Simbiyodinyum - Symbiodinium - Wikipedia
- Bu, bazen Zoox adı verilen cinsle ilgilidir. Şirket için bkz. Zoox (şirket)
| Simbiyodinyum | |
|---|---|
 | |
| bilimsel sınıflandırma | |
| Alan adı: | |
| (rütbesiz): | |
| (rütbesiz): | |
| Şube: | |
| Sınıf: | |
| Sipariş: | |
| Aile: | |
| Cins: | Simbiyodinyum Freudenthal, 1962 [1] |
| Türler | |
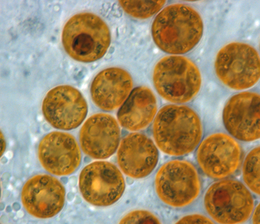
Simbiyodinyum bir cins Dinoflagellatlar en büyük ve en yaygın grubu kapsayan endosimbiyotik dinoflagellatlar bilinmektedir. Bunlar tek hücreli yosun genellikle ikamet endoderm tropikal cnidarians gibi mercanlar, Deniz lalesi, ve Deniz anası ürünleri nerede fotosentetik işleme konakçıda inorganik moleküller ile değiştirilir. Ayrıca çeşitli türler tarafından barındırılırlar. Demosponges, yassı kurtlar, yumuşakçalar benzeri dev istiridye, foraminifera (Soritids ), ve bazı siliatlar. Genellikle bunlar Dinoflagellatlar ana hücreye giriniz fagositoz, devam et hücre içi ortakyaşlar, çoğalır ve çevreye yayılır. Bunun istisnası, bu simbiyoların hücreler arası (hücreler arasında) olduğu çoğu yumuşakçada görülür. Symbiodinium ile ilişkili Cnidarians, çoğunlukla sıcak oligotrofik (besin açısından fakir), genellikle baskın bileşenleri olduğu deniz ortamları Bentik topluluklar. Bu dinoflagellatlar bu nedenle en bol bulunanlar arasındadır ökaryotik bulunan mikroplar mercan kayalığı ekosistemler.
Symbiodinium halk dilinde zooxanthellae ve bu cinsteki alglerle simbiyotik hayvanların "zooksantellat" olduğu söylenir. Terim, diatomlar ve diğer dinoflagellatlar dahil olmak üzere herhangi bir altın-kahverengi endosimbiyiyonu ifade etmek için gevşek bir şekilde kullanılmıştır. Taksonomik olarak çeşitli simbiyotik ilişkilerin aşırı derecede genelleştirilmesinin neden olduğu kafa karışıklığı nedeniyle, terimin bilimsel literatürde kullanılmaya devam edilmesi önerilmez.[2]
2018 yılında sistematiği Sembiyodiniaceae revize edildi ve farklı Clades yedi cinse yeniden atandı.[3] Bu revizyonu takiben isim Simbiyodinyum artık yalnızca daha önce Sınıf A olarak sınıflandırılan türler için bir cins adıdır[3]. Diğer sınıflar, farklı cinsler olarak yeniden sınıflandırıldı (aşağıdaki Moleküler Sistematiğe bakınız).

Hücre içi ortakyaşlar

Birçok Symbiodinium, öncelikle karşılıklı olarak rolleri ile bilinir. endosymbionts. Konakçılarda, genellikle santimetre kare başına yüzbinler ile milyonlar arasında değişen yüksek yoğunluklarda ortaya çıkarlar.[4] Başarılı yüzme kültürü jimnodinioid mercandan alınan hücreler "zooxanthellae" nin aslında dinoflagellat olduğunun keşfedilmesine yol açtı.[5][6] Her biri Simbiyodinyum hücre kokoid misafirperver (bir konak hücrede yaşayan) ve konakçı hücreden kaynaklanan bir zarla çevrili plazmalemma sırasında fagositoz (Şekil 2B ve 3). Bu zar muhtemelen protein içeriğinde bazı modifikasyonlara uğrar ve bu, fago-lizozom füzyonunu sınırlama veya önleme işlevi görür.[7][8][9] Symbiont içeren vakuol yapı bu nedenle simbiyozom. Tek bir ortakyaşam hücresi, her bir simbiyozomu işgal eder. Bu zarın bölünen bir ortakyaşama hücresini barındıracak şekilde nasıl genişlediği belirsizdir. Normal koşullar altında, simbiont ve konak hücreler, her iki partnerin de büyümesini ve çoğalmasını sağlayan organik ve inorganik molekülleri değiştirirler.
Doğal hizmetler ve ekonomik değer
Simbiyodinyum en çok çalışılan ortakyaşarlardan biridir. Resif oluşturan mercanlarla olan karşılıklı ilişkileri, çok çeşitli ve üretken bir ekosistemin temelini oluşturur. Mercan resiflerinin süs, geçimlik ve ticari balıkçılık, turizm ve rekreasyon, fırtınalardan kıyı koruma, farmasötik geliştirme için biyoaktif bileşikler kaynağı ve daha fazlası şeklinde her yıl yüz milyarlarca dolar değerinde ekonomik faydaları vardır.[10]
Mercan ağartma
Çalışma Simbiyodinyum biyoloji, büyük ölçüde küresel mercan resifindeki düşüşü anlama arzusuyla yönlendirilir. Yaygın resif bozulması için ana mekanizma, stres kaynaklı olmuştur. mercan ağartma alışılmadık derecede yüksek deniz suyu sıcaklık. Ağartma, mercanın ve yosun içindeki ortakyaşama ve / veya klorofil kaybının ayrılmasıdır, bu da hayvanın pigmentasyonunda ani bir kayba neden olur. Birçok Simbiyodinyum-cnidarian dernekleri, deniz yüzeyi sıcaklıklarının sürekli yükselmesinden etkilenir,[11] ancak yüksek ışık seviyelerine maruz kalmadan da kaynaklanabilir (dahil UVR ),[12][13] aşırı düşük sıcaklıklar,[14] düşük tuzluluk[15] ve diğer faktörler.[16] Ağartılmış durum, azalmış konak kireçlenmesiyle ilişkilidir,[17] artan hastalık duyarlılığı[18] ve eğer uzun sürerse, kısmi veya toplam ölüm oranı.[19] Tek bir ağartma olayından kaynaklanan ölüm oranı, 2015'te olduğu gibi küresel ölçekte olabilir. Dünya genelinde sıcaklıklar artmaya devam ettikçe bu olayların daha yaygın ve şiddetli olacağı tahmin ediliyor.[20] Bir asistanın fizyolojisi Simbiyodinyum türler genellikle bir mercanın ağartma duyarlılığını düzenler.[21][22] Bu nedenle, önemli miktarda araştırma, termal toleransın fizyolojik temelini karakterize etmeye odaklanmıştır.[23][24][25][26] ve termal olarak toleranslı ortakyaşam türlerinin ekolojisini ve dağılımını belirlemede.[27][28][29]
Simbiyodinyum siper strese dayanıklı bir türdür ve birçok mercan türü ile karşılıklı ilişkiler kurabilir. Küresel olarak mercanda az sayıda bulunur ve Andaman Denizi, suyun diğer bölgelerine göre yaklaşık 4 ° C (7 ° F) daha sıcak olduğu Hint Okyanusu.[30] İçinde Karayib Denizi 2005 yılının sonlarında, su sıcaklığı birkaç ay boyunca yükseldi ve S. trenchiNormalde bol olmayan bir ortakyaşam, daha önce gözlenmediği birçok mercanda ikamet etti. Bu mercanlar ağartmadı. İki yıl sonra, normalde Karayipler'de bulunan türler tarafından büyük ölçüde simbiont olarak değiştirildi.[28]
S. thermophilum yakın zamanda, mercanların içindeki alg popülasyonunun büyük bir kısmını oluşturduğu bulunmuştur. Basra Körfezi. Aynı zamanda Umman Körfezi ve Kızıldeniz'de çok daha düşük bir konsantrasyonda mevcuttur. Bu türe ev sahipliği yapan mercan, küresel olarak mercan resiflerinin 31 ° C (88 ° F) sıcaklığından çok daha sıcak olan Basra Körfezi'nin 35 ° C (95 ° F) sularını tolere edebildi.[31]
Moleküler sistematiği

DNA dizisi karşılaştırmasının ortaya çıkışı, tüm organizmaların sıralanması ve adlandırılmasında bir yeniden doğuş başlattı. Bu metodolojinin uygulanması, uzun süredir devam eden inancın tersine çevrilmesine yardımcı oldu: Simbiyodinyum tek bir türden oluşuyordu, bu süreç, kültürlenmiş izolatların morfolojik, fizyolojik ve biyokimyasal karşılaştırmalarıyla ciddi bir şekilde başladı. Şu anda, genetik belirteçler yalnızca ekolojik kalıpları tanımlamak ve bu grubun morfolojik olarak şifreli üyeleri arasındaki evrimsel ilişkileri çıkarmak için kullanılır. En başta moleküler sistematiği Simbiyodinyum ekolojik olarak ilgili çeşitlilik birimlerini (yani türler) çözmektir.
"Sınıflar" arasındaki filogenetik eşitsizlik
En eski ribozomal gen dizisi verileri şunu gösterdi: Simbiyodinyum farklı cinslerden, ailelerden ve hatta düzenlerden diğer dinoflagellatlarda görülen genetik farklılığa benzer soylara sahipti.[32] A, B, C vb. Sınıflar arasındaki bu büyük filogenetik eşitsizlik, mitokondriyal genin kodlama dizilerinin analizleriyle doğrulanmıştır. sitokrom c oksidaz alt birimi I Dinophyceae arasında.[33] Bu sınıf gruplamalarının çoğu, çok sayıda üremeyle izole edilmiş, genetik olarak farklı soylardan oluşur (bkz. 'Türlerin çeşitliliği' ), farklı ekolojik ve biyocoğrafik dağılımlar sergileyen (bkz. "Coğrafi dağılımlar ve" çeşitlilik "modelleri ).
Yakın zamanda, Sembiyodiniaceae sınıfındaki bu farklı sınıflar, yalnızca olmasa da, yedi cinse yeniden atandı: Simbiyodinyum (A sınıfı), Breviolum (sınıf B), Cladocopium (C sınıfı), Durusdinium (D sınıfı), Efrenyum (E sınıfı), Fugacium (F sınıfı) ve Gerakladyum (G sınıfı).[3]
Türlerin çeşitliliği

Bu cinste tür çeşitliliğinin tanınması, türlerin teşhisinde yararlı olan morfolojik ve biyokimyasal özelliklerin tanımlanmasındaki zorluklar nedeniyle uzun yıllar boyunca sorunlu kaldı.[34] Şu anda, filogenetik, ekolojik ve popülasyon genetik verileri, Simbiyodinyum Biyolojik, Evrimsel ve Ekolojik Tür Kavramları ile uyumlu ayrı varlıklara.[35][36] Genetik temelli çeşitlilik ölçütlerinin çoğu, bir genetik belirtecin (örneğin LSU, ITS2 veya cp23S) analizinden tahmin edilmiştir, ancak son çalışmalarda bunlar ve diğer belirteçler kombinasyon halinde analiz edilmiştir. Nükleer, mitokondriyal ve kloroplast DNA'sı arasında bulunan yüksek uyum, ekolojik ve popülasyon genetik verileriyle birleştirilmiş hiyerarşik bir filogenetik şemanın, üreme yoluyla izole edilmiş soyları, yani türler gibi isimlendirmeyi kesin olarak tanıyabileceğini ve atayabileceğini savunuyor.[kaynak belirtilmeli ]
Ek filogenetik belirteçlerin analizi, bazılarının Simbiyodinyum başlangıçta ITS dizilerindeki küçük farklılıklar ile tespit edilenler aynı türlerin üyelerini içerebilir[36] oysa diğer durumlarda, iki veya daha fazla genetik olarak farklı soy, aynı atalara ait ITS sekansına sahip olabilir.[37][38] Ana tür kavramları bağlamında incelendiğinde,[39] ITS2 sekans verilerinin çoğu tür çeşitliliği için makul bir vekil sağlar.[35][36][40] Şu anda, ITS2 türleri yüzleri buluyor, ancak dünyadaki çoğu simbiyotik cnidaria topluluğu hala kapsamlı örneklemeye ihtiyaç duyuyor. Dahası, soritid foraminiferlerin eşit derecede farklı tür toplulukları ile birlikte bulunan çok sayıda benzersiz tür var gibi görünüyor.[41] yanı sıra diğerleri Simbiyodinyum sadece serbest yaşayan ve çeşitli, genellikle bentik habitatlarda bulunan.[42] Ekolojik olarak şifrelenmiş bu türlerin potansiyel tür çeşitliliği göz önüne alındığında SimbiyodinyumToplam tür sayısı hiçbir zaman doğru bir şekilde değerlendirilemeyebilir.[41]
Klon çeşitliliği ve popülasyon genetiği
Mikro uydu belirteçlerinin kullanımıyla, çok odaklı genotipler tek bir klonal hattını tanımlamak Simbiyodinyum konak doku örneklerinden çözülebilir. Çoğu bireysel koloninin tek bir multilocus genotipi (yani klon) barındırdığı görülmektedir.[43][44] Koloniler içinde kapsamlı örnekleme, birçok koloninin homojen (klonal) Simbiyodinyum nüfus. Bazı kolonilerde ek genotipler meydana gelir, ancak nadiren iki veya üçten fazlası bulunur. Aynı kolonide bulunduğunda, çoklu klonlar genellikle dar örtüşme bölgeleri sergiler.[44] Bir resif üzerinde birbirine bitişik koloniler aynı klonları barındırabilir, ancak konakçı popülasyon boyunca belirli bir klon çeşitliliği Simbiyodinyum tür potansiyel olarak büyüktür ve cinsel rekombinasyonun ürünü olan rekombinant genotipleri içerir. Bir klon, bir kolonide aylarca ve yıllarca baskın kalma eğilimindedir, ancak bazen yerinden olabilir veya değiştirilebilir. Klon dağılımını inceleyen birkaç çalışma, çoğu genotipin sınırlı coğrafi dağılımlara sahip olduğunu, ancak bu dağılımın ve gen akışının muhtemelen konakçı yaşam öyküsünden ve ortakyaşama edinme modundan (örneğin, yatay ve dikey) etkilendiğini bulmuştur.[kaynak belirtilmeli ]
Tür çeşitliliği, ekoloji ve biyocoğrafya
Coğrafi dağılımlar ve çeşitlilik modelleri

Simbiyodinyum mikro-ökaryot fizyolojisi ve ekolojisini çeşitli nedenlerle incelemek için belki de en iyi gruptur. Birincisi, mevcut filogenetik ve popülasyon genetik belirteçleri, genetik çeşitliliğinin geniş uzamsal ve zamansal ölçeklerde ayrıntılı incelenmesine izin verir. Ayrıca, büyük miktarlarda Simbiyodinyum hücreler, kendilerini barındıran konakçıların toplanmasıyla kolayca elde edilir. Son olarak, hayvanlarla ilişkileri, ekolojik dağılımları karşılaştırmak ve karşılaştırmak için ek bir eksen sağlar.[kaynak belirtilmeli ]
Değerlendirme için en eski genetik yöntemler Simbiyodinyum çeşitlilik, cinsi evrimsel olarak farklı soylara ayıran düşük çözünürlüklü moleküler belirteçlere dayanıyordu ve buna "kladlar" deniyordu. Coğrafi dağılım ve baskınlığın önceki karakterizasyonları, genetik çözümlemenin sınıf düzeyine odaklanmıştır, ancak tür düzeyinde çeşitliliğin daha ayrıntılı değerlendirmelerine ihtiyaç vardır. Belirli bir sınıfın üyeleri her yerde bulunabilse de, her bir kuşak içindeki tür çeşitliliği potansiyel olarak büyüktür, her tür genellikle dağılma yetenekleri, ev sahibi biyocoğrafyası ve dış çevre koşulları ile ilgili olarak farklı ekolojik ve coğrafi dağılımlara sahiptir. Az sayıda tür, çok az simbiyotik hayvanın meydana geldiği ılıman ortamlarda ortaya çıkar. Sonuç olarak, bu yüksek enlem ilişkileri, türlere oldukça özel olma eğilimindedir.[kaynak belirtilmeli ]

Farklı ekolojik loncalara atanan tür çeşitliliği
Büyük çeşitlilik Simbiyodinyum Genetik analizlerle ortaya çıkan, rastgele olmayan bir şekilde dağıtılır ve farklı ekolojik alışkanlıklara sahip birkaç loncadan oluşuyor gibi görünmektedir. Birçoğunun Simbiyodinyum genetik olarak karakterize edilen, çoğu konukçuya özgü, karşılıklı görüşlülük ve konukçularına hakimdir.[45] Diğerleri, hakim dış çevre koşulları (örneğin, yüksek ışık ve düşük ışık) altında rekabetçi aşağılık nedeniyle düşük bolluklu arka plan popülasyonları olarak kalan uyumlu ortakyaşları temsil edebilir.[46] Bazıları ayrıca fizyolojik stres dönemlerinde çoğalabilen ve normal yerleşik ortakyaşamın yerini alabilen ve orijinal ortakyaşama ile değiştirilmeden önce konakçının dokularında aylar ila yıllarca bol kalan fırsatçı türleri de içerebilir.[28][47][48] Ayrıca, normal olarak ev sahibi yetişkin kolonilerle ilişkili ortakyaşarlarla değiştirilene kadar, ev sahibi gençlerde hızla enfekte olan ve popülasyonlar oluşturanlar da vardır.[49] Son olarak, başka bir grup var gibi görünüyor Simbiyodinyum Endosimbiyoz oluşturamayan, ancak hayvanın çevresindeki ortamlarda var olan veya diğer substratlarla (yani makro-algal yüzeyler, tortu yüzeyi) yakın ilişki içinde olan[42][50] Simbiyodinyum fonksiyonel gruplardan 2, 3 ve 4, kolayca kültürlendikleri için var oldukları bilinmektedir, ancak bu yaşam öykülerine sahip türler, çevrede düşük bollukları nedeniyle çalışmak zordur.
Serbest yaşayan ve "simbiyotik olmayan" popülasyonlar
Belgelenmiş serbest yaşam popülasyonlarına dair birkaç örnek vardır. Simbiyodinyum.[42] Çoğu konukçu larvanın başlangıçta ortakyaşalarını çevreden alması gerektiği göz önüne alındığında, yaşayabilir Simbiyodinyum hücreler, konağın dışında meydana gelir. Hareketli faz muhtemelen dış çevrede önemlidir ve konakçı larvaların hızlı enfeksiyonunu kolaylaştırır. "Yakalama damarları" olarak yerleştirilen aposymbiyotik konakçı poliplerin kullanımı ve moleküler tekniklerin uygulanması, Symbiodinium'un çevresel kaynaklarının saptanmasına izin vermiştir.[48][51] Araştırmacılar, kullanılan bu yöntemlerle farklı türlerin çeşitli bentik yüzeylerdeki dağılımını çözebilir.[50] ve su kolonunda asılı hücre yoğunlukları.[52] Çevreden kültürlenen hücrelerin genetik kimlikleri genellikle konakçılarda bulunanlardan farklıdır. Bunlar muhtemelen endosimbiyoz oluşturmazlar ve tamamen özgür yaşarlar; "dağılan" simbiyotik türlerden farklıdırlar.[45] Bu çevresel popülasyonların "özel hayatları" ve bunların ekolojik işlevleri hakkında daha fazla bilgi edinmek, bu büyük cins içindeki üyeler arasındaki çeşitlilik, dağılma başarısı ve evrim hakkındaki bilgilerimizi daha da ileriye taşıyacaktır.
Kültür
Belirli Simbiyodinyum suşlar ve / veya türler daha kolay kültürlenir ve suni veya takviye edilmiş deniz suyu ortamında (ör. ASP – 8A, F / 2) onlarca yıl sürebilir. Kültürlenmiş izolatların aynı koşullar altında karşılaştırılması, morfoloji, boyut, biyokimya, gen ifadesi, yüzme davranışı, büyüme oranları vb.[53][54][55] Bu öncü karşılaştırmalı yaklaşım, bu cinsin tek bir türden daha fazlasını içerdiğini kabul etmede yavaş bir paradigma değişimi başlattı.
Kültür, seçici bir süreçtir ve birçok Simbiyodinyum Yapay ortamda büyüyen izolatlar, normal olarak belirli bir konakçı ile ilişkili türler için tipik değildir. Aslında, çoğu konakçıya özgü tür henüz kültürlenmemiştir. Ortaya çıkan kültürü baskın ve ekolojik olarak ilgili ortakyayın kimliğiyle eşleştirmek için kaynak koloniden genetik analiz numuneleri alınmalıdır ve başlangıçta hayvan tarafından barındırılan ekolojik olarak ilgili ortakyaşam.[45][56][57]
Yaşam döngüsü

Yaşam döngüsü Simbiyodinyum ilk olarak kültür ortamında büyüyen hücrelerden tanımlanmıştır. Log faz büyümesinde olan izolatlar için, bölünme oranları her 1-3 günde bir gerçekleşir. Simbiyodinyum küresel veya kokoid, morfoloji ve daha küçük kamçılı hareketli mastigot aşaması arasında değişen hücreler (bkz. "Morfolojik Özellikler"). Her bir morfolojik durumun diğerine nasıl geçtiğini açıklayan birkaç benzer şema yayınlanırken, en zorlayıcı yaşam öyküsü rekonstrüksiyonu, ışık ve elektron mikroskobu ve nükleer boyama kanıtlarından çıkarıldı.[58] Eşeysiz yayılma sırasında (bazen mitotik veya bitkisel büyüme olarak da adlandırılır), hücreler karanlıkta bir diel karyokinesis (kromozom / nükleer bölünme) döngüsüne maruz kalır. Ana hücre daha sonra ışığa maruz kaldıktan hemen sonra bölünür (sitokinez) ve iki hareketli hücre salar. Hareketliliğin başlangıcı ve süresi türler arasında değişir.[58] Fotoperiyodun yaklaşırken veya sonunda, mastigotlar yüzmeyi durdurur, kamçısını salar ve kokoid formuna hızlı bir başkalaşım geçirir. Kültürler durağan büyüme aşamasına ulaştıkça, daha az ve daha az hareketli hücre gözlenir, bu da daha yavaş bölünme oranlarını gösterir.
Özellikle durağan büyüme aşamasındaki hücreler taze ortama aktarıldığında bazen büyük tetradlar gözlemlenir. Bununla birlikte, bu aşamanın ardışık iki mitotik bölünmenin ürünü mü, yoksa cinsel açıdan yetkin hareketli hücreler (yani gametler) üreten bir süreç mi yoksa gamet füzyonunu takiben mayozun nihai sonucu mu olduğu bilinmemektedir. Cinsel rekombinasyon için sitolojik bir kanıt yoktur ve mayoz hiçbir zaman gözlemlenmemiştir, ancak popülasyon genetik kanıtı görüşünü desteklemektedir. Simbiyodinyum periyodik olarak cinsel rekombinasyon olaylarına maruz kalır.[37][59][60] Yaşam öykülerindeki cinsel evrenin nasıl, ne zaman ve nerede gerçekleştiği bilinmemektedir.
Morfoloji



Cinsin morfolojik tanımı Simbiyodinyum başlangıçta türlere (holotip) dayanmaktadır Symbiodinium microadriaticum.[34][61] Bu dinoflagellatlar, yaşam öykülerinde (yukarıya bakınız) iki ana aşamaya sahip oldukları için, yani mastigot (hareketli) ve kokoid (hareketsiz) aşamalar, organizmanın tam teşhisini sağlamak için her ikisinin de morfolojisi açıklanmıştır.
Flagellated (mastigot) hücre
Hareketli kamçılı form, gymnodinioid ve athekattır.[62] Epikonun ve hipokonun göreceli boyutları türler arasında farklılık gösterir.[34] Alveoller en çok hareketli fazda görülür, ancak kate ("zırhlı") dinoflagellatlarda bulunan lifli selülozik yapılardan yoksundur. İki flagella'nın başlangıç noktaları arasında pedinkül adı verilen, bilinmeyen işlevin genişletilebilir bir yapısı vardır. Diğer dinoflagellatlarda, heterotrofik beslenme ve cinsel rekombinasyonda benzer bir yapı rol oynamaktadır. İçinde Simbiyodinyum, bazı hücrelerin neden yerinde döndüğünü açıklayarak, pedinkülün substrat bağlanmasına dahil olabileceği öne sürülmüştür.[61] Diğer gymnodinioid cinslere kıyasla, sigulum oluğunun uçlarının birleştiği sulkusta çok az yer değiştirme vardır veya hiç yoktur.
Mastigotun iç organelleri, esasen kokoid hücrede tarif edilenlerle aynıdır (aşağıya bakınız). Mastigottan kokoid aşamasına geçiş Simbiyodinyum hızla oluşur, ancak hücresel değişikliklerle ilgili ayrıntılar bilinmemektedir. Mukokistler (bir çıkarma organel[63]) plazmalemmanın altında bulunan S. pilosum ve işlevleri bilinmemektedir, ancak heterotrofik beslenmeyle ilgili olabilir.
Kokoid hücresi
Kokoid hücresi Simbiyodinyum küreseldir ve türlere bağlı olarak ortalama çapı 6 ila 13 um arasında değişir (Blank ve diğerleri, 1989). Bu aşama genellikle yanlış bir şekilde bir dinokist; bu nedenle, yayınlanmış literatürde, misafirhanedeki alg genellikle vejetatif kist olarak anılır.[61] Dönem kist genellikle diğer dinoflagellatların yaşam öyküsünde, besin mevcudiyeti, sıcaklık ve gün uzunluğu gibi çeşitli faktörlerle başlatılan, uykuda, metabolik olarak durgun bir aşamayı ifade eder.[64] Bu tür kistler, olumsuz çevre koşullarına karşı uzun süreli direnç sağlar. Kokoit Simbiyodinyum hücreler, fotosentez yaptıkları, mitoza girdikleri ve proteinleri ve nükleik asitleri aktif olarak sentezledikleri için metabolik olarak aktiftir. Çoğu dinoflagellat bir mastigot olarak mitoza uğrarken, Simbiyodinyummitoz sadece kokoid hücrede meydana gelir.[58]
Hücre çeperi
Kokoid hücre, büyük moleküler ağırlıklı proteinler ve glikoproteinler içeren, genellikle yumuşak bir hücre duvarı ile çevrilidir.[34][65] Hücre duvarları kültürde olduğundan daha kalın büyür misafirperver (Şekil 11B).[7] Hücre zarı (plazmalemma) hücre duvarının altında yer alır, ancak metabolitlerin trans-membran taşınmasının düzenlenmesi açısından bileşimi ve işlevi hakkında çok az şey bilinmektedir. Karyokinezi ve sitokinez sırasında, mastigotlar ana hücreden kaçana kadar hücre duvarı bozulmadan kalır. Kültürde, atılan duvarlar kültür kabının dibinde birikir. Bölünmüş hücrelerden duvarlara ne olduğu bilinmemektedir. misafirperver.[66] Bir tür, S. pilosum, hücre duvarından saç benzeri çıkıntılar kümelerine sahiptir; bu, cins içindeki bir türü teşhis etmek için kullanılan bilinen tek yüzey özelliğidir.
Kloroplast
Tanımlanan türlerin çoğu, üç zar ile sınırlanmış tek, çevresel, ağsı bir kloroplasta sahiptir. Kloroplastın işgal ettiği hücrenin hacmi türler arasında değişir.[34] Lameller, birbirine yakın yerleştirilmiş (istiflenmiş) üç tilakoid içerir ve pirenoid'e iki sapla bağlanır.[34] nişasta kılıfı ile çevrili. Tarif edilen türlerin üçünde tilakoidler paralel diziler halindedir, ancak S. pilosum'da periferik lameller de vardır. Diğer simbiyotik dinoflagellatların aksine, pirenoidi istila eden tilakoid membranlar yoktur.[67][68] Tilakoidlerin lipid bileşenleri arasında galaktolipidler (monogalaktosil-digliseridler, digalaktosil-digliseridler), sülfolipid, (sülfokinovosil-digliserit), fosfatidil gliserol ve fosfatidil kolin bulunmaktadır. Bunlarla ilişkili çeşitli yağ asitleridir.[69] Tilakoid membrandaki ışık toplama ve reaksiyon merkezi bileşenleri, suda çözünür bir peridinin-klorofil (kl.) A-protein kompleksi (PCP) ve bir zara bağlı kl. a-chl. c2-peridinin-protein kompleksi (acpPC), fotosistem II reaksiyon merkezi ve chl gibi tipik fotosentetik elektron taşıma sistemleri ile birlikte. fotosistem I'in aP700 reaksiyon merkezi kompleksi[70][71] Ayrıca tilakoidlerle bağlantılı olan ksantofiller dinoxanthin, diadinoxanthin, diatoxanthin ve carotene, B-carotene'dir. Pirenoid, nükleer kodlu enzim tip II Ribuloz-bis-fosfat-karboksilaz-oksijenaz (Rubisco) içerir,[72] inorganik karbon dioksitin organik bileşiklere katalizinden sorumlu olan.
Kültürlenmiş tüm izolatlar (yani suşlar), hücresel Chla ve peridinin kotasının yanı sıra fotosentetik birimlerin boyutu ve sayısını değiştirerek olduğu gibi, ışık hasadı (yani fotoklimasyon) kapasitelerinde fenotipik ayarlama yapabilirler.[73] Bununla birlikte, alışma yeteneği, belirli bir fotik ortama farklı şekilde uyarlanmış (evrimleşmiş) türler arasındaki genetik farklılıkların bir yansımasıdır.[74][75] Örneğin, S. pilosum diğer türler düşük ışığa adapte edilmiş (S. kawagutii) veya değişen ışık alanlarında daha geniş bir aralığa adapte edilmişken (S. microadriaticum).

Çekirdek
Genel olarak, çekirdek merkezi olarak konumlandırılmıştır ve çekirdekçik genellikle iç çekirdek zarıyla ilişkilidir. Kromozomlar, diğer dinoflagellatlarda olduğu gibi, transmisyon elektron mikrograflarında (TEM) "kalıcı olarak süper sargılı" DNA olarak görülür.[76] Tanımlanan türleri Simbiyodinyum farklı kromozom sayılarına sahiptir (26 ile 97 arasında değişir)[34]), nükleer döngünün tüm aşamaları boyunca sabit kalan. Bununla birlikte, M-fazı sırasında, her bir kromozomun hacmi, ortaya çıkan iki çekirdeğin her birinin hacmi gibi yarı yarıya azalır. Böylece, kromozom hacminin nükleer hacme oranı sabit kalır. Bu gözlemler, alglerin haploid olduğu yorumuyla tutarlıdır ve bu, moleküler genetik verilerle desteklenen bir sonuçtur.[77] Nükleer döngünün S fazı sırasında, kromozomlar DNA sentezini kolaylaştırmak için çözülürler ve hem kromozomların hem de çekirdeğin hacimleri G2 aşamasında görülenlere geri döner.[76]
Diğer sitoplazmik organeller
Sitoplazmasında bulunan birkaç ek organel vardır. Simbiyodinyum. Bunlardan en bariz olanı, "birikim gövdesi" olarak adlandırılan yapıdır. Bu, içeriği tanınmayan ancak ışık mikroskobu altında kırmızı veya sarı görünen zara bağlı bir keseciktir (vakuol). Hücre döküntülerini biriktirmeye hizmet edebilir veya fonksiyonel olmayan organellerin sindirildiği ve bileşenlerinin geri dönüştürüldüğü bir otofajik vakuol görevi görebilir. Mitoz sırasında, sadece bir yavru hücre bu yapıyı elde ediyor gibi görünmektedir. Membranöz kapanımlar içerebilen başka vakuoller vardır,[78] yine diğerleri oksalat kristalleri veya kristal ürik asit olarak çeşitli yorumlanan kristalli malzeme içerir.
Türler

Aşağıdaki türler, Dünya Deniz Türleri Kaydı:[1]
- Symbiodinium bermudense R.K.Trench, 1993
- Simbiyodinyum kaliforniyum A.T.Banaszak, R.Iglesias-Prieto ve R.K.Trench, 1993
- Symbiodinium cariborum R.K.Trench, 1993
- Symbiodinium corculorum R.K.Trench, 1993
- Simbiyodinyum glynnii D.C.Wham, G.Ning, T.C. LaJeunesse, 2017 [79]
- Symbiodinium goreaui Hendek ve Boşluk, 2000
- Simbiyodinyum kawagutii Hendek ve Boşluk, 2000
- Simbiyodinyum meandrinae R.K.Trench, 1993
- Symbiodinium microadriaticum Freudenthal, 1962
- Simbiyodinyum minutum T.C. LaJeunesse, J.E. Parkinson ve J.D. Reimer, 2012
- Simbiyodinyum pilosum Hendek ve Boşluk, 2000
- Simbiyodinyum psygmophilum LaJeunesse, T.C., Parkinson, J.E. & Reimer, J.D., 2012
- Symbiodinium pulchrorum R.K.Trench, 1993
- Simbiyodinyum termofil, yeni türler [80]
Referanslar
- ^ a b Guiry, Michael D. (2014). "Simbiyodinyum Freudenthal, 1962 ". WoRMS. Dünya Deniz Türleri Kaydı. Alındı 2015-01-29.
- ^ Boş, Rudolf J .; Açma, Robert K. (Mayıs 1986). "Endosymbiyotik Dinoflagellatların İsimlendirilmesi". Takson. 35 (2): 286–94. doi:10.2307/1221270. JSTOR 1221270.
- ^ a b c LaJeunesse, Todd C .; Parkinson, John E .; Gabrielson, Paul W .; Jeong, Hae Jin; Reimer, James D .; Voolstra, Christian R .; Santos, Scott R. (2018). "Symbiodiniaceae'nin sistematik revizyonu, mercan endosymbiontlarının antik dönemini ve çeşitliliğini vurguluyor". Güncel Biyoloji. 28 (16): P2570–2580. doi:10.1016 / j.cub.2018.07.008. PMID 30100341.
- ^ Stimson, J .; Sakai, K .; Sembali, H. (Aralık 2002). "Mercanlarda simbiyotik ilişkinin yüksek ve düşük ağartmaya bağlı ölüm oranlarıyla karşılaştırılması". Mercan resifleri. 21 (4): 409–21. doi:10.1007 / s00338-002-0264-3.
- ^ Kawaguti, Siro (1944). "Resif mercanlarının fizyolojisi üzerine VI. Pigmentler üzerinde çalışın". Palau Tropikal Biyolojik İstasyon Çalışmaları. 2: 617–74.
- ^ McLaughlin, John J. A .; Zahl, Paul A. (1959). "Çeşitli Omurgasız Konakçılardan Axenic Zooxanthellae". New York Bilimler Akademisi Yıllıkları. 77 (2): 55–72. Bibcode:1959 NYASA..77 ... 55M. doi:10.1111 / j.1749-6632.1959.tb36892.x.
- ^ a b Colley, Nansi J .; Açma, R. K. (1983). "Fagositozda Seçicilik ve Denizanasının Scyphistoma Aşamasında Simbiyotik Alglerin Kalıcılığı Cassiopeia xamachana". Londra Kraliyet Cemiyeti Bildirileri. B serisi. 219 (1214): 61–82. Bibcode:1983RSPSB.219 ... 61C. doi:10.1098 / rspb.1983.0059. JSTOR 35678. PMID 22470960.
- ^ Wakefield, Timothy S .; Kempf, Stephen C. (2001). "Konakçıya ve ortakyaşaya özgü monoklonal antikorların geliştirilmesi ve ortakyaşar-dinoflagellat simbiyozunda simbiyozom zarının kökeninin doğrulanması". Biyolojik Bülten. 200 (2): 127–43. doi:10.2307/1543306. JSTOR 1543306. PMID 11341574.
- ^ Peng, Shao-En; Wang, Yu-Bao; Wang, Li-Hsueh; Chen, Wan-Nan Uang; Lu, Chi-Yu; Fang, Lee-Shing; Chen, Chii-Shiarng (2010). "Cnidaria-dinoflagellate endosymbiosis'te simbiyozom membranlarının proteomik analizi". Proteomik. 10 (5): 1002–16. doi:10.1002 / pmic.200900595. PMID 20049864.
- ^ Moberg, Fredrik; Folke, Carl (1999). "Mercan resif ekosistemlerinin ekolojik ürünleri ve hizmetleri". Ekolojik Ekonomi. 29 (2): 215–33. doi:10.1016 / S0921-8009 (99) 00009-9.
- ^ Jokiel, P. L .; Coles, S.L. (1990). "Hawaii ve diğer Hint-Pasifik resif mercanlarının yüksek sıcaklığa tepkisi". Mercan resifleri. 8 (4): 155–62. Bibcode:1990CorRe ... 8..155J. doi:10.1007 / BF00265006.
- ^ Küçük, Michael P (1996). "Yüksek sıcaklıklar ve ultraviyole radyasyon oksidatif strese neden olur ve simbiyotik dinoflagellatlarda fotosentezi engeller". Limnoloji ve Oşinografi. 41 (2): 271–83. Bibcode:1996LimOc..41..271L. doi:10.4319 / lo.1996.41.2.0271.
- ^ Fitt, William; Brown, Barbara; Warner, Mark; Dunne Richard (2001). "Mercan ağartma: Tropikal mercanlarda termal tolerans limitlerinin ve termal eşiklerin yorumlanması". Mercan resifleri. 20 (1): 51–65. doi:10.1007 / s003380100146.
- ^ Lajeunesse, Todd C .; Smith, Robin; Walther, Mariana; Pinzon, Jorge; Pettay, Daniel T .; McGinley, Michael; Aschaffenburg, Matthew; Medine-Rosas, Pedro; Cupul-Magana, Amilcar L .; Pérez, Andrés López; Reyes-Bonilla, Hector; Warner, Mark E. (2010). "Host-symbiont recombination versus natural selection in the response of coral-dinoflagellate symbioses to environmental disturbance". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 277 (1696): 2925–34. doi:10.1098/rspb.2010.0385. JSTOR 27862400. PMC 2982020. PMID 20444713.
- ^ Goreau, Thomas F (1964). "Mass Expulsion of Zooxanthellae from Jamaican Reef Communities after Hurricane Flora". Bilim. 145 (3630): 383–6. Bibcode:1964Sci...145..383G. doi:10.1126/science.145.3630.383. PMID 17816975.
- ^ Brown, Barbara E. (2000). "The significance of pollution in eliciting the 'bleaching' response in symbiotic cnidarians". International Journal of Environment and Pollution. 13 (1–6): 392–415. doi:10.1504/IJEP.2000.002328.
- ^ Colombo-Pallotta et al. 2010
- ^ Brandt, Marilyn E.; McManus, John W. (2009). "Disease incidence is related to bleaching extent in reef-building corals". Ekoloji. 90 (10): 2859–67. doi:10.1890/08-0445.1. JSTOR 25592820. PMID 19886494.
- ^ Baker, Andrew C .; Glynn, Peter W.; Riegl, Bernhard (2008). "Climate change and coral reef bleaching: An ecological assessment of long-term impacts, recovery trends and future outlook". Nehir ağzı. 80 (4): 435–71. Bibcode:2008ECSS...80..435B. doi:10.1016/j.ecss.2008.09.003.
- ^ Hoegh-Guldberg, O .; Mumby, P. J .; Hooten, A. J .; Steneck, R. S.; Greenfield, P .; Gomez, E .; Harvell, C. D .; Sale, P. F .; Edwards, A. J .; Caldeira, K .; Knowlton, N.; Eakin, C. M .; Iglesias-Prieto, R .; Muthiga, N .; Bradbury, R. H.; Dubi, A .; Hatziolos, M.E. (2007). "Hızlı İklim Değişikliği ve Okyanus Asitlenmesi Altındaki Mercan Resifleri". Bilim. 318 (5857): 1737–42. Bibcode:2007Sci ... 318.1737H. CiteSeerX 10.1.1.702.1733. doi:10.1126 / science.1152509. PMID 18079392.
- ^ Berkelmans, R.; Van Oppen, M. J.H (2006). "The role of zooxanthellae in the thermal tolerance of corals: A 'nugget of hope' for coral reefs in an era of climate change". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 273 (1599): 2305–12. doi:10.1098/rspb.2006.3567. PMC 1636081. PMID 16928632.
- ^ Sampayo, E. M.; Ridgway, T.; Bongaerts, P.; Hoegh-Guldberg, O. (2008). "Bleaching susceptibility and mortality of corals are determined by fine-scale differences in symbiont type". Ulusal Bilimler Akademisi Bildiriler Kitabı. 105 (30): 10444–9. Bibcode:2008PNAS..10510444S. doi:10.1073/pnas.0708049105. JSTOR 25463173. PMC 2492480. PMID 18645181.
- ^ Robison, Jennifer D.; Warner, Mark E. (2006). "Differential Impacts of Photoacclimation and Thermal Stress on the Photobiology of Four Different Phylotypes of Simbiyodinyum (Pyrrhophyta)". Journal of Phycology. 42 (3): 568–79. doi:10.1111/j.1529-8817.2006.00232.x.
- ^ Warner, Mark E.; Lajeunesse, Todd C.; Robison, Jennifer D.; Thur, Rebecca M. (2006). "The ecological distribution and comparative photobiology of symbiotic dinoflagellates from reef corals in Belize: Potential implications for coral bleaching". Limnoloji ve Oşinografi. 51 (4): 1887–97. Bibcode:2006LimOc..51.1887W. CiteSeerX 10.1.1.322.1206. doi:10.4319/lo.2006.51.4.1887.
- ^ Ragni, Maria; Airs, Ruth L.; Hennige, Sebastian J .; Suggett, David J.; Warner, Mark E.; Geider, Richard J. (2010). "PSII photoinhibition and photorepair in Simbiyodinyum (Pyrrhophyta) differs between thermally tolerant and sensitive phylotypes". Deniz Ekolojisi İlerleme Serisi. 406: 57–70. Bibcode:2010MEPS..406...57R. doi:10.3354/meps08571.
- ^ Takahashi, Shunichi; Whitney, Spencer; Itoh, Shigeru; Maruyama, Tadashi; Badger, Murray (2008). "Heat stress causes inhibition of the de novo synthesis of antenna proteins and photobleaching in cultured Simbiyodinyum". Ulusal Bilimler Akademisi Bildiriler Kitabı. 105 (11): 4203–8. Bibcode:2008PNAS..105.4203T. doi:10.1073/pnas.0708554105. JSTOR 25461395. PMC 2393757. PMID 18322010.
- ^ Lien, Yi-T.; Nakano, Y.; Plathong, S.; Fukami, H.; Wang, Jih-T.; Chen, C. A. (2007). "Occurrence of the putatively heat-tolerant Symbiodinium phylotype D in high-latitudinal outlying coral communities". Coral Reefs. 26 (1): 35–44. Bibcode:2007CorRe..26...35L. doi:10.1007/s00338-006-0185-7.
- ^ a b c Lajeunesse, Todd C.; Smith, Robin T.; Finney, Jennifer; Oxenford, Hazel (2009). "Outbreak and persistence of opportunistic symbiotic dinoflagellates during the 2005 Caribbean mass coral 'bleaching' event". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 276 (1676): 4139–48. doi:10.1098/rspb.2009.1405. JSTOR 40506039. PMC 2821356. PMID 19740874.
- ^ Lajeunesse, Todd C.; Pettay, Daniel T.; Sampayo, Eugenia M .; Phongsuwan, Niphon; Brown, Barbara; Obura, David O.; Hoegh-Guldberg, Ove; Fitt, William K. (2010). "Long-standing environmental conditions, geographic isolation and host-symbiont specificity influence the relative ecological dominance and genetic diversification of coral endosymbionts in the genus Simbiyodinyum". Biyocoğrafya Dergisi. 37 (5): 785–800. doi:10.1111/j.1365-2699.2010.02273.x.
- ^ David DeFranza (2010-02-17). "Andaman Sea Coral May Hold the Secret to Warm Water Reef Survival". Treehugger. Alındı 2015-02-02.[kendi yayınladığı kaynak? ]
- ^ "A hot survivor". Ekonomist. 9 Nisan 2016. ISSN 0013-0613. Alındı 2016-04-30.
- ^ Rowan, Rob; Powers, Dennis A. (1992). "Ribosomal RNA Sequences and the Diversity of Symbiotic Dinoflagellates (Zooxanthellae)". Amerika Birleşik Devletleri Ulusal Bilimler Akademisi Bildirileri. 89 (8): 3639–43. Bibcode:1992PNAS...89.3639R. doi:10.1073/pnas.89.8.3639. JSTOR 2359156. PMC 48924. PMID 1565660.
- ^ Stern, Rowena F.; Horak, Ales; Andrew, Rose L.; Coffroth, Mary-Alice; Andersen, Robert A.; Küpper, Frithjof C.; Jameson, Ian; Hoppenrath, Mona; Véron, Benoît; Kasai, Fumai; Brand, Jerry; James, Erick R.; Keeling, Patrick J. (2010). "Environmental Barcoding Reveals Massive Dinoflagellate Diversity in Marine Environments". PLoS ONE. 5 (11): e13991. Bibcode:2010PLoSO...513991S. doi:10.1371/journal.pone.0013991. PMC 2981561. PMID 21085582.
- ^ a b c d e f g Trench, Robert K.; Blank, Rudolf J. (1987). "Symbiodinium Microadriaticum Freudenthal, S. Goreauii Sp. Nov., S. Kawagutii Sp. Nov. And S. Pilosum Sp. Nov.: Gymnodinioid Dinoflagellate Symbionts of Marine Invertebrates". Journal of Phycology. 23 (3): 469–81. doi:10.1111/j.1529-8817.1987.tb02534.x.
- ^ a b Sampayo, E. M.; Dove, S.; Lajeunesse, T. C. (2009). "Cohesive molecular genetic data delineate species diversity in the dinoflagellate genus Simbiyodinyum". Moleküler Ekoloji. 18 (3): 500–19. doi:10.1111/j.1365-294X.2008.04037.x. PMID 19161470.
- ^ a b c Lajeunesse, Todd C.; Thornhill, Daniel J. (2011). "Improved Resolution of Reef-Coral Endosymbiont (Symbiodinium) Species Diversity, Ecology, and Evolution through psbA Non-Coding Region Genotyping". PLoS ONE. 6 (12): e29013. Bibcode:2011PLoSO...629013L. doi:10.1371/journal.pone.0029013. PMC 3247227. PMID 22216157.
- ^ a b Santos, S. R .; Shearer, T. L.; Hannes, A. R.; Coffroth, M. A. (2004). "Fine-scale diversity and specificity in the most prevalent lineage of symbiotic dinoflagellates (Symbiodinium, Dinophyceae) of the Caribbean". Moleküler Ekoloji. 13 (2): 459–69. doi:10.1046/j.1365-294X.2003.02058.x. PMID 14717900.
- ^ Finney, J. Christine; Pettay, Daniel Tye; Sampayo, Eugenia M .; Warner, Mark E.; Oxenford, Hazel A.; Lajeunesse, Todd C. (2010). "The Relative Significance of Host–Habitat, Depth, and Geography on the Ecology, Endemism, and Speciation of Coral Endosymbionts in the Genus Symbiodinium". Mikrobiyal Ekoloji. 60 (1): 250–63. doi:10.1007/s00248-010-9681-y. JSTOR 40802290. PMID 20502891.
- ^ De Queiroz, Kevin (2007). "Species Concepts and Species Delimitation". Sistematik Biyoloji. 56 (6): 879–86. doi:10.1080/10635150701701083. PMID 18027281.
- ^ Thornhill, Daniel J.; Lajeunesse, Todd C.; Santos, Scott R. (2007). "Measuring rDNA diversity in eukaryotic microbial systems: How intragenomic variation, pseudogenes, and PCR artifacts confound biodiversity estimates". Moleküler Ekoloji. 16 (24): 5326–40. doi:10.1111/j.1365-294X.2007.03576.x. PMID 17995924.
- ^ a b Pochon, X.; Garcia-Cuetos, L .; Baker, A. C.; Castella, E.; Pawlowski, J. (2007). "One-year survey of a single Micronesian reef reveals extraordinarily rich diversity of Simbiyodinyum types in soritid foraminifera" (PDF). Coral Reefs. 26 (4): 867–82. Bibcode:2007CorRe..26..867P. doi:10.1007/s00338-007-0279-x.
- ^ a b c Reimer, James Davis; Shah, Md Mahfuzur Rahman; Sinniger, Frederic; Yanagi, Kensuke; Suda, Shoichiro (2010). "Preliminary analyses of cultured Symbiodinium isolated from sand in the oceanic Ogasawara Islands, Japan". Deniz Biyoçeşitliliği. 40 (4): 237–47. doi:10.1007/s12526-010-0044-1.
- ^ Andras, Jason P.; Kirk, Nathan L.; Drew Harvell, C. (2011). "Range-wide population genetic structure of Symbiodinium associated with the Caribbean Sea fan coral, Gorgonia ventalina". Moleküler Ekoloji. 20 (12): 2525–42. doi:10.1111/j.1365-294X.2011.05115.x. PMID 21545573.
- ^ a b Pettay, Daniel T.; Wham, Drew C.; Pinzón, Jorge H.; Lajeunesse, Todd C. (2011). "Genotypic diversity and spatial-temporal distribution of Symbiodinium clones in an abundant reef coral". Moleküler Ekoloji. 20 (24): 5197–212. doi:10.1111/j.1365-294X.2011.05357.x. PMC 5957298. PMID 22082053.
- ^ a b c LaJeunesse, T. (2002). "Karayip mercan resiflerinden simbiyotik dinoflagellatların çeşitliliği ve topluluk yapısı". Deniz Biyolojisi. 141 (2): 387–400. doi:10.1007 / s00227-002-0829-2.
- ^ Rowan, Rob; Knowlton, Nancy; Baker, Andrew; Jara, Javier (1997). "Landscape ecology of algal symbionts creates variation in episodes of coral bleaching". Doğa. 388 (6639): 265–9. Bibcode:1997Natur.388..265R. doi:10.1038/40843. PMID 9230434.
- ^ Toller, W. W.; Rowan, R; Knowlton, N (2001). "Repopulation of Zooxanthellae in the Caribbean corals Montastraea annularis and M. Faveolata following experimental and disease-associated bleaching". Biyolojik Bülten. 201 (3): 360–73. doi:10.2307/1543614. JSTOR 1543614. PMID 11751248.
- ^ a b Thornhill, Daniel J.; LaJeunesse, Todd C.; Kemp, Dustin W.; Fitt, William K.; Schmidt, Gregory W. (2005). "Multi-year, seasonal genotypic surveys of coral-algal symbioses reveal prevalent stability or post-bleaching reversion". Deniz Biyolojisi. 148 (4): 711–22. doi:10.1007/s00227-005-0114-2.
- ^ Coffroth, Mary Alice; Santos, Scott R.; Goulet, Tamar L. (2001). "Early ontogenetic expression of specificity in a cnidarian-algal symbiosis" (PDF). Deniz Ekolojisi İlerleme Serisi. 222: 85–96. Bibcode:2001MEPS..222...85C. doi:10.3354/meps222085.
- ^ a b Porto, Isabel; Granados, Camila; Restrepo, Juan C.; Sánchez, Juan A. (2008). "Macroalgal-Associated Dinoflagellates Belonging to the Genus Symbiodinium in Caribbean Reefs". PLoS ONE. 3 (5): e2160. Bibcode:2008PLoSO...3.2160P. doi:10.1371/journal.pone.0002160. PMC 2364641. PMID 18478069.
- ^ Coffroth, Mary Alice; Lewis, Cynthia F.; Santos, Scott R.; Weaver, Jessica L. (2006). "Environmental populations of symbiotic dinoflagellates in the genus Symbiodinium can initiate symbioses with reef cnidarians". Güncel Biyoloji. 16 (23): R985–7. doi:10.1016/j.cub.2006.10.049. PMID 17141602.
- ^ Manning, Mackenzie M.; Gates, Ruth D. (2008). "Diversity in populations of free-living Symbiodinium from a Caribbean and Pacific reef". Limnoloji ve Oşinografi. 53 (5): 1853–61. Bibcode:2008LimOc..53.1853M. doi:10.4319/lo.2008.53.5.1853.
- ^ Schoenberg, D. A.; Trench, R. K. (1980). "Genetic Variation in Simbiyodinyum (=Gymnodinium) mikroadriatikum Freudenthal, and Specificity in its Symbiosis with Marine Invertebrates. I. Isoenzyme and Soluble Protein Patterns of Axenic Cultures of Symbiodinium microadriaticum". Londra Kraliyet Cemiyeti Bildirileri. B serisi. 207 (1169): 405–27. Bibcode:1980RSPSB.207..405S. doi:10.1098/rspb.1980.0031. JSTOR 35362.
- ^ Schoenberg, D. A.; Trench, R. K. (1980). "Genetic Variation in Simbiyodinyum (=Gymnodinium) mikroadriatikum Freudenthal, and Specificity in its Symbiosis with Marine Invertebrates. II. Morphological Variation in Symbiodinium microadriaticum". Kraliyet Topluluğu B Bildirileri: Biyolojik Bilimler. 207 (1169): 429–44. Bibcode:1980RSPSB.207..429S. doi:10.1098/rspb.1980.0032. JSTOR 35363.
- ^ Schoenberg, D. A.; Trench, R. K. (1980). "Genetic Variation in Simbiyodinyum (=Gymnodinium) mikroadriatikum Freudenthal, and Specificity in its Symbiosis with Marine Invertebrates. III. Specificity and Infectivity of Symbiodinium microadriaticum". Londra Kraliyet Cemiyeti Bildirileri. B serisi. 207 (1169): 445–60. Bibcode:1980RSPSB.207..445S. doi:10.1098/rspb.1980.0033. JSTOR 35364.
- ^ Santos, Scott R.; Taylor, Derek J.; Coffroth, Mary Alice (2001). "Genetic Comparisons of Freshly Isolated Versus Cultured Symbiotic Dinoflagellates: Implications for Extrapolating to the Intact Symbiosis". Journal of Phycology. 37 (5): 900–12. doi:10.1046/j.1529-8817.2001.00194.x.
- ^ Goulet, T.; Coffroth, M. (February 2003). "Genetic composition of zooxanthellae between and within colonies of the octocoral Plexaura Kuna, based on small subunit rDNA and multilocus DNA fingerprinting". Deniz Biyolojisi. 142 (2): 233–9. doi:10.1007/s00227-002-0936-0.
- ^ a b c Fitt, W. K.; Trench, R. K. (1983). "The Relation of Diel Patterns of Cell Division to Diel Patterns of Motility in the Symbiotic Dinoflagellate Symbiodinium Microadria Ticum Freudenthal in Culture". Yeni Fitolog. 94 (3): 421–32. doi:10.1111/j.1469-8137.1983.tb03456.x. JSTOR 2432757.
- ^ Baillie, B. K.; Belda-Baillie, C. A.; Silvestre, V.; Sison, M.; Gomez, A. V.; Gomez, E. D.; Monje, V. (2000). "Genetic variation in Symbiodinium isolates from giant clams based on random-amplified-polymorphic DNA (RAPD) patterns". Deniz Biyolojisi. 136 (5): 829–36. doi:10.1007/s002270000290.
- ^ Lajeunesse, Todd C. (2001). "Investigating the Biodiversity, Ecology, and Phylogeny of Endosymbiotic Dinoflagellates in the Genus Symbiodinium Using the Its Region: In Search of A 'species' Level Marker". Journal of Phycology. 37 (5): 866–80. doi:10.1046/j.1529-8817.2001.01031.x.
- ^ a b c Freudenthal, Hugo D. (1962). "Simbiyodinyum gen. Nov. And Symbiodinium microadriaticum sp. nov., a Zooxanthella: Taxonomy, Life Cycle, and Morphology". The Journal of Protozoology. 9 (1): 45–52. doi:10.1111/j.1550-7408.1962.tb02579.x.
- ^ Taylor, FJR (1987). "Dinoflagellate morphology". In Taylor, F. J. R. (ed.). The Biology of Dinoflagellates. Botanical Monographs, vol. 21. Oxford: Blackwell Scientific Publications. pp.24–91. ISBN 978-0-632-00915-2.
- ^ Dodge, JD; Greuet, C (1987). "Dinoflagellate ultrastructure and complex organelles". In Taylor, F. J. R. (ed.). The Biology of Dinoflagellates. Botanical Monographs, vol. 21. Oxford: Blackwell Scientific Publications. pp.92–142. ISBN 978-0-632-00915-2.
- ^ Lee, Edward Lee (2008). Psikoloji (4. baskı). New York: Cambridge University Press. ISBN 978-1-139-46987-6.[sayfa gerekli ]
- ^ Markell, DA; Trench, RK; Iglesias-Prieto, R (1992). "Macromolecules associated with the cell-walls of symbiotic dinoflagellates". Ortak yaşam. 12 (1): 19–31. INIST:5092729.
- ^ Wakefield, Timothy S.; Farmer, Mark A.; Kempf, Stephen C. (August 2000). "Revised description of the fine structure of yerinde 'zooxanthellae' genus Simbiyodinyum". Biyolojik Bülten. 199 (1): 76–84. doi:10.2307/1542709. JSTOR 1542709. PMID 10975645.
- ^ Trench, RK; Winsor, H (1987). "Symbiosis with dinoflagellates in two pelagic flatworms, Amphiscolops sp. and Haplodiscus sp". Ortak yaşam. 3 (1): 1–21. INIST:8265704.
- ^ Banaszak, Anastazia T.; Iglestas-Prieto, Roberto; Trench, Robert K. (1993). "Scrippsiella velellae sp. kas. (Peridiniales) and Gloeokinium viscum sp. kas. (Phytodiniales), Dinoflagellate Symbionts of Two Hydrozoans (Cnidiaria)". Journal of Phycology. 29 (4): 517–28. doi:10.1111/j.1529-8817.1993.tb00153.x.
- ^ Díaz-Almeyda, E.; Thomé, P. E.; El Hafidi, M.; Iglesias-Prieto, R. (2011). "Differential stability of photosynthetic membranes and fatty acid composition at elevated temperature in Symbiodinium". Coral Reefs. 30 (1): 217–25. Bibcode:2011CorRe..30..217D. doi:10.1007/s00338-010-0691-5.
- ^ Iglesias-Prieto, R .; Govind, N. S.; Trench, R. K. (1991). "Apoprotein Composition and Spectroscopic Characterization of the Water-Soluble Peridinin–Chlorophyll a–Proteins from Three Symbiotic Dinoflagellates". Bildiriler: Biyolojik Bilimler. 246 (1317): 275–83. Bibcode:1991RSPSB.246..275I. doi:10.1098/rspb.1991.0155. JSTOR 76745.
- ^ Iglesias-Prieto, R .; Govind, N. S.; Trench, R. K. (1993). "Isolation and Characterization of Three Membrane-Bound Chlorophyll-Protein Complexes from Four Dinoflagellate Species". Kraliyet Topluluğu'nun Felsefi İşlemleri B: Biyolojik Bilimler. 340 (1294): 381–92. doi:10.1098/rstb.1993.0080. JSTOR 3030171.
- ^ Rowan, Rob; Whitney, Spencer M .; Fowler, Amanda; Yellowlees, David (1996). "Rubisco in marine symbiotic dinoflagellates: Form II enzymes in eukaryotic oxygenic phototrophs encoded by a nuclear multigene family". The Plant Cell Online. 8 (3): 539–53. doi:10.1105/tpc.8.3.539. JSTOR 3870331. PMC 161119. PMID 8721755.
- ^ Hennige, S. J.; Suggett, D. J.; Warner, M. E.; McDougall, K. E.; Smith, D. J. (2009). "Photobiology of Symbiodinium revisited: Bio-physical and bio-optical signatures". Coral Reefs. 28 (1): 179–95. Bibcode:2009CorRe..28..179H. doi:10.1007/s00338-008-0444-x.
- ^ Iglesias-Prieto, Roberto; Trench, Robert K. (1994). "Acclimation and adaptation to irradiance in symbiotic dinoflagellates. I. Responses of the photosynthetic unit to changes in photon flux density". Deniz Ekolojisi İlerleme Serisi. 113 (1): 163–75. Bibcode:1994MEPS..113..163I. doi:10.3354/meps113163.
- ^ Iglesias-Prieto, R .; Trench, R. K. (1997). "Acclimation and adaptation to irradiance in symbiotic dinoflagellates. II. Response of chlorophyll-protein complexes to different photon-flux densities". Deniz Biyolojisi. 130 (1): 23–33. doi:10.1007/s002270050221.
- ^ a b Blank, Rudolf J.; Trench, Robert K. (1985). "Speciation and Symbiotic Dinoflagellates". Bilim. 229 (4714): 656–8. Bibcode:1985Sci...229..656B. doi:10.1126/science.229.4714.656. PMID 17739379.
- ^ Santos, Scott R.; Coffroth, Mary Alice (February 2003). "Molecular genetic evidence that dinoflagellates belonging to the genus Simbiyodinyum freudenthal are haploid". Biyolojik Bülten. 204 (1): 10–20. doi:10.2307/1543491. JSTOR 1543491. PMID 12588740.
- ^ Trench, R. K. (1974). "Nutritional potentials in Zoanthus sociathus (Coelenterata, Anthozoa)". Helgoländer Wissenschaftliche Meeresuntersuchungen. 26 (2): 174–216. Bibcode:1974HWM....26..174T. doi:10.1007/BF01611382.
- ^ [Drew C. Wham, Gang Ning, and Todd C. LaJeunesse (2017) Symbiodinium glynnii sp. nov., a species of stress-tolerant symbiotic dinoflagellates from pocilloporid and montiporid corals in the Pacific Ocean. Phycologia: 2017, Vol. 56, No. 4, pp. 396-409.]
- ^ Algal species helps corals survive in Earth's hottest reefs
Dış bağlantılar
- "Symbiodinium" -de Yaşam Ansiklopedisi
- An image nın-nin Simbiyodinyum -de Smithsonian Ocean Portal

| Stony corals |  | |
|---|---|---|
| Soft corals | ||
| Mercan resifleri | ||
| Coral regions |
| |
| Coral diseases | ||
| Koruma | ||
| Symbiotic algae | ||
| Diğer | ||