlevi için çok az çalışma yapılmıştır, ancak bakterilerin rolü bir bölgedeki besin maddelerinin geri dönüşümü için kritik olmayabilir. ötrofik sistemi. Bunun yerine, doğrudan otlatma yoluyla alternatif bir besin zinciri sağlayabilirler. kamçılılar, rotiferler ve siliatlar.[36]
Yüksek bolluk siklopoid kopepod Limnoithona tetraspina birincil besin kaynağı olarak fitoplankton yerine kirpiklere olan bağımlılığından kaynaklanıyor olabilir.[37] Başlıca türleri kalanoid kopepodlar ayrıca kirpikleri tamamlayıcı veya hatta birincil besin kaynağı olarak kullanabilir, ancak ne derece bilinmemektedir.[38][39]
İkincil üretim
İkincil üretim, birincil üretimle beslenen ve enerjiyi daha yükseğe aktaran organizmaları ifade eder. trofik seviyeler nehir ağzı foodweb. Tarihsel olarak, San Francisco Halici'ndeki ikincil üretime, mysid karides üretim.[40][41] Ancak, yerel mysid Neomysis mercedis büyük ölçüde yerine getirildi Acanthomysis bowmani, daha düşük yoğunluklarda devam eder. Tanıtılan amfipod Gammurus daiberi bu nişin bir kısmını devralmış olabilir, ancak büyük ölçüde tatlı su ile sınırlıdır.
Bugün, ikincil üretimin ana kaynağı, kopepodlar. Vatandaşlığa kabul edilen yerli kalanoid kopepod Eurytemora affinis 19. yüzyılın sonlarına doğru tanıtıldığına inanılıyor.[42] Düşük tuzluluk bölgesinin zooplanktonuna, 1980'lere kadar büyük ölçüde başka bir kalanoid kopepod ile değiştirildiği zamana kadar hakim oldu. Pseudodiaptomus forbesi.[16][43] P. forbesi, amur nehri istiridyesinin tuzluluk toleransı aralığının dışında, özellikle Deltada olmak üzere, haliçin tatlı su, yüksek yerleşim bölgelerinde bir kaynak popülasyonu sürdürerek devam etmektedir.[44] Bir zamanlar baskın olan E. affinis, yukarı akış menzilinden yoksun olduğundan, midye tarafından avlanmaya karşı daha savunmasızdır ve P. forbesi ile bariz rekabetten muzdariptir.
Önemli olabilecek diğer kalanoid kopepodlar, son zamanlarda Sinokalanus doerri ve Acartiella sinensis. Morfolojilerine bağlı olsalar da, bu organizmaların yaşam öyküleri hakkında çok az şey biliniyor, diğer kopepodları avlayabilirler. Düzensiz bolluk döngülerinde ortaya çıkarlar ve bu sırada zooplanktona hakim olabilirler.[45]
Yine başka bir istilacı kopepod, çok küçük siklopoid Limnoithona tetraspina, 1990'larda Düşük Tuzluluk Bölgesinde ortaya çıktı. O zamandan beri, L. tetraspina sayısal olarak baskın kopepod haline geldi ve 10.000 / m3 civarında yoğunluklara ulaştı. Güveniyor mikrobiyal döngü besin kaynağı olarak bakteriler, siliatlar ve rotiferlerle beslenir.[37] Ek olarak, bilinmeyen nedenlerden dolayı Amur Nehri istiridye tarafından avlanmaya karşı savunmasız görünüyor. Küçük boyutundan dolayı, L. tetraspina genellikle daha büyük avcılar, özellikle balıklar tarafından tüketilemez ve bu onu enerjik bir çıkmaz yapar.
Tüketiciler
Birincil tüketiciler
Birincil tüketiciler güvenmek birincil üretim ana besin kaynağı olarak. LSZ'nin pelajik ağının en önemli tüketicileri, yukarıda bahsedilen rotiferler, kamçılılar ve siliatların yanı sıra kopepodlardır. Tüm kalanoid kopepod türleri, yakın zamanda piyasaya sürülen Amur Nehri istiridyesinden kaynaklanan yüksek avlanma baskısı altında azalmıştır (Corbula amurensis).[16] Bu nedenle ve kopepodlar hem fotosentetik hem de parçalayıcı gıda kaynaklarına dayandıkları için, LSZ'deki kopepodlar, kopepodların çiçeklenmeleri birkaç gün içinde otlatabildiği deniz ve lentik sistemlerin aksine, birincil üretim konusunda sınırlı geribildirime sahiptir.
Pseudodiaptomus forbesi biyokütle açısından LSZ'nin baskın kalanoid kopepodudur. Hem düşük tuzlulukta hem de tatlı suda kalabilecek kadar geniş bir tuzluluk toleransına sahiptir. Bu geniş dağılım, daha dar tuzluluk toleranslarına sahip diğer türlerin aksine, popülasyonun avlanmadan yukarı akış sığınağı sağlamasına yardımcı olur.[44]
Limnoithona tetraspina 1993 yılında piyasaya sürülmesinden bu yana sayısal olarak baskın siklopoid kopepod haline geldi. Öncelikle siliatlar ve mikroflagellatlar ile beslenir, ancak bunun aksine P. forbesiistiridye veya balık tarafından avlanmaya nispeten dayanıklıdır, dolayısıyla bolluğu vardır. Enerjik, L. tetraspina besin ağı için bir çıkmaz olabilir; bu kopepodlar ya gelgitler ve akıntılar tarafından sistemin dışına yönlendirilir ya da ölür ve mikrobiyal döngü ya da detritivorlar için mevcut olabilecekleri benthos'a düşer.[46]
Yırtıcı kopepodlar
Delta boyunca, hakkında nispeten az şey bilinen bir dizi yırtıcı kopepod bulunmaktadır. Sinokalanus doerri, Acartiella sinensis, ve Tortanus dextrilobatus hepsi morfolojik olarak diğer kopepodlar üzerinde avlanma yeteneğine sahip gibi görünmektedir. Her biri, muhtemelen balast suyu 1980'lerden beri değişim. Genel olarak, kopepod tüketicilerini olumsuz etkilemeye yetecek miktarda değildirler; ancak, periyodik çiçek açan S. doerri ve A. sinensis iyi çalışılmamış olan meydana gelir.[47]
Makro omurgasızlar

Yapabilirken filtre besleme, mysids (minik karides benzeri yaratıklar) büyük ölçüde etoburdur ve kopepodlu yetişkinlerle beslenir. Yavru balıklar da dahil olmak üzere plankton ve planktivor balıklar arasında enerjik bir kanal sağladılar. mersin balığı, Chinook somonu, ve Amerikan shad. Mysids bir zamanlar yerli olana kadar boldu. Neomysis mercedis 1980'lerin ortasında istilacı tarafından değiştirildi Acanthomysis bowmanidaha küçük ve daha az bol olan. Mysid düşüşü, 1980'lerde ve 90'larda haliçteki bazı balık türlerindeki müteakip düşüşle ilişkilendirildi.[48][49]
Karides büyük ölçüde mysids üzerinde avlanan genelci etoburlar ve amfipodlar. Crangon franciscorum Haliçte kalan iki ticari balıkçılıktan birini temsil etmektedir. Artık "San Francisco Körfezi karides kokteylleri" için kullanılmasa da, yem olarak hasat ediliyorlar. Diğer avcılar arasında çizgili levrek ve Chinook somonu yetişkinler ve smolts.[50]
Balık

Çünkü balıklar taksonomik olarak ve morfolojik olarak çeşitli grup, türler kendilerine göre değişir trofik ekolojiler. Genel olarak, balıklar dört geniş besleme kategorisine ayrılabilir: Filtre besleyicileri planktivorlar piskivorlar ve Bentik besleyiciler.
Filtre besleyiciler, tipik olarak fito ve zooplankton olmak üzere küçük avlar için su kolonunu gelişigüzel zorlar. Bu balık kategorisi şunları içerir: threadfin shad (Dorosoma petenense), Amerikan shad (Alosa sapidissima), iç gümüşler (Menidia beryllina), ve hamsi (Engraulis mordax). Bazı kanıtlar, bu türlerin bazılarının, Amur Nehri midyesinin ortaya çıkmasından sonra çökmüş plankton seviyeleri nedeniyle yiyecekle sınırlı olduğunu göstermektedir. Hamsiler, LSZ'yi San Pablo ve Merkez Körfezlerdeki haliçin daha üretken bölgelerine bıraktı.[17]
Planktivorlar seçici olarak kopepodlar, mysidler ve mysidler gibi bireysel zooplanktonları avlar. Gammarids. Bu grupta çoğu balık larvası bulunur, delta kokusu (Hypomesus transpacificus ) ve longfin kokusu (Spirinchus thaleichthys ), tule levrek (Hysterocarpus traski), ve Somon smolts. Delta kokusu nedeniyle özellikle ilgi çekicidir. tehlike durumu. Yiyeceklerle sınırlı olabilir, ancak kanıtlar biraz çelişkili. Delta'dan gelen tatlı suyun ihracatı pompalanmasında yumurta ve larvaların sürüklenmesi gibi diğer faktörler de düşüşü açıklayabilir.[11]

LSZ'nin ana piskivoru, çizgili levrek (Morone saksati ), 1890'larda tanıtılan ve ağırlıklı olarak yerli balıkları avlayan. Çizgili levrek, San Francisco Haliç'teki önemli bir spor balıkçılığıdır ve bu nedenle, haliçten biyokütlenin küçük bir geri çekilmesini temsil eder.[51]
Bentik veya dipte yaşayan balıklar şunları içerir: beyaz mersin balığı (Acipenser transmontanus), beyaz yayın balığı (Ameiurus catus), ve yıldızlı pisi balığı (Platichthys stellatus). Onların yüzünden yetişme ortamı oryantasyon, öncelikle amfipodlar, defne karidesi ve çift kabuklular gibi epibentik organizmalardan beslenirler. Bu balıkların en azından ara sıra Amur Nehri istiridye ile beslendiği bilinmektedir; bu, kırıntılı üretim dışında, bu türden enerji akışı için birkaç kanaldan birini temsil eder.[52]
Tek ticari balıkçılık LSZ'deki yem karidesleri içindir. Çeşitli var spor balıkçılığı Bu, küçük bir karbon akışını, ancak haliç çevresindeki yerel ekonomilere önemli miktarda sermaye akışını temsil eder. Eğlence amaçlı balıkçılığın çoğu çizgili levrek, mersin balığı çevreliyor ve tatlı su Deltası'nda tatlı su levreği tanıttı. Bu balıkçılık kıtlığı San Francisco Haliç'i benzersiz kılıyor. Neredeyse tüm dünyadaki haliçler, en azından önemli balıkçılık kalıntılarını destekliyor.[53] San Francisco Halici bir zamanlar somon, hamsi ve hamsi için büyük balıkçılığı destekledi. Dungeness yengeçleri 1950'lere kadar. Bu balıkçılığın ölümü muhtemelen daha çok Habitat kaybı -den aşırı hasat.[2]
Kuş
San Francisco Halici, Pasifik geçiş yolu su kuşları göç etmek için. Yine de, haliçin içinden veya dışına karbon akışı hakkında çok az şey bilinmektedir. kuşlar. Milyonlarca su kuşları her yıl sığ sığınakları sığınak olarak kullanın. Kuşların çoğu dabbling ördekler batık sucul bitkilerle beslenen. Dalış ördekler (örneğin Scaup'lar ) C. amurensis gibi epibentik organizmalarla beslenmek, aksi takdirde çıkmazdan olası bir karbon akışını temsil eder.[54][55] İki nesli tükenmekte olan türler kuşların yüzdesi burada bulunur: California en az sumru ve California tokmağı rayı. Maruz defne çamurları için önemli beslenme alanları sağlamak kıyı kuşları ancak altta yatan defne çamur katmanları, körfez çevresinin birçok kısmına yakın yapılar için jeolojik tehlikeler oluşturmaktadır. Piscivorous gibi kuşlar karabataklar ve pelikanlar Haliç de yaşamaktadır, ancak trofik etkileri üzerinde yeterince çalışılmamıştır.
Ocak 2015'te bilim adamları, San Francisco Körfezi kıyı şeridi boyunca kuşların üzerinde gri, kalın, yapışkan, kokusuz bir madde tabakası belirlemek için çalışıyorlardı. Yüzlerce kuş öldü ve yüzlercesi de bu maddeyle kaplandı. Bilim adamları, bu maddeden dolayı risk altında olabilecek diğer yaban hayatı konusunda endişeli.[56]
Memeliler


1825'ten önce, İspanyollar, Fransızlar, İngilizler, Ruslar ve Amerikalılar, muazzam miktarlarda hasat için Körfez Bölgesi'ne çekildiler. kunduz (Castor canadensis), nehir su samuru, sansar, balıkçı, vizon, tilki, sansar, liman ve kürklü foklar (Callorhinus ursinus) ve Deniz su samuru (Enhydra lutris). Bu erken kürk ticareti, California Fur Rush Batı'nın ve Batı'nın açılmasından sorumlu olan diğer tek faktörlerden daha fazlasıydı. San Francisco Körfez Bölgesi özellikle dünya ticaretine.[57] 1817'de bölgedeki su samuru pratikte ortadan kaldırıldı.[58] Ruslar yakınlarda bir mühür istasyonu kurdular. Farallon Adaları 1812'den 1840'a kadar, Amerikan gemileri zaten adaları istismar etmiş olsa da, yılda 1.200 ila 1.500 kürk fok alıyor.[59] 1818'e gelindiğinde, mühürler, yılda sadece 500 kadar alınabilene kadar hızla azaldı ve önümüzdeki birkaç yıl içinde, kürklü fok, 1996'da adaları yeniden kolonileştirmeye başlayana kadar adalardan çıkarıldı.[60] 20. yüzyıl doğa bilimcileri, kunduzun kıyı akarsularında veya Körfez'in kendisinde tarihsel olarak var olduğu konusunda şüpheci olsalar da,[61] daha önceki kayıtlar, Kaliforniya altın kunduzunun (Castor canadensis ssp. subauratus) alınan hayvanların en değerlilerinden biriydi ve görünüşe göre çokça bulundu.[57] Thomas McKay Hudson's Bay Company'nin bir yıl içinde San Francisco Körfezi kıyılarında 4.000 kunduz derisi aldığını bildirdi. Son zamanlarda, kunduz acı suyu yeniden kolonileştirdi Napa Sonoma Bataklığı Kuzeyde San Pablo Körfezi ve Onun Sonoma Deresi ve Napa Nehri kolları. Ayrıca yakın zamanda Körfezi ve kollarını yeniden kolonize eden Kuzey Amerika nehir su samuru (Lontra canadensis) ilk olarak rapor edildi Redwood Creek -de Muir Plajı 1996'da[62] ve son zamanlarda Corte Madera Deresi ve güney körfezde Coyote Creek,[63] ve 2010'da San Francisco Körfezi'nde Richmond Yat Limanı.
Deniz anası
Deniz anası Haliç yakın zamana kadar yaygın değildi. Doğu Kıyısı haliçlerinde, örneğin Chesapeake Körfezi sık sık üst düzey avcılar, hem balık hem de zooplanktonla gelişigüzel besleme. LSZ ve tatlı su bölgelerinde birkaç küçük istilacı takson tespit edilmiştir. Bu türler yazın okşar, ancak polipler içinde Benthos yıl boyunca.
Üzerindeki etkileri plankton bilinmiyor, ancak ölçmek için araştırmalar devam ediyor. Yeterli yoğunlukta jöleler, planktonik türlerin yırtıcı olmayan bir sığınağa sahip olduğu istiridye aralığı dışında düşük tuzlu alanlarda yaşayarak zooplanktonun baskılanmasında C. amurensis için tamamlayıcı bir role sahip olabilir.
Bentik tüketiciler
Bentik topluluk, önemli istilalar nedeniyle haliçin gıda ağı ekolojisinde orantısız şekilde büyük bir rol üstlenmiştir. çift kabuklular.[16] Bu istiridyelerin kullanımı, Amur Nehri istiridyesi (Corbula amurensis) tatlı suya değil, düşük tuzluluk bölgesine uzanan geniş bir tuzluluk toleransına sahiptir. Filtre, fitoplankton ve kalanoid kopepod gibi küçük zooplanktonlarla beslenir. Nauplii. İstiridyenin San Francisco Haliçinde birkaç yırtıcı hayvanı vardır ve bu, yüksek yoğunluklara (onbinlerce / m2 mertebesinde) büyümesine izin verir. Yüksek temizlenme oranları nedeniyle, birkaç gün içinde haliçin tüm su kolonunu birkaç gün içinde temizleyebilir ve bu da büyük ölçüde tükenmiş plankton popülasyonlarına yol açar.[64] Bu, istiridyelerin 1980'lerin ortalarında istilasından sonra ekosistem üretkenliğindeki düşüşün ana nedeni olduğu düşünülüyor.[16][65][66]
Üretkenlikteki bu düşüş, esasen pelajik ağın bu tür tarafından bir bentik zincire yeniden yönlendirilmesinden kaynaklanmaktadır.[16] Amur Nehri deniz tarağı birincil üreticiler, tüketiciler ve yırtıcılardan beslendiği için birden çok trofik seviyeler. Sonuç olarak, neredeyse tüm planktonlar, bariz rekabet bir trofik seviyedeki üretim, midye bolluğunu artırarak diğerlerini etkiler. Bu, olumsuz sonuçlanır geribildirim döngüsü: C. amurensis plankton biyokütlesini sınırlar, bu da sırayla sınırlar C. amurensis. Ancak, gelgit ilerlemesi veya yukarı akış kaynakları nedeniyle sistem dışından gelen girdiler artabilir. C. amurensis biyokütle, plankton sınırlamasını daha da artırır. Bu geri bildirim döngüsü daha da güçlendirilir çünkü midye bir veya iki yıldan fazla sürebilir ve bu da düşük verimlilik döngüleri sırasında plankton popülasyonları üzerinde ek baskı oluşturur.
Karbonun C. amurensis tarafından benthos'a yeniden yönlendirilmesi sınırlı bir zincir oluşturarak pelajik ağın depauperate olmasını sağladı. İstiridye atılımından ve ölümünden kaynaklanan detrital üretim, bakteri üretimini tetikleyebilir, bu da kırıntılı besin ağı veya mikrobiyal döngü içinde dolaştırılabilir.
Geri dönüştürülen besinler bazı fitoplankton büyümesini destekleyebilirken, sonuçta artan C. amurensis popülasyonlarına geri beslenir. Limnoithona tetraspina'nın son işgal başarısı, bu fenomen açısından anlaşılabilir. Besleniyor siliatlar ve midye tarafından otlatılamayacak kadar küçük olan mikroflagellatlar, böylece rekabeti önler. Bunlara ek olarak, L. tetraspina istiridye veya (neredeyse) başka herhangi bir şey tarafından avlanmaya karşı dayanıklı görünmektedir. Mikrobiyal besin ağının yükselişi ve işgali L. tetraspina C. amurensis tarafından kolaylaştırılan, besin ağındaki enerji akışı için kullanılmayan alternatif bir yolun sonucudur. Sonraki istila modelleri benzer bir modeli yansıtabilir.
Tanıtılan türler
Ticareti ve trafiği artırmanın bir işlevi olarak türlerin tanıtımı en az 19. yüzyıldan beri artmaktadır. Girişler, kopepodlar dahil olmak üzere çok sayıda taksonu içerir, karides, amfipodlar, çift kabuklular, balık ve hem köklü hem de yüzen bitkiler. Birçok pelajik tür, son zamanlarda balast suyu büyük gemilerden doğrudan haliçlere salınır.[67] Sonuç olarak, ortaya çıkan bu türlerin çoğu Pasifik Kıyısı çevresindeki haliçlerden, özellikle P. forbesi ve L. tetraspina gibi kopepodlardan kaynaklanmaktadır. Amur Nehri istiridyesi Asya'dan gelmektedir ve öncelikle pelajik besinleri bentoslara ve hızlandırılmış bir mikrobiyal döngüye yönlendirerek LSZ ekolojisinde önemli ve şiddetli değişiklikler yaratmıştır.[68]
Türler, bölgeler arasında römorkla çekilen spor teknelerine bağlanarak tanıtıldı.[67] Bu, bir dizi düşük tuzluluk bitkisinin olası kaynağıdır. Egeria densa ve su sümbülü (Eichhornia crassipes ). Bu bitkiler, su akışını bozarak, fitoplanktonları gölgelendirerek ve balıkçıl gibi balık çizgili levrek, Morone saksati 1800'lerin sonlarında kasıtlı olarak Chesapeake Körfezi.[51][67] Tatlı su quagga midye aslen Avrupa'dan olan, ihtiyati tedbirlere rağmen, kayıkçılar tarafından önümüzdeki iki ila on yıl içinde tanıtılması bekleniyor.[69] Dahası, San Francisco halicinin genç yaşı ve izole ortamı, biyolojik çeşitlilik eksikliği nedeniyle istilacı omurgasızların başarısıyla sonuçlandı.[53]
Gelecek istilalar
Modern besin ağı, bir dizi istiladan ve trofik ikamelerden türetilmiştir.
Yeni organizmalar tesadüfen veya kasıtlı girişlerle geldikçe bu sürecin devam etmesi bekleniyor. Daha az açık olan şey, önceki girişlerin gelecekteki istilalara ne ölçüde yol açtığıdır. Bu, üç yoldan biriyle gerçekleşebilir.
- Erken bir istilacı, yeni bir avcı tanıtılıncaya kadar yeni sistemde kullanılmayan bir kaynak sağlayabilir (L. tetraspina ve yukarıda açıklandığı gibi mikrobiyal döngü).
- Erken istilacılar, habitatları değiştirerek ve daha sonraki istilalar için uygun hale getirerek yenilerini kolaylaştırabilir (substrat için Amur Nehri istiridye kabukları kullanan jöle polipleri).
- Apparent competition between old and new residents may increase the possibilities for invasion and settlement of new organisms that can capitalize on unexploited resources (the subsidization of the Amur River clam by upstream populations of the introduced copepod P. forbesi, creating pressure on native copepods).
Özet
The LSZ food web of the San Francisco Estuary operates in two parallel and asymmetrical directions. The bulk of carbon is assimilated into the benthic and microbial loops, which represent energetic dead ends. A smaller fraction is delivered to higher pelagic trophic levels which may support copepods, fish, birds and fisheries. This redirection of the food web into these two narrow loops may be responsible for the decline in macroinvertebrates and fishes in the estuary, which operate outside of these chains. Restoration of the estuary to a higher degree of function relies upon the probability of delivering increased benefits to the pelagic web without subsidizing the benthic.
Future ecology
The ecology of the Low Salinity Zone of the San Francisco Estuary is difficult to characterize because it is the result of a complex synergy of both abiotic and biotic factors. In addition, it continues to undergo rapid change resulting from newly introduced species, direct anthropogenic influences and iklim değişikliği. Future ecological changes will be driven on an ecosystem wide scale, particularly as Deniz seviyesi yükselmesi, tektonik instability and infrastructure decline cause levee failure in the Delta.[70] The resulting back-surge in water flow is expected to force X2 into the Delta, jeopardizing spatially oriented habitat (like freshwater marshes), channelizing the low salinity zone, and threatening southern California's water supply, with unknown and unforeseeable consequences for the natural and human ecology of the West coast's largest estuary.
Ayrıca bakınız
- San Francisco Körfez Bölgesi'ndeki su yollarının listesi
- San Francisco Körfez Bölgesi Hidrografisi
- Watershed Projesi
- San Francisco Haliç ve Havza Bilimi
Notlar
- ^ a b c d e Kimmerer 2004
- ^ a b Conomos 1979
- ^ Vayssieres 2006
- ^ Jassby 2000
- ^ Deverel 1998
- ^ Philip 2007
- ^ Lund 2007
- ^ a b Kimmerer 2002
- ^ Gunther, Andrew (2011). "The State of the San Francisco Bay 2011, San Francisco Estuary Partnership" (PDF). The State of the San Francisco Bay 2011. Alındı 6 Ekim 2017.
- ^ Moyle 1992
- ^ a b Bennett 2006
- ^ Monismith 1996
- ^ Burau 1998
- ^ Schoellhamer 2001
- ^ Orsi 1986
- ^ a b c d e f Kimmerer 1996
- ^ a b Kimmerer 2006
- ^ Davis, Jay (September 2006). "The Pulse of the Estuary 2006" (PDF). sfei.org. San Francisco Haliç Enstitüsü. Alındı 2 Kasım 2016.
- ^ Taugher 2005
- ^ Smith 2002
- ^ a b Dugdale 2003
- ^ Jassby 2002
- ^ Nichols 1986
- ^ Kimmerer 1994
- ^ a b Jassby and Cloern 2000
- ^ Anderson vd. 2002
- ^ a b Jassby et al. 2002
- ^ Dugdale et al. 2003
- ^ Cloern 1987
- ^ Cole and Cloern 1987
- ^ Ball and Arthur 1979
- ^ Lehman and Waller 2003
- ^ Murrell et al. 1999
- ^ Murrell and Hollibaugh 2000
- ^ Jassby et al. 1993
- ^ Hollibaugh and Wong 1996
- ^ a b Bouley 2006
- ^ Rollwagen-Bollens and Penry 2003
- ^ Rollwagen-Bollens et al. 2006
- ^ Modlin 1997
- ^ Kimmerer 1998
- ^ Lee 1999
- ^ Orsi 1991
- ^ a b Durand2006
- ^ Orsi 1999
- ^ Bouley and Kimmerer 2006
- ^ Orsi and Ohtsuka 1999
- ^ Orsi and Knutson 1979
- ^ Modlin and Orsi 1997
- ^ Sitts and Knight 1979
- ^ a b Radovich 1963
- ^ Peterson 1997
- ^ a b Nichols et al. 1986
- ^ Poulton et al. 2002
- ^ Richman and Lovvorn 2004
- ^ Scientists race to identify goop on birds along San Francisco Bay shorelines (Ocak 2015), Hıristiyan Bilim Monitörü
- ^ a b Skinner, John E. (1962). San Francisco Körfez Bölgesi'ndeki Balık ve Vahşi Yaşam Kaynaklarının Tarihsel İncelemesi (Memeli Kaynakları). California Balık ve Av Hayvanları Bakanlığı, Su Projeleri Şubesi Rapor no. 1. Sacramento, California: California Balık ve Av Hayvanları Bakanlığı. Arşivlenen orijinal 2011-07-26 tarihinde.
- ^ Suzanne Stewart ve Adrian Praetzellis (Kasım 2003). Archeological Research Issues for the Point Reyes National Seashore - Golden Gate National Recreation Area (PDF) (Bildiri). Anthropological Studies Center, Sonoma State University. s. 335. Alındı 10 Ocak 2010.
- ^ Thompson, R. A. (1896). The Russian Settlement in California Known as Fort Ross, Founded 1812...Abandoned 1841: Why They Came and Why They Left. Santa Rosa, California: Sonoma Democrat Publishing Company. s. 7. ISBN 0-559-89342-6.
- ^ White, Peter (1995). The Farallon Islands: Sentinels of the Golden Gate. San Francisco, California: Scottwall Associates. ISBN 0-942087-10-0.
- ^ Tappe Donald T. (1942). "Kaliforniya'daki Kunduzların Durumu". Oyun Bülteni No. 3. Kaliforniya Balık ve Avlanma Bölümü.
- ^ "Dışarı çık!". San Francisco Chronicle. Nisan 1966. Alındı 2011-03-06.
- ^ "Blue Oak Ranch Koruma Alanı". Kaliforniya Üniversitesi. Arşivlenen orijinal 2011-05-12 tarihinde. Alındı 2011-03-06.
- ^ Werner and Hollibaugh 1993
- ^ Carlton et al. 1990
- ^ Nichols et al. 1990
- ^ a b c Carlton 1996
- ^ Cole 1992
- ^ Dey 2007
- ^ Epstein 2006
Referanslar
- Anderson, D. M., P. M. Glibert, and J. M. Burkholder. 2002. Harmful Algal Blooms and Eutrophication: Nutrient Sources, Composition, and Consequences. Estuaries 25:704-726.
- Ball, M. D., and J. F. Arthur. 1979. Planktonic chlorophyll dynamics in the northern San Francisco Bay and Delta. Pages 265–286 in T. J. Conomos, editor. San Francisco Bay: The urbanized estuary. American Association for the Advancement of Science, Pacific Division, San Francisco, CA.
- Bennett, W. A. (2006). Delta Smelt Growth and Survival During the Recent Pelagic Organism Decline: What Causes Them Summer Time Blues? CALFED Science Conference. Sacramento, CA.
- Bouley P., W. J. K. (2006). "Ecology of a highly abundant, introduced cyclopoid copepod in a temperate estuary. " Marine Ecology Progress Series 324: 219–228.
- Burau, J. R. (1998). "Results from the hydrodynamic element of the 1994 entrapment zone study in Suisun Bay, Sacramento, CA. " Interagency Ecological Program for the San Francisco Bay-Delta Estuary Technical Report 56: 13–62.
- Carlton, J. T., J. K. Thompson, L. E. Schemel, and F. H. Nichols. 1990. Remarkable invasion of San Francisco Bay (California, USA) by the Asian clam Potamocorbula amurensis. 1. Introduction and dispersal. Deniz Ekolojisi İlerleme Serisi. 66:81-94.
- Carlton, J. (1996). "Biological Invasions and Cryptogenic Species. " Ecology 77(6).
- Cole, B. E., and J. E. Cloern. 1987. An empirical model for estimating phytoplankton productivity in estuaries. Deniz Ekolojisi İlerleme Serisi. 36:299-305.
- Cole, B. E., J. K. Thompson, and J. E. Cloern (1992). "Measurement of filtration rates by infaunal bivalves in a recirculating flume. " Marine Biology 113: 219–225.
- Cloern, J. E. 1987. Turbidity as a control on phytoplankton biomass and productivity in estuaries. Continental Shelf Research 7:1367-1381.
- Conomos, T. J., Ed. (1979). San Francisco Bay: The urbanized estuary. San Francisco, CA, Pacific Division, American Association for the Advancement of Science.
- Deverel, S. (1998). Subsidence mitigation in the Sacramento-San Joaquin Delta., CALFED Bay-Delta Program.
- Dey, R. (2007, January 16, 2007). "Invasive Mussel Update from Lake Mead National Recreation Area. " 100th Meridian Initiative, from http://100thmeridian[kalıcı ölü bağlantı ]. org/mead. asp.
- Dugdale, R. C., V. Hogue, A. Marchi, A. Lassiter, and F. Wilkerson. 2003. Effects of anthropogenic ammonium input and flow on primary production in the San Francisco Bay. CALFED Science Program, Sacramento, CA.
- Durand, J. (2006). Determinants of calanoid copepod recruitment failure in the San Francisco Estuary Calfed Science Program. Sacramento, CA.
- Epstein, E. (2006). Creaky levees ripe for disaster. San Francisco Chronicle. San Francisco.
- Hollibaugh, J. T., and P. S. Wong. 1996. Distribution and activity of bacterioplankton in San Francisco Bay. Pages 263–288 in J. T. Hollibaugh, editor. San Francisco Bay: The ecosystem. American Association for the Advancement of Science, San Francisco, CA.
- Jassby, A. D., J. E. Cloern, and B. E. Cole. 2002. Annual primary production: Patterns and mechanisms of change in a nutrient-rich tidal ecosystem. Limnology and Oceanography 47:698-712.
- Jassby, A. D., J. E. Cloern, and T. M. Powell. 1993. Organic carbon sources and sinks in San Francisco Bay - variability induced by river flow. Deniz Ekolojisi İlerleme Serisi. 95.
- Jassby, A. D., and J. E. Cloern. 2000. Organic matter sources and rehabilitation of the Sacramento-San Joaquin Delta (California, USA). Aquatic Conservation: Marine and Freshwater Ecosystems 10:323-352.
- Jassby, A. D., J. E. Cloern and B. E. Cole. (2002). "Annual primary production: Patterns and mechanisms of change in a nutrient-rich tidal ecosystem. " Limnology and Oceanography 47(3): 698–712.
- Kimmerer, W. J., E. Gartside, and J. J. Orsi. (1994). "Predation by an introduced clam as the likely cause of substantial declines in zooplankton of San Francisco Bay. " Mar. Ecol. Prog. Ser. 113: 81–93.
- Kimmerer, W. J., J. J. Orsi. 1996. Changes in the zooplankton of the San Francisco Bay estuary since the introduction of the clam Potamocorbula amurensis. Pages 403–425 in J. T. Hollibaugh, editor. San Francisco Bay: The Ecosystem. American Association for the Advancement of Science, San Francisco, CA.
- Kimmerer, W. J., J. R. Burau, and W. A. Bennett. (1998). "Tidally oriented vertical migration and position maintenance of zooplankton in a temperate estuary. " Limnol. Oceanogr. 43(7): 1697–1709.
- Kimmerer, W. J. (2002). "Physical, biological, and management responses to variable freshwater flow into the San Francisco Estuary. " Estuaries 25: 1275–1290.
- Kimmerer, W. (2004). "San Francisco Halici'nin açık su süreci: fiziksel zorlamadan biyolojik tepkilere." San Francisco Haliç ve Su Havzası Bilimi 2 (1).
- Kimmerer, W. (2006). "Response of anchovies dampens effects invasive bivalve Corbula amurensis on San Francisco Estuary foodweb. " Marine Ecology Progress Series 324: 207–218.
- Lee, C. E. (1999). "Rapid and repeated invasions of fresh water by the copepod Eurytermora affinis. " Evolution 53(5): 1423–1434.
- Lehman, P. W., and S. Waller. 2003. Microcystis blooms in the delta. IEP Newsletter 16:18-19.
- Lund, J., E. Hanak, W. Fleenor, R. Howitt, J. Mount, P. Moyle (2007). Envisioning Futures for the Sacramento-San Joaquin Delta. San Francisco, Public Policy Institute.
- Modlin, R. F., and J. J. Orsi (1997). "Acanthomysis bowmani, a new species, and A. asperali, Mysidacea newly reported from the Sacramento-San Joaquin Estuary, California (Crustacea: Mysidae). " Proceedings of the Biological Society of Washington 110(3): 439–446.
- Monismith, S. G., J. R. Burau, and M. Stacey (1996). Stratification dynamics and gravitational circulation on northern San Francisco Bay. San Francisco Bay: The ecosystem. J. T. Hollibaugh. San Francisco, CA, American Association for the Advancement of Science: 123–153.
- Moyle, P. B., B. Herbold, D. E. Stevens, and L. W. Miller. (1992). "Life history and status of delta smelt in the Sacramento-San Joaquin Estuary, California. " Trans. Am. Balık. Soc. 121: 67–77.
- Murrell, M. C., J. T. Hollibaugh, M. W. Silver, and P. S. Wong. 1999. Bacterioplankton dynamics in northern San Francisco Bay: Role of particle association and seasonal freshwater flow. Limnoloji ve Oşinografi. 44:295-308.
- Murrell, M. C., and J. T. Hollibaugh. 2000. Distribution and composition of dissolved and particulate organic carbon in northern San Francisco Bay during low flow conditions. Estuarine, Coastal and Shelf Science 51:75-90.
- Nichols, F. H., J. E. Cloern, S. Luoma, and D. H. Peterson. 1986. The modification of an estuary. Science 231:567-573.
- Nichols, F. H., J. K. Thompson, and L. E. Schemel. 1990. Remarkable invasion of San Francisco Bay (California, USA) by the Asian clam Potamocorbula amurensis. 2. Displacement of a former community. Deniz Ekolojisi İlerleme Serisi. 66:95-101.
- Orsi, J. J., and A. C. Knutson. 1979. The role of mysid shrimp in the Sacramento-San Joaquin Estuary and factors affecting their abundance and distribution. Pages 401–408 in T. J. Conomos, editor. San Francisco Bay: the urbanized estuary. American Association for the Advancement of Science, Pacific Division, San Francisco.
- Orsi, J. J., and W. L. Mecum. (1986). "Zooplankton distribution and abundance in the Sacramento-San Joaquin Delta in relation to certain environmental factors. " Estuaries 9(4 B): 326–339.
- Orsi, J. J., and T. C. Walter. (1991). "Pseudodiaptomus forbesi and P. marinus (Copepoda: Calanoida), the latest copepod immigrants to California's Sacramento-San Joaquin Estuary. " Bull. Plankton Soc. Japan Spec. Cilt (Proceedings of the Fourth International Conference on Copepoda): 553–562.
- Orsi, J. J., and S. Ohtsuka. (1999). "Introduction of the Asian copepods Acartiella sinensis, Tortanus dextrilobatus (Copepoda: Calanoida), and Limnoithona tetraspina (Copepoda: Cyclopoida) to the San Francisco Estuary, California, USA. " Plank. Bio. Ecol. 46: 128–131.
- Peterson, H. 1997. Clam-stuffed sturgeon. IEP Newsletter 9:19-19.
- Philip, T. (2007). A time to deliver water solutions. Sacramento Bee. Sacramento, CA.
- Poulton, V. K., J. R. Lovvorn, and J. Y. Takekawa. 2002. Clam density and scaup feeding behavior in San Pablo Bay, California. Condor. 104:518-527.
- Radovich, J. (1963). "Effects of ocean temperature on the seaward movement of striped bass, Roccus saxatilis, on the Pacific coast. " California Department of Fish and Game 49: 191–205.
- Richman, S. E., and J. R. Lovvorn. 2004. Relative foraging value to lesser scaup ducks of native and exotic clams from San Francisco Bay. Ecological Applications 14:1217-1231.
- Rollwagen-Bollens, G. C., and D. L. Penry. 2003. Feeding dynamics of Acartia sp. in a large, temperate estuary (San Francisco Bay, CA). Marine Ecology Progress Series 257:139-158.
- Rollwagen-Bollens, G., S. Gifford, A. Slaughter, and S. M. Bollens. 2006. Protists in a Temperate Estuary: Diversity, Grazing and Consumption by Metazoans. Eos, Transactions of the American Geophysical Union, Ocean Sciences Meeting Supplement. 87:36 OS11D-05.
- Schoellhamer, D. H. (2001). Influence of salinity, bottom topography, and tides on locations of estuarine turbidity maxima in northern San Francisco Bay. Coastal and Estuarine Fine SedimentProcesses. W. H. McAnally and A. J. Mehta. Amsterdam, Netherlands, Elsevier: 343–356.
- Sitts R. M., A. W. Knight. 1979. Predation by the estuarine shrimps Crangon franciscorum Stimpson and Palaemon macrodactylus Rathbun. Biological Bulletin 156:356-368.
- Smith, S. V., and J. T. Hollibaugh (2002). "Water, salt, and nutrient exchanges in the San Francisco Bay. " Interagency Ecological Program for the San Francisco Bay-Delta Estuary TechnicalReport 66.
- Taugher, Mike (2005). Environmental sirens in Delta are screaming. Contra Costa Times.
- Trott, J. S. S., Warren J. Ferguson, and Andrew J. Kleinfeld, (2006). Central Delta Water Agency v. Bureau of Reclamation. Dokuzuncu Daire için Amerika Birleşik Devletleri Temyiz Mahkemesi. San Francisco, CA.
- Vayssieres, M. P. (2006). Trends in the benthos of the upper San Francisco Esutary. CALFED Science Conference. Sacramento, CA.
- Werner, I., and J. T. Hollibaugh. 1993. Potamocorbula amurensis (Mollusca, Pelecyopoda): Comparison of clearance rates and assimilation efficiencies for phytoplankton and bacterioplankton. Limnology and Oceanography 38:949-964.


