Deniz birincil üretimi - Marine primary production

Deniz birincil üretimi okyanustaki kimyasal sentezdir organik bileşikler atmosferik veya çözünmüş karbon dioksit. Esas olarak şu süreçte gerçekleşir: fotosentez ışığı enerji kaynağı olarak kullanan, ancak aynı zamanda kemosentez, enerji kaynağı olarak inorganik kimyasal bileşiklerin oksidasyonunu veya indirgenmesini kullanan. Neredeyse hepsi hayat Dünyada doğrudan veya dolaylı olarak birincil üretim. Birincil üretimden sorumlu organizmalara birincil üreticiler veya ototroflar.
Deniz birincil üretiminin çoğu, çeşitli deniz mikroorganizmaları aranan yosun ve siyanobakteriler. Bunlar birlikte, temel birincil üreticileri oluştururlar. okyanus besin zinciri ve dünyadaki oksijenin yarısını üretir. Deniz birincil üreticileri, deniz hayvanlarının var olmak için ihtiyaç duydukları neredeyse tüm oksijeni ve besinleri üreterek neredeyse tüm deniz hayvanı yaşamının temelini oluşturur. Bazı birincil denizcilik üreticileri de ekosistem mühendisleri çevreyi değiştiren ve sağlayan habitatlar diğer deniz yaşamı için.
Okyanustaki birincil üretim, karadaki birincil üretimle karşılaştırılabilir. Küresel olarak okyanus ve toprağın her biri yaklaşık aynı miktarda birincil üretim üretir, ancak okyanusta birincil üretim esas olarak siyanobakteriler ve alglerden gelirken, karada ise esas olarak damarlı Bitkiler.
Deniz yosunları büyük ölçüde görünmez olanları içerir ve çoğu zaman tek hücreli mikroalg ile birlikte siyanobakteriler okyanusu oluşturmak fitoplankton ve daha büyük, daha görünür ve karmaşık çok hücreli makroalg Yaygın olarak adlandırılan Deniz yosunu. Deniz yosunları kıyı bölgelerinde bulunur ve kıta rafları ve yıkanmış gelgit bölgeleri. Bazı deniz yosunları, güneşli yüzey sularında planktonla sürüklenir (epipelajik bölge ) açık okyanus.
Geri Silüriyen bazı fitoplanktonlar, kırmızı, Kahverengi ve yeşil alg. Bu algler daha sonra toprağı istila ettiler ve kara bitkileri bugün biliyoruz. Daha sonra Kretase bu kara bitkilerinden bazıları denize geri döndü mangrovlar ve Deniz çayırları. Bunlar kıyılarda bulunur gelgit bölgeleri ve acı sularında haliçler. Ek olarak, deniz yosunları gibi bazı deniz otları kıta sahanlığının hem yumuşak hem de sert tabanlarında 50 metreye kadar derinliklerde bulunabilir.
| Bir dizi genel bakışın parçası |
| Deniz yaşamı |
|---|
Deniz birincil üreticileri
| Bir dizinin parçası |
| Karbon döngüsü |
|---|
 |

• Kırmızı = diyatomlar (silikaya ihtiyaç duyan büyük fitoplankton)
• Sarı = kamçılılar (diğer büyük fitoplankton)
• Yeşil = proklorokok (nitrat kullanamayan küçük fitoplankton)
• Camgöbeği = Synechococcus (diğer küçük fitoplankton)
Opaklık, karbon biyokütlesinin konsantrasyonunu gösterir. Özellikle girdapların ve ipliklerin rolü (orta ölçekli özellikler) okyanustaki yüksek biyolojik çeşitliliğin korunmasında önemli görünmektedir.[2][3]
Birincil üreticiler bunlar ototrof diğer organizmaları yemek yerine kendi yiyeceklerini yapan organizmalar. Bu, birincil üreticilerin başlangıç noktası haline geldiği anlamına gelir. besin zinciri için heterotrof diğer organizmaları yiyen organizmalar. Bazı birincil denizcilik üreticileri, özelleşmiş bakteri ve arkelerdir. kemotroflar etrafında toplanarak kendi yiyeceklerini yapıyorlar hidrotermal menfezler ve soğuk sızıntılar ve kullanarak kemosentez. Ancak çoğu denizci birincil üretim kullanan organizmalardan gelir fotosentez suda çözünen karbondioksit üzerinde. Bu işlem, suyu dönüştürmek için güneş ışığından enerji kullanır ve karbon dioksit[4]:186–187 hem kimyasal enerji kaynağı olarak hem de hücrelerin yapısal bileşenlerinde kullanılan organik moleküllerin şekeri haline getirilir.[4]:1242 Deniz birincil üreticileri önemlidir, çünkü deniz canlılarının çoğunu üreterek hemen hemen tüm deniz hayvanı yaşamını desteklerler. oksijen ve diğer organizmalara var olmak için ihtiyaç duydukları kimyasal enerjiyi sağlayan yiyecekler.
Başlıca denizcilik birincil üreticileri siyanobakteriler, yosun ve deniz bitkileri. oksijen fotosentezin bir yan ürünü olarak piyasaya sürülen neredeyse tüm canlılar gerçekleştirilecek hücresel solunum. Ek olarak, birincil üreticiler küresel ölçekte etkilidir. karbon ve Su döngüleri. Kıyı bölgelerini stabilize ederler ve deniz hayvanları için habitat sağlayabilir. Dönem bölünme geleneksel olarak yerine kullanılmıştır filum birincil üreticileri tartışırken, Algler, mantarlar ve bitkiler için Uluslararası Adlandırma Kodu şimdi şartları eşdeğer olarak kabul ediyor.[5]
Karada, okyanuslarda, hemen hemen tüm fotosentez, algler ve siyanobakteriler tarafından gerçekleştirilir; damarlı Bitkiler ve diğer gruplar. Algler, tek yüzen hücrelerden tutunmuş hücrelere kadar çok çeşitli organizmaları kapsar. yosunlar. Çeşitli gruplardan fotoototrofları içerirler. Öbakteriler hem okyanus hem de karasal ekosistemlerde önemli fotosentezleyicilerdir ve bazıları Archaea vardır fototrofik hiçbirinin oksijenle gelişen fotosentezi kullandığı bilinmemektedir.[6] Bir dizi ökaryotlar okyanustaki birincil üretime önemli katkılarda bulunur; yeşil alg, kahverengi algler ve kırmızı yosun ve çeşitli tek hücreli gruplar grubu. Vasküler bitkiler de okyanusta şu gruplarla temsil edilir: Deniz çayırları.
Karasal ekosistemlerin aksine, okyanustaki birincil üretimin çoğunluğu serbest yaşam tarafından gerçekleştirilir. mikroskobik organizmalar aranan fitoplankton. Dünyadaki oksijenin yarısının fitoplankton tarafından üretildiği tahmin edilmektedir.[7][8] Deniz otları ve makroalgler gibi daha büyük ototroflar (yosunlar ) genel olarak sınırlıdır kıyı bölge ve bitişik sığ sular eklemek temeldeki alt tabakaya, ancak yine de fotik bölge. Gibi istisnalar vardır Sargassum ancak serbest yüzen üretimin büyük çoğunluğu mikroskobik organizmalar içinde gerçekleşir.
Okyanustaki birincil üretimi sınırlayan faktörler de karadakilerden çok farklı. Açıktır ki, suyun mevcudiyeti bir sorun değildir (yine de tuzluluk olabilir). Benzer şekilde, sıcaklık etkilerken metabolik oranları (bakınız Q10 ), okyanusta karada olduğundan daha az yaygındır çünkü ısı kapasitesi deniz suyu tamponlarının sıcaklık değişimleri ve oluşumu Deniz buzu yalıtır daha düşük sıcaklıklarda. Bununla birlikte, ışığın mevcudiyeti, fotosentez için enerji kaynağı ve mineral besinler, yeni büyümenin yapı taşları, okyanustaki birincil üretimin düzenlenmesinde çok önemli roller oynamaktadır.[9] Mevcut Dünya Sistem Modelleri, okyanuslarda devam eden biyo-jeokimyasal değişikliklerin, emisyon senaryosuna bağlı olarak okyanus NPP'sinde mevcut değerlerin% 3 ila% 10'u arasında azalmaları tetikleyebileceğini öne sürmektedir.[10]
2020'de araştırmacılar, son yirmi yılda birincil üretimin Kuzey Buz Denizi daha yüksek konsantrasyonlar nedeniyle yaklaşık% 60'lık bir artış göstermektedir. fitoplankton. Diğer okyanuslardan yeni besinlerin aktığını varsayıyorlar ve bunun Arktik okyanusunun destekleyebileceği anlamına geldiğini öne sürüyorlar. daha yüksek trofik seviye üretimi ve ek karbon fiksasyonu gelecekte.[11][12]
Siyanobakteriler
Siyanobakteriler, tek hücreliden tek hücrelere kadar değişen bir bakteri filumudur (bölümü). ipliksi ve dahil sömürge türleri, hangi inorganik karbonu düzeltmek organik karbon bileşiklerine dönüştürülür. Dünyanın hemen hemen her yerinde bulunurlar: nemli toprakta, hem tatlı su hem de deniz ortamlarında ve hatta Antarktika kayalarında.[19] Özellikle, bazı türler okyanusta yüzen sürüklenen hücreler olarak meydana gelir ve bu nedenle, fitoplankton. Bu bakteriler, okyanusun içinde olmadığında atmosferdeki nitrojeni işleyebildikleri için algler gibi işlev görürler.

Fotosentez kullanan ilk birincil üreticiler okyanus siyanobakteriler yaklaşık 2,3 milyar yıl önce.[20][21] Moleküler salınım oksijen tarafından siyanobakteriler fotosentezin bir yan ürünü olarak Dünya'nın çevresinde küresel değişikliklere neden oldu. Oksijen o sırada Dünya'daki çoğu yaşam için zehirli olduğu için, bu durum neredeyse yok olmaya yol açtı. oksijene tahammülsüz organizmalar, bir dramatik değişim ana hayvan ve bitki türlerinin evrimini yeniden yönlendirdi.[22]
Küçük deniz siyanobakteri Proklorokok 1986'da keşfedilen, bugün okyanus tabanının bir parçasını oluşturuyor besin zinciri ve açık okyanusun fotosentezinin yarısından fazlasını oluşturur[23] ve Dünya atmosferindeki oksijenin tahmini% 20'si.[24] Muhtemelen dünyadaki en bol cinstir: tek bir mililitre yüzey deniz suyu, 100.000 veya daha fazla hücre içerebilir.[25]
Biyologlar başlangıçta siyanobakteriler yosundu ve "mavi-yeşil alg" olarak anılıyordu. Daha yeni görüş, siyanobakterilerin bir bakteri olduğu ve dolayısıyla aynı şey olmadığıdır. Krallık yosun olarak. Çoğu otorite hepsini dışlar prokaryotlar ve dolayısıyla alg tanımından siyanobakteriler.[26][27]
Biyolojik pigmentler
Biyolojik pigmentler bitki veya hayvan hücrelerindeki herhangi bir renkli materyaldir. Tüm biyolojik pigmentler seçici olarak emmek belirli dalga boylarında ışık yansıtan diğerleri.[28][29] Bitkilerdeki pigmentlerin temel işlevi fotosentez yeşil pigmenti kullanan klorofil ve mümkün olduğunca fazla ışık enerjisini emen birkaç renkli pigment. Klorofil bitkilerdeki birincil pigmenttir; bu bir klor ışığı yansıtırken sarı ve mavi dalga boylarını emen yeşil. Bitkilere yeşil rengini veren klorofilin varlığı ve görece bolluğudur. Yeşil alg ve bitkiler bu pigmentin iki formuna sahiptir: klorofil a ve klorofil b. Kelps, diyatomlar ve diğer fotosentetik heterokontlar klorofil içerir c onun yerine bkırmızı algler sadece klorofile sahipken a. Tüm klorofiller, bitkilerin fotosentezi beslemek için ışığı kesmek için kullandıkları birincil araç görevi görür.
Kloroplastlar


Kloroplastlar (Yunanca'dan Chloros yeşil için ve plastes "oluşturan" için[31]) organeller bu davranış fotosentez fotosentetik pigmentin klorofil yakalar enerji itibaren Güneş ışığı, dönüştürür ve serbest bırakırken enerji depolama moleküllerinde depolar oksijen sudan bitki ve alg hücreler. Daha sonra depolanan enerjiyi organik moleküller yapmak için kullanırlar. karbon dioksit olarak bilinen bir süreçte Calvin döngüsü.
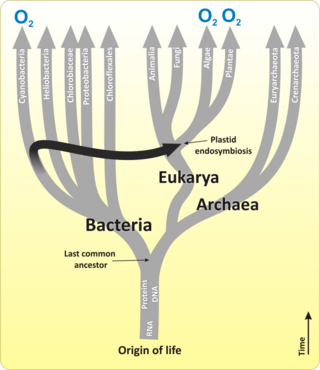
Bir kloroplast, bir tür organeldir. plastid, ile karakterize edilen onun iki zarı ve yüksek konsantrasyon klorofil. Oldukça dinamiktirler - bitki hücreleri içinde dolaşırlar ve hareket ederler ve bazen ikiye sıkıştır yeniden üretmek. Davranışları, açık renk ve yoğunluk gibi çevresel faktörlerden büyük ölçüde etkilenir. Kloroplastlar gibi mitokondri, kendi DNA atalarından miras kaldığı düşünülen fotosentetik siyanobakteri bu ... idi yutulmuş erkenden ökaryotik hücre.[32] Kloroplastlar bitki hücresi tarafından yapılamaz ve hücre bölünmesi sırasında her bir yavru hücre tarafından miras alınmalıdır.
Çoğu kloroplast muhtemelen tek bir endosimbiyotik olay, bir siyanobakteri ökaryot tarafından yutulduğunda. Buna rağmen, kloroplastlar son derece geniş bir organizma kümesinde bulunabilir, bazıları doğrudan birbiriyle ilişkili bile değildir. ikincil ve hatta üçüncül endosimbiyotik olaylar.

Mikrobiyal rodopsin

(2) konfigürasyonunu değiştirir, böylece bir proton hücreden atılır
(3) kimyasal potansiyel, protonun hücreye geri akmasına neden olur
(4) böylece enerji üretir
(5) şeklinde adenozin trifosfat.[34]
Fototrofik metabolizma, üç enerji dönüştürücü pigmentten birine dayanır: klorofil, bakterioklorofil, ve retina. Retinal kromofor içinde bulunan Rodopinler. Klorofilin ışık enerjisini dönüştürmedeki önemi onlarca yıldır yazılmıştır, ancak retina pigmentlerine dayalı fototrofi henüz araştırılmaya başlanmıştır.[35]

2000 yılında, liderliğindeki bir mikrobiyolog ekibi Edward DeLong deniz karbon ve enerji döngülerinin anlaşılmasında çok önemli bir keşif yaptı. Birkaç bakteri türünde bir gen keşfettiler[37][38] protein üretiminden sorumlu Rodopsin, daha önce bakterilerde duyulmamış. Hücre zarlarında bulunan bu proteinler, güneş ışığı çarptığında rodopsin molekülünün konfigürasyonundaki bir değişiklik nedeniyle ışık enerjisini biyokimyasal enerjiye dönüştürebilmektedir. proton içten dışa ve ardından enerjiyi üreten bir giriş.[39] Arkeal benzeri rodopinler daha sonra farklı taksonlarda, protistlerde ve ayrıca bakteri ve arkelerde bulunmuştur, ancak kompleks olarak nadirdirler. Çok hücreli organizmalar.[40][41][42]
2019'da yapılan araştırmalar, bu "güneşten kaçan bakterilerin" daha önce düşünülenden daha yaygın olduğunu ve okyanusların küresel ısınmadan nasıl etkilendiğini değiştirebileceğini gösteriyor. "Bulgular, ders kitaplarında bulunan ve okyanustaki neredeyse tüm güneş ışığının alglerdeki klorofil tarafından yakalandığını belirten geleneksel deniz ekolojisi yorumundan kopuyor. çoğu bakteri - ve besinler kıt olduğunda güneş ışığı ile. "[43][35]
Bir astrobiyolojik varsayım denilen Mor Dünya hipotezi Bu, Dünya'daki orijinal yaşam formlarının klorofil bazlı değil retina bazlı olduğunu ve Dünya'nın yeşil yerine mor görünmesini sağlayacağını düşünüyor.[44][45]
Deniz yosunları
| Bir serinin parçası |
| Plankton |
|---|
 |
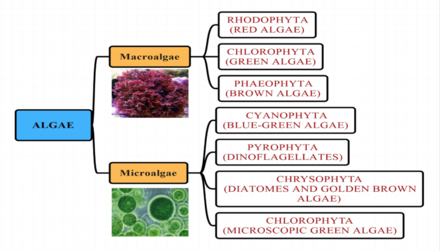
Yosun yaygın ve çeşitli bir fotosentetik koleksiyonu için resmi olmayan bir terimdir ökaryotik mutlaka yakından ilişkili olmayan ve bu nedenle polifirik. Daha yüksek bitkilerin aksine, yosunların kökleri, gövdeleri veya yaprakları yoktur.
Alg grupları
Deniz yosunları geleneksel olarak aşağıdaki gibi gruplar halinde yerleştirilmiştir: yeşil alg, kırmızı yosun, kahverengi algler, diyatomlar, kokolitoforlar ve Dinoflagellatlar.
Yeşil alg
Yeşil alg hayatlarının çoğunu tek hücreler halinde veya filamentli yaşarken diğerleri oluşur koloniler uzun hücre zincirlerinden oluşur veya oldukça farklılaşmış makroskopik deniz yosunlarıdır. Yaklaşık 8.000 tanınmış türü içeren gayri resmi bir grup oluştururlar.[47]
Kırmızı algler
Modern kırmızı yosun çoğunlukla çok hücreli farklılaşmış hücrelerle ve birçok dikkate değer yosunlar.[48][49] Gibi mercan yosunu mercan resiflerinin ekolojisinde önemli bir rol oynarlar. Yaklaşık 7.000 tanınmış tür içeren (tartışmalı) bir filum oluştururlar.[48]

Cyanidiophyceae koloni, tek hücreli kırmızı algler sınıfı

Yosun Porphyra umbilicalis

Kahverengi algler
Kahverengi algler çoğunlukla çok hücreli ve dahil olmak üzere birçok deniz yosunu içerir yosun. Oluştururlar sınıf yaklaşık 2.000 tanınmış tür içeren.[50]
Diyatomlar
Toplamda yaklaşık yüzde 45 birincil üretim okyanuslarda katkıda bulunan diyatomlar.[51]

Diyatomlar en yaygın fitoplankton türlerinden biridir

Dünya oksijen üretiminin yaklaşık% 20'sini oluşturan önemli bir alg grubudur.[52]




Kokolitoforlar

Her yerde bulunan Emiliania huxleyi

Emiliania huxleyi güney İngiltere'de çiçek açar

Kokolitoforlar neredeyse tamamen denizeldir ve tüm dünyada çok sayıda bulunur. güneş ışığı bölgesi of okyanus. Onlarda var kalsiyum karbonat denilen belirsiz işlevli plakalar (veya ölçekler) kokolitler önemli olan mikrofosiller. Coccolithophores, küresel araştırma yapanların ilgisini çekiyor iklim değişikliği Çünkü çünkü okyanus asitliği arttıkça, kokolitleri daha da önemli hale gelebilir. karbon yutağı.[55] Kokolitoforun en bol bulunan türleri, Emiliania huxleyi plankton tabanının her yerde bulunan bir bileşenidir. deniz besin ağları.[56] Ötrofikasyonla ilişkili kokolitofor çiçeklenmelerini önlemek için yönetim stratejileri uygulanmaktadır, çünkü bu çiçekler, okyanusun daha düşük seviyelerine besin akışında bir azalmaya yol açmaktadır.[57]
Dinoflagellat

Dinoflagellatlar

Karenia brevis insanlar için oldukça zehirli kırmızı gelgitler üretir[58]



Mixotrophic algler
Diğer gruplar

Diplonemidler dünya okyanuslarında bol olabilir

Geleneksel olarak soyoluş yukarıda tartışılan alg grupları gibi mikroorganizmaların taksonomi çalışmalarına dayanarak kuruldu morfoloji. Ancak gelişmeler moleküler filogenetik türlerin evrimsel ilişkilerinin analiz edilerek kurulmasına izin vermiştir. DNA ve protein diziler.[59] Yukarıda tartışılan alg grupları da dahil olmak üzere birçok takson, moleküler filogenetik kullanılarak yeniden sınıflandırılma veya yeniden tanımlanma sürecindedir. Son gelişmeler moleküler sıralama iyileşmesine izin verdi genomlar doğrudan çevresel numunelerden ve kültürleme ihtiyacından kaçınarak. Bu, örneğin, zenginlik ve çeşitlilik bilgisinde hızlı bir genişlemeye yol açmıştır. deniz mikroorganizmaları. Genom çözümlemeli gibi moleküler teknikler metagenomik ve tek hücre genomiği ile birlikte kullanılıyor yüksek verim teknikleri.
2009-2013 yılları arasında Tara Okyanuslar seferi planktonları toplayarak ve bunları çağdaş moleküler tekniklerle analiz ederek dünya okyanuslarını dolaştı. Daha önce bilinmeyen çok çeşitli fotosentetik ve miksotrofik algler buldular.[60] Bulguları arasında şunlar vardı: diplonemidler. Bu organizmalar genellikle renksiz ve dikdörtgen şeklindedir, tipik olarak yaklaşık 20 um uzunluğunda ve iki kamçıya sahiptir.[61] Kanıt DNA barkodlama diplonemidlerin, tüm deniz ökaryot gruplarının en bol ve tür açısından en zengini olabileceğini öne sürmektedir.[62][63]
Boyuta göre
Algler boyutlarına göre sınıflandırılabilir: mikroalg veya makroalg.
Mikroalg
Mikroalg çıplak gözle görülemeyen mikroskobik alg türleridir. Çoğunlukla tek hücreli Bireyler veya zincirler veya gruplar halinde var olan türler, ancak bazıları çok hücreli. Mikroalgler, deniz protestocuları yanı sıra deniz fitoplanktonu. Onlar çok çeşitli. Yaklaşık 50.000 türü tanımlanmış olan 200.000-800.000 türün olduğu tahmin edilmektedir.[64] Türlere bağlı olarak boyutları birkaç mikrometre (µm) ile birkaç yüz mikrometre arasında değişir. Viskoz kuvvetlerin hakim olduğu bir ortama özel olarak uyarlanmıştır.
- Mikroalg

Zooxanthellae ev sahiplerinin içinde yaşayan fotosentetik bir yosundur. mercan

Tek hücreli kirpik yeşil ile zoochlorellae içeride yaşamak endosimbiyotik olarak

Euglena mutabilis fotosentetik kamçılı



Makroalg

Makroalg daha büyük çok hücreli ve genellikle adı verilen daha görünür alg türleri yosunlar. Deniz yosunları genellikle sığ kıyı sularında büyür ve burada deniz tabanına bir Dayan. Suda sürüklenen deniz yosunu kumsallarda yıkanabilir. Kelp büyük bir su altında oluşan büyük bir kahverengi deniz yosunu ormanlar dünya kıyı şeridinin yaklaşık% 25'ini kaplar.[65] Dünyadaki en üretken ve dinamik ekosistemler arasındadırlar.[66] Biraz Sargassum yosunlar planktoniktir (serbest yüzer) ve yüzen sürüklenmeler oluşturur.[67](pp246–255) Mikroalgler gibi, makroalgler (deniz yosunları) da teknik olarak deniz protestocuları gerçek bitkiler olmadıkları için.

- Makroalg

Dev yosun gerçek bir bitki olmadığı için teknik olarak bir protisttir, ancak çok hücreli ve 50 m'ye kadar büyüyebilir

Sargassum deniz yosunu, yüzmesine yardımcı olan hava keseleri olan kahverengi bir yosundur

Sargassum balığı sürüklenmek için kamufle edilmiş Sargassum Deniz yosunu

Bu tek hücreli kabarcık yosunu yaşıyor gelgit bölgeleri. 4 cm çapında olabilir.[68]




Deniz bitkileri

Geri Silüriyen bazı fitoplanktonlar, kırmızı, Kahverengi ve yeşil alg. Bu algler daha sonra toprağı istila ettiler ve kara bitkileri bugün biliyoruz. Daha sonra Kretase bu kara bitkilerinin bir kısmı denize geri döndü mangrovlar ve Deniz çayırları.[69]
Bitki yaşamı dünyanın acı sularında gelişebilir. haliçler, nerede mangrovlar veya Cordgrass veya plaj çimi büyüyebilir. Çiçekli bitkiler şeklinde kumlu sığ alanlarda büyür deniz çayırları,[70] mangrovlar tropikal ve subtropikal bölgelerde sahili hizalayın[71] ve tuza toleranslı bitkiler düzenli olarak su altında kalır tuz bataklıkları.[72] Tüm bu habitatlar, büyük miktarlarda karbon tutabilir ve biyolojik çeşitlilik daha büyük ve daha küçük hayvan yaşamı aralığı.[73] Deniz bitkileri bulunabilir gelgit bölgeleri ve sığ sular, örneğin Deniz çayırları sevmek Eelgrass ve kaplumbağa otu, Talasya. Bu bitkiler okyanus ortamının yüksek tuzluluğuna uyum sağlamıştır.
Işık yalnızca en üstteki 200 metreyi (660 ft) geçebilir, bu nedenle denizin bitkilerin büyüyebileceği tek kısmı burasıdır.[74] Yüzey katmanları genellikle biyolojik olarak aktif nitrojen bileşiklerinde yetersizdir. Deniz nitrojen döngüsü karmaşık mikrobiyal dönüşümlerden oluşur. nitrojen fiksasyonu, asimilasyon, nitrifikasyon, anammox ve denitrifikasyon.[75] Bu işlemlerin bazıları derin suda gerçekleşir, böylece soğuk suların yükseldiği yerlerde ve ayrıca kara kaynaklı besinlerin bulunduğu haliçlerin yakınında bitki büyümesi daha yüksektir.Bu, plankton açısından ve dolayısıyla balık bakımından da zengin olan en verimli alanların çoğunlukla kıyı bölgeleri olduğu anlamına gelir.[76](pp160–163)
Mangrovlar
Mangrovlar, büyük balıkların ve omurgasızların larva ve genç formları için saklanma ve yiyecek arama yerleri olarak hareket ederek, deniz yaşamı için önemli fidanlık habitatları sağlar. Uydu verilerine dayanarak, mangrov ormanlarının toplam dünya alanı 2010 yılında 134.257 kilometre kare (51.837 sq mi) olarak tahmin edilmiştir.[77][78]

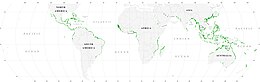
- Spalding, M. (2010) Mangrovların dünya atlası, Routledge. ISBN 9781849776608. doi:10.4324/9781849776608.
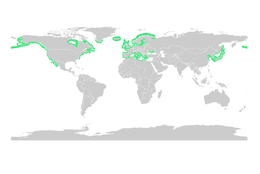
Deniz çayırı
Mangrovlar gibi, deniz otları da büyük balıkların ve omurgasızların larva ve genç formları için önemli fidanlık habitatları sağlar. Deniz çayırlarının toplam dünya alanını belirlemek mangrov ormanlarından daha zordur, ancak 2003 yılında ihtiyatlı bir şekilde 177.000 kilometre kare (68.000 mil kare) olarak tahmin edilmiştir.[79]

Deniz ejderhaları Yüzen deniz yosunu gibi görünmek için kamufle edilmiş, yosun ormanlarında ve deniz çayırı çayırlarında yaşar[80]

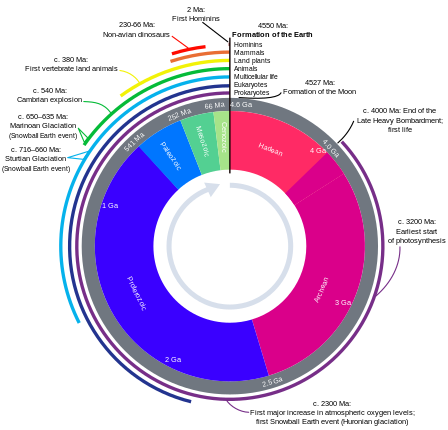
Evrimsel zaman çizelgesi
Ayrıca bakınız
- Yosun
- Su bitkileri
- Bitkilerin evrimsel tarihi
- Bitki evrimi
- Bitki evriminin zaman çizelgesi
- Fotosentezin evrimi
Referanslar
- ^ Klorofil NASA Dünya Gözlemevi. 30 Kasım 2019'da erişildi.
- ^ Küresel Okyanusta Modellenmiş Fitoplankton Toplulukları NASA Hyperwall, 30 Eylül 2015.
 Bu makale, bu kaynaktan alınan metni içermektedir. kamu malı.
Bu makale, bu kaynaktan alınan metni içermektedir. kamu malı. - ^ Darwin Projesi Massachusetts Teknoloji Enstitüsü.
- ^ a b Campbell, Neil A .; Reece, Jane B .; Urry, Lisa Andrea; Cain, Michael L .; Wasserman, Steven Alexander; Minorsky, Peter V .; Jackson, Robert Bradley (2008). Biyoloji (8 ed.). San Francisco: Pearson - Benjamin Cummings. ISBN 978-0-321-54325-7.
- ^ McNeill, J .; ve diğerleri, eds. (2012). Algler, mantarlar ve bitkiler için Uluslararası İsimlendirme Kodu (Melbourne Kodu), Eighteenth International Botanical Congress Melbourne, Avustralya, Temmuz 2011 tarafından kabul edilmiştir. (elektronik ed.). Uluslararası Bitki Taksonomisi Derneği. Alındı 2017-05-14.
- ^ Schäfer G, Engelhard M, Müller V (1 Eylül 1999). "Arkenin Biyoenerjetiği". Microbiol. Mol. Biol. Rev. 63 (3): 570–620. doi:10.1128 / MMBR.63.3.570-620.1999. PMC 103747. PMID 10477309.
- ^ Roach, John (7 Haziran 2004). "Yarım Dünya'nın Oksijen Kaynağı Çok Az Kredi Alır". National Geographic Haberleri. Alındı 2016-04-04.
- ^ Lin, I .; Liu, W. Timothy; Wu, Chun-Chieh; Wong, George T. F .; Hu, Chuanmin; Chen, Zhiqiang; Wen-Der, Liang; Yang, Yih; Liu, Kon-Kee (2003). "Tropikal siklon tarafından tetiklenen gelişmiş okyanus birincil üretimi için yeni kanıtlar". Jeofizik Araştırma Mektupları. 30 (13). doi:10.1029 / 2003GL017141.
- ^ Sigman, D.M .; Hain, M.P. (2012). "Okyanusun Biyolojik Verimliliği" (PDF). Doğa Eğitimi Bilgisi. 3 (6): 1–16. Alındı 2015-06-01.
Derin klorofil maksimum (DCM), fotosentez için yeterli ışığın olduğu ve yine de aşağıdan önemli miktarda besin tedarikinin olduğu temas noktasında meydana gelir.
- ^ Mora, C .; et al. (2013). "21. Yüzyılda Okyanus Biyojeokimyasında Öngörülen Değişikliklere Karşı Biyotik ve İnsani Hassasiyet". PLOS Biyoloji. 11 (10): e1001682. doi:10.1371 / journal.pbio.1001682. PMC 3797030. PMID 24143135.
- ^ "Bilim adamları, Arktik Okyanusu'nda bir 'rejim değişikliği' oluyor diyor". phys.org. Alındı 16 Ağustos 2020.
- ^ Lewis, K. M .; Dijken, G. L. van; Arrigo, K. R. (10 Temmuz 2020). "Fitoplankton konsantrasyonundaki değişiklikler artık Arktik Okyanusu birincil üretiminin artmasına neden oluyor". Bilim. 369 (6500): 198–202. doi:10.1126 / science.aay8380. ISSN 0036-8075. PMID 32647002. S2CID 220433818. Alındı 16 Ağustos 2020.
- ^ Olson, J.M. ve Blankenship, R.E. (2005) "Fotosentezin evrimini düşünmek". İçinde: Fotosentezde Keşifler, 1073–1086. sayfalar, Springer. ISBN 9781402033247. doi:10.1007/1-4020-3324-9_95.
- ^ Blankenship, R. E., Sadekar, S. ve Raymond, J. (2007) "Anoksijenikten oksijenik fotosenteze evrimsel geçiş". İçinde: Sucul Fotoautotrofların Evrimi, eds P. G. Falkowski ve A. N. Knoll, New York: Academic Press, sayfalar 21–35. doi:10.1016 / B978-012370518-1 / 50004-7.
- ^ Hohmann-Marriott, M.F. ve Blankenship, R.E. (2011) "Fotosentezin Evrimi". Bitki biyolojisinin yıllık incelemesi, 62: 515-548. doi:10.1146 / annurev-arplant-042110-103811.
- ^ Kim, E., Harrison, J.W., Sudek, S., Jones, M.D., Wilcox, H.M., Richards, T.A., Worden, A.Z. ve Archibald, J.M. (2011) "Ökaryotik hayat ağacında yeni tanımlanmış ve çeşitli plastid taşıyan dal". Ulusal Bilimler Akademisi Bildiriler Kitabı, 108(4): 1496–1500. doi:10.1073 / pnas.1013337108.
- ^ Garcia-Mendoza, E. ve Ocampo-Alvarez, H. (2011) "Kahverengi alglerde ışık koruma Macrocystis pyrifera: evrimsel çıkarımlar ". Fotokimya ve Fotobiyoloji B Dergisi: Biyoloji, 104(1-2): 377–385. doi:10.1016 / j.jphotobiol.2011.04.004.
- ^ Shevela, D. (2011) "Siyanobakterilerle Maceralar: kişisel bir bakış açısı". Bitki biliminde sınırlar, 2: 28. doi:10.3389 / fpls.2011.00028.
- ^ Walsh PJ, Smith S, Fleming L, Solo-Gabriele H, Gerwick WH, editörler. (2 Eylül 2011). "Siyanobakteriler ve siyanobakteriyel toksinler". Okyanuslar ve İnsan Sağlığı: Denizlerden Kaynaklanan Riskler ve Çözümler. Akademik Basın. s. 271–296. ISBN 978-0-08-087782-2.
- ^ "Oksijenin Yükselişi - Astrobiology Magazine". Astrobiology Dergisi. 30 Temmuz 2003. Alındı 2016-04-06.
- ^ Flannery, D. T .; R.M. Walter (2012). "Archean püsküllü mikrobiyal paspaslar ve Büyük Oksidasyon Olayı: eski bir soruna yeni bakış açıları". Avustralya Yer Bilimleri Dergisi. 59 (1): 1–11. Bibcode:2012AuJES..59 .... 1F. doi:10.1080/08120099.2011.607849. S2CID 53618061.
- ^ Rothschild Lynn (Eylül 2003). "Yaşamın evrimsel mekanizmalarını ve çevresel sınırlarını anlayın". NASA. Arşivlenen orijinal 11 Mart 2012 tarihinde. Alındı 13 Temmuz 2009.
- ^ Nadis S (Aralık 2003). "Denizleri yöneten hücreler" (PDF). Bilimsel amerikalı. 289 (6): 52–3. Bibcode:2003SciAm.289f..52N. doi:10.1038 / bilimselamerican1203-52. PMID 14631732. Arşivlenen orijinal (PDF) 2014-04-19 tarihinde. Alındı 2019-07-11.
- ^ "Hiç Duymadığınız En Önemli Mikrop". npr.org.
- ^ Flombaum, P .; Gallegos, J. L .; Gordillo, R. A .; Rincon, J .; Zabala, L. L .; Jiao, N .; Karl, D. M .; Li, W. K. W .; Lomas, M. W .; Veneziano, D .; Vera, C. S .; Vrugt, J. A .; Martiny, A.C. (2013). "Deniz Siyanobakterileri Prochlorococcus ve Synechococcus'un mevcut ve gelecekteki küresel dağıtımları". Ulusal Bilimler Akademisi Bildiriler Kitabı. 110 (24): 9824–9829. Bibcode:2013PNAS..110.9824F. doi:10.1073 / pnas.1307701110. PMC 3683724. PMID 23703908.
- ^ Nabors, Murray W. (2004). Botanik'e Giriş. San Francisco, CA: Pearson Education, Inc. ISBN 978-0-8053-4416-5.
- ^ Allaby, M., ed. (1992). "Yosun". Kısa Botanik Sözlüğü. Oxford: Oxford University Press.
- ^ Grotewold, E. (2006). "Çiçek Pigmentlerinin Genetiği ve Biyokimyası". Bitki Biyolojisinin Yıllık İncelemesi. 57: 761–780. doi:10.1146 / annurev.arplant.57.032905.105248. PMID 16669781.
- ^ Lee, DW (2007) Doğanın paleti - bitki rengi bilimi. Chicago Press Üniversitesi
- ^ Biyoloji Kavramları: Ökaryotik Kökenler. OpenStax CNX. Erişim tarihi: 16 Temmuz 2020.
 Materyal, bir altında bulunan bu kaynaktan kopyalandı. Creative Commons Attribution 4.0 Uluslararası Lisansı.
Materyal, bir altında bulunan bu kaynaktan kopyalandı. Creative Commons Attribution 4.0 Uluslararası Lisansı. - ^ "kloroplast". Çevrimiçi Etimoloji Sözlüğü.
- ^ Temel Biyoloji (18 Mart 2016). "Bakteriler".
- ^ Patrick J. Keeling (2004). "Plastidlerin ve ev sahiplerinin çeşitliliği ve evrimsel tarihi". Amerikan Botanik Dergisi. 91 (10): 1481–1493. doi:10.3732 / ajb.91.10.1481. PMID 21652304.
- ^ DeLong, E.F .; Beja, O. (2010). "Işıkla çalışan proton pompası proteorhodopsin, zor zamanlarda bakteriyel hayatta kalmayı artırır". PLOS Biyoloji. 8 (4): e1000359. doi:10.1371 / journal.pbio.1000359. PMC 2860490. PMID 20436957. e1000359.
- ^ a b Gómez-Consarnau, L .; Raven, J.A .; Levine, N.M .; Cutter, L.S .; Wang, D .; Seegers, B .; Arístegui, J .; Fuhrman, J.A .; Gasol, J.M .; Sañudo-Wilhelmy, S.A. (2019). "Mikrobiyal rodopinler, denizde tutulan güneş enerjisine en büyük katkı sağlayan unsurlardır". Bilim Gelişmeleri. 5 (8): eaaw8855. Bibcode:2019SciA .... 5.8855G. doi:10.1126 / sciadv.aaw8855. PMC 6685716. PMID 31457093.
- ^ Oren, Aharon (2002). "Aşırı derecede halofilik Arkea ve Bakterilerin moleküler ekolojisi". FEMS Mikrobiyoloji Ekolojisi. 39 (1): 1–7. doi:10.1111 / j.1574-6941.2002.tb00900.x. ISSN 0168-6496. PMID 19709178.
- ^ Béja, O .; Aravind, L .; Koonin, E.V .; Suzuki, M.T .; Hadd, A .; Nguyen, L.P .; Jovanovich, S.B .; Gates, C.M .; Feldman, R.A .; Spudich, J.L .; Spudich, E.N. (2000). "Bakteriyel rodopsin: denizde yeni bir fototrofi türü için kanıt". Bilim. 289 (5486): 1902–1906. Bibcode:2000Sci ... 289.1902B. doi:10.1126 / science.289.5486.1902. PMID 10988064. S2CID 1461255.
- ^ "Arkadaşlarla Röportajlar: Ed Delong". Amerikan Mikrobiyoloji Akademisi. Arşivlenen orijinal 7 Ağustos 2016. Alındı 2 Temmuz 2016.
- ^ Pilli Bakteriler, Popular Science, Ocak 2001, Sayfa 55.
- ^ Béja, O .; Aravind, L .; Koonin, E.V .; Suzuki, M.T .; Hadd, A .; Nguyen, L.P .; Jovanovich, S.B .; Gates, C.M .; Feldman, R.A .; Spudich, J.L .; Spudich, E.N. (2000). "Bakteriyel rodopsin: denizde yeni bir fototrofi türü için kanıt". Bilim. 289 (5486): 1902–1906. Bibcode:2000Sci ... 289.1902B. doi:10.1126 / science.289.5486.1902. PMID 10988064.
- ^ Boeuf, Dominique; Audic, Stéphane; Brillet-Guéguen, Loraine; Caron, Christophe; Jeanthon, Hıristiyan (2015). "MicRhoDE: mikrobiyal rodopsin çeşitliliği ve evriminin analizi için küratörlü bir veritabanı". Veri tabanı. 2015: bav080. doi:10.1093 / veritabanı / bav080. ISSN 1758-0463. PMC 4539915. PMID 26286928.
- ^ Yawo, Hiromu; Kandori, Hideki; Koizumi, Amane (5 Haziran 2015). Optogenetik: Işığı Algılayan Proteinler ve Uygulamaları. Springer. s. 3–4. ISBN 978-4-431-55516-2. Alındı 30 Eylül 2015.
- ^ Küçük bir deniz mikropu iklim değişikliğinde büyük rol oynayabilir Güney Kaliforniya Üniversitesi, Basın Odası, 8 Ağustos 2019.
- ^ DasSarma, Shiladitya; Schwieterman, Edward W. (11 Ekim 2018). "Dünya üzerindeki mor retina pigmentlerinin erken evrimi ve dış gezegen biyo-imzalarının etkileri". Uluslararası Astrobiyoloji Dergisi: 1–10. arXiv:1810.05150. Bibcode:2018arXiv181005150D. doi:10.1017 / S1473550418000423. ISSN 1473-5504. S2CID 119341330.
- ^ Sparks, William B .; DasSarma, S .; Reid, I.N. (Aralık 2006). "İlkel Fotosentetik Sistemler Arasındaki Evrimsel Rekabet: Erken bir mor Dünyanın varlığı mı?". Amerikan Astronomi Derneği Toplantısı Özetleri. 38: 901. Bibcode:2006AAS ... 209.0605S.
- ^ Javed, M.R., Bilal, M.J., Ashraf, M.U.F., Waqar, A., Mehmood, M.A., Saeed, M. and Nashat, N. (2019) "Biyoyakıt Üretimi için Hammadde Olarak Mikroalg: Mevcut Durum ve Gelecek Beklentiler" İçinde: Enerji Araştırma ve Geliştirmede İlk 5 Katkı, üçüncü baskı, bölüm 2, Avid Science. ISBN 978-93-88170-77-2. Materyal, bir altında bulunan bu kaynaktan kopyalandı. Creative Commons Attribution 4.0 Uluslararası Lisansı.
- ^ Guiry MD (Ekim 2012). "Kaç yosun türü var?". Journal of Phycology. 48 (5): 1057–63. doi:10.1111 / j.1529-8817.2012.01222.x. PMID 27011267. S2CID 30911529.
- ^ a b Guiry, M.D .; Guiry, G.M. (2016). "Algaebase". www.algaebase.org. Alındı 20 Kasım 2016.
- ^ D. Thomas (2002). Yosunlar. Yaşam Serisi. Doğal Tarih Müzesi, Londra. ISBN 978-0-565-09175-0.
- ^ Hoek, Christiaan; den Hoeck, Hoeck Van; Mann, David; Jahns, H.M. (1995). Yosun: psikolojiye giriş. Cambridge University Press. s. 166. ISBN 9780521316873. OCLC 443576944.
- ^ Yool, A .; Tyrrell, T. (2003). "Okyanusun silikon döngüsünü düzenlemede diatomların rolü". Küresel Biyojeokimyasal Çevrimler. 17 (4): yok. Bibcode:2003GBioC..17.1103Y. CiteSeerX 10.1.1.394.3912. doi:10.1029 / 2002GB002018.
- ^ Soluduğunuz Hava? Bunu Bir Diatom Yaptı
- ^ "Diatomlar hakkında daha fazla bilgi". California Üniversitesi Paleontoloji Müzesi. Arşivlenen orijinal 2012-10-04 tarihinde. Alındı 2019-07-11.
- ^ Bu alacakaranlık kuşağı karanlık, sulu ve evet aynı zamanda entrikalarla dolu NASA Blogu, 21 Ağustos 2018.
- ^ Smith, H.E.K .; et al. (2012), "Biscay Körfezi'nde kış aylarında düşük CaCO3 doygunluğunda yoğun şekilde kalsifiye kokolitoforların baskınlığı", Ulusal Bilimler Akademisi Bildiriler Kitabı, 109 (23): 8845–8849, Bibcode:2012PNAS..109.8845S, doi:10.1073 / pnas.1117508109, PMC 3384182, PMID 22615387
- ^ "Biyocoğrafya ve mikroorganizmaların yayılması: protistleri vurgulayan bir inceleme", Acta Protozoologica, 45 (2): 111–136, 2005
- ^ Yunev, O.A .; et al. (2007), "Kültürel ötrofikasyon ve iklim değişikliklerine yanıt olarak Batı Karadeniz sahanlığındaki besin ve fitoplankton eğilimleri", Nehir Ağzı, Kıyı ve Raf Bilimi, 74 (1–2): 63–67, Bibcode:2007ECSS ... 74 ... 63Y, doi:10.1016 / j.ecss.2007.03.030
- ^ Marka, Larry E .; Campbell, Lisa; Bresnan, Eileen (2012). "Karenia: Toksik bir cinsin biyolojisi ve ekolojisi ". Zararlı Algler. 14: 156–178. doi:10.1016 / j.hal.2011.10.020.
- ^ Olsen GJ, Woese CR, Overbeek R (1994). "(Evrimsel) değişim rüzgarları: mikrobiyolojiye yeni yaşam soluyor". Bakteriyoloji Dergisi. 176 (1): 1–6. doi:10.2172/205047. PMC 205007. PMID 8282683.
- ^ Bork, P., Bowler, C., De Vargas, C., Gorsky, G., Karsenti, E. ve Wincker, P. (2015) "Tara Oceans, planktonu gezegen ölçeğinde inceliyor ". doi:10.1126 / science.aac5605.
- ^ Gawryluk, Ryan M.R .; Del Campo, Javier; Okamoto, Noriko; Strassert, Jürgen F.H .; Lukeš, Julius; Richards, Thomas A .; Worden, Alexandra Z .; Santoro, Alyson E .; Keeling, Patrick J. (2016). "Deniz Diplonemidlerinin Morfolojik Tanımlanması ve Tek Hücreli Genomiği". Güncel Biyoloji. 26 (22): 3053–3059. doi:10.1016 / j.cub.2016.09.013. PMID 27875688.
- ^ Faktorová, D., Dobáková, E., Peña-Diaz, P. ve Lukeš, J., 2016. Basitten süper karmaşıklığa: öglenozoan protistlerinin mitokondriyal genomları. F1000Research, 5. doi:10.12688 / f1000research.8040.1. Materyal, bir altında bulunan bu kaynaktan kopyalandı. Creative Commons Attribution 4.0 Uluslararası Lisansı.
- ^ De Vargas, C., Audic, S., Henry, N., Decelle, J., Mahé, F., Logares, R., Lara, E., Berney, C., Le Bescot, N., Probert, I ., Carmichael, M. ve 44 diğerleri (2015) "Güneşli okyanusta ökaryotik plankton çeşitliliği. Bilim", 348(6237): 1261605. doi:10.1126 / science.1261605.
- ^ Starckx, Senne (31 Ekim 2012) Güneşte bir yer - Geel'deki araştırmacılara göre algler geleceğin mahsulü Flanders Today, Erişim tarihi: 8 Aralık 2012
- ^ Wernberg, T., Krumhansl, K., Filbee-Dexter, K. ve Pedersen, M.F. (2019) "Dünyadaki kelp ormanlarının durumu ve eğilimleri". İçinde: Dünya denizleri: çevresel bir değerlendirme, 57–78. sayfalar). Akademik Basın. doi:10.1016 / B978-0-12-805052-1.00003-6.
- ^ Mann, K.H. 1973. Deniz yosunları: üretkenlikleri ve büyüme stratejileri. Science 182: 975-981.
- ^ Kindersley Dorling (2011). Okyanusun Resimli Ansiklopedisi. Dorling Kindersley. ISBN 978-1-4053-3308-5.
- ^ Tunnell, John Wesley; Chávez, Ernesto A .; Withers, Kim (2007). Güney Meksika Körfezi'ndeki mercan resifleri. Texas A&M University Press. s. 91. ISBN 978-1-58544-617-9.
- ^ Orth, R.J., Carruthers, T.J., Dennison, W.C., Duarte, C.M., Fourqurean, J.W., Heck, K.L., Hughes, A.R., Kendrick, G.A., Kenworthy, W.J., Olyarnik, S. and Short, F.T. (2006) "Deniz çayırı ekosistemleri için küresel bir kriz". Biyobilim, 56(12): 987–996. Sayfalar. doi:10.1641 / 0006-3568 (2006) 56 [987: AGCFSE] 2.0.CO; 2
- ^ van der Heide, T .; van Nes, E. H .; van Katwijk, M. M .; Olff, H .; Smolders, A.J.P. (2011). Romanuk, Tamara (ed.). "Deniz çayırı ekosistemlerinde olumlu geri bildirimler: büyük ölçekli ampirik verilerden kanıtlar". PLOS ONE. 6 (1): e16504. Bibcode:2011PLoSO ... 616504V. doi:10.1371 / journal.pone.0016504. PMC 3025983. PMID 21283684.
- ^ "Mangal (Mangrov)". Mildred E. Mathias Botanik Bahçesi. Alındı 11 Temmuz 2013.
- ^ "Kıyı Tuzlu Bataklığı". Mildred E. Mathias Botanik Bahçesi. Alındı 11 Temmuz 2013.
- ^ "Deniz biyoçeşitliliği ile ilgili gerçekler ve rakamlar". Deniz biyoçeşitliliği. UNESCO. 2012. Alındı 11 Temmuz 2013.
- ^ Russell, F. S .; Yonge, C.M. (1928). Denizler. Frederick Warne. s. 225–227.
- ^ Voss, Maren; Bange, Hermann W .; Dippner, Joachim W .; Middelburg, Jack J .; Montoya, Joseph P .; Ward, Bess (2013). "Deniz nitrojen döngüsü: son keşifler, belirsizlikler ve iklim değişikliğinin potansiyel ilgisi". Royal Society B'nin Felsefi İşlemleri. 368 (1621): 20130121. doi:10.1098 / rstb.2013.0121. PMC 3682741. PMID 23713119.
- ^ Stow, Dorrik (2004). Okyanusların Ansiklopedisi. Oxford University Press. ISBN 978-0-19-860687-1.
- ^ Giri C, Ochieng E, Tieszen LL, Zhu Z, Singh A, Loveland T, ve diğerleri. (2011) "Dünya gözlem uydu verilerini kullanarak dünyadaki mangrov ormanlarının durumu ve dağılımı". Küresel Ekoloji ve Biyocoğrafya, 20(1):154–159. doi:10.1111 / j.1466-8238.2010.00584.x
- ^ Thomas, N., Lucas, R., Bunting, P., Hardy, A., Rosenqvist, A. ve Simard, M. (2017) "Küresel mangrov ormanı değişiminin dağıtımı ve etmenleri, 1996–2010". PLOS ONE, 12(6): e0179302. doi:10.1371 / journal.pone.0179302
- ^ Kısa, F.T. ve Frederick, T. (2003) Deniz çayırlarının dünya atlası, University of California Press, sayfa 24. ISBN 9780520240476
- ^ Froese, Rainer ve Pauly, Daniel, editörler. (2009). "Phycodurus eques" içinde FishBase. Temmuz 2009 versiyonu.
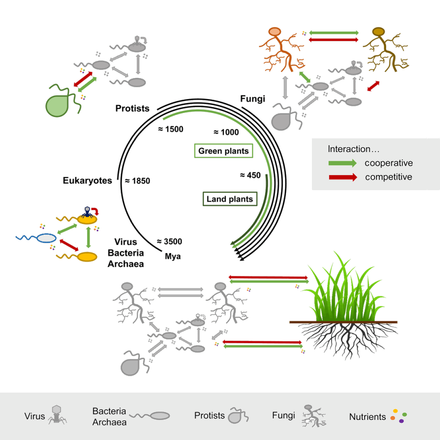
- ^ Hassani, M.A., Durán, P. and Hacquard, S. (2018) "Bitki holobiont içindeki mikrobiyal etkileşimler". Mikrobiyom, 6(1): 58. doi:10.1186 / s40168-018-0445-0. Materyal, bir altında bulunan bu kaynaktan kopyalandı. Creative Commons Attribution 4.0 Uluslararası Lisansı.
- ^ Lucking, R., Huhndorf, S., Pfister, D.H., Plata, E.R. ve Lumbsch, H.T. (2009) "Mantarlar doğru yolda gelişti". Mikoloji, 101(6): 810–822. doi:10.3852/09-016.
- ^ Heckman, D.S., Geiser, D.M., Eidell, B.R., Stauffer, R.L., Kardos, N.L. ve Hedges, S.B. (2001) "Toprağın mantar ve bitkiler tarafından erken kolonileştirilmesine ilişkin moleküler kanıtlar". Bilim, 293(5532): 1129–1133. doi:10.1126 / bilim.1061457.
daha fazla okuma
- Falkowski, Paul (Ed.) (2013) Denizde Birincil Verimlilik Springer. ISBN 9781468438901.
- Falkowski, Paul ve Kuzgun, John A. (2013) Sucul Fotosentez İkinci baskı revize edildi, Princeton University Press. ISBN 9781400849727.
- Falkowski P ve Knoll AH (2011) Denizdeki Birincil Üreticilerin Gelişimi Akademik Basın. ISBN 9780080550510.
- Kirk, John T. O. (2010) Sucul Ekosistemlerde Işık ve Fotosentez Üçüncü baskı revize edildi, Cambridge University Press. ISBN 9781139493918.

