Niş bölme modelleri - Niche apportionment models
Niş bölüştürme için mekanik modeller, açıklamak için kullanılan biyolojik modellerdir. bağıl türlerin bolluğu dağılımlar. Bunlar niş bölme modelleri türlerin nasıl ayrıldığını tarif edin kaynak çok boyutlu uzayda havuz, bireylerin bolluğunun türler arasındaki dağılımını belirleme. Türlerin göreli bolluğu genellikle bir Whittaker grafiği veya türlerin x eksenindeki bireylerin sayısına göre sıralandığı ve y eksenindeki her türün log göreli bolluğuna karşı çizilen sıralama bolluğu grafiği olarak ifade edilir. Nispi bolluk, türler içindeki göreceli birey sayısı veya türler içindeki bireylerin göreceli biyokütlesi olarak ölçülebilir.
Tarih
Niş bölüştürme modelleri geliştirildi çünkü ekolojistler biyolojik açıklamalar için bağıl türlerin bolluğu dağılımlar. MacArthur (1957, 1961),[1][2] tamamen ile ilgili memnuniyetsizliği en erken ifade edenlerden biriydi istatistiksel modeller yerine 3 mekanik niş paylaştırma modeli sunuyor. MacArthur buna inanıyordu Ekolojik nişler bir kaynak havuzunun içinde bir çubuk gibi parçalanabilir ve çubuğun her bir parçası toplulukta bulunan nişleri temsil eder. Sugihara'nın (1980) katkılarıyla,[3] Tokeshi (1990, 1993, 1996)[4][5][6] kabaca 7 mekanik niş dağıtma modeli ürettiğinde, kırık çubuk modelini genişletti. Bu mekanik modeller, toplulukların tür bileşimini tanımlamak için yararlı bir başlangıç noktası sağlar.
Açıklama

Bir kaynak havuzunun, türleri kolonileştirerek veya türleme yoluyla sıralı olarak veya eş zamanlı olarak daha küçük nişlere bölündüğü durumlarda (kaynak kullanımına ilişkin açıklama: bir loncadaki türler aynı kaynakları kullanırken, bir topluluk içindeki türler bunu kullanamayabilir) ).
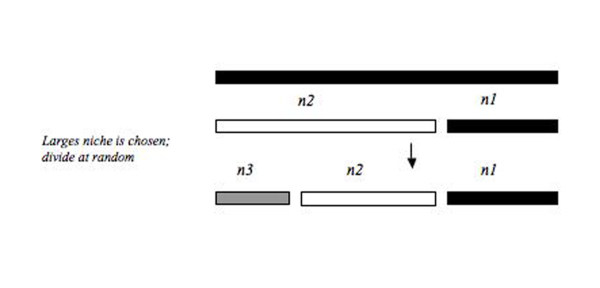
Bu modeller, aynı kaynak havuzundan (ör. lonca (ekoloji) ) nişlerini böl. Kaynak havuzu, sırayla veya eşzamanlı olarak kırılır ve nişin parçalanma sürecinin iki bileşeni, hangi parçanın seçildiğini ve elde edilen parçanın boyutunu içerir (Şekil 2).
Birincil literatürde, tatlı su böcekleri, balıklar, briyofit böcekleri, hymenopteran parazitleri, plankton toplulukları ve tuzlu bataklık otları dahil olmak üzere çeşitli taksonların nispi bolluk dağılımlarındaki değişiklikleri açıklamak ve tanımlamak için niş bölme modelleri kullanılmıştır.
Varsayımlar

Bu arazileri tanımlayan mekanik modeller, sıralama bolluğu grafiklerinin türler içindeki bireylerin bolluğunun titiz bir tahminine dayandığı ve bu önlemlerin gerçek tür bolluğu dağılımını temsil ettiği varsayımı altında çalışır. Dahası, ister bolluk ölçüsü olarak birey sayısını ister bireylerin biyokütlesini kullanın, bu modeller bu miktarın bir organizmanın kapladığı nişin büyüklüğü ile doğru orantılı olduğunu varsayar. Bir öneri, bireylerin sayısı olarak ölçülen bolluğun, kullananlara göre daha düşük varyanslar gösterebileceğidir. biyokütle. Bu nedenle, bolluğu niş tahsisi için bir vekil olarak kullanan bazı çalışmalar, eşitlik bir topluluk. Bunun nedeni, vücut büyüklüğü arasındaki ilişkinin net bir ayrımının olmaması, bolluk (ekoloji), ve kaynak kullanın. Çoğunlukla çalışmalar, büyüklük yapısını veya biyokütle tahminlerini gerçek bolluk ölçümlerine dahil etmekte başarısız olur ve bu ölçüm, niş dağıtma modellerinde, kesinlikle birey sayısı olarak ölçülen bolluktan daha yüksek bir varyans yaratabilir.[7][8]
Tokeshi'nin niş bölüştürme mekanistik modelleri
Niş paylaştırmayı tanımlayan yedi mekanik model aşağıda açıklanmaktadır. Modeller, Baskınlık Önleme modelinden, Baskınlık Azalması ve MacArthur Kesir modellerine en azından eşitlik artırma sırasına göre sunulur.
Hakimiyet önleme
Bu model, ilk kolonizasyondan (veya türleşmeden) sonra her yeni türün kalan en küçük nişin% 50'sinden fazlasını önceden boşalttığı bir durumu tanımlar. Bir Dominance preemption modelinde niş dağıtma modelinde türler, kalan en küçük nişin% 50 ila% 100'ü arasındaki rastgele bölümü kolonileştirerek bu modeli doğası gereği stokastik yapar. Yakından ilişkili bir model olan Geometrik Seri,[9] Baskınlık ön empresyon modelinin deterministik bir versiyonudur, burada yeni türün kapladığı kalan niş alanı yüzdesi (k) her zaman aynıdır. Aslında, baskınlık ön-empresyonu ve geometrik seri modelleri kavramsal olarak benzerdir ve doldurulan daha küçük nişin oranı her zaman 0,75 olduğunda aynı göreceli bolluk dağılımını üretecektir. Baskınlık ön empoze modeli, bazı akarsu balığı topluluklarının göreli bolluk dağılımlarına en uygun olanıdır. Teksas, bazı taksonomik gruplamalar ve belirli işlevsel gruplamalar dahil.[10]
Geometrik (k = 0.75)


Rastgele çeşitler
Rastgele çeşitlilik modelinde kaynak havuzu, eşzamanlı veya sıralı olarak kolonileşen türler arasında rastgele bölünür. Bu model, bolluk ölçüsü bir türün işgal ettiği niş miktarına göre ölçeklenmediğinden veya türlerin bolluğundaki veya niş genişliğindeki zamansal varyasyonun zaman içinde niş dağılımında süreksizliğe neden olduğu ve bu nedenle türlerin doluluk kapsamı arasında hiçbir ilişkiye sahip olmadığı için ortaya çıkabilir. ve nişleri. Tokeshi (1993)[5] bu modelin birçok yönden Caswell'in tarafsız biyoçeşitlilik teorisine benzediğini, çünkü türlerin birbirinden bağımsız hareket ediyor gibi göründüğünü açıkladı.
Rastgele kesir
Rastgele fraksiyon modeli, niş büyüklüğünün, türleri sıralı olarak kolonize ederek rastgele seçildiği bir süreci açıklar. İlk tür, toplam nişin rastgele bir bölümünü seçer ve sonraki kolonileşen türler de toplam nişin rastgele bir bölümünü seçer ve tüm türler kolonileşene kadar rastgele böler. Tokeshi (1990)[4] bu modeli bazı epifitik Chiromonid karides toplulukları ile uyumlu bulmuş ve son zamanlarda fitoplankton topluluklarının göreceli bolluk dağılımlarını, tuzlu çayır bitki örtüsünü, Diptera düzenindeki bazı böcek topluluklarını, bazı yer böceği topluluklarını, işlevsel ve işlevselliği açıklamak için kullanılmıştır. Teksas biyo-bölgelerindeki akarsu balıklarının taksonomik gruplamaları ve iknömonid parazitoitler. Benzer bir model, Preston'ın (1948) log normal dağılımı için biyolojik bir açıklama sağlamak amacıyla Sugihara tarafından geliştirilmiştir.[11] Sugihara'nın (1980)[3] Sabit Bölme Modeli, rastgele kesir modeline benzerdi, ancak modelin rasgeleliği bir üçgen dağılım rasgele kesirde ortalama 0,5 olan normal dağılımdan ziyade ortalama 0,75 ile. Sugihara, rastgele değişkenleri çizmek için üçgen bir dağılım kullandı çünkü bazı doğal popülasyonların rasgeleliği, ortalama 0.75 olan üçgen bir dağılımla eşleşti.
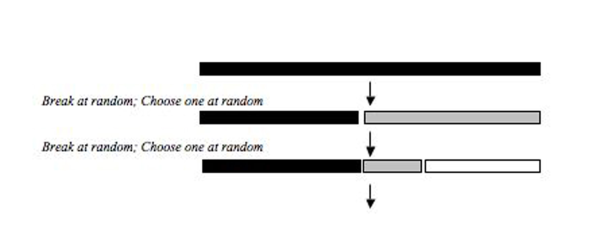
Güç oranı
Bu model, bir kaynak havuzundaki mevcut bir nişin kolonizasyon olasılığı o nişin boyutuyla pozitif olarak ilişkili olduğunda (bolluk, biyokütle vb. Ölçülür) göreceli bir bolluk dağılımını açıklayabilir. Nişin bir kısmının kolonileşme olasılığı, oluşturulan nişlerin göreli boyutlarına bağlıdır ve bir k üssü ile ölçeklenir. k, 0 ile 1 arasında bir değer alabilir ve k> 0 ise, her zaman biraz daha yüksek bir olasılıkla daha büyük niş kolonileştirilir. Bu model biyolojik olarak daha gerçekçi olarak düşünülmüştür çünkü daha fazla kaynak oranına sahip nişin işgal edilme olasılığının daha yüksek olduğu, çünkü bu nişin daha fazla kaynak alanına ve dolayısıyla daha fazla edinme fırsatına sahip olduğu birçok durum hayal edilebilir. Niş bölüşümünün rastgele kesir modeli, k = 0 olduğu güç kesir modelinin bir aşırısıdır ve k = 1 olduğunda güç kesirinin diğer ucu, kolonizasyon olasılığının niş boyutuyla doğru orantılı olduğu MacArthur Kesir modeline benzer.[6][12]

MacArthur kesri
Bu model, ilk nişin rastgele kırılmasını ve ardışık nişlerin büyüklükleriyle orantılı bir olasılıkla seçilmesini gerektirir. Bu modelde, en büyük niş, kaynak havuzundaki daha küçük nişlere göre her zaman daha büyük bir kırılma olasılığına sahiptir. Bu model, daha büyük nişlerin kırılma olasılığının daha yüksek olduğu ve eşdeğer büyüklükteki nişlerdeki türler arasında bir arada yaşamayı kolaylaştıran daha eşit bir dağılıma yol açabilir. MacArthur Kesir modelinin temeli, MacArthur (1957) tarafından geliştirilen Kırık Çubuk'tur. Bu modeller benzer sonuçlar üretir, ancak temel kavramsal farklılıklardan biri, MacArthur Fraksiyonunda olduğu gibi sıralı olarak değil, Kırık Çubuk modelinde nişlerin aynı anda doldurulmasıdır. Tokeshi (1993)[5] Bir kaynak havuzunu sıralı olarak istila etmenin aynı anda niş alanını kırmaktan daha biyolojik olarak gerçekçi olduğunu savunuyor. Teksas'taki tüm biyo-bölgelerden gelen balıkların bolluğu birleştirildiğinde, dağılım, Teksas'taki tatlı su balık türlerinin nispeten eşit dağılımını düşündüren, niş dağılımının kırık çubuk modeline benziyordu.[10]

Hakimiyet çürümesi
Bu model, Baskınlık ön empresyon modelinin tersi olarak düşünülebilir. İlk olarak, ilk kaynak havuzu rastgele kolonize edilir ve geri kalan, sonraki koloniler, kolonize edilmiş olsun ya da olmasın her zaman en büyük nişi kolonize eder. Bu model, yukarıda açıklanan niş paylaştırma modellerine göre en eşit topluluğu oluşturur, çünkü en büyük niş her zaman kırılmamış daha küçük nişin boyutuna eşdeğer olma olasılığı daha yüksek olan iki küçük parçaya bölünür. Bu "düzlük" düzeyindeki toplulukların doğal sistemlerde nadir olduğu görülmektedir. Bununla birlikte, böyle bir topluluk, Avusturya'daki Tuna Nehri içindeki bir sahadaki filtre besleyicilerin göreceli bolluk dağılımını içermektedir.[13]
Bileşik
Kaynak havuzunun farklı bölümlerinde niş dağıtma modellerinin bir kombinasyonu hareket ettiğinde bir bileşik model mevcuttur. Fesl (2002).[13] tatlı su çalışmasında bir kompozit modelin nasıl görünebileceğini gösterir Diptera, farklı niş dağıtma modellerinin verilerin farklı işlevsel gruplarına uyması bakımından. Higgins ve Strauss'un (2008) balık topluluklarını modelleyen bir başka örneği, farklı habitatlardan ve farklı tür bileşimlerine sahip balık topluluklarının farklı niş dağıtma modellerine uyduğunu, bu nedenle tüm tür topluluğunun tür aralığının farklı bölgelerindeki modellerin bir kombinasyonu olduğunu bulmuştur. .
Niş dağılımının mekanik modellerini ampirik verilere uydurma
Mekanistik niş paylaştırma modelleri toplulukları tanımlamayı amaçlamaktadır. Araştırmacılar, türlerin bolluğundaki zamansal ve coğrafi eğilimleri araştırmak için bu modelleri birçok şekilde kullandılar.
Uzun yıllar niş paylaştırma modellerinin uyumu gözle yapılmış ve modellerin grafikleri deneysel verilerle karşılaştırılmıştır.[5] Daha yakın zamanlarda, niş paylaştırma modellerinin ampirik verilere uygunluğunun istatistiksel testleri geliştirilmiştir.[14][15] Daha sonraki yöntem (Etienne ve Ollf 2005)[15] Ampirik verilere uyumlarını test etmek için modellerin Bayes simülasyonunu kullanır. Halen yaygın olarak kullanılan eski yöntem, ampirik verilerle aynı sayıda türe verilen her modelin normal dağılımından beklenen göreli bolluklarını simüle eder. Her model birden çok kez simüle edilir ve ortalama ve standart sapma, her bir göreli bolluk dağılımı etrafında güven aralıkları atamak için hesaplanabilir. Model uyumunu belirlemek için her bir seviye etrafındaki güven, her model için deneysel verilere karşı test edilebilir. Güven aralıkları aşağıdaki şekilde hesaplanır.[4][12] Niş paylaştırma modellerinin simülasyonu hakkında daha fazla bilgi için web sitesi [1][kalıcı ölü bağlantı ], Power Niche programını açıklıyor.[14]

r = simüle edilen verilerin güven limiti σ = simüle edilen veranın standart sapması = ampirik numunenin tekrarlarının sayısı
Referanslar
- ^ MacArthur, R.H. (1957). Kuş türlerinin göreceli bolluğu üzerine. Proc. Natl. Acad. Sci. 43, 293-295.
- ^ MacArthur, R.H. MacArthur, J.W. (1961). Kuş türleri çeşitliliği üzerine. Ekoloji. 42, 594-598.
- ^ a b Sugihara, G. (1980). Minimal topluluk yapısı: türlerin bolluk modellerinin açıklaması. Am. Nat. 116. 770-787.
- ^ a b c Tokeshi, M. (1990). Niş bölüştürme veya rastgele çeşitlilik: türlerin bolluğu kalıpları yeniden ziyaret edildi. Hayvan Ekolojisi Dergisi. 59, 1129-1146.
- ^ a b c d Tokeshi, M. (1993). Tür bolluğu kalıpları ve topluluk yapısı. Adv. Ecol. Res. 24, 112-186.
- ^ a b Tokeshi, M. (1996). Güç Fraksiyonu: Tür açısından zengin topluluklardaki tür bolluğu modellerine yeni bir açıklama. Oikos. 75, 543-550.
- ^ Gaston, K.W.Blackburn, T.M. (2000). Macroecology, Oxford, UK: Blackwell Science.
- ^ Taper, M.L. Marquet, P.A. (1996). Türler kaynakları gerçekten nasıl böler? Amerikan Doğabilimci. 147, 1072-1086.
- ^ Mayıs, R.M. (1975). Türlerin bolluğu ve çeşitliliği kalıpları. Ekoloji ve Toplulukların Evrimi'nde. 81-120, Cambridge, MA: Harvard University Press.
- ^ a b Higgins, C.L. Strauss, R. E. (2008). Akarsu balığı topluluklarının niş dağıtma modelleriyle modellenmesi: desenler. süreçler ve ölçek bağımlılığı. Amerikan Balıkçılık Derneği'nin İşlemleri. 137, 696-706.
- ^ Prestion, F.W. (1948). Türlerin yaygınlığı ve enderliği. Ekoloji. 29, 254-253.
- ^ a b Magurrran, A. E. (2004). Biyolojik Çeşitliliğin Ölçülmesi. Oxford, İngiltere: Blackwell Science
- ^ a b Fesl, C. (2002). Niş yönelimli türlerin bolluk modelleri: farklı yaklaşımlar ve bunların büyük bir nehirdeki larva chironomid (Diptera) topluluklarına uygulamaları. J. Anim. Ecol. 71, 1085-1094.
- ^ a b Drozd, P. Novotny, V. (2000). Güç Niş: Topluluk analizi için Niş Bölme modelleri. Sürüm 1. Kılavuz ve program www.entu.cas / png / PowerNiche adresinde yayınlanmıştır.
- ^ a b Etienne, R.S. Ollf, H. (2005). Tür bolluğu verileriyle farklı topluluk yapısı modelleriyle yüzleşmek: Bayes modeli karşılaştırması. Ekoloji Mektupları. 8, 493-504.