Mercan kayalığı - Coral reef - Wikipedia
| Deniz habitatları |
|---|
 Biyoçeşitlilik mercan kayalığı |
Bir mercan kayalığı bir su altı ekosistem resif yapımı ile karakterize mercanlar. Resifler oluşur koloniler nın-nin mercan polipler tarafından bir arada tutuldu kalsiyum karbonat. Mercan resiflerinin çoğu taş mercanlar, polipleri gruplar halinde kümelenir.
Mercan sınıf Anthozoa hayvanda filum Cnidaria, içerir Deniz lalesi ve Deniz anası. Deniz anemonlarının aksine mercanlar sert karbonat salgılarlar dış iskeletler mercanı destekleyen ve koruyan. Resiflerin çoğu en iyi ılık, sığ, berrak, güneşli ve çalkalanmış suda büyür. Mercan resifleri ilk olarak 485 milyon yıl önce, Erken Ordovisyen, mikrobiyal ve sünger resifleri Kambriyen.[1]
Bazen aranır denizin yağmur ormanları,[2] sığ mercan resifleri, dünyanın en çeşitli ekosistemlerinden bazılarını oluşturur. Dünya okyanus alanının% 0.1'inden azını, Fransa'nın yaklaşık yarısını kaplarlar, ancak tüm denizcilerin en az% 25'ine ev sağlarlar. Türler,[3][4][5][6] dahil olmak üzere balık, yumuşakçalar, solucanlar, kabuklular, ekinodermler, süngerler, tunikatlar ve diğeri cnidarians.[7] Mercan resifleri, az besin sağlayan okyanus sularında gelişir. Genellikle tropikal sularda sığ derinliklerde bulunurlar, ancak derin su ve soğuk su mercan resifleri diğer bölgelerde daha küçük ölçeklerde bulunur.
Mercan resifleri sunar ekosistem servisleri turizm, balıkçılık ve kıyı şeridi koruması. Mercan resiflerinin yıllık küresel ekonomik değerinin 30–375 milyar ABD Doları arasında olduğu tahmin edilmektedir.[8][9] ve 9,9 trilyon ABD doları.[10] Mercan resifleri kırılgandır, çünkü kısmen su koşullarına duyarlıdırlar. Tehdit altındalar fazla besin (azot ve fosfor), artan sıcaklıklar, okyanus asitlenmesi aşırı avlanma (ör. patlama balıkçılığı, siyanür avcılığı, zıpkınla balık avlama açık tüplü dalış ), güneş kremi kullanımı,[11] ve zararlı arazi kullanımı uygulamaları dahil akış ve sızıntılar (ör. enjeksiyon kuyuları ve fosseptikler).[12][13][14]
Oluşumu
Mercan resiflerinin çoğu, Son Buzul Dönemi buz eridiğinde Deniz seviyesi yükselmek ve sel kıta rafları. Mercan resiflerinin çoğu 10.000 yaşın altındadır. Topluluklar kendilerini kurdukça, resifler yükseliyor, hızlanıyor Deniz seviyesi. Çok yavaş yükselen resifler, yeterli ışık olmazsa boğulabilir.[15] Mercan resifleri, denizin derinliklerinde bulunur. kıta rafları, etrafında okyanus adaları ve mercan adaları. Bu adaların çoğu volkanik kökeninde. Diğerleri var tektonik köken nerede tabak hareketleri derin okyanus tabanını kaldırdı.
İçinde Mercan Resiflerinin Yapısı ve Dağılımı,[16] Charles Darwin atol resiflerinin oluşumu hakkındaki teorisini ortaya koydu. Yolculuğu Beagle. O teorileştirdi canlanma ve çökme Dünyanın kabuk okyanusların altında atoller oluşturdu.[17] Darwin, atol oluşumunda üç aşamalı bir dizi oluşturdu. Bir saçak resif soyu tükenmiş bir etrafında oluşur volkanik ada ada ve okyanus tabanı azaldıkça. Çökme devam ettikçe, kenarlardaki resif bir bariyer resifine ve nihayetinde bir atol resifine dönüşür.

Darwin'in teorisi bir volkanik ada yok olan

Ada ve okyanus tabanı çökerken, mercan büyümesi bir saçak resif, genellikle kara ile ana resif arasında sığ bir lagün içerir.

Çökme devam ederken, kıyıdan uzaktaki resif daha büyük ve daha derin bir bariyer resifine dönüşür. lagün içeride.

Nihayetinde ada denizin altına batar ve set resifi bir mercan adası açık bir lagünü çevreleyen.


Darwin, her birinin altında lagün öyle olabilir mi ana kaya taban, orijinal yanardağın kalıntıları.[18] Sonraki araştırmalar bu hipotezi destekledi. Darwin'in teorisi, mercan poliplerinin, tropik suyun çalkalandığı, ancak yalnızca düşük bir derinlik aralığında başlayarak sınırlı bir derinlik aralığında yaşayabildiği gelgit. Altta yatan toprak seviyesinin izin verdiği yerlerde, mercanlar kıyı boyunca büyür ve saçak resifler oluşturur ve sonunda bir bariyer resifine dönüşebilirler.

Dibin yükseldiği yerde, kıyıların etrafında dağınık resifler büyüyebilir, ancak deniz seviyesinin üzerinde yükselen mercanlar ölür. Arazi yavaş yavaş azalırsa, saçaklı resifler, resif ile kara arasında bir lagünü çevreleyen bir bariyer resifini oluşturan, yaşlı, ölü mercanların tabanında yukarı doğru büyüyerek hızlanır. Bir bariyer resifi bir adayı çevreleyebilir ve ada deniz seviyesinin altına battığında, kabaca dairesel bir büyüyen mercan, merkezi bir lagün oluşturarak deniz seviyesine ayak uydurmaya devam eder. Bariyer resifleri ve atoller genellikle tam çember oluşturmazlar, ancak yer yer fırtınalarla kırılırlar. Deniz seviyesinin yükselmesi gibi, hızla düşen bir dip, mercan büyümesini bastırabilir, mercan ve resifi öldürür. mercan boğulma.[20] Güvenen mercanlar zooxanthellae su onlar için çok derin olduğunda ölebilir ortakyaşlar yeterince fotosentez yapmak, azalan ışık maruziyeti nedeniyle.[21]
Belirleyen iki ana değişken jeomorfoloji mercan resiflerinin şekli veya şekli, substrat üzerinde durdukları ve bu alt tabakaya göre deniz seviyesindeki değişimin geçmişi.
Yaklaşık 20.000 yaşındaki Büyük Set Resifi kıta raflarında mercan resiflerinin nasıl oluştuğuna dair bir örnek sunar. Deniz seviyesi 21. yüzyıldakinden 120 m (390 ft) daha alçaktı.[22][23] Deniz seviyesi yükseldikçe, su ve mercanlar Avustralya kıyı düzlüğünün tepeleri olan bölgeyi işgal etti. 13.000 yıl önce, deniz seviyesi şu andan 60 m'ye (200 ft) yükseldi ve kıyı ovalarının birçok tepesi, kıtasal adalar. Deniz seviyesinin yükselmesi devam ederken, su kıtasal adaların çoğunu aştı. Mercanlar daha sonra tepeleri büyütebilir ve cays ve resifler. Büyük Set Resifi'ndeki deniz seviyesi son 6.000 yılda önemli ölçüde değişmedi.[23] Yaşayan resif yapısının yaşının 6.000 ila 8.000 yıl arasında olduğu tahmin edilmektedir.[24] Büyük Set Resifi, volkanik bir ada çevresinde değil, kıta sahanlığı boyunca oluşmuş olsa da, Darwin'in ilkeleri geçerlidir. Avustralya batmak üzere olmadığı için, kalkınma bariyer resif aşamasında durdu. Kıyıdan 300-1.000 m (980-3.280 ft) uzaklıkta 2.000 km (1.200 mi) boyunca uzanan dünyanın en büyük bariyer resifini oluşturdu.[25]
Sağlıklı tropikal mercan resifleri yatay olarak yılda 1 ila 3 cm (0,39 ila 1,18 inç) arasında büyür ve her yıl 1 ila 25 cm (0,39 ila 9,84 inç) arasında dikey olarak büyür; ancak, güneş ışığına ihtiyaç duydukları için yalnızca 150 m'den (490 ft) daha sığ derinliklerde büyürler ve deniz seviyesinin üzerinde büyüyemezler.[26]
Malzeme
Adından da anlaşılacağı gibi, mercan resifleri çoğunlukla sağlam mercan kolonilerinden mercan iskeletlerinden oluşur. Mercanlarda bulunan diğer kimyasal elementler kalsiyum karbonat birikintilerine dahil edildikçe, aragonit oluşturulmuş. Ancak, kabuk parçaları ve kalıntılar mercan yosunu yeşil bölümlü gibi cins Halimeda resifin fırtınalardan ve diğer tehditlerden kaynaklanan hasara karşı koyma yeteneğini artırabilir. Bu tür karışımlar aşağıdaki gibi yapılarda görülebilir Eniwetok Atolü.[27]
Türler
Darwin'in üç klasik resif oluşumunu tanımlamasından bu yana - volkanik bir adanın etrafındaki saçaklı resif, bir bariyer resif ve ardından bir atole dönüşüyor[28] - bilim adamları daha fazla resif türü belirlediler. Bazı kaynaklar yalnızca üç tane bulurken,[29][30] Thomas ve Goudie, dört "ana büyük ölçekli mercan resif türünü" listeliyor - saçaklı resif, bariyer resifi, atol ve masa resifi[31] - Spalding sırasında et al. beş "ana türü" listeleyiniz - dağınık resif, bariyer resifi, atol, "banka veya platform resifi" ve yama resifi.[32]
Saçaklı resif


Bir kıyı resifi olarak da adlandırılan bir saçak resif,[33] doğrudan kıyıya bağlı,[34] veya araya giren dar, sığ bir kanal veya lagünle sınırlar.[35] En yaygın resif türüdür.[35] Saçaklı resifler kıyı şeridini takip eder ve kilometrelerce uzayabilir.[36] Genellikle 100 metreden daha az genişliktedir, ancak bazıları yüzlerce metre genişliğindedir.[37] Saçaklı resifler başlangıçta kıyıda oluşur. alçak su boyut olarak büyüdükçe denizleri düzleştirin ve genişletin. Nihai genişlik, deniz yatağının dik bir şekilde düşmeye başladığı yere bağlıdır. Kenar resifinin yüzeyi genellikle aynı yükseklikte kalır: su seviyesinin hemen altında. Dış bölgeleri denizin derinliklerine doğru ilerleyen daha eski saçaklı resiflerde, iç kısım erozyonla derinleşir ve sonunda bir lagün.[38] Saçaklı resif lagünleri, 100 metreden genişliğe ve birkaç metre derinliğe ulaşabilir. Saçaklı resifin kendisi gibi, kıyıya paralel uzanırlar. Kıyı şeridi resifleri Kızıl Deniz "dünyadaki en gelişmiş yerlerden bazıları" dır ve kumlu koylar dışında tüm kıyılarında bulunur.[39]
Avustralya'nın doğusundaki mercan kayalıkları

Bariyer resifleri, bir anakaradan veya ada kıyısından derin bir kanal veya lagün.[35] Göl kenarındaki bir resifin sonraki aşamalarına benziyorlar, ancak ikincisinden esas olarak boyut ve köken olarak farklılık gösteriyorlar. Lagünleri birkaç kilometre genişliğinde ve 30 ila 70 metre derinliğinde olabilir. Her şeyden önce, açık deniz dış resif kenarı bir kıyı şeridinin yanında değil, açık suda oluşmuştur. Bir atol gibi, bu resiflerin ya deniz tabanı alçaldıkça ya da deniz seviyesi yükseldikçe oluştuğu düşünülmektedir. Oluşum, sınırdaki bir resiften çok daha uzun sürer, bu nedenle bariyer resifleri çok daha nadirdir.
Bir bariyer resifinin en iyi bilinen ve en büyük örneği Avustralya Büyük Set Resifi.[35][40] Diğer önemli örnekler Belize Bariyer Resifi ve Yeni Kaledonya Bariyer Resifi.[40] Bariyer resifleri de kıyılarda bulunur. Providencia,[40] Mayotte, Gambier Adaları güneydoğu kıyısında Kalimantan sahil kesimlerinde Sulawesi, güneydoğu Yeni Gine ve güney sahili Louisiade Takımadaları.
Platform resifi

Çeşitli şekillerde banka veya masa resifleri olarak adlandırılan platform resifleri, kıta sahanlığı açık okyanusta olduğu gibi, aslında deniz tabanının okyanus yüzeyine yeterince yakın yükseldiği ve zooksantemik, resif oluşturan mercanların büyümesini mümkün kılan her yerde.[41] Swain'in güneyindeki Great Barrier Reef'te platform resifleri bulunur[42] ve Oğlak Grubu[43] kıta sahanlığında, kıyıdan yaklaşık 100–200 km. Kuzeydeki bazı platform resifleri Maskarenler anakaradan birkaç bin kilometre uzakta. Sadece deniz kenarına uzanan saçaklı ve bariyer resiflerinin aksine, platform resifleri her yönde büyür.[41] Boyutları birkaç yüz metreden birçok kilometreye kadar değişkendir. Normal şekilleri ovalden uzamasına kadardır. Bu resiflerin bazı kısımları yüzeye ulaşabilir ve etraflarında saçaklı resifler oluşturabilecek kumluklar ve küçük adalar oluşturabilir. Bir platform resifinin ortasında bir lagün oluşabilir.
Platform resifleri atollerde bulunabilir. Orada yama resifleri olarak adlandırılırlar ve çapları yalnızca birkaç düzine metreye ulaşabilirler. Uzatılmış bir yapı üzerinde platform resiflerinin oluştuğu yerlerde, e. g. eski, aşınmış bir bariyer resifi, doğrusal bir düzenleme oluşturabilirler. Bu durum, örneğin, doğu kıyısında Kızıl Deniz yakın Cidde. Eski platform resiflerinde, iç kısım o kadar ağır bir şekilde aşınabilir ki sahte bir atol oluşturur.[41] Bunlar gerçek mercan adalarından ancak muhtemelen karot sondajı da dahil olmak üzere ayrıntılı inceleme ile ayırt edilebilir. Bazı platform resifleri Lakadifler rüzgar ve su akışı nedeniyle U şeklindedir.
Mercan Adası
Atolls veya atol resifleri merkezi bir ada olmadan bir lagünün her yerine uzanan aşağı yukarı dairesel veya sürekli bir bariyer resifidir.[44] Genellikle volkanik adaların etrafındaki dağınık resiflerden oluşurlar.[35] Zamanla ada aşınmalar uzakta ve deniz seviyesinin altına batıyor.[35] Atoller, deniz tabanının batması veya deniz seviyesinin yükselmesi ile de oluşturulabilir. Bir lagünü çevreleyen bir resif halkası ortaya çıkar. Güney Pasifik'te mercan adaları sayısızdır ve genellikle okyanus ortasında meydana gelirler. Caroline Adaları, Cook Adaları, Fransız Polinezyası, Marşal Adaları ve Mikronezya.[40]
Mercan adaları Hint Okyanusu'nda, örneğin Maldivler, Chagos Adaları, Seyşeller Ve çevresinde Cocos Adası.[40] Maldivler'in tamamı 26 atolden oluşmaktadır.[45]
Diğer resif türleri veya çeşitleri


- Apron resifi - saçak bir resifi andıran, ancak daha eğimli olan kısa resif; bir noktadan veya yarımada kıyıdan dışarı ve aşağı doğru uzanan. Bir saçak resifinin ilk aşaması.[33]
- Banka resifi - bir yama resifinden daha büyük olan ve genellikle orta sahanlık bölgelerinde bulunan ve doğrusal veya yarı dairesel şekilli izole, düz tepeli resif; bir tür platform resifi.[40]
- Yama resifi - Yaygın, izole, nispeten küçük resif çıkıntısı, genellikle bir lagün veya gömme, genellikle dairesel ve kumla çevrili veya deniz çayırı. Bir tür platform resifi olarak düşünülebilir[DSÖ? ] ya da çevredeki resiflerin, atollerin ve bariyer resiflerinin özellikleri olarak.[40] Yamalar, deniz çayırı olarak anılan azaltılmış deniz çayırı örtüsü ile çevrelenebilir. otlayan hale.[46]
- Şerit resif - genellikle bir atol lagünüyle ilişkilendirilen uzun, dar, muhtemelen dolambaçlı resif. Raf kenarı resif veya eşik resifi olarak da adlandırılır.[33]
- Habili - özel resif Kızıl Deniz; görünür hale getirmek için yüzeye yeterince yaklaşmıyor sörf; gemiler için bir tehlike olabilir ( Arapça "doğmamış" için)
- Microatoll - mercan türleri topluluğu; ortalama gelgit yüksekliği ile sınırlı dikey büyüme; büyüme morfolojileri, deniz seviyesi değişim modellerinin düşük çözünürlüklü bir kaydını sunar; fosilleşmiş kalıntılar kullanılarak tarihlendirilebilir radyoaktif karbon yaş tayini ve yeniden inşa etmek için kullanıldı Holosen Deniz seviyesi[47]
- Cays - mercan resiflerinin yüzeyinde biriken ve deniz seviyesinin üzerinde bir alan oluşturan aşınmış malzemeden oluşan küçük, düşük yükseklikte kumlu adalar; bitkiler tarafından yaşanabilir hale gelmek için stabilize edilebilir; boyunca tropikal ortamlarda meydana gelir Pasifik, Atlantik ve Hint Okyanusları (Karayipler ve Büyük Set Resifi ve Belize Barrier Reef), yaşanabilir ve tarım arazisi sağladıkları
- Seamount veya Guyot - volkanik bir adadaki mercan resifi çöktüğünde oluşmuştur; deniz dağlarının üst kısımları yuvarlatılmış ve gergiler düzdür; düz adamlar veya masa üstüdalgalar, rüzgarlar ve atmosferik süreçlerin neden olduğu erozyon nedeniyle
Bölgeler

Mercan resif ekosistemleri, farklı habitat türlerine ev sahipliği yapan farklı bölgeler içerir. Genellikle üç büyük bölge tanınır: ön resif, resif tepesi ve arka resif (genellikle resif lagünü olarak anılır).
Üç bölge fiziksel ve ekolojik olarak birbirine bağlıdır. Resif yaşamı ve okyanus süreçleri, deniz suyu, sedimanlar, besinler ve deniz yaşamı.
Mercan resiflerinin çoğu 50 metreden daha derin sularda bulunur. Bazıları, soğuk, besin açısından zengin tropikal kıta raflarında yaşar. yükselen gibi oluşmaz Büyük Set Resifi. Diğerleri, adaları çevreleyen derin okyanusta veya atoller olarak bulunur. Maldivler. Adaları çevreleyen resifler, adalar okyanusa düştüğünde oluşur ve atoller, bir ada deniz yüzeyinin altına düştüğünde oluşur.
Alternatif olarak, Moyle ve Cech altı bölgeyi birbirinden ayırır, ancak çoğu resifte yalnızca bazı bölgeler bulunur.[48]

Resif yüzeyi resifin en sığ kısmıdır. Tabi dalgalanma ve gelgit. Dalgalar sığ alanların üzerinden geçerken, sürgün, yandaki diyagramda gösterildiği gibi. Bu, suyun genellikle çalkalandığı anlamına gelir. Bunlar, mercanların geliştiği kesin durumdur. Işık için yeterlidir fotosentez simbiyotik zooxanthellae tarafından ve çalkalanmış su mercanları beslemek için plankton getirir.
Resif dışı zemin bir resifi çevreleyen sığ deniz tabanıdır. Bu bölge, kıta raflarındaki resiflerin yanında meydana gelir. Tropik adalar ve atoller etrafındaki resifler aniden büyük derinliklere iner ve böyle bir zemine sahip değildir. Genellikle kumludur, zemin genellikle deniz çayırları resif balıkları için önemli yiyecek arama alanlarıdır.
Resif düşüşü ilk 50 metrede uçurumun kenarında barınak bulan resif balıklarının yaşam alanı ve plankton yakındaki suda. Bırakma bölgesi esas olarak okyanus adalarını ve atolleri çevreleyen resifler için geçerlidir.
Resif yüzü resif tabanı veya resif düşüşü üzerindeki bölgedir. Bu bölge genellikle resifin en çeşitli alanıdır. Mercan ve kireçli algler, çatlaklar ve yarıklar gibi koruma sağlayan karmaşık habitatlar ve alanlar sağlar. Omurgasızlar ve epifitik algler diğer organizmalar için yiyeceklerin çoğunu sağlar.[48] Bu ön sahadaki ortak bir özellik, mahmuz ve oluk oluşumları Çökeltinin aşağı doğru taşınmasına hizmet eder.
Resif düzlüğü ana resifin arkasında yer alan, içinde mercan parçaları bulunan kumlu düzlüktür. Bu bölge bir lagünü çevreleyebilir ve koruma alanı olarak hizmet verebilir veya resif ile kıyı arasında uzanabilir ve bu durumda düz, kayalık bir alandır. Balık, mevcut olduğunda onu tercih etme eğilimindedir.[48]
Resif lagünü dalga hareketinden daha az etkilenen ve genellikle küçük resif yamaları içeren, tamamen kapalı bir bölgedir.[48]
Ancak, "mercan resiflerinin topografyası sürekli değişmektedir. Her resif düzensiz yosun yamalarından oluşur. sapsız omurgasızlar ve çıplak kaya ve kum. Bu yamaların boyutu, şekli ve göreceli bolluğu, bir yamayı diğerine tercih eden çeşitli faktörlere yanıt olarak yıldan yıla değişir. Örneğin büyüyen mercan, resiflerin ince yapısında sürekli bir değişim yaratır. Daha büyük ölçekte, tropikal fırtınalar resifin büyük bölümlerini yerle bir edebilir ve kumlu alanlardaki kayaların hareket etmesine neden olabilir. "[49]
Konumlar
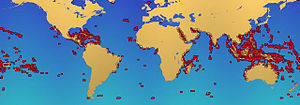
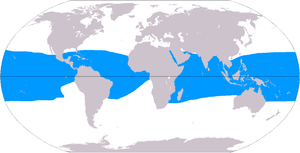

Mercan resiflerinin 284.300 km kapladığı tahmin ediliyor2 (109.800 mil kare),[50] okyanusların yüzey alanının sadece% 0,1'inin biraz altında. Hint-Pasifik bölge (dahil Kızıl Deniz, Hint Okyanusu, Güneydoğu Asya ve Pasifik ) bu toplamın% 91,9'unu oluşturmaktadır. Güneydoğu Asya bu rakamın% 32,3'ünü oluştururken, Pasifik Avustralya % 40,8'dir. Atlantik ve Karayipler mercan resifleri% 7,6'yı oluşturmaktadır.[4]
Mercanlar hem ılıman hem de tropikal sularda bulunmasına rağmen, sığ su resifleri yalnızca ekvatorun yaklaşık 30 ° N'den 30 ° G'sine uzanan bir bölgede oluşur. Tropikal mercanlar 50 metreden (160 ft) daha derinlerde büyümez. Çoğu mercan resifleri için optimum sıcaklık 26–27 ° C'dir (79–81 ° F) ve 18 ° C'nin (64 ° F) altındaki sularda birkaç resif bulunur.[51] Ancak, Basra Körfezi kışın 13 ° C (55 ° F) ve yazın 38 ° C (100 ° F) sıcaklıklara uyum sağlamıştır.[52] 37 tür skleraktin mercan böyle bir ortamda yaşar Larak Adası.[53]
Derin su mercanı Norveç kadar kuzeyde, çok daha yüksek enlemlerde daha büyük derinliklerde ve daha soğuk sıcaklıklarda yaşar.[54] Derin su mercanları resif oluşturabilmesine rağmen, onlar hakkında çok az şey bilinmektedir.
Mercan resifleri, Amerika ve Afrika öncelikle yükselen ve bu bölgelerdeki su sıcaklıklarını düşüren güçlü soğuk kıyı akıntıları ( Peru, Benguela ve Kanarya Akıntıları sırasıyla).[55] Mercanlar nadiren kıyı şeridinde bulunur. Güney Asya - Hindistan'ın doğu ucundan (Chennai ) için Bangladeş ve Myanmar sınırlar[4]- kuzeydoğu kıyılarında olduğu gibi Güney Amerika ve Bangladeş'ten gelen tatlı su çıkışı nedeniyle Amazon ve Ganj Sırasıyla nehirler.
- Büyük Set Resifi - 2.900'den fazla ayrı resif ve 2.600 kilometreden (1.600 mil) fazla uzanan 900 adadan oluşan en büyük alan Queensland, Avustralya
- Mezoamerikan Bariyer Resif Sistemi - en büyük ikinci, 1000 kilometre (620 mil) Isla Contoy ucunda Yucatan yarımadası aşağı Honduras Körfezi Adaları
- Yeni Kaledonya Bariyer Resifi - 1.500 kilometre (930 mil) kapsayan ikinci en uzun çift bariyer resifi
- Andros, Bahamalar Bariyer Resifi - Bahamalar'daki Andros Adası'nın doğu kıyısından sonra üçüncü en büyük Andros ve Nassau
- Kızıl Deniz - 2.000 km'lik (1.240 mil) bir kıyı şeridinde bulunan 6.000 yıllık kıyı resifleri içerir
- Florida Resif Yolu —En büyük kıta ABD resifi ve üçüncü en büyük mercan bariyer resifi, Asker Anahtarı, konumlanmış Biscayne Körfezi, için Kuru Tortugalar Meksika Körfezi'nde[56]
- Kasnak Sırtı - en derin fotosentetik mercan resifi, Florida
- Çevresinde çok sayıda resif Maldivler
- Filipinler Güneydoğu Asya'nın ikinci en büyüğü olan mercan kayalığı alanının 26.000 kilometre kare olduğu tahmin ediliyor. 915 resif balığı türü ve 12'si endemik olmak üzere 400'den fazla skleraktin mercan türü burada bulunur.
- Raja Ampat Adaları içinde Endonezya 's Batı Papua il, bilinen en yüksek deniz çeşitliliğini sunar.[57]
- Bermuda en kuzeydeki mercan resif sistemi ile tanınır. 32 ° 24′K 64 ° 48′W / 32.4 ° K 64.8 ° B. Bu yüksek enlemde mercan resiflerinin varlığı, deniz kıyısının yakınlığından kaynaklanmaktadır. Gulf Stream. Bermuda mercan türleri, Büyük Karayipler'de bulunanların bir alt kümesini temsil eder.[58]
- Dünyanın en kuzeyindeki mercan kayalığı, Japonya'nın bir koyunda yer almaktadır. Tsushima Adası içinde Kore Boğazı.[59]
- Dünyanın en güneydeki mercan resifi Lord Howe Adası, Avustralya'nın doğu kıyısındaki Pasifik Okyanusu'nda.
Mercan

Yaşarken mercanlar koloniler Gömülü küçük hayvanların kalsiyum karbonat kabukları. Mercan başları, adı verilen bireysel hayvanların birikimlerinden oluşur. polipler, çeşitli şekillerde düzenlenmiştir.[60] Polipler genellikle küçüktür, ancak boyutları toplu iğne başından 12 inç'e (30 cm) kadar değişebilir.
Resif yapımı veya hermatipik mercanlar sadece içinde yaşamak fotik bölge (50 m'nin üzerinde), suya yeterli güneş ışığının girdiği derinlik.
Zooxanthellae
Mercan polipleri fotosentez yapmaz, ancak mikroskobik ile simbiyotik bir ilişkiye sahiptir. yosun (Dinoflagellatlar ) cinsin Simbiyodinyum, genellikle şu şekilde anılır zooxanthellae. Bu organizmalar poliplerin dokuları içinde yaşar ve polipi şu şekilde besleyen organik besinler sağlar. glikoz, gliserol ve amino asitler.[61] Bu ilişki nedeniyle mercan resifleri berrak suda çok daha hızlı büyür ve bu da daha fazla güneş ışığı alır. Ortakyaşları olmadan mercan büyümesi önemli resif yapıları oluşturmak için çok yavaş olacaktır. Mercanlar besinlerinin% 90'ına kadar ortakyaşlarından alırlar.[62] Buna karşılık, bir örnek olarak karşılıklılık mercanlar zooxanthellae'yi barındırır, her santimetreküp mercan için ortalama bir milyondur ve sürekli bir karbon dioksit fotosenteze ihtiyaçları var.
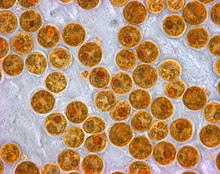



Farklı zooksantel türlerindeki değişen pigmentler, onlara genel olarak kahverengi veya altın-kahverengi bir görünüm verir ve kahverengi mercanlara renklerini verir. Kırmızılar, maviler, yeşiller vb. Gibi diğer pigmentler mercan hayvanları tarafından yapılan renkli proteinlerden gelir. Zooksantellerinin büyük bir kısmını kaybeden mercan beyaz olur (veya bazen mercanlarda kendi proteinleriyle pigmentlenen pastel tonlar) ve olduğu söylenir. ağartılmış düzeltilmezse mercanı öldürebilecek bir durum.
Sekiz tane var Clades nın-nin Simbiyodinyum filotipler. Çoğu araştırma A – D sınıflarında yapılmıştır. Her grup, kendi yararlarının yanı sıra mercan konukçularının hayatta kalmasına daha az uyumlu niteliklere katkıda bulunur. Her fotosentetik organizmanın, proteinler gibi hayatta kalmak için gerekli bileşiklere karşı fotohasarlara karşı belirli bir duyarlılığı vardır. Yenilenme ve çoğaltma hızları, organizmanın hayatta kalma yeteneğini belirler. Filotip A daha çok sığ sularda bulunur. Üretebilir mikosporin benzeri amino asitler bunlar UV ışınlarına dayanıklı türevini kullanarak Gliserin UV radyasyonunu emmek ve daha yüksek su sıcaklıklarına daha iyi uyum sağlamalarını sağlamak için. UV veya termal hasar durumunda, onarım meydana gelirse ve meydana geldiğinde, konağın ve simbiyonun hayatta kalma olasılığını artıracaktır. Bu, evrimsel olarak, A sınıfının diğer sınıflardan daha UV dirençli ve termal olarak daha dirençli olduğu fikrine yol açar.[64]
Clades B ve C, daha derin sularda daha sık bulunur ve bu, artan sıcaklıklara karşı daha yüksek hassasiyetlerini açıklayabilir. Çalılıklarda bulundukları için daha az güneş ışığı alan karasal bitkiler, B, C ve D sınıflarına benzer. . UV dalga boylarında yüksek absorpsiyon oranları ile bu filotipler, sığ clade A'ya kıyasla mercan ağartmaya daha yatkındır.
Clade D'nin yüksek sıcaklığa toleranslı olduğu ve modern dönemlerde B ve C sınıflarından daha yüksek bir hayatta kalma oranına sahip olduğu gözlemlenmiştir. beyazlatma olayları.[64]
İskelet
Resifler polipler ve diğer organizmalar kalsiyum karbonat biriktirdikçe büyür,[65][66] Mercanların temeli, altlarında ve çevrelerinde iskelet yapısı olarak mercan başının tepesini yukarı ve dışa doğru iter.[67] Dalgalar, otlayan balıklar (ör. papağan balığı ), Deniz kestaneleri, süngerler ve diğer kuvvetler ve organizmalar, Bioeroders mercan iskeletlerini resif yapısındaki boşluklara yerleşen veya ilgili resif lagünlerinde kumlu taban oluşturan parçalara ayırır.
Mercan için tipik şekiller Türler karasal nesnelere benzerlikleriyle adlandırılırlar. buruşuk beyinler lahana masa başında, boynuzları, tel şeritleri ve sütunlar. Bu şekiller, ışığa maruz kalma ve dalga hareketi gibi mercanın yaşam geçmişine bağlı olabilir.[68] ve kırılmalar gibi olaylar.[69]
Üreme
Mercanlar hem eşeyli hem de eşeysiz olarak ürerler. Bireysel bir polip, yaşamı boyunca her iki üreme modunu da kullanır. Mercanlar, iç veya dış döllenmeyle cinsel olarak çoğalırlar. Üreme hücreleri, Mezenterler mide boşluğunu çevreleyen doku katmanından içeri doğru yayılan zarlar. Bazı yetişkin mercanlar hermafrodittir; diğerleri sadece erkek veya kadındır. Birkaç Türler büyüdükçe cinsiyeti değiştirin.
İçten döllenmiş yumurtalar, polipte günlerden haftalara kadar değişen bir süre boyunca gelişir. Sonraki gelişme, küçük bir larva, olarak bilinir planula. Senkronize yumurtlama sırasında dışarıdan döllenmiş yumurtalar gelişir. Resifteki polipler, aynı anda yumurta ve spermi toplu halde suya bırakır. Geniş bir alana yayılır. Yumurtlamanın zamanlaması yılın zamanına, su sıcaklığına ve gelgit ve ay döngülerine bağlıdır. Yumurtlama, yüksek ve düşük arasındaki küçük varyasyon göz önüne alındığında en başarılıdır gelgit. Ne kadar az su hareketi olursa, döllenme şansı o kadar artar. İdeal zamanlama ilkbaharda gerçekleşir. Yumurta veya planula salımı genellikle geceleri meydana gelir ve bazen ay döngüsü ile aynı fazdadır (dolunaydan üç ila altı gün sonra). Serbest bırakılmasından yerleşime kadar geçen süre yalnızca birkaç gün sürer, ancak bazı planula'lar birkaç hafta boyunca ayakta kalabilir. Bu işlem sırasında larvalar yerleşim için uygun bir yer bulmak için birkaç farklı ipucu kullanabilir. Uzun mesafelerde, mevcut resiflerden gelen sesler muhtemelen önemlidir,[70] kısa mesafelerde kimyasal bileşikler önemli hale gelir.[71] Larvalar avlanma ve çevresel koşullara karşı savunmasızdır. Alt tabakaya başarıyla yapışan şanslı birkaç planula daha sonra yiyecek ve alan için rekabet eder.[kaynak belirtilmeli ]
Diğer resif yapıcıları
Mercanlar en muhteşem resif yapıcılarıdır. Bununla birlikte, resif topluluğunda yaşayan diğer birçok organizma, mercanlarla aynı şekilde iskelet kalsiyum karbonatına katkıda bulunur. Bunlar arasında mercan yosunu ve bazı süngerler.[72] Resifler her zaman bu farklıların ortak çabaları ile inşa edilir. filum, farklı organizmaların farklı bölgelerde resif inşasına yol açmasıyla jeolojik dönemler.[kaynak belirtilmeli ]
Mercan yosunu

Mercan yosunu resif yapısına önemli katkı sağlayanlardır. Mineral biriktirme hızları mercanlardan çok daha yavaş olsa da, sert dalga hareketine daha toleranslıdırlar ve bu nedenle resifin en büyük kuvvetlere maruz kalan resif kısımları üzerinde koruyucu bir kabuk oluşturmaya yardımcı olurlar. açık okyanus. Ayrıca resif yüzeyinin üzerine tabakalar halinde kireçtaşı bırakarak resif yapısını güçlendirirler.[kaynak belirtilmeli ]
Süngerler

"Sklerospong "herkes için açıklayıcı isimdir Porifera o yapı resifler. Erken Kambriyen dönemi, Archaeocyatha süngerler dünyanın ilk resif oluşturan organizmalarıydı ve süngerler, o döneme kadar tek resif yapıcıydı. Ordovisyen. Sklerosponglar hala mercanların modern resifler inşa etmesine yardımcı oluyor, ancak mercan yosunu mercanlardan çok daha yavaş büyürler ve katkıları (genellikle) küçüktür.[kaynak belirtilmeli ]
Kuzey Pasifik Okyanusu'nda bulut süngerleri Yapılar tropikal resifler gibi yüzeyden tanınmasa da, yine de merceksiz derin su mineral yapıları oluşturur. Onlar tek kaybolmamış soğuk suda resif benzeri yapılar inşa ettiği bilinen organizmalar.[kaynak belirtilmeli ]
Resif yapan mercanlar ve resif inşa eden asistanlarının galerisi

Floresan mercan[73]

Spiral tel mercan

Mercan yosunu Mezofilum sp.

Mercan yosunlarını saran

mercan yosunu Corallina officinalis











Darwin paradoksu
Son oşinografik araştırmalar, bu paradoksun gerçekliğini gün ışığına çıkardı. oligotrofi okyanusun öfotik bölge, kabarık resif tepesine kadar varlığını sürdürür. Açık denizin quasidesert'inden resif kenarlarına ve atollere yaklaştığınızda, canlı maddenin neredeyse yokluğu birdenbire, geçiş olmaksızın bir yaşam bolluğu haline gelir. Öyleyse neden hiçbir şey yerine bir şey var ve daha doğrusu, bu olağanüstü mercan resif makinesinin çalışması için gerekli besinler nereden geliyor? "
- Francis Rougerie[74]İçinde Mercan Resiflerinin Yapısı ve Dağılımı Darwin, 1842'de yayınlanan bir makalesinde, açık bir neden olmaksızın mercan resiflerinin bazı tropik bölgelerde bulunup diğerlerinde bulunmadığını açıkladı. En büyük ve en güçlü mercanlar, resifin en şiddetli sörflere maruz kalan kısımlarında büyüdü ve mercanlar, gevşek tortunun biriktiği yerlerde zayıfladı veya yoktu.[75]
Tropikal sular çok az besin içerir[76] yine de bir mercan kayalığı "çölde bir vaha" gibi gelişebilir.[77] Bu, bazen "Darwin paradoksu" olarak adlandırılan ekosistem muammasına yol açtı: "Bu kadar yüksek üretim, bu kadar besin değeri düşük koşullarda nasıl gelişebilir?"[78][79][80]
Mercan resifleri, tüm deniz türlerinin dörtte birinden fazlasını barındırır. Bu çeşitlilik, karmaşık besin ağları, büyük yırtıcı balık daha küçük yemek yem balığı daha küçük yemek Zooplankton ve benzeri. Bununla birlikte, tüm besin ağları sonunda bitkiler hangileri birincil üreticiler. Mercan resifleri tipik olarak günde metrekare başına 5–10 gram karbon üretir (gC · m−2·gün−1) biyokütle.[81][82]
Tropikal suların olağandışı berraklığının bir nedeni, besin eksiklikleri ve sürüklenmeleridir. plankton. Dahası, güneş tropik bölgelerde yıl boyunca parlayarak yüzey katmanını ısıtır ve onu yeraltı katmanlarından daha az yoğun hale getirir. Daha sıcak su, daha derin, daha soğuk sudan, sabit bir su ile ayrılır. termoklin, sıcaklığın hızlı bir değişim yaptığı yerde. Bu, ılık yüzey sularının daha soğuk ve daha derin suların üzerinde yüzmesini sağlar. Okyanusun çoğu bölgesinde, bu katmanlar arasında çok az değişim vardır. Su ortamlarında ölen organizmalar genellikle dibe çöker, burada ayrışırlar ve besinleri şu şekilde salarlar. azot (N), fosfor (P) ve potasyum (K). Bu besinler bitki büyümesi için gereklidir, ancak tropik bölgelerde doğrudan yüzeye geri dönmezler.[kaynak belirtilmeli ]
Bitkiler besin zincirinin temelini oluşturur ve büyümek için güneş ışığına ve besin maddelerine ihtiyaç duyar. Okyanusta, bu bitkiler çoğunlukla mikroskobiktir. fitoplankton hangi sürüklenme su sütunu. Güneş ışığına ihtiyaçları var fotosentez hangi güçler karbon fiksasyonu, bu nedenle sadece nispeten yüzeye yakın bulunurlar, ancak besin maddelerine de ihtiyaçları vardır. Fitoplankton, besinleri yüzey sularında hızla kullanır ve tropik bölgelerde, bu besinler genellikle termoklin.[83]

Açıklamalar
Mercan resiflerinin etrafındaki lagünler, resiften ve adadan aşınmış malzemelerle dolar. Deniz yaşamı için sığınaklar haline gelirler, dalgalardan ve fırtınalardan koruma sağlarlar.
En önemlisi resifler geri dönüştürmek açık okyanusta çok daha az olan besinler. Mercan resiflerinde ve lagünlerde üreticiler, fitoplanktonun yanı sıra deniz yosunu ve mercan yosunu, özellikle de besinleri mercanlara geçiren çim algleri adı verilen küçük türleri içerir.[84] Fitoplankton, besin zincirinin temelini oluşturur ve balıklar ve kabuklular tarafından yenir. Geri dönüşüm, toplumu desteklemek için genel olarak ihtiyaç duyulan besin girdilerini azaltır.[62]

Mercanlar ayrıca inorganik nitrojen ve fosfor da dahil olmak üzere besinleri doğrudan sudan emer. Birçok mercan, yakalamak için geceleri dokunaçlarını uzatır. Zooplankton yakın geçer. Zooplankton, polipe nitrojen sağlar ve polip, nitrojenin bir kısmını zooxanthellae ile paylaşır ve bu da bu elementi gerektirir.[84]
Resiflerdeki yarıklarda süngerler yaşar. Etkilidirler Filtre besleyicileri, Ve içinde Kızıl Deniz sürüklenen fitoplanktonun yaklaşık% 60'ını tüketirler. Süngerler sonunda besinleri mercanların kullanabileceği bir biçimde salgılar.[85]

Mercan yüzeylerinin sertliği, çalkalanmış sularda mercanların hayatta kalmasının anahtarıdır. Normalde, durgun sudan oluşan bir sınır tabakası, bir bariyer görevi gören batık bir nesneyi çevreler. Mercanların aşırı pürüzlü kenarlarında kırılan dalgalar, sınır tabakasını bozarak mercanların geçen besin maddelerine erişmesine izin verir. Turbulent water thereby promotes reef growth. Without the access to nutrients brought by rough coral surfaces, even the most effective recycling would not suffice.[86]
Deep nutrient-rich water entering coral reefs through isolated events may have significant effects on temperature and nutrient systems.[87][88] This water movement disrupts the relatively stable thermocline that usually exists between warm shallow water and deeper colder water. Temperature regimes on coral reefs in the Bahamas and Florida are highly variable with temporal scales of minutes to seasons and spatial scales across depths.[89]
Water can pass through coral reefs in various ways, including current rings, surface waves, internal waves and tidal changes.[87][90][91][92] Movement is generally created by tides and wind. As tides interact with varying bathymetry and wind mixes with surface water, internal waves are created. An internal wave is a gravity wave that moves along density stratification within the ocean. When a water parcel encounters a different density it oscillates and creates internal waves.[93] While internal waves generally have a lower frequency than surface waves, they often form as a single wave that breaks into multiple waves as it hits a slope and moves upward.[94] This vertical breakup of internal waves causes significant diapycnal mixing and turbulence.[95][96] Internal waves can act as nutrient pumps, bringing plankton and cool nutrient-rich water to the surface.[87][92][97][98][99][100][101][102][103][104][105]
The irregular structure characteristic of coral reef bathymetry may enhance mixing and produce pockets of cooler water and variable nutrient content.[106] Arrival of cool, nutrient-rich water from depths due to internal waves and tidal bores has been linked to growth rates of suspension feeders and benthic algae[92][105][107] as well as plankton and larval organisms.[92][108] The seaweed Codium isthmocladum reacts to deep water nutrient sources because their tissues have different concentrations of nutrients dependent upon depth.[105] Aggregations of eggs, larval organisms and plankton on reefs respond to deep water intrusions.[99] Similarly, as internal waves and bores move vertically, surface-dwelling larval organisms are carried toward the shore.[108] This has significant biological importance to cascading effects of food chains in coral reef ecosystems and may provide yet another key to unlocking the paradox.
Siyanobakteriler provide soluble nitratlar üzerinden nitrojen fiksasyonu.[109]
Coral reefs often depend on surrounding habitats, such as seagrass meadows ve mangrov ormanları, for nutrients. Seagrass and mangroves supply dead plants and animals that are rich in nitrogen and serve to feed fish and animals from the reef by supplying wood and vegetation. Reefs, in turn, protect mangroves and seagrass from waves and produce tortu in which the mangroves and seagrass can root.[52]
Biyoçeşitlilik



Coral reefs form some of the world's most productive ecosystems, providing complex and varied marine habitats that support a wide range of other organisms.[110][111] Saçaklı resifler hemen aşağıda düşük gelgit level have a mutually beneficial relationship with mangrov forests at high tide level and deniz otu meadows in between: the reefs protect the mangroves and seagrass from strong currents and waves that would damage them or aşındırmak the sediments in which they are rooted, while the mangroves and sea grass protect the coral from large influxes of alüvyon, fresh water and kirleticiler. This level of variety in the environment benefits many coral reef animals, which, for example, may feed in the sea grass and use the reefs for protection or breeding.[112]
Reefs are home to a variety of animals, including fish, Deniz kuşları, süngerler, cnidarians (which includes some types of corals and Deniz anası ), solucanlar, kabuklular (dahil olmak üzere karides, temizleyici karides, spiny lobsters ve Yengeçler ), yumuşakçalar (dahil olmak üzere kafadanbacaklılar ), ekinodermler (dahil olmak üzere denizyıldızı, Deniz kestaneleri ve deniz hıyarı ), deniz dölleri, Deniz kaplumbağaları ve deniz yılanları. Aside from humans, memeliler are rare on coral reefs, with visiting deniz memelileri gibi yunuslar the main exception. A few species feed directly on corals, while others graze on algae on the reef.[4][84] Reef biomass is positively related to species diversity.[113]
The same hideouts in a reef may be regularly inhabited by different species at different times of day. Nighttime predators such as cardinalfish ve squirrelfish hide during the day, while damselfish, surgeonfish, tetik balığı, Wrasses ve papağan balığı hide from yılanbalığı ve köpekbalıkları.[27]:49
The great number and diversity of hiding places in coral reefs, i.e. sığınaklar, are the most important factor causing the great diversity and high biyokütle of the organisms in coral reefs.[114][115]
Yosun
Reefs are chronically at risk of algal encroachment. Overfishing and excess nutrient supply from onshore can enable algae to outcompete and kill the coral.[116][117] Increased nutrient levels can be a result of sewage or chemical fertilizer runoff. Runoff can carry nitrogen and phosphorus which promote excess algae growth. Algae can sometimes out-compete the coral for space. The algae can then smother the coral by decreasing the oxygen supply available to the reef.[118] Decreased oxygen levels can slow down calcification rates, weakening the coral and leaving it more susceptible to disease and degradation.[119] Algae inhabit a large percentage of surveyed coral locations.[120] The algal population consists of turf algae, mercan yosunu ve macro algae. Some sea urchins (such as Diadema antillarum ) eat these algae and could thus decrease the risk of algal encroachment.
Süngerler
Sponges are essential for the functioning of the coral reef that system. Algae and corals in coral reefs produce organic material. This is filtered through sponges which convert this organic material into small particles which in turn are absorbed by algae and corals.[121]
Balık
Over 4,000 species of fish inhabit coral reefs.[4] The reasons for this diversity remain unclear. Hypotheses include the "lottery", in which the first (lucky winner) recruit to a territory is typically able to defend it against latecomers, "competition", in which adults compete for territory, and less-competitive species must be able to survive in poorer habitat, and "predation", in which population size is a function of postsettlement piscivore mortality.[122] Healthy reefs can produce up to 35 tons of fish per square kilometer each year, but damaged reefs produce much less.[123]
Omurgasızlar
Sea urchins, Dotidae ve deniz sümüklü böcekleri eat seaweed. Some species of sea urchins, such as Diadema antillarum, can play a pivotal part in preventing algae from overrunning reefs.[124] Researchers are investigating the use of native collector urchins, Tripneustes gratilla, for their potential as biocontrol agents to mitigate the spread of invasive algae species on coral reefs.[125][126] Nudibranchia ve Deniz lalesi eat sponges.
A number of invertebrates, collectively called "cryptofauna," inhabit the coral skeletal substrate itself, either boring into the skeletons (through the process of bioerosion ) or living in pre-existing voids and crevices. Animals boring into the rock include sponges, çift kabuklu mollusks, and Sipunculans. Those settling on the reef include many other species, particularly crustaceans and polychaete solucanlar.[55]
Deniz kuşları
Coral reef systems provide important habitats for deniz kuşu species, some endangered. Örneğin, Midway Atolü içinde Hawaii supports nearly three million seabirds, including two-thirds (1.5 million) of the global population of Laysan albatros, and one-third of the global population of black-footed albatross.[127] Each seabird species has specific sites on the atoll where they nest. Altogether, 17 species of seabirds live on Midway. kısa kuyruklu albatros is the rarest, with fewer than 2,200 surviving after excessive feather hunting in the late 19th century.[128]
Diğer
Deniz yılanları feed exclusively on fish and their eggs.[129][130][131] Marine birds, such as balıkçıl, gannets, pelikanlar ve göğüs, feed on reef fish. Some land-based sürüngenler intermittently associate with reefs, such as kertenkeleleri izlemek, marine crocodile and semiaquatic snakes, such as Laticauda colubrina. Deniz kaplumbağaları, özellikle hawksbill deniz kaplumbağaları, feed on sponges.[132][133][134]

Soft coral, cup coral, sponges and ascidians

Kabuğu Latiaxis wormaldi, a coral snail









Ekosistem servisleri
Coral reefs deliver ekosistem servisleri to tourism, fisheries and coastline protection. The global economic value of coral reefs has been estimated to be between US$29.8 billion[8] and $375 billion per year.[9]
The economic cost over a 25-year period of destroying one kilometer of coral reef has been estimated to be somewhere between $137,000 and $1,200,000.[135]
To improve the management of coastal coral reefs, the Dünya Kaynakları Enstitüsü (WRI) developed and published tools for calculating the value of coral reef-related tourism, shoreline protection and fisheries, partnering with five Caribbean countries. As of April 2011, published working papers covered St. Lucia, Tobago, Belize, ve Dominik Cumhuriyeti. The WRI was "making sure that the study results support improved coastal policies and management planning".[136] The Belize study estimated the value of reef and mangrove services at $395–559 million annually.[137]
Bermuda's coral reefs provide economic benefits to the Island worth on average $722 million per year, based on six key ecosystem services, according to Sarkis ve diğerleri (2010).[138]
Shoreline protection
Coral reefs protect shorelines by absorbing wave energy, and many small islands would not exist without reefs. Coral reefs can reduce wave energy by 97%, helping to prevent loss of life and property damage. Coastlines protected by coral reefs are also more stable in terms of erosion than those without. Reefs can attenuate waves as well as or better than artificial structures designed for kıyı savunması such as breakwaters.[139] An estimated 197 million people who live both below 10 m elevation and within 50 km of a reef consequently may receive risk reduction benefits from reefs. Restoring reefs is significantly cheaper than building artificial breakwaters in tropical environments. Expected damages from flooding would double, and costs from frequent storms would triple without the topmost meter of reefs. For 100-year storm events, flood damages would increase by 91% to $US 272 billion without the top meter.[140]
Balıkçılık
About six million tons of fish are taken each year from coral reefs. Well-managed reefs have an average annual yield of 15 tons of seafood per square kilometer. Southeast Asia's coral reef fisheries alone yield about $2.4 billion annually from seafood.[135]
Tehditler

Since their emergence 485 million years ago, coral reefs have faced many threats, including disease,[142] predation,[143] invasive species, bioerosion by grazing fish,[144] alg çiçekleri, jeolojik tehlikeler, and recent human activity.
This include coral mining, dip trolü,[145] and the digging of kanallar and accesses into islands and bays, all of which can damage marine ecosystems if not done sustainably. Other localized threats include blast fishing, aşırı avlanma, coral overmining,[146] ve Deniz kirliliği, including use of the banned anti-fouling biyosit tributyltin; although absent in developed countries, these activities continue in places with few environmental protections or poor regulatory enforcement.[147][148][149] Chemicals in sunscreens may awaken latent viral infections in zooxanthellae[11] and impact reproduction.[150] However, concentrating tourism activities via offshore platforms has been shown to limit the spread of coral disease by tourists.[151]
Sera gazı emissions present a broader threat through sea temperature rise and sea level rise,[152] though corals adapte olmak their calcifying fluids to changes in deniz suyu pH and carbonate levels and are not directly threatened by okyanus asitlenmesi.[153] Volcanic and manmade aerosol pollution can modulate regional sea surface temperatures.[154]
In 2011, two researchers suggested that "extant marine invertebrates face the same synergistic effects of multiple stressors" that occurred during the end-Permian extinction, and that genera "with poorly buffered respiratory physiology and calcareous shells", such as corals, were particularly vulnerable.[155][156][157]

Corals respond to stress by "bleaching," or expelling their colorful zooksantellat endosymbionts. Corals with Clade C zooxanthellae are generally vulnerable to heat-induced bleaching, whereas corals with the hardier Clade A or D are generally resistant,[158] as are tougher coral genera like Porit ve Montipora.[159]
Every 4–7 years, an El Niño event causes some reefs with heat-sensitive corals to bleach,[160] with especially widespread bleachings in 1998 and 2010.[161][162] However, reefs that experience a severe bleaching event become resistant to future heat-induced bleaching,[163][164][159] due to rapid yönlü seçim.[164] Similar rapid adaption may protect coral reefs from global warming.[165]
A large-scale systematic study of the Jarvis Adası coral community, which experienced ten El Niño-coincident mercan ağartma events from 1960 to 2016, found that the reef recovered from almost complete death after severe events.[160]
Koruma

Deniz koruma alanları (MPAs) are areas designated because they provide various kinds of protection to ocean and/or estuarine areas. They are intended to promote responsible fishery management ve habitat protection. MPAs can also encompass social and biological objectives, including reef restoration, aesthetics, biodiversity and economic benefits.
The effectiveness of MPAs is still debated. For example, a study investigating the success of a small number of MPAs in Endonezya, Filipinler ve Papua Yeni Gine found no significant differences between the MPAs and an unprotected sites.[166][167] Furthermore, in some cases they can generate local conflict, due to a lack of community participation, clashing views of the government and fisheries, effectiveness of the area and funding.[168] In some situations, as in the Phoenix Adaları Koruma Alanı, MPAs provide revenue to locals. The level of income provides is similar to the income they would have generated without controls.[169] Overall, it appears the MPA's can provide protection to local coral reefs, but that clear management and sufficient funds are required.
The Caribbean Coral Reefs - Status Report 1970–2012, states that coral decline may be reduced or even reversed. For this aşırı avlanma needs to be stopped, especially fishing on species key to coral reefs, gibi papağan balığı. Direct human pressure on coral reefs should also be reduced and the inflow of kanalizasyon should be minimised. Measures to achieve this could include restricting coastal settlement, development and turizm. The report shows that healthier reefs in the Caribbean are those with large, healthy populations of parrotfish. These occur in countries that protect parrotfish and other species, like Deniz kestaneleri. They also often ban banning fish trapping ve zıpkınla balık avlama. Together these measures help creating "resilient reefs".[170][171]
Protecting networks of diverse and healthy reefs, not only climate refugia, helps ensure the greatest chance of genetik çeşitlilik, which is critical for coral to adapt to new climates.[172] A variety of conservation methods applied across marine and terrestrial threatened ecosystems makes coral adaption more likely and effective.[172]
Designating a reef as a biyosfer rezervi, deniz parkı, ulusal anıt veya Dünya Mirası site can offer protections. For example, Belize's barrier reef, Sian Ka'an, Galapagos islands, Büyük Set Resifi, Henderson Adası, Palau ve Papahānaumokuākea Ulusal Denizcilik Anıtı are world heritage sites.[173]
In Australia, the Great Barrier Reef is protected by the Great Barrier Reef Marine Park Authority, and is the subject of much legislation, including a biodiversity action plan.[174] Australia compiled a Coral Reef Resilience Action Plan. This plan consists of uyarlanabilir yönetim strategies, including reducing carbon footprint. A public awareness plan provides education on the "rainforests of the sea" and how people can reduce carbon emissions.[175]
Inhabitants of Ahus Island, Manus Bölgesi, Papua Yeni Gine, have followed a generations-old practice of restricting fishing in six areas of their reef lagoon. Their cultural traditions allow line fishing, but no net or spear fishing. Her ikisi de biyokütle and individual fish sizes are significantly larger than in places where fishing is unrestricted.[176][177]
Increased levels of atmospheric CO2 contribute to ocean acidification, which in turn damages coral reefs. To help combat ocean acidification, several countries have put laws in place to reduce greenhouse gases such as carbon dioxide. Many land use laws aim to reduce CO2 emissions by limiting deforestation. Deforestation can release significant amounts of CO2 absent sequestration via active follow-up forestry programs. Deforestation can also cause erosion, which flows into the ocean, contributing to ocean acidification. Incentives are used to reduce miles traveled by vehicles, which reduces carbon emissions into the atmosphere, thereby reducing the amount of dissolved CO2 in the ocean. State and federal governments also regulate land activities that affect coastal erosion.[178] High-end satellite technology can monitor reef conditions.[179]
Birleşik Devletler Temiz Su Yasası puts pressure on state governments to monitor and limit run-off of polluted water.
Restorasyon
Coral reef restoration has grown in prominence over the past several decades because of the unprecedented reef die offs around the planet. Coral stressors can include pollution, warming ocean temperatures, extreme weather events, and overfishing. With the deterioration of global reefs, fish nurseries, biodiversity, coastal development and livelihood, and natural beauty are under threat. Fortunately, researchers have taken it upon themselves to develop a new field, coral restoration, in the 1970s-1980s[180]
Coral farming
Mercan yetiştiriciliği, also known as coral farming or coral gardening, is showing promise as a potentially effective tool for restoring coral reefs.[181][182][183] The "gardening" process bypasses the early growth stages of corals when they are most at risk of dying. Coral seeds are grown in nurseries, then replanted on the reef.[184] Coral is farmed by coral farmers whose interests range from reef koruma to increased income. Due to its straight forward process and substantial evidence of the technique having a significant effect on coral reef growth, coral nurseries became the most widespread and arguably the most effective method for coral restoration.[185]

Coral gardens take advantage of a coral's natural ability to fragment and continuing to grow if the fragments are able to anchor themselves onto new substrates. This method was first tested by Baruch Rinkevich [186] in 1995 which found success at the time. By today's standards, coral farming has grown into a variety of different forms, but still have the same goals of cultivating corals. Consequently, coral farming quickly replaced previously used transplantation methods, or the act of physically moving sections or whole colonies of corals into a new area.[185] Transplantation has seen success in the past and decades of experiments have led to a high success and survival rate. However, this method still requires the removal of corals from existing reefs. With the current state of reefs, this kind of method should generally be avoided if possible. Saving healthy corals from eroding substrates or reefs that are doomed to collapse could be a major advantage of utilizing transplantation.
Coral gardens generally take on the safe forms no matter where you go. It begins with the establishment of a nursery where operators can observe and care for coral fragments.[185] It goes without saying that nurseries should be established in areas that are going to maximize growth and minimize mortality. Floating offshore coral trees or even aquariums are possible locations where corals can grow. After a location has been determined, collection and cultivation can occur.
The major benefit for using coral farms is it lowers polyp and juvenile mortality rates. By removing predators and recruitment obstacles, corals are able to mature without much hindrance. However, nurseries cannot stop climate stressors. Warming temperatures or hurricanes can still disrupt or even kill nursery corals.
Creating substrates

Efforts to expand the size and number of coral reefs generally involve supplying substrate to allow more corals to find a home. Substrate materials include discarded vehicle tires, scuttled ships, subway cars and formed concrete, such as reef balls. Reefs grow unaided on marine structures such as petrol kuleleri. In large restoration projects, propagated hermatipik mercan on substrate can be secured with metal pins, superglue veya milliput. Needle and thread can also attach A-hermatype coral to substrate.
Biorock is a substrate produced by a patented process that runs low voltage elektrik akımları through seawater to cause dissolved minerals to precipitate onto çelik yapılar. The resultant white carbonate (aragonit ) is the same mineral that makes up natural coral reefs. Corals rapidly colonize and grow at accelerated rates on these coated structures. The electrical currents also accelerate formation and growth of both chemical limestone rock and the skeletons of corals and other shell-bearing organisms, such as oysters. Çevresi anot ve katot provides a high-pH environment which inhibits the growth of competitive filamentous and fleshy algae. The increased growth rates fully depend on the accretion activity. Under the influence of the electric field, corals display an increased growth rate, size and density.
Simply having many structures on the ocean floor is not enough to form coral reefs. Restoration projects must consider the complexity of the substrates they are creating for future reefs. Researchers conducted an experiment near Ticao Island in the Philippines in 2013[187] where several substrates in varying complexities were laid in the nearby degraded reefs. Large complexity consisted of plots that had both a man-made substrates of both smooth and rough rocks with a surrounding fence, medium consisted of only the man-made substrates, and small had neither the fence or substrates. After one month, researchers found that there was positive correlation between structure complexity and recruitment rates of larvae.[187] The medium complexity performed the best with larvae favoring rough rocks over smooth rocks. Following one year of their study, researchers visited the site and found that many of the sites were able to support local fisheries. They came to the conclusion that reef restoration could be done cost-effectively and will yield long term benefits given they are protected and maintained.[187]
Yer değiştirme
One case study with coral reef restoration was conducted on the island of Oahu içinde Hawaii. Hawaii Üniversitesi operates a Coral Reef Assessment and Monitoring Program to help relocate and restore coral reefs in Hawaii. A boat channel from the island of Oahu to the Hawaii Institute of Marine Biology açık Hindistan Cevizi Adası was overcrowded with coral reefs. Many areas of coral reef patches in the channel had been damaged from past dredging in the channel.
Dredging covers corals with sand. Coral larvae cannot settle on sand; they can only build on existing reefs or compatible hard surfaces, such as rock or concrete. Because of this, the University decided to relocate some of the coral. They transplanted them with the help of Amerikan ordusu divers, to a site relatively close to the channel. They observed little if any damage to any of the colonies during transport and no mortality of coral reefs was observed on the transplant site. While attaching the coral to the transplant site, they found that coral placed on hard rock grew well, including on the wires that attached the corals to the site.
No environmental effects were seen from the transplantation process, recreational activities were not decreased, and no scenic areas were affected.
As an alternative to transplanting coral themselves, juvenile fish can also be encouraged to relocate to existing coral reefs by auditory simulation. In damaged sections of the Great Barrier Reef, loudspeakers playing recordings of healthy reef environments, were found to attract fish twice as often as equivalent patches where no sound was played, and also increased species biodiversity by 50%.
Heat-tolerant symbionts
Another possibility for coral restoration is gene therapy: inoculating coral with genetically modified bacteria, or naturally-occurring heat-tolerant varieties of coral symbiotes, may make it possible to grow corals that are more resistant to climate change and other threats.[188] Warming oceans are forcing corals to adapt to unprecedented temperatures. Those that do not have a tolerance for the elevated temperatures experience coral bleaching and eventually mortality. There is already research that looks to create genetically modified corals that can withstand a warming ocean. Madeleine J. H. van Oppen, James K. Oliver, Hollie M. Putnam, and Ruth D. Gates described four different ways that gradually increase in human intervention to genetically modify corals.[189] These methods focus on altering the genetics of the zooxanthellae within coral rather than the alternative.
The first method is to induce acclimatization of the first generation of corals.[189] The idea is that when adult and offspring corals are exposed to stressors, the zooxanthellae will gain a mutation. This method is based mostly on the chance that the zooxanthellae will acquire the specific trait that will allow it to better survive in warmer waters. The second method focuses on identifying what different kinds of zooxanthellae are within the coral and configuring how much of each zooxanthella lives within the coral at a given age.[189] Use of zooxanthellae from the previous method would only boost success rates for this method. However, this method would only be applicable to younger corals, for now, because previous experiments of manipulation zooxanthellae communities at later life stages have all failed. The third method focuses on selective breeding tactics.[189] Once selected, corals would be reared and exposed to simulated stressors in a laboratory. The last method is to genetically modify the zooxanthellae itself.[189] When preferred mutations are acquired, the genetically modified zooxanthellae will be introduced to an aposymbiotic poly and a new coral will be produced. This method is the most laborious of the fourth, but researchers believe this method should be utilized more and holds the most promise in genetic engineering for coral restoration.
Invasive algae
Hawaiian coral reefs smothered by the spread of invasive algae were managed with a two-prong approach: divers manually removed invasive algae, with the support of super-sucker barges. Grazing pressure on invasive algae needed to be increased to prevent the regrowth of the algae. Researchers found that native collector urchins were reasonable candidate grazers for algae biocontrol, to extirpate the remaining invasive algae from the reef.[125]
Invasive algae in Caribbean reefs

Macroalgae, or better known as seaweed, has to potential to cause reef collapse because they can outcompete many coral species. Macroalgae can overgrow on corals, shade, block recruitment, release biochemicals that can hinder spawning, and potentially form bacteria harmful to corals.[190][191] Historically, algae growth was controlled by herbivorous fish and sea urchins. Parrotfish are a prime example of reef caretakers. Consequently, these two species can be considered as keystone species for reef environments because of their role in protecting reefs.
Before the 1980s, Jamaica's reefs were thriving and well cared for, however, this all changed after Hurricane Allen occurred in 1980 and an unknown disease spread across the Caribbean. In the wake of these events, massive damage was caused to both the reefs and sea urchin population across Jamaican's reefs and into the Caribbean Sea. As little as 2% of the original sea urchin population survived the disease.[191] Primary macroalgae succeeded the destroyed reefs and eventually larger, more resilient macroalgae soon took its place as the dominant organism.[191][192] Parrotfish and other herbivorous fish were few in numbers because of decades of overfishing and bycatch at the time.[192] Historically, the Jamaican coast had 90% coral cover and was reduced to 5% in the 1990s.[192] Eventually, corals were able to recover in areas where sea urchin populations were increasing. Sea urchins were able to feed and multiply and clear off substrates, leaving areas for coral polyps to anchor and mature. However, sea urchin populations are still not recovering as fast as researchers predicted, despite being highly fecundate.[191] It is unknown whether or not the mysterious disease is still present and preventing sea urchin populations from rebounding. Regardless, these areas are slowly recovering with the aid of sea urchin grazing. This event supports an early restoration idea of cultivating and releasing sea urchins into reefs to prevent algal overgrowth.[193][194]
Microfragmentation and fusion
In 2014, Christopher Page, Erinn Muller, and David Vaughan from the International Center for Coral Reef Research & Restoration at Mote Marine Laboratory in Summerland Key, Florida developed a new technology called "microfragmentation," in which they use a specialized diamond band saw to cut corals into 1 cm2 fragments instead of 6 cm2 to advance the growth of brain, boulder, and star corals.[195] Mercanlar Orbicella faveolata ve Montastraea cavernosa were outplanted off the Florida's shores in several microfragment arrays. After two years, O. faveolata had grown 6.5x its original size while M. cavernosa had grown nearly twice its size.[195] Under conventional means, both corals would have required decades to reach the same size. It is suspected that if predation events had not occurred near the beginning of the experiment O. faveolata would have grown at least ten times its original size.[195] By using this method, Mote Marine Laboratory produced 25,000 corals and planted 10,000 in the Florida Keys in only one year. Shortly after, they discovered that these microfragments fused with other microfragments from the same parent coral. Typically, corals that are not from the same parent fight and kill nearby corals in an attempt to survive and expand. This new technology is known as "fusion" and has been shown to grow coral heads in just two years instead of the typical 25–75 years. After fusion occurs, the reef will act as a single organism rather than several independent reefs. Currently, there has been no published research into this method.[195]
Tarih

The times of maximum reef development were in the Orta Kambriyen (513–501 Anne ), Devoniyen (416–359 Ma) and Karbonifer (359–299 Ma), owing to order Rugosa nesli tükenmiş corals and Geç Kretase (100–66 Ma) and all Neojen (23 Ma–present), owing to sipariş Scleractinia mercanlar.
Not all reefs in the past were formed by corals: those in the Erken Kambriyen (542–513 Ma) resulted from calcareous yosun ve arkeosikitler (small animals with conical shape, probably related to süngerler ) Ve içinde Geç Kretase (100–66 Ma), when reefs formed by a group of çift kabuklular aranan rudistler existed; one of the valves formed the main conical structure and the other, much smaller valve acted as a cap.
Measurements of the oxygen isotopic composition of the aragonitic skeleton of coral reefs, such as Porit, can indicate changes in sea surface temperature and sea surface salinity conditions during the growth of the coral. This technique is often used by climate scientists to infer a region's paleoiklim.[196]
Ayrıca bakınız
- Catlin Seaview Survey
- Mercan Resifleri Sayımı – A field project of the Census of Marine Life
- Mercan resif organizasyonları
- Deniz Biyolojisi – The scientific study of organisms that live in the ocean
- Sünger resif
- Sözde atol – An island that encircles a lagoon, either partially or completely that is not formed by subsidence or coral reefs
Referanslar
- ^ Lee, Jeong-Hyun; Chen, Jitao; Chough, Sung Kwun (June 1, 2015). "The middle–late Cambrian reef transition and related geological events: A review and new view". Yer Bilimi Yorumları. 145: 66–84. Bibcode:2015ESRv..145...66L. doi:10.1016/j.earscirev.2015.03.002. ISSN 0012-8252.
- ^ Mercan resifleri NOAA National Ocean Service. Accessed: 10 January 2020.
- ^ Spalding MD, Grenfell AM (1997). "New estimates of global and regional coral reef areas". Mercan resifleri. 16 (4): 225–230. doi:10.1007/s003380050078. S2CID 46114284.
- ^ a b c d e Spalding, Mark, Corinna Ravilious, and Edmund Green (2001). World Atlas of Coral Reefs. Berkeley, CA: University of California Press and UNEP/WCMC ISBN 0520232550.
- ^ Mulhall M (Spring 2009) Saving rainforests of the sea: An analysis of international efforts to conserve coral reefs Arşivlendi 6 Ocak 2010, Wayback Makinesi Duke Environmental Law and Policy Forum 19:321–351.
- ^ Where are Corals Found? NOAA. Reviewed: 13 May 2011. Retrieved: 24 March 2015.
- ^ Hoover, John (November 2007). Hawaiʻi's Sea Creatures. Mutual. ISBN 978-1-56647-220-3.
- ^ a b Cesar, H.J.S.; Burke, L.; Pet-Soede, L. (2003). The Economics of Worldwide Coral Reef Degradation. The Netherlands: Cesar Environmental Economics Consulting. s. 4. (pdf: bağlantı ). Alındı 21 Eylül 2013.
- ^ a b Costanza, Robert; Ralph d'Arge; Rudolf de Groot; Stephen Farber; Monica Grasso; Bruce Hannon; Karin Limburg; Shahid Naeem; Robert V. O'Neill; Jose Paruelo; Robert G. Raskin; Paul Sutton; Marjan van den Belt (May 15, 1997). "The value of the world's ecosystem services and natural capital". Doğa. 387 (6630): 253–260. Bibcode:1997Natur.387..253C. doi:10.1038/387253a0. S2CID 672256.
- ^ Costanza, Robert; de Groot, Rudolph; Sutton, Paul (2014). "Changes in the global value of ecosystem services". Küresel Çevresel Değişim. 26 (1): 152–158. doi:10.1016/j.gloenvcha.2014.04.002.
- ^ a b Danovaro, Roberto; Bongiorni, Lucia; Corinaldesi, Cinzia; Giovannelli, Donato; Damiani, Elisabetta; Astolfi, Paola; Greci, Lucedio; Pusceddu, Antonio (Nisan 2008). "Güneş Koruyucuları Viral Enfeksiyonları Teşvik Ederek Mercan Ağartmasına Neden Olur". Çevre Sağlığı Perspektifleri. 116 (4): 441–447. doi:10.1289 / ehp.10966. PMC 2291018. PMID 18414624.
- ^ "Mercanlar arazi kullanımının etkisini ortaya koyuyor". Mercan Resifi Çalışmaları için ARC Mükemmeliyet Merkezi. Alındı 21 Eylül 2013.
- ^ Minato, Charissa (1 Temmuz 2002). "Kentsel yüzey akışı ve kıyı suyu kalitesi mercan resifleri üzerindeki etkiler için araştırılıyor" (PDF). Arşivlenen orijinal (PDF) 10 Haziran 2010.
- ^ "Kıyı Havzası Bilgi Formları - Mercan Resifleri ve Kıyı Havzanız". Çevre Koruma Kurumu Su Ofisi. Temmuz 1998.
- ^ Kleypas Joanie (2010). "Mercan resifi". Dünya Ansiklopedisi. Arşivlenen orijinal 15 Ağustos 2010. Alındı 4 Nisan, 2011.
- ^ Darwin, Charles (1843). "Mercan Resiflerinin Yapısı ve Dağılımı. 1832-1836 yılları arasında Yüzbaşı Fitzroy, R.N. komutasındaki Beagle yolculuğunun jeolojisinin ilk bölümü olan". Londra: Smith Elder and Co. Alıntı dergisi gerektirir
| günlük =(Yardım) - ^ Şansölye Gordon (2008). "Mercan Resiflerine Giriş". Darwin Çevrimiçi. Alındı 20 Ocak 2009. Alıntı dergisi gerektirir
| günlük =(Yardım) - ^ "Mercan Resifleri ve Atollerin 4 Ana Teorisi | Okyanuslar | Coğrafya". Coğrafya Notları. Mart 11, 2017. Alındı 1 Ağustos, 2020.
- ^ Mercan atol oluşumunun animasyonu Arşivlendi 14 Temmuz 2012, Wayback Makinesi NOAA Okyanus Eğitim Hizmeti. Erişim tarihi: January 9, 2010.
- ^ Webster, Jody M .; Braga, Juan Carlos; Clague, David A .; Gallup, Christina; Hein, James R .; Potts, Donald C .; Renema, Willem; Binicilik, Robert; Riker-Coleman, Kristin; Gümüş, Eli; Wallace, Laura M. (1 Mart 2009). "Hızla azalan kenar boşluklarında mercan kayalıklarının evrimi". Küresel ve Gezegensel Değişim. 66 (1–2): 129–148. Bibcode:2009GPC .... 66..129W. doi:10.1016 / j.gloplacha.2008.07.010.CS1 bakimi: birden çok ad: yazarlar listesi (bağlantı)
- ^ Webster, Jody M .; Clague, David A .; Riker-Coleman, Kristin; Gallup, Christina; Braga, Juan C .; Potts, Donald; Moore, James G .; Winterer, Edward L .; Paull, Charles K. (1 Ocak 2004). "Hawaii açıklarında -150 metrelik resifin boğulması: 1A küresel eriyik su darbesi zayiatı mı?". Jeoloji. 32 (3): 249. Bibcode:2004Geo .... 32..249W. doi:10.1130 / G20170.1.
- ^ Great Barrier Reef Marine Park Authority (2006). "Büyük Set Resifi'nin" büyük resim "görüntüsü" (PDF). Tur Rehberleri için Resif Gerçekleri. Arşivlenen orijinal (PDF) 20 Haziran 2007. Alındı 18 Haziran 2007.
- ^ a b Tobin Barry (2003) [1998]. "Büyük Set Resifi nasıl oluştu". Avustralya Deniz Bilimleri Enstitüsü. Arşivlenen orijinal 5 Ekim 2006. Alındı 22 Kasım, 2006.
- ^ CRC Reef Araştırma Merkezi Ltd. "Büyük Set Resifi nedir?". Arşivlenen orijinal 22 Ağustos 2006. Alındı 28 Mayıs 2006.
- ^ Dört Tür Mercan Kayalığı Microdocs, Stanford Education. Erişim tarihi: January 10, 2010.
- ^ MSN Encarta (2006). Büyük Set Resifi. Arşivlenen orijinal 28 Ekim 2009. Alındı 11 Aralık 2006.
- ^ a b Murphy, Richard C. (2002). Mercan Resifleri: Denizlerin Altındaki Şehirler. Darwin Press, Inc. ISBN 978-0-87850-138-0.
- ^ Hopley, David (ed.) Modern Mercan Resifleri Ansiklopedisi Dordrecht: Springer, 2011. s. 40.
- ^ Örneğin. Ünite 10: Resif Türleri Coral Reef Ekoloji Müfredatında. Erişim tarihi: 1 Şubat 2018.
- ^ Whittow, John (1984). Fiziki Coğrafya Sözlüğü. Londra: Penguin, 1984, s. 443. ISBN 0-14-051094-X.
- ^ Thomas David S.G. ve Andrew Goudie (editörler) (2000), Fiziki Coğrafya Sözlüğü, 3. baskı, Oxford, Blackwell, s. 403. ISBN 0-631-20473-3.
- ^ Spalding, Mark, Corinna Ravilious ve Edmund P. Green. Dünya Mercan Resifleri Atlası. Berkeley: California Üniversitesi, 2001, s. 16.
- ^ a b c Ulusal Okyanus ve Atmosfer İdaresi. Mercan Resifi Bilgi Sistemi Sözlüğü, 2014.
- ^ Saçaklı Resifler (Kıyı Resifleri) www.pmfias.com adresinde. 2 Şubat 2018 alındı.
- ^ a b c d e f Mercan Resifi Oluşum Türleri coral.org'da. 2 Şubat 2018 alındı.
- ^ McClanahan, C.R.C. Sheppard ve D.O. Obura. Hint Okyanusu'nun Mercan Resifleri: Ekolojisi ve Korunması. Oxford: OUP, 2000, s. 136.
- ^ Goudie, Andrew. Jeomorfoloji Ansiklopedisi, Londra: Routledge, 2004, s. 411.
- ^ Ghiselin, Michael T. Darwinci Metodun Zaferi. Berkeley, Kaliforniya Üniversitesi, 1969, s. 22.
- ^ Hanauer, Eric. Mısır Kızıldeniz: Bir Dalgıç Rehberi. San Diego: Watersport, 1988, s. 74.
- ^ a b c d e f g Mercan Resifi Türleri Arşivlendi 13 Eylül 2017, at Wayback Makinesi www.coral-reef-info.com adresinde. 2 Şubat 2018 alındı.
- ^ a b c Leser, Hartmut, ed. (2005). Wörterbuch Allgemeine Geographie (Almanca) (13. dtv ed.). Münih, DE. s. 685. ISBN 978-3-423-03422-7.
- ^ Scoffin TP, Dixon JE (1983). "Mercan resiflerinin dağılımı ve yapısı: Darwin'den yüz yıl". Linnean Society Biyolojik Dergisi. 20: 11–38. doi:10.1111 / j.1095-8312.1983.tb01587.x.
- ^ Jell JS, Flood PG (Nisan 1978). "Oğlak burcu ve Bunker gruplarının resiflerinin jeolojisi rehberi, Büyük Set Resifi bölgesi". Makaleler, Jeoloji Bölümü. 8 (3). sayfa 1-85, lütfen. 1-17. Alındı 28 Haziran 2018.
- ^ Hopley, David. Modern Mercan Resifleri Ansiklopedisi: Yapı, Form ve Süreç. Dordrecht: Springer, 2011, s. 51.
- ^ Maldivler Mercan Adası www.mymaldives.com adresinde. 2 Şubat 2018 alındı.
- ^ Sweatman, Hugh; Robertson, D. Ross (1994), "Yavru Karayip cerrah balıklarında otlayan haleler ve avlanma" (PDF), Deniz Ekolojisi İlerleme Serisi, 111 (1–6): 1, Bibcode:1994MEPS..111 .... 1S, doi:10.3354 / meps111001, alındı 24 Nisan 2019
- ^ Smithers, S.G .; Woodroffe, C.D. (2000). "Orta okyanus atolünde deniz seviyesi göstergesi olarak mikroatolllar". Deniz Jeolojisi. 168 (1–4): 61–78. Bibcode:2000MGeol.168 ... 61S. doi:10.1016 / S0025-3227 (00) 00043-8.
- ^ a b c d Moyle, Peter B .; Joseph J. Cech (2004). Balıklar: iktiyolojiye giriş (Beşinci baskı). Upper Saddle Nehri, NJ: Pearson / Prentice Hall. s. 556. ISBN 978-0-13-100847-2.
- ^ Connell, Joseph H. (24 Mart 1978). "Tropikal Yağmur Ormanlarında ve Mercan Resiflerinde Çeşitlilik". Bilim. 199 (4335): 1302–1310. Bibcode:1978Sci ... 199.1302C. doi:10.1126 / science.199.4335.1302. PMID 17840770.
- ^ UNEP (2001) UNEP-WCMC Dünya Mercan Resifleri Atlası Mercan Kayalığı Birimi
- ^ Achituv, Y. ve Dubinsky, Z. 1990. Dünyadaki Mercan Resifleri Ekosistemlerinin Evrimi ve Zoocoğrafyası. Cilt 25: 1–8.
- ^ a b Wells, Sue; Hanna, Nick (1992). Greenpeace Mercan Resifleri Kitabı. Sterling Yayıncılık Şirketi. ISBN 978-0-8069-8795-8.
- ^ Vajed Samiei, J .; Dab K .; Ghezellou P .; Shirvani A. (2013). "Larak Adası, Basra Körfezi'nin Bazı Skleractinian Mercanları (Sınıf: Anthozoa)". Zootaxa. 3636 (1): 101–143. doi:10.11646 / zootaxa.3636.1.5. PMID 26042286.
- ^ Gunnerus, Johan Ernst (1768). Om Nogle Norske Coraller.
- ^ a b Nybakken, James. 1997. Deniz Biyolojisi: Ekolojik Bir Yaklaşım. 4. baskı Menlo Park, CA: Addison Wesley.
- ^ NOAA CoRIS - Bölgesel Portal - Florida. Coris.noaa.gov (16 Ağustos 2012). Erişim tarihi: 3 Mart 2013.
- ^ NGM.nationalgeographic.com, Ultra Marine: Uzak doğu Endonezya'da Raja Ampat adaları olağanüstü bir mercan vahşi doğasını kucaklıyor, David Doubilet, National Geographic, Eylül 2007
- ^ Yaşayan Resifler Vakfı. Erişim tarihi: Mayıs 28, 2015.
- ^ LiveScience. Erişim tarihi: 14 Nisan 2016.
- ^ Sherman, C.D.H. (2006). Genotipik Çeşitlilik ve Yerel Uyum Düzeylerinin Belirlenmesinde İnce Ölçekli Çevresel Heterojenliğin Önemi (PDF) (Doktora tezi). Wollongong Üniversitesi. Arşivlenen orijinal (PDF) 24 Temmuz 2008. Alındı 7 Haziran 2009.
- ^ Zooxanthellae… Bu Nedir?. Oceanservice.noaa.gov (25 Mart 2008). Erişim tarihi: 1 Kasım 2011.
- ^ a b Marshall, Paul; Schuttenberg, Heidi (2006). Mercan Ağartma için Resif Yöneticisinin Kılavuzu. Townsville, Avustralya: Great Barrier Reef Marine Park Authority. ISBN 978-1-876945-40-4.
- ^ Mercanlar hayvanlar mı yoksa bitkiler mi? NOAA: Ulusal Okyanus Hizmeti. Erişim tarihi 11 Şubat 2020. Güncellenme tarihi: 7 Ocak 2020.
- ^ a b Reynolds J, Bruns B, Fitt W, Schmidt G (2008). "Sığ su mercanlarının ve diğer cnidaryanların simbiyotik dinoflagellatlarında gelişmiş ışık koruma yolları". Ulusal Bilimler Akademisi Bildiriler Kitabı. 105 (36): 13674–13678. Bibcode:2008PNAS..10513674R. doi:10.1073 / pnas.0805187105. PMC 2527352. PMID 18757737.
- ^ Stacy J, Marion G, McCulloch M, Hoegh-Guldberg O (Mayıs 2007). "Mackay Whitsunday su kalitesinde ve karasal, mangrov ve mercan resif ekosistemleri arasındaki bağlantıda uzun vadeli değişiklikler: Mercan vekillerinden ve uzaktan algılama kayıtlarından ipuçları" (PDF). Deniz Araştırmaları Merkezi. ARC Bağlantı Hibesinden (2004–2007) araştırmanın sentezi. Queensland Üniversitesi. Arşivlenen orijinal (PDF) 30 Ağustos 2007. Alındı 7 Haziran 2009.
- ^ Nothdurft, L.D. (2007). Son resif binası Scleractinian mercanlarının mikro yapısı ve erken diyajenezleri, Heron Resifi, Büyük Bariyer Resifi: Paleoiklim analizi için çıkarımlar (PDF) (Doktora tezi). Queensland Teknoloji Üniversitesi. Alındı 7 Haziran 2009.[kalıcı ölü bağlantı ]
- ^ Wilson RA (9 Ağustos 2007). "Bireyin Biyolojik Kavramı". Stanford Felsefe Ansiklopedisi. Alındı 7 Haziran 2009.
- ^ Chappell, John (17 Temmuz 1980). "Mercan morfolojisi, çeşitliliği ve resif büyümesi". Doğa. 286 (5770): 249–252. Bibcode:1980Natur.286..249C. doi:10.1038 / 286249a0. S2CID 4347930.
- ^ Jackson, Jeremy B.C. (1 Temmuz 1991). "Resif Mercanlarının Adaptasyonu ve Çeşitliliği". BioScience. 41 (7): 475–482. doi:10.2307/1311805. JSTOR 1311805.
- ^ Vermeij, Mark J. A .; Marhaver, Kristen L .; Huijbers, Chantal M .; Nagelkerken, Ivan; Simpson, Stephen D. (2010). "Mercan larvaları resif seslerine doğru hareket ediyor". PLOS ONE. 5 (5): e10660. doi:10.1371 / journal.pone.0010660. PMC 2871043. PMID 20498831.
- ^ Gleason, D. F .; Danilowicz, B. S .; Nolan, C.J. (2009). "Resif suları, kuluçka dönemindeki Karayip mercanlarından gelen düzlemlerde substrat keşfini teşvik eder". Mercan resifleri. 28 (2): 549–554. doi:10.1007 / s00338-009-0480-1. S2CID 39726375.
- ^ Jennings S, Kaiser MJ, Reynolds JD (2001). Deniz Balıkçılığı Ekolojisi. Wiley-Blackwell. s. 291–293. ISBN 978-0-632-05098-7.
- ^ "Floresan mercan". fotoğrafçılık. Mercan krallıkları. National Geographic Topluluğu.
- ^ Rougerier, F (1998). "Mercan resiflerinin ve atollerin işleyişi: paradokstan paradigmaya". Jost, Christian (ed.). Fransızca Konuşan Pasifik: Nüfus, Çevre ve Kalkınma Sorunları. Boombana Yayınları. ISBN 978-1-876542-02-3. (pdf: bağlantı ).
- ^ Darwin, Charles R. (1842). Mercan Resiflerinin Yapısı ve Dağılımı. Kaptan Fitzroy komutasındaki Beagle yolculuğunun jeolojisinin ilk bölümü olan R.N. 1832 ile 1836 yılları arasında. Londra: Smith Elder and Co. s.61–71.
- ^ Crossland CJ (1983). "Mercan resif sularında çözünmüş besinler". Barnes, D. J. (ed.). Mercan Resifleri Üzerine Perspektifler. Avustralya Deniz Bilimleri Enstitüsü. s. 56–68. ISBN 9780642895851.
- ^ Odum, E.P. (1971). Ekolojinin Temelleri. 3.ed. Saunders.
- ^ Sammarco, PW; Risk, MJ; Schwarcz, HP; Heikoop, JM (1999). "Great Barrier Reef üzerindeki mercan δ15N'de kıtalar arası sahanlık eğilimleri: resif besin paradoksunun daha fazla değerlendirilmesi" (PDF). Mar Ecol Prog Ser. 180: 131–138. Bibcode:1999MEPS..180..131S. doi:10.3354 / meps180131.
- ^ Rougerie, F; Wauthy, B (1993). "Endo-upwelling konsepti: jeotermal konveksiyondan resif inşasına" (PDF). Mercan resifleri. 12 (1): 19–30. Bibcode:1993 CorRe..12 ... 19R. doi:10.1007 / bf00303781. S2CID 27590358.
- ^ De Goeij, Jasper M (2009) "Tropikal mercan resiflerinde element döngüsü: kriptik karbon şantı ortaya çıktı" Doktora tezi, sayfa 13. Groningen Üniversitesi.
- ^ Sorokin, Yuri I. (1993). Mercan Kayalığı Ekolojisi. Almanya: Springer-Verlag, Berlin Heidelberg. ISBN 978-0-387-56427-2.
- ^ Hatcher, Bruce Gordon (1 Mayıs 1988). "Mercan kayalığı birincil üretkenliği: Bir dilencinin ziyafeti". Ekoloji ve Evrimdeki Eğilimler. 3 (5): 106–111. doi:10.1016/0169-5347(88)90117-6. PMID 21227159.
- ^ Ross, On; Sharples, J (11 Ekim 2007). "Fitoplankton hareketliliği ve termoklinde besinler için rekabet". Deniz Ekolojisi İlerleme Serisi. 347: 21–38. Bibcode:2007MEPS..347 ... 21R. doi:10.3354 / meps06999. ISSN 0171-8630.
- ^ a b c Castro, Peter ve Huber, Michael (2000) Deniz Biyolojisi. 3. baskı Boston: McGraw-Hill.
- ^ Roach, John (7 Kasım 2001). "Besleyici-Zayıf Sudaki Zengin Mercan Resifleri: Paradoks Açıklandı mı?". National Geographic Haberleri. Alındı 5 Nisan, 2011.
- ^ Nowak, Rachel (21 Eylül 2002). "Mercanlar, Darwin paradoksu üzerinde sert oynuyor". Yeni Bilim Adamı (2361).
- ^ a b c Leichter, J .; Wing S .; Miller S .; Denny M. (1996). "İç gelgit sondajlarından Conch Reef'e (Florida Keys) alt termoklin suyunun darbeli teslimi". Limnoloji ve Oşinografi. 41 (7): 1490–1501. Bibcode:1996LimOc..41.1490L. doi:10.4319 / lo.1996.41.7.1490.
- ^ Wolanski, E .; Pickard, G.L. (1983). "Büyük Set Resifi'ndeki kıta sahanlığındaki iç gelgitler ve kelvin dalgaları ile yükselme". Deniz ve Tatlı Su Araştırmaları. 34: 65. doi:10.1071 / MF9830065.
- ^ Leichter, J .; Helmuth B .; Fischer A. (2006). "Yüzeyin altındaki varyasyon: Karayipler, Bahamalar ve Florida'daki mercan resiflerindeki karmaşık termal ortamların nicelendirilmesi". Deniz Araştırmaları Dergisi. 64 (4): 563–588. doi:10.1357/002224006778715711.
- ^ Ezer, T .; Heyman W .; Houser C .; Kjerfve B. (2011). "Karayip resiflerinin yumurtlama kümelenme bölgesinde yüksek frekanslı akış değişkenliği ve iç dalgaların modellenmesi ve gözlemleri". Okyanus Dinamikleri. 61 (5): 581–598. Bibcode:2011OcDyn..61..581E. doi:10.1007 / s10236-010-0367-2. S2CID 55252988.
- ^ Fratantoni, D .; Richardson P. (2006). "Kuzey Brezilya Güncel Halkalarının Evrimi ve Ölümü" (PDF). Fiziksel Oşinografi Dergisi. 36 (7): 1241–1249. Bibcode:2006JPO .... 36.1241F. doi:10.1175 / JPO2907.1. hdl:1912/4221.
- ^ a b c d Leichter, J .; Shellenbarger G .; Genovese S .; Wing S. (1998). "Florida (ABD) mercan resifinde iç dalgaları kırmak: işte bir plankton pompası mı?". Deniz Ekolojisi İlerleme Serisi. 166: 83–97. Bibcode:1998MEPS..166 ... 83L. doi:10.3354 / meps166083.
- ^ Talley, L. (2011). Tanımlayıcı Fiziksel Oşinografi: Giriş. Oxford Birleşik Krallık: Elsevier Inc. ISBN 978-0750645522.
- ^ Helfrich, K. (1992). "Tek tip bir eğimde dahili tek dalga kırılması ve yükselmesi". Akışkanlar Mekaniği Dergisi. 243: 133–154. Bibcode:1992JFM ... 243..133H. doi:10.1017 / S0022112092002660.
- ^ Gregg, M. (1989). "Termoklinde ölçeklenen türbülans yayılımı". Jeofizik Araştırmalar Dergisi. 9686–9698. 94 (C7): 9686. Bibcode:1989JGR .... 94.9686G. doi:10.1029 / JC094iC07p09686.
- ^ Taylor, J. (1992). "Rezonant olarak zorlanan bir iç dalga alanında olayları kırmanın enerjisi". Akışkanlar Mekaniği Dergisi. 239: 309–340. Bibcode:1992JFM ... 239..309T. doi:10.1017 / S0022112092004427.
- ^ Andrews, J .; Gentien P. (1982). "Büyük Set Resifi ekosistemleri için bir besin kaynağı olarak yukarı çıkma: Darwin'in sorusuna bir çözüm mü?". Deniz Ekolojisi İlerleme Serisi. 8: 257–269. Bibcode:1982MEPS .... 8..257A. doi:10.3354 / meps008257.
- ^ Sandstrom, H .; Elliott J. (1984). "İskoç sahanlığındaki iç gelgitler ve solitonlar: İş başında bir besin pompası". Jeofizik Araştırmalar Dergisi. 89 (C4): 6415–6426. Bibcode:1984JGR .... 89.6415S. doi:10.1029 / JC089iC04p06415.
- ^ a b Wolanski, E .; Hamner W. (1988). "Okyanustaki topografik olarak kontrol edilen cepheler ve biyolojik önemi". Bilim. 241 (4862): 177–181. Bibcode:1988Sci ... 241..177W. doi:10.1126 / science.241.4862.177. PMID 17841048. S2CID 19757639.
- ^ Rougerie, F .; Fagerstrom J .; Andrie C. (1992). "Jeotermal endo-yükselme: Resif besin paradoksuna bir çözüm mü?" (PDF). Kıta Sahanlığı Araştırması. 12 (7–8): 785–798. Bibcode:1992CSR .... 12..785R. doi:10.1016 / 0278-4343 (92) 90044-K.
- ^ Wolanski, E .; Delesalle B. (1993). "İç dalgaların yükselişi, Tahiti, Fransız Polinezyası". Kıta Sahanlığı Araştırması. 15 (2–3): 357–368. Bibcode:1995CSR .... 15..357W. doi:10.1016 / 0278-4343 (93) E0004-R.
- ^ Szmant, A. M .; Forrester, A. (1996). "Florida Keys, ABD'deki su sütunu ve tortu nitrojen ve fosfor dağılım modelleri". Mercan resifleri. 15 (1): 21–41. Bibcode:1996CorRe..15 ... 21S. doi:10.1007 / BF01626075. S2CID 42822848.
- ^ Furnas, M. J .; Mitchell, A.W. (1996). "Mercan Denizi sularının yüzey altı izinsiz girişlerinden merkezi Büyük Set Resifi'ne (Avustralya) besin girdileri: İki boyutlu bir yer değiştirme modeli". Kıta Sahanlığı Araştırması. 16 (9): 1127–1148. Bibcode:1996CSR .... 16.1127F. doi:10.1016/0278-4343(95)00060-7.
- ^ Leichter, J .; Miller S. (1999). "Yüksek frekanslı yükselmeyi tahmin etmek: Florida mercan resifindeki sıcaklık anormalliklerinin mekansal ve zamansal modelleri". Kıta Sahanlığı Araştırması. 19 (7): 911–928. Bibcode:1999CSR .... 19..911L. doi:10.1016 / s0278-4343 (99) 00004-7.
- ^ a b c Leichter, J .; Stewart H .; Miller S. (2003). "Florida mercan resiflerine dönemsel besin nakliyesi". Limnoloji ve Oşinografi. 48 (4): 1394–1407. Bibcode:2003LimOc..48.1394L. doi:10.4319 / lo.2003.48.4.1394. S2CID 15125174.
- ^ Leichter, J .; Deane G .; Stokes M. (2005). "Mercan Resifi Üzerindeki İç Dalga Zorlamasının Uzamsal ve Zamansal Değişkenliği" (PDF). Fiziksel Oşinografi Dergisi. 35 (11): 1945–1962. Bibcode:2005JPO .... 35.1945L. doi:10.1175 / JPO2808.1.
- ^ Smith, J .; Smith C .; Vroom P .; Sahil K .; Miller S. (2004). "Florida Keys, Conch Reef'te Halimeda orkinosunun besin ve büyüme dinamikleri: İç gelgitlerin besin durumu ve fizyolojisi üzerindeki olası etkisi". Limnoloji ve Oşinografi. 49 (6): 1923–1936. Bibcode:2004LimOc..49.1923S. doi:10.4319 / lo.2004.49.6.1923.
- ^ a b Pineda, J. (1994). "Yakın kıyıdaki iç gelgit sondajları: Sıcak su cepheleri, denizdeki yerçekimi akımları ve neustonik larvaların karada taşınması". Deniz Araştırmaları Dergisi. 52 (3): 427–458. doi:10.1357/0022240943077046.
- ^ Wilson, E (2004). "Coral's Symbiotic Bacteria Fluoresce, Fix Nitrogen". Kimya ve Mühendislik Haberleri. 82 (33): 7. doi:10.1021 / cen-v082n033.p007a.
- ^ Barnes, R.S.K .; Mann, K.H. (1991). Sucul Ekolojinin Temelleri. Blackwell Publishing. s. 217–227. ISBN 978-0-632-02983-9.
- ^ Fuchs. T (2013). "Mercan Resifi Karmaşıklığının Omurgasız Biyoçeşitliliği Üzerindeki Etkileri". Immediate Science Ecology Publishing: 1–10. Arşivlenen orijinal 2 Nisan 2015.
- ^ Hatcher, B.G. Johannes, R.E .; Robertson, A.J. (1989). "Sığ Su Deniz Ekosistemlerinin Korunması". Oşinografi ve Deniz Biyolojisi: Yıllık İnceleme. 27. Routledge. s. 320. ISBN 978-0-08-037718-6.CS1 bakimi: birden çok ad: yazarlar listesi (bağlantı)
- ^ "Dünya'nın Resif Balıkları İnsan Aşırı Nüfusuyla Boğuşuyor". Günlük Bilim. 5 Nisan 2011.
- ^ Gratwicke, B .; Speight, M.R. (2005). "Bir dizi sığ tropikal deniz habitatında balık türlerinin zenginliği, bolluğu ve habitat karmaşıklığı arasındaki ilişki". Balık Biyolojisi Dergisi. 66 (3): 650–667. doi:10.1111 / j.0022-1112.2005.00629.x. ISSN 0022-1112.
- ^ Fontaneto, Diego; Sanciangco, Jonnell C .; Carpenter, Kent E .; Etnoyer, Peter J .; Moretzsohn, Fabio (2013). "Tropikal Hint-Pasifik'teki Deniz Türlerinin Zenginliğinin Belirleyicileri Olarak Habitat Bulunabilirliği ve Heterojenlik ve Hint-Pasifik Sıcak Havuzu". PLOS ONE. 8 (2): e56245. Bibcode:2013PLoSO ... 856245S. doi:10.1371 / journal.pone.0056245. ISSN 1932-6203. PMC 3574161. PMID 23457533.
- ^ "Mercan Resifi Biyolojisi". NOAA. Arşivlenen orijinal 27 Eylül 2011. Alındı 6 Nisan 2011.
- ^ Glynn, P.W. (1990). Dubinsky, Z. (ed.). Ecosystems of the World / 25-Mercan Resifleri. New York, NY: Elsevier Science. ISBN 978-0-444-87392-7.
- ^ Murphy, James W.A .; Richmond, Robert H. (19 Nisan 2016). "Oksijen yoksunluğu altında mercan sağlığı ve metabolik aktivitede değişiklikler". PeerJ. 4: e1956. doi:10.7717 / peerj.1956. ISSN 2167-8359. PMC 4841221. PMID 27114888.
- ^ "SEDİMENTLERİN, BESİNLERİN VE DİĞER KİRLETİCİLERİN KARASAL AKIŞININ MERCAN REEFLERİNE ETKİSİ" (PDF). Arşivlenen orijinal (PDF) Mart 4, 2016. Alındı 5 Aralık 2015.
- ^ Vroom, Peter S .; Page, Kimberly N .; Kenyon, Jean C .; Brainard, Russell E. (2006). "Yosun Etkili Resifler". Amerikalı bilim adamı. 94 (5): 430–437. doi:10.1511/2006.61.1004.
- ^ Kaplan Matt (2009). "Sünger nasıl ince kalır". Doğa. doi:10.1038 / haber.2009.1088.
- ^ Buchheim, Jason. "Mercan Resifi Balık Ekolojisi". marinebiology.org. Alındı 5 Nisan, 2011.
- ^ McClellan, Kate; Bruno, John (2008). "Yıkıcı balıkçılık uygulamaları yoluyla mercanların bozulması". Dünya Ansiklopedisi. Alındı 25 Ekim 2008.
- ^ Osborne, Patrick L. (2000). Tropikal Ekosistem ve Ekolojik Kavramlar. Cambridge: Cambridge University Press. s. 464. ISBN 978-0-521-64523-2.
- ^ a b Westbrook, Charley E .; Ringang, Rory R .; Cantero, Sean Michael A .; Toonen, Robert J .; Team, HDAR & TNC Urchin (15 Eylül 2015). "Dışarıda bırakılmış deniz kestaneleri, Tripneustes gratilla'nın boyut sınıfları arasında hayatta kalma ve beslenme tercihleri ve istilacı yabancı algler için biyolojik kontrol olarak olası kullanım". PeerJ. 3: e1235. doi:10.7717 / peerj.1235. ISSN 2167-8359. PMC 4579015. PMID 26401450.
- ^ Conklin, Eric J .; Smith, Jennifer E. (1 Kasım 2005). "Kane'ohe Körfezi, Hawai'i'de İstilacı Kırmızı Alglerin Bolluğu ve Yayılması, Kappaphycus spp. Ve Yönetim Seçeneklerinin Deneysel Değerlendirmesi". Biyolojik İstilalar. 7 (6): 1029–1039. doi:10.1007 / s10530-004-3125-x. ISSN 1387-3547. S2CID 33874352.
- ^ Midway'in albatros nüfusu sabit. The.honoluluadvertiser.com (17 Ocak 2005). Erişim tarihi: 1 Kasım 2011.
- ^ "ABD Balık ve Vahşi Yaşam Servisi - Birds of Midway Atoll". Arşivlenen orijinal 22 Mayıs 2013. Alındı 19 Ağustos 2009.
- ^ Heatwole Harold (1999). Deniz yılanları (2. baskı). Malabar, Fla: Krieger. ISBN 978-1-57524-116-6.
- ^ Li, Min; Fry, B.G .; Kini, R. Manjunatha (1 Ocak 2005). "Sadece Yumurta Diyeti: Mermer Deniz Yılanının (Aipysurus eydouxii) Toksin Profil Değişiklikleri ve Ekolojisi için Etkileri". Moleküler Evrim Dergisi. 60 (1): 81–89. Bibcode:2005JMolE..60 ... 81L. doi:10.1007 / s00239-004-0138-0. PMID 15696370. S2CID 17572816.
- ^ Voris Harold K. (1 Ocak 1966). "Deniz Yılanı Cinsi, Emydocephalus (Krefft) için Görünür Tek Gıda Öğesi Olarak Balık Yumurtaları". Ekoloji. 47 (1): 152–154. doi:10.2307/1935755. JSTOR 1935755.
- ^ McClenachan, Loren; Jackson, Jeremy BC; Newman, Marah JH (1 Ağustos 2006). "Tarihi deniz kaplumbağasının yuva yapan sahil kaybının koruma etkileri". Ekoloji ve Çevrede Sınırlar. 4 (6): 290–296. doi:10.1890 / 1540-9295 (2006) 4 [290: ciohst] 2.0.co; 2.
- ^ Lutz, Peter L .; Musick, John A. (1996). Deniz kaplumbağalarının biyolojisi. Boca Raton, Fla: CRC Press. ISBN 978-0849384226.
- ^ Meylan, Anne (22 Ocak 1988). "Hawksbill Kaplumbağalarında Süngerimsi: Camdan Bir Diyet". Bilim. 239 (4838): 393–395. Bibcode:1988Sci ... 239..393M. doi:10.1126 / science.239.4838.393. PMID 17836872. S2CID 22971831.
- ^ a b "Mercanın İnsanlar İçin Önemi". Dünya Vahşi Yaşam Fonu. Alındı 7 Nisan 2011.
- ^ "Kıyı Başkenti: Karayipler'deki Kıyı Ekosistemlerinin Ekonomik Değerlendirmesi". Dünya Kaynakları Enstitüsü.
- ^ Cooper, Emily; Burke, Lauretta; Bood, Nadia (2008). "Kıyı Başkenti: Belize: Belize Mercan Resiflerinin ve Mangrovlarının Ekonomik Katkısı" (PDF). Alındı 6 Nisan 2011.
- ^ Sarkis, Samia; van Beukering, Pieter J.H .; McKenzie Emily (2010). "Bermuda Mercan Resiflerinin Toplam Ekonomik Değeri. Ekosistem Hizmetlerinin Değerlemesi" (PDF). Alındı 29 Mayıs 2015.
- ^ Ferarrio, F .; et al. (2014). "Mercan resiflerinin kıyı tehlike riskinin azaltılması ve adaptasyonu için etkinliği". Doğa İletişimi. 5:3794: 3794. Bibcode:2014NatCo ... 5.3794F. doi:10.1038 / ncomms4794. PMC 4354160. PMID 24825660.
- ^ Beck, M .; et al. (2018). "Mercan resifleri tarafından sağlanan küresel sel koruma tasarrufu". Doğa İletişimi. 9:2186 (1): 2186. Bibcode:2018NatCo ... 9.2186B. doi:10.1038 / s41467-018-04568-z. PMC 5997709. PMID 29895942.
- ^ "Dünyadaki mercan resifleri". Guardian.co.uk. 2 Eylül 2009.
- ^ Peters, Esther C. (2015). "Mercan Resifi Organizmalarının Hastalıkları". Antroposen'de Mercan Resifleri. Springer Hollanda: 147–178. doi:10.1007/978-94-017-7249-5_8. ISBN 978-94-017-7248-8.
- ^ Bradbury, R. H .; Hammond, L. S .; Moran, P. J .; Reichelt, R. E. (7 Mart 1985). "Mercan resif toplulukları ve dikenli denizyıldızı: Niteliksel olarak kararlı döngüler için kanıt". Teorik Biyoloji Dergisi. 113 (1): 69–80. doi:10.1016 / S0022-5193 (85) 80076-X. ISSN 0022-5193.
- ^ Hutchings, P.A. (1986). "Mercan resiflerinin biyolojik olarak yok edilmesi". Mercan resifleri. 12 (1): 1–17. Bibcode:1986 CorRe ... 4..239H. doi:10.1007 / BF00298083. S2CID 34046524.
- ^ Clark, Malcolm R .; Tittensor, Derek P. (2010). "Deniz dağlarında dip trolü yapan mercanların taşlanma riskini değerlendirmek için bir indeks". Deniz Ekolojisi. 31 (s1): 200–211. Bibcode:2010MarEc..31..200C. doi:10.1111 / j.1439-0485.2010.00392.x. ISSN 1439-0485.
- ^ Caras, Tamir; Pasternak, Zohar (1 Ekim 2009). "Endonezya, Wakatobi deniz parkında mercan madenciliğinin uzun vadeli çevresel etkisi". Okyanus ve Kıyı Yönetimi. 52 (10): 539–544. doi:10.1016 / j.ocecoaman.2009.08.006. ISSN 0964-5691.
- ^ "Şok balıkçılığı". Yasadışı Balıkçılığı Durdurun. Alındı 15 Kasım 2019.
- ^ "Magnuson-Stevens Yasası: Sürdürülebilir deniz ürünleri için benzersiz bir ücret | Ulusal Okyanus ve Atmosfer İdaresi". www.noaa.gov. Alındı 15 Kasım 2019.
- ^ "Mercan". ABD Balık ve Vahşi Yaşam Servisi. Alındı 15 Kasım 2019.
- ^ Stierwalt, Everyday Einstein Sabrina. "Hawaii Güneş Koruyucu Neden Yasaklıyor?". Bilimsel amerikalı. Alındı 19 Ağustos 2018.
- ^ Kuzu, Joleah; Bette Willis (16 Ağustos 2011). "Tropikal bir deniz parkındaki açık deniz resifleri üzerindeki turizm faaliyetlerini yoğunlaştırmanın etkilerini değerlendirmek için mercan hastalığı yaygınlığını kullanmak". Koruma Biyolojisi. 25 (5): 1044–1052. doi:10.1111 / j.1523-1739.2011.01724.x. PMID 21848962.
- ^ "Karayip mercan resifleri 20 yıl içinde yok olabilir: Rapor". Ian. news.biharprabha.com. Alındı 3 Temmuz, 2014.
- ^ McCulloch, Malcolm T .; D'Olivo, Juan Pablo; Falter, James; Holcomb, Michael; Trotter, Julie A. (30 Mayıs 2017). "Değişen bir Dünyada mercan kalsifikasyonu ve pH ve DIC yukarı regülasyonunun etkileşimli dinamikleri". Doğa İletişimi. 8 (1): 15686. Bibcode:2017NatCo ... 815686M. doi:10.1038 / ncomms15686. ISSN 2041-1723. PMC 5499203. PMID 28555644.
- ^ Kwiatkowski, Lester; Cox, Peter M .; Economou, Theo; Halloran, Paul R .; Mumby, Peter J .; Booth, Ben B. B .; Carilli, Jessica; Guzman, Hector M. (Mayıs 2013). "Antropojenik aerosol emisyonlarından etkilenen Karayip mercanlarının büyümesi". Doğa Jeolojisi. 6 (5): 362–366. Bibcode:2013NatGe ... 6..362K. doi:10.1038 / ngeo1780. ISSN 1752-0908.
- ^ Clapham ME ve Payne (2011). "Asidifikasyon, anoksi ve yok olma: Orta ve Geç Permiyen sırasında yok olma seçiciliğinin çoklu lojistik regresyon analizi". Jeoloji. 39 (11): 1059–1062. Bibcode:2011Geo .... 39.1059C. doi:10.1130 / G32230.1.
- ^ Payne JL, Clapham ME (2012). "Okyanuslarda Permiyen Sonu Kitlesel Yokoluş: Yirmi Birinci Yüzyıl İçin Eski Bir Analog mu?". Yeryüzü ve Gezegen Bilimleri Yıllık İncelemesi. 40 (1): 89–111. Bibcode:2012AREPS..40 ... 89P. doi:10.1146 / annurev-earth-042711-105329.
- ^ Denizdeki Yaşam Kaderini Yok Olmanın Paroksizminde Buldu New York Times, 30 Nisan 2012.
- ^ Abrego, D .; Ulstrup, K. E; Willis, B. L; van Oppen, M. J.H (7 Ekim 2008). "Alg endosimbiyomları ve mercan konakçılar arasındaki türe özgü etkileşimler, ısıya ve ışık stresine karşı ağartma tepkilerini tanımlar". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 275 (1648): 2273–2282. doi:10.1098 / rspb.2008.0180. ISSN 0962-8452. PMC 2603234. PMID 18577506.
- ^ a b Konuk, James R .; Baird, Andrew H .; Maynard, Jeffrey A .; Muttaqin, Efin; Edwards, Alasdair J .; Campbell, Stuart J .; Yewdall, Katie; Affendi, Yang Amri; Chou, Loke Ming (9 Mart 2012). "2010'da Mercan Ağartma Duyarlılığının Zıt Modelleri Termal Strese Uyarlanabilir Bir Tepki Öneriyor". PLOS ONE. 7 (3): e33353. Bibcode:2012PLoSO ... 733353G. doi:10.1371 / journal.pone.0033353. ISSN 1932-6203. PMC 3302856. PMID 22428027.
- ^ a b Barkley, Hannah C .; Cohen, Anne L .; Mollica, Nathaniel R .; Brainard, Russell E .; Rivera, Hanny E .; DeCarlo, Thomas M .; Lohmann, George P .; Drenkard, Elizabeth J .; Alpert, Alice E. (8 Kasım 2018). "Geçtiğimiz altmış yılda (1960-2016) bir Orta Pasifik mercan resifinin ağartılmasını tekrarlayın". İletişim Biyolojisi. 1 (1): 177. doi:10.1038 / s42003-018-0183-7. ISSN 2399-3642. PMC 6224388. PMID 30417118.
- ^ Ritter, Karl (8 Aralık 2010). −goal-coral-reefs.html "İklim hedefi bazı mercan resifleri için sona erebilir". İlişkili basın.[kalıcı ölü bağlantı ]
- ^ Markey, Sean (16 Mayıs 2006). "Küresel Isınmanın Mercan Resifleri Üzerinde Yıkıcı Etkisi Var, Çalışma Gösterileri". National Geographic Haberleri.
- ^ Maynard, J. A .; Anthony, K.R.N .; Marshall, P. A .; Masiri, I. (1 Ağustos 2008). "Büyük ağartma olayları, mercanlarda termal toleransın artmasına neden olabilir". Deniz Biyolojisi. 155 (2): 173–182. doi:10.1007 / s00227-008-1015-y. ISSN 1432-1793. S2CID 85935124.
- ^ a b Thompson, D. M .; van Woesik, R. (22 Ağustos 2009). "Mercanlar, son zamanlarda ve tarihsel olarak sık termal stres yaşayan bölgelerde ağarmadan kaçıyor". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 276 (1669): 2893–2901. doi:10.1098 / rspb.2009.0591. PMC 2817205. PMID 19474044.
- ^ Matz, Mikhail V .; Treml, Eric A .; Aglyamova, Galina V .; Bay, Line K. (19 Nisan 2018). "Bir Great Barrier Reef mercanında ısınmaya hızlı genetik adaptasyon potansiyeli ve sınırları". PLOS Genetiği. 14 (4): e1007220. doi:10.1371 / journal.pgen.1007220. ISSN 1553-7404. PMC 5908067. PMID 29672529.
- ^ McClanahan, Timothy; Marnane, Michael; Cinner, Joshua E .; Kiene, William E. (2006). "Deniz Koruma Alanları ve Mercan-Resif Yönetimine Alternatif Yaklaşımların Karşılaştırması". Güncel Biyoloji. 16 (14): 1408–13. doi:10.1016 / j.cub.2006.05.062. PMID 16860739. S2CID 17105410.
- ^ Christie, P. (2004). "Güneydoğu Asya'da biyolojik başarılar ve sosyal başarısızlıklar olarak deniz koruma alanları". Amerikan Balıkçılık Derneği Sempozyumu. 2004 (42): 155-164. Arşivlenen orijinal 16 Aralık 2013.
- ^ McClanahan, Timothy; Davies, Jamie; Maina Joseph (2005). "Kaynak kullanıcılarının ve yöneticilerin Kenya'da deniz koruma alanı yönetimine yönelik algılarını etkileyen faktörler". Çevresel koruma. 32: 42–49. doi:10.1017 / S0376892904001791.
- ^ Stone, Gregory (Ocak 2011). "Anka kuşu yükseliyor". National Geographic Dergisi.
- ^ Ewa Magiera; Sylvie Rockel (2 Temmuz 2014). "Umutsuzluktan onarıma: Karayip mercanlarının dramatik düşüşü tersine çevrilebilir". Alındı 8 Haziran 2015.
- ^ "Karayip Mercan Resifleri - Durum Raporu 1970-2012" (PDF). IUCN.org. 2013. Arşivlenen orijinal (PDF) 11 Ocak 2015.
- ^ a b Walsworth, T.E .; Schindler, D.E .; Colton, M.A .; Webster, M.S .; Palumbi, S.R .; Mumby, P.J .; Essington, T.E .; Pinsky, M.L. (1 Temmuz 2019). "Ağ çeşitliliğinin yönetimi, iklim değişikliğine evrimsel uyumu hızlandırıyor". Doğa Araştırması. 9: 632–636.CS1 bakimi: birden çok ad: yazarlar listesi (bağlantı)
- ^ "Dünya Mirası Listesi". UNESCO. Alındı 18 Aralık 2016.
- ^ "Büyük Set Resifi için bir biyolojik çeşitlilik stratejisi". Great Barrier Reef Marine Park Authority, Avustralya Hükümeti. Alındı 20 Eylül 2013.
- ^ "Büyük Bariyer Resifi İklim Değişikliği Eylem Planı 2007–2011" (PDF). Great Barrier Reef Marine Park Authority. 2007.
- ^ Cinner, Joshua E .; Marnane, Michael J .; McClanahan, Tim R. (2005). "Papua Yeni Gine'deki Ahus Adası'ndaki geleneksel mercan resif yönetiminden koruma ve topluluk faydaları". Koruma Biyolojisi. 19 (6): 1714–1723. doi:10.1111 / j.1523-1739.2005.00209.x-i1.
- ^ "Mercan Kayalıkları Yönetimi, Papua Yeni Gine". Nasa 's Dünya Gözlemevi. Alındı 2 Kasım, 2006.
- ^ Kelly, R.P; Foley; Fisher, WS; Feely, RA; Halpern, BS; Waldbusser, GG; Caldwell, MR; et al. (2011). "Okyanus asitlenmesinin yerel nedenlerini mevcut yasalarla hafifletmek" (PDF). Bilim. 332 (6033): 1036–1037. Bibcode:2011Sci ... 332.1036K. doi:10.1126 / science.1203815. PMID 21617060. S2CID 206533178.
- ^ Mallikarjun, Y. (10 Aralık 2014). "Mercan resiflerinin sağlığını değerlendirmek için uydular". Hindu. Alındı 13 Aralık, 2014.
- ^ "Mercan Restorasyonu - Köpekbalığı Araştırma ve Koruma Programı (SRC) | Miami Üniversitesi". Alındı 3 Mayıs, 2020.
- ^ Horoszowski-Fridman YB, Izhaki I, Rinkevich B (2011). "Fidanlıklarda yetiştirilen gravid kolonilerin transplantasyonu yoluyla mercan resifi larvası tedarikinin mühendisliği". Deneysel Deniz Biyolojisi ve Ekoloji Dergisi. 399 (2): 162–166. doi:10.1016 / j.jembe.2011.01.005.
- ^ Pomeroy RS, Parks JE, Balboa CM (2006). "Resifin yetiştirilmesi: Su ürünleri yetiştiriciliği, mercan resifleri üzerindeki balıkçılık baskısını azaltmak için bir çözüm müdür?". Denizcilik Politikası. 30 (2): 111–130. doi:10.1016 / j.marpol.2004.09.001.
- ^ Rinkevich, B (2008). "Mercan resiflerinin yönetimi: Aktif resif restorasyonunu ihmal ederken yanlış yaptık" (PDF). Deniz Kirliliği Bülteni. 56 (11): 1821–1824. doi:10.1016 / j.marpolbul.2008.08.014. PMID 18829052. Arşivlenen orijinal (PDF) 23 Mayıs 2013.
- ^ Ferse, S.C.A. (2010). "Kısa Dayanıklı Alt Tabakalara Nakledilen Mercanların Kötü Performansı". Restorasyon Ekolojisi. 18 (4): 399–407. doi:10.1111 / j.1526-100X.2010.00682.x.
- ^ a b c Lirman, Diego; Schopmeyer, Stephanie (20 Ekim 2016). "Resif bozulmasına ekolojik çözümler: Karayipler ve Batı Atlantik'te mercan resif restorasyonunu optimize etme". PeerJ. 4: e2597. doi:10.7717 / peerj.2597. ISSN 2167-8359. PMC 5075686. PMID 27781176.
- ^ Rinkevich, Baruch (1995). "Eğlence Faaliyetlerinden Zarar Gören Mercan Resifleri İçin Restorasyon Stratejileri: Cinsel ve Cinsel Olmayan Acemilerin Kullanımı". Restorasyon Ekolojisi. 3 (4): 241–251. doi:10.1111 / j.1526-100X.1995.tb00091.x. ISSN 1526-100X.
- ^ a b c Yanovski, Roy; Abelson, Avigdor (1 Temmuz 2019). "Potansiyel bir mercan resifi restorasyon aracı olarak yapısal karmaşıklığın iyileştirilmesi". Ekolojik Mühendislik. 132: 87–93. doi:10.1016 / j.ecoleng.2019.04.007. ISSN 0925-8574.
- ^ "Gen Terapisi Mercanların İklim Değişikliğinden Kurtulmasına Yardımcı Olabilir". Bilimsel amerikalı. 29 Şubat 2012.
- ^ a b c d e Oppen, Madeleine J. H. van; Oliver, James K .; Putnam, Hollie M .; Gates, Ruth D. (24 Şubat 2015). "Yardımlı evrim yoluyla mercan kayalığı direnci oluşturma". Ulusal Bilimler Akademisi Bildiriler Kitabı. 112 (8): 2307–2313. Bibcode:2015PNAS..112.2307V. doi:10.1073 / pnas.1422301112. ISSN 0027-8424. PMC 4345611. PMID 25646461.
- ^ Vieira, Christophe; Payri, Claude; Clerck, Olivier (8 Eylül 2016). "Makroalgal-mercan etkileşimlerine yeni bir bakış: makroalg mercanlar için bir tehdit mi?". Psikolojide Perspektifler. 3 (3): 129–140. doi:10.1127 / pip / 2016/0068.
- ^ a b c d Knowlton, N. (24 Nisan 2001). "Deniz kestanesinin toplu ölümden kurtulması: Karayip mercan resifleri için yeni bir umut mu?". Ulusal Bilimler Akademisi Bildiriler Kitabı. 98 (9): 4822–4824. Bibcode:2001PNAS ... 98.4822K. doi:10.1073 / pnas.091107198. ISSN 0027-8424. PMC 33118. PMID 11320228.
- ^ a b c Edmunds, P. J .; Carpenter, R.C. (27 Mart 2001). "Diadema antillarum geri kazanımı, makroalgal örtüyü azaltır ve bir Karayip resifindeki yavru mercanların bolluğunu artırır". Ulusal Bilimler Akademisi Bildiriler Kitabı. 98 (9): 5067–5071. doi:10.1073 / pnas.071524598. ISSN 0027-8424. PMC 33164. PMID 11274358.
- ^ McClanahan, T.R .; Kaunda-Arara, B. (Ağustos 1996). "Mercan Resifli Deniz Parkında Balıkçılık Geri Kazanımı ve Bitişik Balıkçılık Üzerindeki Etkisi". Koruma Biyolojisi. 10 (4): 1187–1199. doi:10.1046 / j.1523-1739.1996.10041187.x. ISSN 0888-8892.
- ^ Sammarco, Paul W. (1 Ocak 1980). "Diadema ve mercan tükürük ölüm oranıyla ilişkisi: Otlatma, rekabet ve biyolojik rahatsızlık". Deneysel Deniz Biyolojisi ve Ekoloji Dergisi. 45 (2): 245–272. doi:10.1016/0022-0981(80)90061-1. ISSN 0022-0981.
- ^ a b c d Sayfa, Christopher A .; Muller, Erinn M .; Vaughan, David E. (1 Kasım 2018). "Yavaş büyüyen masif mercanların başarılı bir şekilde restorasyonu için mikro parçalama". Ekolojik Mühendislik. 123: 86–94. doi:10.1016 / j.ecoleng.2018.08.017. ISSN 0925-8574.
- ^ Cobb, K .; Charles, Christopher D .; Cheng, Hai; Edwards, R. Lawrence (2003). "El Nino / Güney Salınımı ve son milenyum boyunca tropikal Pasifik iklimi" (PDF). Doğa. 424 (6946): 271–6. Bibcode:2003Natur.424..271C. doi:10.1038 / nature01779. PMID 12867972. S2CID 6088699. Arşivlenen orijinal (PDF) 11 Ocak 2012.
Diğer referanslar
- Mercan Resiflerinin Korunması: Mercan Resifleri Nedir?. ABD EPA.
- UNEP. 2004. Güney Çin Denizi'ndeki Mercan Resifleri. UNEP / GEF / SCS Teknik Yayını No. 2.
- UNEP. 2007. Güney Çin Denizi'ndeki Mercan Resifleri Gösteri Alanları. UNEP / GEF / SCS Teknik Yayını No. 5.
- UNEP, 2007. Güney Çin Denizi'nin Kıyı Sularındaki Mercan Resifleri Üzerine Ulusal Raporlar. UNEP / GEF / SCS Teknik Yayını No. 11.
Dış bağlantılar
| Harici Görsel | |
|---|---|
- "Mercan Resifi Bilgi Formu". Waitt Enstitüsü. Arşivlenen orijinal 9 Haziran 2015. Alındı 8 Haziran 2015.
- Mercanlar ve Mercan Resifleri Smithsonian Okyanus Portalı'na genel bakış
- Mercanlar hakkında Avustralya Deniz Bilimleri Enstitüsü.
- Uluslararası Mercan Resifi Girişimi
- Moorea Mercan Resifi Uzun Dönemli Ekolojik Araştırma Sitesi (ABD NSF)
- Mercan Resifi Çalışmaları için ARC Mükemmeliyet Merkezi
- NOAA'nın Mercan Kayalığı Bilgi ve Haberleri için Mercan Listesi Liste Sunucusu
- NOAA'nın Mercan Resifi Koruma Programı
- NOAA'nın Mercan Resifi Bilgi Sistemi
- ReefBase: Mercan Resifleri Üzerine Küresel Bir Bilgi Sistemi
- Ulusal Mercan Resifi Enstitüsü Nova Güneydoğu Üniversitesi
- Deniz Akvaryumu Konseyi
- NCORE Ulusal Mercan Resifi Araştırma Merkezi Miami Üniversitesi
- Güney Çin Denizi ve Tayland Körfezi'ndeki Mercan Resiflerinin Bilimi ve Yönetimi
- Mikro Dokümanlar: 4 çeşit Resif & Resif yapısı
- Resif Kabartması Mercan resifleri eğitimi ve korumasına odaklanan, aktif Florida çevreci, kar amacı gütmeyen kuruluş
- Küresel Resif Kaydı - Catlin Seaview Resif Araştırması, bir görüntü veri tabanı ve diğer bilgiler
- "Mercanlar ve Mercan Resifleri "(arşivlendi). Nancy Knowlton, iBioSeminars, 2011.
- Nancy Knowlton Semineri: "Mercanlar ve Mercan Resifleri ". Nancy Knowlton, iBioSeminars, 2011.
- Mercan resifleri hakkında Yaşayan Resifler Vakfı, Bermuda
- Karayip Mercan Resifleri - Durum Raporu 1970-2012 tarafından IUCN. - Video açık Youtube, raporu öne çıkarıyor.



| Yer şekilleri |
| 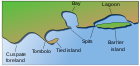  |
|---|---|---|
| Sahiller | ||
| Süreçler |
| |
| Yönetim | ||
| İlişkili | ||
| ||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| Yetki kontrolü |
|---|