Gallimimus - Gallimimus
| Gallimimus | |
|---|---|
 | |
| Yeniden yapılandırılmış iskelet (yetişkinlere göre holotip ve bir çocuk örneği), Doğal Tarih Müzesi, Londra | |
| bilimsel sınıflandırma | |
| Krallık: | Animalia |
| Şube: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | †Ornitomimosauria |
| Aile: | †Ornithomimidae |
| Cins: | †Gallimimus Osmólska et al., 1972 |
| Türler: | †G. bullatus |
| Binom adı | |
| †Gallimimus bullatus Osmólska et al., 1972 | |
| Eş anlamlı | |
| |
Gallimimus (/ˌɡælɪˈmaɪməs/ GAL-ben-BENİM-məs ) bir cins nın-nin Theropod Dinozor şimdi ne yaşadı Moğolistan esnasında Geç Kretase dönem, yaklaşık yetmiş milyon yıl önce (mya). Polonya-Moğol keşif gezileri tarafından çeşitli büyüme aşamalarında bulunan birkaç fosil keşfedildi. Gobi Çölü 1960'larda Moğolistan; bu bölgede keşfedilen büyük bir iskelet, holotip yeni cins ve türlerin örneği Gallimimus bullatus 1972'de. Genel ad "tavuk taklit ", boyun omurları ile boyun omurları arasındaki benzerliklere atıfta bulunur. Galliformlar. Spesifik ad türetilmiştir mühür baskısı giydiği altın bir kapsül Roma gençlik, kafatasının tabanındaki soğanlı bir yapıya referansla Gallimimus. Adını verildiği sırada fosilleri Gallimimus en eksiksiz ve en iyi korunmuş olanı temsil etti ornitomimid ("devekuşu dinozor ") materyali keşfedildi ve cins, grubun en bilinen üyelerinden biri olmaya devam ediyor.
Gallimimus bilinen en büyüğü ornitomimid; yetişkinler kalçada yaklaşık 6 metre (20 ft) uzunluğunda, 1.9 metre (6 ft 3 inç) uzunluğundaydı ve yaklaşık 440 kilogram (970 lb) ağırlığındaydı. Akrabası tarafından kanıtlandığı gibi Ornitomimus, olurdu tüyleri vardı. Baş, yanlara bakan iri gözlerle küçük ve hafifti. Burun, diğer ornitomimidlere kıyasla uzundu, ancak diğer türlere göre daha geniş ve uçta daha yuvarlaktı. Gallimimus ile dişsizdi keratinli (azgın) gaga ve narin bir alt çenesi vardı. Birçok omur olduğunu gösteren açıklıklar vardı pnömatik (hava dolu). Boyun gövdeye göre orantılı olarak uzundu. Eller oransal olarak herhangi bir ornitomimozorun en kısasıydı ve her birinin kavisli pençeli üç rakamı vardı. Arka ayaklar orantılı olarak uzun iken ön ayaklar zayıftı. Ornithomimidae ailesi grubun bir parçasıdır Ornitomimosauria. Anserimimus Moğolistan'dan da en yakın akrabası olduğu düşünülüyor. Gallimimus.
Ornitomimid olarak, Gallimimus bir filo olabilirdi (veya cursorial ) avcılardan kaçmak için hızını kullanan hayvan; hızının 42–56 km / saat (29–34 mil / saat) olduğu tahmin edilmektedir. Şununla karşılaştırılabilir iyi bir vizyon ve zekaya sahip olabilir ratit kuşlar. Gallimimus bir grupta korunmuş birkaç örneğin keşfine dayanarak gruplar halinde yaşamış olabilir. kemik yatağı. Diyetle ilgili çeşitli teoriler önerilmiştir. Gallimimus ve diğer ornitomimidler. Oldukça hareketli olan boyun, küçük bir avın yerdeki yerini tespit etmeye yardımcı olmuş olabilir, ancak aynı zamanda fırsatçı da olabilir. Hepçil. Ayrıca gagasında küçük sütunlu yapılar kullandığı öne sürülmüştür. filtre besleme su içinde, ancak bu yapılar bunun yerine sert bitki materyalini beslemek için kullanılan sırtlar olabilir; otçul diyet. Gallimimus içinde en yaygın bulunan ornitomimozordur Nemegt Oluşumu akrabalarıyla birlikte yaşadığı yer Anserimimus ve Deinocheirus. Gallimimus filmde yer aldı Jurassic Park tarihi için önemli olan bir sahnede özel efektler ve dinozorların kuş benzeri hayvanlar olarak genel anlayışını şekillendirmede.
Keşif tarihi

1963 ile 1965 arasında Polonya Bilimler Akademisi ve Moğol Bilimler Akademisi Polonya-Moğol paleontolojik seferlerini düzenledi. Gobi Çölü Moğolistan. Kum yataklarında keşfedilen dinozor kalıntıları arasında Nemegt Havzası sayısızdı ornitomimidler Nemegt, Tsaagan Khushuu, Altan Ula IV ve Naran Bulak yörelerinden farklı büyüme aşamalarında. İkisi kafataslı, kısmen tamamlanmış üç iskelet ve birçok parçalı kalıntı toplandı. En büyük iskelet (daha sonra holotip olacak Gallimimus bullatus) paleontolog tarafından keşfedildi Zofia Kielan-Jaworowska 1964'te Tsaagan Khushuu'da; sırt üstü yatarak korunmuştur ve kafatası pelvisinin altında bulunmuştur. Aynı yıl Tsaagan Khushuu'da da küçük bir örnek bulundu ve Nemegt yöresinde küçük bir örnek daha bulundu. Nemegt Havzası dışındaki Bugeen Tsav'da Moğol paleontoloji keşif gezisi tarafından 1967'de ön ayakları olmayan küçük bir iskelet keşfedildi. Fosiller, Moğol, Polonya ve SSCB Bilimler Akademisi.[1][2][3] Polonya-Moğol keşif gezileri, bazıları yeni dinozorlara isim veren ilk kadınlar arasında yer alan kadınlar tarafından yönetilmesiyle dikkat çekiyordu. Bu keşif gezilerinde bulunan fosiller, Asya ve Kuzey Amerika arasındaki fauna değişimine yeni ışık tuttu. Kretase dönem.[4][5][6] İskeletlerin bir kısmı 1968'de Varşova'da sergilendi, yüksek, yarı dik duruşlarda monte edildi ve o zamanlar kabul edildi, ancak bugün daha yatay duruşlar tercih ediliyor.[6]

1972'de paleontologlar Halszka Osmólska, Ewa Roniewicz ve Rinchen Barsbold yeni isimlendirildi cins ve türler Gallimimus bullatus, toplanan en büyük iskelet olan IGM 100/11 örneğini (daha önce G.I.No.DPS 100/11 ve MPD 100/11 olarak anılan Tsaagan Khushuu'dan) kullanarak, holotip. Genel ad, Latince safra, "tavuk" ve Yunanca Mimolar, "mimik", boyun omurlarının ön kısmına atıfta bulunarak Galliformlar. belirli isim Latince'den türetilmiştir mühür baskısı giydiği altın bir kapsül Roma dinozorun kafatasının tabanındaki parasfenoid üzerinde bulunan soğanlı kapsüle referansla boyundaki gençlik. Böyle bir özellik, o zamanlar diğer sürüngenlerden tarif edilmemişti ve olağandışı kabul edildi. Holotip, burnu bozuk, tam olmayan alt çene, omurga serisi, pelvis ve bazı eksik el ve ayak kemiklerine sahip neredeyse eksiksiz bir iskeletten oluşur.[1][7] Diğer kısmen tamamlanmış iskeletler gençti; ZPAL MgD-I / 1 (Tsaagan Khushuu'dan) eksik ucu, hasarlı omurları, parçalanmış kaburgaları, pektoral kuşağı ve ön ayakları olan ezilmiş bir kafatasına ve tamamlanmamış bir sol arka uzuv, ZPAL MgD-I / 94'e (Nemegt bölgesinden) sahiptir. kafatası, atlas, kuyruğun ucu, pektoral kuşak ve ön ayaklardan yoksundur, en küçük örnek olan IGM 100/10 (Bugeen Tsav'dan) pektoral kuşak, ön ayaklar ve birkaç omur ve kaburgadan yoksundur. Osmólska ve meslektaşları, dokuzu yalnızca tek kemiklerle temsil edilen yirmi beş bilinen örneği listeledi.[1][8]

Adını verildiği sırada fosilleri Gallimimus Henüz keşfedilmiş en eksiksiz ve en iyi korunmuş ornitomimid materyali temsil ediyordu ve cins, grubun en iyi bilinen üyelerinden biri olmaya devam ediyor. Ornitomimidler daha önce esas olarak Kuzey Amerika'dan biliniyordu. Archaeornithomimus Asya'dan bilinen tek üye (kafatası olmasa da). İlk keşiflerden bu yana, Moğol liderliğindeki uluslararası keşif gezilerinde daha fazla örnek bulundu.[1][8][5][6] Üçü Gallimimus iskeletler (holotip dahil) daha sonra çeşitli ülkeleri gezen Moğol dinozor fosillerinin gezici sergisinin bir parçası haline geldi.[9][10] Fosil kaçakçılığı 21. yüzyılda Moğolistan'da ciddi bir sorun haline geldi ve Gallimimus örnekler yağmalandı. 2017'de Hang-Jae Lee ve meslektaşları, 2009'da keşfedilen bir fosil yolunun sıkılmış bir Gallimimus ayak (örnek MPC-D100F / 17). İskeletin geri kalanı daha önce kaçak avcılar tarafından ve diğer birkaç kişi tarafından çıkarılmış gibi görünüyordu. Gallimimus örnekler (boş kazı çukurları, çöpler ve taş ocağındaki dağınık kırık kemiklerle gösterildiği gibi). Vücut fosilleriyle yakından ilişkili izler bulmak alışılmadık bir durumdur; izlerin bazıları ornitoimid ayaklarla uyumluyken diğerleri farklı dinozorlara aittir.[11][12] 2014 yılında, iki Gallimimus ABD'ye kaçırıldıktan sonra örnekler diğer dinozor iskeletleriyle birlikte Moğolistan'a geri gönderildi.[13]
1988'de paleontolog Gregory S. Paul Ornitomimidlerin kafataslarının önceden düşünüldüğünden daha benzer olduğu ve çoğu türü aynı cinse taşıdığı sonucuna vardı, Ornitomimus, sonuçta yeni kombinasyon O. bullatus.[14] 2010'da bunun yerine "Gallimimus (veya Struthiomimus) bullatus", ancak yalnızca cins adını kullanmaya geri döndü Gallimimus 2016 yılında.[15][16] İlgili türler genellikle diğer yazarlar tarafından ayrı cinslerde tutulmuştur.[17] Japonya'dan bir ornitoimid omur gayri resmi olarak adlandırılmış "Sanchusaurus" 1988 tarihli bir dergide yayınlandı, ancak Gallimimus sp. (belirsiz türlerin) paleontolog tarafından Dong Zhiming ve meslektaşları 1990'da.[7] Barsbold gayri resmi olarak neredeyse eksiksiz bir iskelete (IGM 100/14) "Gallimimus mongoliensis" olarak atıfta bulundu, ancak Gallimimus Bazı detaylarda, Yoshitsugu Kobayashi ve Barsbold, 2006 yılında muhtemelen farklı bir cinse ait olduğunu öne sürmüşlerdir. 2000 yılında paleontolog Philip J. Currie bunu önerdi Anserimimus Moğolistan'dan sadece bir iskeletten bilinen bir küçük eşanlamlı nın-nin Gallimimus, ancak bu, ikisi arasındaki birkaç farklılığa işaret eden Kobayashi ve Barsbold tarafından reddedildi. Barsbold bazılarını kaydetti morfolojik yeniler arasında varyasyon Gallimimus örnekler, ancak bu asla yayınlanmadı.[8]
Açıklama
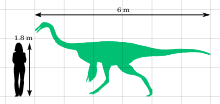
Gallimimus bilinen en büyük üyesidir aile Ornithomimidae. Yetişkin holotipi (örnek IGM 100/11) kalçada yaklaşık 6 metre (20 ft) uzunluğunda ve 1.9 metre (6.2 ft) uzunluğundaydı; kafatası 330 milimetre (1.08 ft) uzunluğundaydı ve uyluk (uyluk kemiği) 660 milimetre (2.17 ft) idi. Yaklaşık 440 kilogram (970 lb) ağırlığındaydı. Karşılaştırıldığında, bir genç örnek (ZPAL MgD-I / 94) yaklaşık 2.15 metre (7.1 ft) uzunluğunda, kalçada 0.79 metre (2.6 ft) uzunluğundaydı ve yaklaşık 26 kilogram (57 lb) ağırlığındaydı.[14] İlgili fosillere dayanarak Ornitomimus biliniyor ki ornitomimozorlar -di tüylü ve yetişkinlerin kanat benzeri yapılar taşıdığını, bunun da kanıtlandığı gibi, üzerinde tüy uçlarının bulunmasıyla kanıtlanmıştır. ulna alt kolun kemiği, tüylerin nereye yapışacağını gösteren tümsekler.[18]
Kafatası

Başı Gallimimus omurgaya göre çok küçük ve hafifti. Burnunun uzunluğu nedeniyle, kafatası diğer ornitomimidlere kıyasla uzundu ve burnun hafifçe dışbükey eğimli bir üst profili vardı. Burnun yan profili, diğer ornitomimidlerden, ön yarısına ve alt ön kenarına doğru daralmaması bakımından farklıydı. premaksilla üst çenenin önünde yatay olmak yerine yukarı doğru yükseldi. Yukarıdan bakıldığında, burun neredeyse spatulaydı (kaşık şeklinde), uçta geniş ve yuvarlaktı (veya U şeklinde ), Kuzey Amerika ornitomimidlerinde akut (veya V şeklinde) idi. yörüngeler (göz yuvaları) büyüktü ve diğer ornitomimidlerde olduğu gibi yanlara dönüktü. zamansal bölge kafatasının yanında gözlerin arkasında derin ve zamansız fenestra (yörüngenin arkasındaki alt açıklık) neredeyse üçgen şeklindeydi ve ilişkili olandan daha küçüktü. Struthiomimus. Kafatasının çatısının arka kısmında derin kas izleri vardı. paryetal kemik. parasfenoid (kafatasının tabanının altındaki beyin kasasının kemiği) ince duvarlı, oyuk ve armut biçimli, soğanlı bir yapı oluşturuyordu. Yapı, öne doğru açılan sığ bir oluğa sahipti. iç burun (burun geçişi için iç açıklıklar) büyüktü ve sırtın çok gerisine yerleştirilmişti. damak kapsamlı bir ikincil damak ornitomimidlerde ortak olan.[1][17][19][20]
İnce kemiklerden oluşan hassas alt çene, önde ince ve sığdı, arkaya doğru derinleşiyordu. Mandibulanın ön tarafı kürek şeklindeydi ve kapatıldığında çenelerin uçları arasında bir boşluk oluşmasına neden oluyordu. Kürek benzeri şekli, kürek şeklininkine benziyordu. ortak martı ve alt gaga bu kuşunkine benzer bir şekle sahip olabilir. Çenenin arkasındaki retroartiküler süreç (gagayı açan çene kaslarının bağlandığı yer) iyi gelişmiştir ve esas olarak açısal kemik. yuvarlak theropodlarda olağan olan alt çenenin en büyük kemiğiydi. mandibular fenestra alt çenede yana bakan bir açıklık uzamış ve nispeten küçüktür. Alt çenede bir koronoid süreç veya bir supradentary kemik, eksikliği gagalı theropodların (ornitomimozorlar, Oviraptorozorlar, Therizinosaurlar ve kuşlar), ancak genel olarak theropodlar arasında sıra dışıdır.[1][21] Çeneleri Gallimimus -di dişsiz (dişsiz) ve ön kısım bir keratinli hayatta rhamphotheca (azgın gaga). Gaga, maksillada besleyici foramina bulunmamasına bağlı olarak Kuzey Amerika'daki akrabalardan daha küçük bir alanı kaplamış olabilir. Gaganın iç tarafı, önde en uzun olan ve arkaya doğru kısalan küçük, sıkıca paketlenmiş ve eşit aralıklı sütunlu yapılara (kesin doğası tartışılmaktadır) sahipti.[19][21][22]
Postkraniyal iskelet

Gallimimus 64–66 vardı omur onun içinde omurga, diğer ornitomimidlerden daha az. Omurların merkezi (veya gövdeleri), ilk altı tanesi hariç, düz bir ön yüzeye ve içbükey bir arka yüzeye sahip, platikelözdü. kuyruk (kuyruk) omur - arka yüzeyin de düz olduğu - ve kuyruğun sonundakiler - her iki yüzeyi de düz olan amfiplatyan. Centra'nın çoğu vardı foramina ("açıklıklar" olarak da adlandırılır)pleurocoels ") ve bu nedenle muhtemelen pnömatik (oyuk odaları tarafından işgal edilmiş hava keseciklerinin ). Boyun 10'dan oluşuyordu boyun omurları, hepsi uzun ve genişti, hariç atlas kemiği (kafatasının arkasına bağlanan ilk omur). Atlas, diğer ornitomimidlerden farklıydı, çünkü intercentrumunun ön yüzeyi, içbükey olmak ve yukarı bakacak şekilde arkaya doğru eğimliydi. oksipital kondil. Boyun diğer ornitomimidlere göre gövdeye göre orantılı olarak daha uzun görünmektedir. Boyun iki ayrı bölüme ayrıldı: öndeki servikal omurlar, yan görünümde neredeyse üçgen olan ve arkaya doğru sivrilen bir merkeze sahipti, ayrıca düşük sinir kemerleri ve kısa, geniş zigapofizler ( süreçler omurlar arasında eklemlenen); arkadaki servikal omurlar, giderek yükselen makara benzeri merkezlere ve uzun, ince zigapofizlere sahipti. Buradaki pnömatik foramina küçük ve ovaldi ve merkezden dışarıya doğru çıkıntı yapan sinir dikenleri, en arkadaki boyun omurları dışında uzun, alçak ve keskin sırtlar oluşturdu.[1][23]

Arkası Gallimimus 13 vardı sırt omurları kısa, ancak arkaya doğru daha derin ve daha uzun olma eğiliminde olan makara benzeri bir merkez ile. Enine süreçleri (kaburgalarla eklemlenen süreçler) arkaya doğru biraz arttı. İlk iki dorsal merkezde derin pnömatik foraminalar varken, geri kalanı sadece sığdı. fossae (çöküntüler) ve sinir dikenleri, bir şekilde üçgen veya dikdörtgen şeklinde belirgindi. sakrum (pelvik kemikler arasındaki kaynaşmış omurlar) beşten oluşuyordu sakral omur yaklaşık olarak eşit uzunluktaydı. Buradaki merkez, makaraya benzer, yanlara doğru düzleştirilmiş ve bazı örneklerde derin foramina olarak devam ettiği görülen fossalara sahipti. Buradaki sinir dikenleri dikdörtgen, geniş ve sırt omurundakilerden daha yüksekti. Yükseklikleri üst kenar boşluğuna eşit veya daha yüksekti. İliak bıçak ağzı ve ayrıydılar, diğer ornitomimidlerde ise birbirine kaynaşmışlardı. Kuyruğun 36-39 kaudal omurları vardı, öndekilerin merkezi makara şeklindeydi, arkadakiler ise neredeyse üçgen şeklindeydi ve boydan boya uzamıştı. Buradaki sinir dikenleri yüksek ve düzdü, ancak sırt benzeri hale geldikleri yerde geriye doğru azaldılar. Kuyruktaki tek pnömatiklik işareti, sinir dikenleri arasındaki derin çukurlar ve iki ilk kuyruk omurunun enine süreci idi. Sakrumun önündeki tüm omurlar deliği pirzola Atlas ve son dorsal vertebra hariç.[1][23]
kürek kemiği (kürek kemiği) kısa ve kavisli, ön uçta ince ve arkada kalındı. İle nispeten zayıf bir şekilde bağlantılıydı korakoid, yukarıdan aşağıya geniş ve derindi. Genel olarak, ön ayaklar, hepsi nispeten zayıf olan diğer ornitomimidlerden çok farklı değildi. humerus Enine kesite yakın dairesel bir kesite sahip olan (üst kol kemiği) uzun ve bükülmüştü. Humerusun üst ön kısmındaki deltopektoral tepe nispeten küçüktü ve bu nedenle üst kol kaslarının bağlanması için çok az yüzey sağladı. Ulna ince, uzun ve zayıf kıvrımlıydı ve neredeyse üçgen bir şaftı vardı. Olekranon (dirsekten çıkıntı) yetişkinlerde belirgindi, ancak gençlerde iyi gelişmedi. yarıçap (alt koldaki diğer kemik) uzun ve inceydi ve alt koldan daha genişletilmiş bir üst uç vardı. Manus (el), diğer ornitomimozorlarınkilerle orantılı olarak kısaydı, grubun herhangi bir üyesinin en küçük manus / humerus uzunluğu oranına sahipti, ancak başka türlü yapı olarak benzerdi. Benzer şekilde geliştirilmiş üç parmağı vardı; birincisi ("başparmak") en güçlüydü, üçüncüsü en zayıftı ve ikincisi en uzundu. toynaklılar (pençe kemikleri) güçlü, biraz kavisli (ilk parmağınki en kavisliydi) ve her iki tarafta derin bir olukla yana doğru sıkıştırılmıştı. Üçüncüsü biraz daha küçük olmasına rağmen, toynaklar da benzer şekilde geliştirildi.[1][8][17]
kasık (kasık kemiği) uzun ve inceydi, ornitomimozorların genel bir özelliği olan öne ve arkaya doğru genişleyen bir kasık botu ile bitiyordu. Arka uzuvlar, diğer ornitomimidlerden çok az farklıydı ve orantılı olarak diğer theropodlardan daha uzundu. Femur, yana doğru düzleştirilmiş bir şaft ile neredeyse düz, uzun ve inceydi. tibia düz, uzun ve iyi gelişmiş iki Kondiller (bir kemiğin yuvarlak ucu) üst uçta ve düzleştirilmiş bir alt uçta. fibula Alt bacak düz, ince ve genişti, üst uçta alt uca doğru daraldı. Üçüncünün alt yarısı metatarsal Uçtan bakıldığında genişti, kısmen her iki tarafa bitişik iki metatarsalı kapladı, ancak orta uzunlukta aniden daraldı, bu kemikler arasında sıkışarak ve kayboldu (bir Arctometatarsalian ayak yapısı). Üçüncü ayak parmağı, diğer ornitomimidlere göre uzuvla orantılı olarak daha kısaydı. Diğer ornitomimidlerde olduğu gibi, ayak halluks (veya diğer birçok theropodun ilk parmağı olan dewclaw). Ayak parmaklarının toynakları alt taraflarında düzdü; dıştaki ikisi rakamlarından biraz dışa doğru düştü.[1][17]
Sınıflandırma


Osmólska ve meslektaşları atandı Gallimimus Ornithomimidae ailesine 1972'de Kuzey Amerikalı ile birlikte Struthiomimus en yakın akraba olarak, arasındaki karşılaştırmanın takson zordu çünkü o sırada bilinen diğer ornitomimidler ya yetersiz korunmuş ya da yetersiz tanımlanmıştı.[1] 1975'te Kielan-Jaworowska, Asya'dan birçok dinozorun Kuzey Amerika akrabalarıyla aynı ailelere yerleştirilmesine rağmen, bu sınıflandırma kategorisinin modern kuşlarda kullanılandan daha kapsamlı olma eğiliminde olduğunu belirtti. O sırada vurguladı Gallimimus yuvarlatılmış bir gagası vardı (bir kaz veya ördeğe benzer), Kuzey Amerika ornitomimidleri sivri gagalara sahipti; bu, aksi takdirde taksonomistlerin modern kuşları farklı ailelere yerleştirmelerine yol açacak bir farktı.[5] 1976'da Barsbold, Ornithomimidae'yi yeni Ornithomimosauria grubuna ("devekuşu dinozorları") yerleştirdi. 2003 yılında Kobayashi ve Jun-Chang Lü şunu buldu: Anserimimus oldu kardeş takson -e Gallimimus, her ikisi de bir türetilmiş (veya "gelişmiş") clade 2006 yılında Kobayashi ve Barsbold tarafından onaylanan Kuzey Amerika cinsi ile.[8][19]
Aşağıdaki kladogram yerleşimini gösterir Gallimimus Li Xu ve meslektaşlarına göre Ornithomimidae arasında, 2011:[24]
| Ornithomimidae |
| ||||||||||||||||||||||||||||||||||||
Ornitomimozorlar klada aitti Maniraptoriformes nın-nin Coelurosaurian modern kuşları da içeren theropodlar. İlk ornitomimozorlar, grubun daha türetilmiş üyelerinde kaybolan dişlere sahipti.[25] 2004'te Makovicky, Kobayashi ve Currie, ornitomimozorların erken evrimsel tarihinin çoğunun, çoğu cinsin keşfedildiği Asya'da gerçekleştiğini öne sürdüler. baz alınan (veya "ilkel") taksonlar, bazalin varlığını kabul etseler de Pelecanimimus Avrupa'da sınıflandırmada bir komplikasyon ortaya çıkar. Grup, Asya'dan Kuzey Amerika'ya bir veya iki kez dağılmış olmalı. Beringia orada bulunan Geç Kretase cinslerini açıklamak için. Diğer bazı dinozor gruplarında görüldüğü gibi, ornitomimozorlar, Avrupa Asya'dan ayrıldıktan sonra büyük ölçüde Asya ve Kuzey Amerika ile sınırlıydı. Turgai Boğazı.[17]
1994 yılında paleontolog Thomas R. Holtz gruplanmış ornitomimozorlar ve Troodontidler parasfenoid üzerinde soğanlı bir kapsülün varlığı gibi paylaşılan özelliklere dayanan bir sınıfta. O clade Bullatosauria adını özel ismine dayanarak Gallimimus bullatus, bu da kapsüle atıfta bulunuyordu.[26] 1998'de Holtz, bunun yerine troodontidlerin bazal maniraptoranlar olduğunu buldu; bu, bu sınıftaki tüm üyelerin Bullatosauria'ya gireceği ve bu nedenle Maniraptoriformes'in küçük bir eşanlamlısı haline geleceği ve sınıfın o zamandan beri kullanım dışı kaldığı anlamına geldiğini buldu.[27][28]
Paleobiyoloji

Servikal vertebra Gallimimus boynunu eğik tuttuğunu ve 35 derecelik bir açıyla yukarı doğru indiğini gösterir. Osmólska ve meslektaşları, Gallimimus değildi kavrayıcı (ya da kavrayabilen) ve başparmağın karşı çıkmadığını. Ayrıca, kolların, örneğin ornitomimozorunkilere kıyasla zayıf olduğunu öne sürdüler. Deinocheirus. Ornitoimid biyolojinin paleontolog tarafından yapılan yorumlarına katıldılar. Dale Russell 1972'nin başlarından itibaren, çok filo (veya cursorial ) hayvanlar, büyük, modern kara kuşlarından daha az çevik olsalar da hızlarını avcılardan kaçmak için kullanırlardı. Russell ayrıca, modern teknolojiye kıyasla iyi bir vizyon ve zekaya sahip olduklarını öne sürdü. ratit kuşlar. Yırtıcıları sahip olabileceğinden renkli görüş, renklerini etkileyeceğini, belki de kamuflaj.[1][29] 1982'de paleontolog Richard A. Thulborn tahmin etti Gallimimus 42–56 km / sa (29–34 mil / sa) hızlarda koşmuş olabilir. Ornitimidlerin, kısmen kolları ve kuyruklarının ağırlıklarını artırması nedeniyle 70-80 km / saate (43-49 mph) ulaşabilen devekuşları kadar hızlı olmayacağını buldu.[30]
1988'de Paul, ornitomimidlerin gözlerinin düzleştirildiğini ve soketler içinde minimum hareketliliğe sahip olduğunu, bu da nesneleri görmek için kafanın hareket etmesini gerektirdiğini öne sürdü. Gözleri diğer kuş benzeri theropodlara göre daha yana baktığından dürbün görüşü daha sınırlı olurdu, bu da bazı hayvanlarda arkalarındaki yırtıcıları görme yeteneklerini geliştiren bir adaptasyon. Paul, ağırlığı azaltan ve ornitomimidlerin hallukslerini kaçıran görece kısa kuyrukların hız için uyarlamalar olduğunu düşünüyordu. Gagalama ve tekme atarak kendilerini savunabileceklerini, ancak esas olarak kaçış hızlarına güvenebileceklerini ileri sürdü.[14] 2015 yılında, Akinobu Watanabe ve meslektaşları bunu, Deinocheirus ve Archaeornithomimus, Gallimimus ornitomimozorlar arasında en fazla pnömatik hale gelen iskelete sahipti. Pnömatizasyonun modern kuşlarda uçuş için avantajlı olduğu düşünülmektedir, ancak bunun kuş olmayan dinozorlardaki işlevi kesin olarak bilinmemektedir. Büyük kemiklerin kütlesini azaltmak için pnömatizasyonun kullanıldığı, yüksek metabolizma, hareket sırasında denge veya termoregülasyon.[23]

2017'de Lee ve meslektaşları çeşitli olasılıklar önerdiler tafonomik nasıl olduğunu açıklamak için koşullar (çürüme ve fosilleşme sırasındaki değişiklikler) Gallimimus 2009'da keşfedilen ayak bir yolla ilişkilendirildi. Parkur, şurada korunur: kumtaşı ayak saklanırken çamurtaşı, izlerin bulunduğu katmanın 20 santimetre (7,9 inç) altına uzanır. Fosil, izlerinde ölen bir hayvanı temsil ediyor olabilir, ancak çamurdaki ayağın derinliği, çamurun batması için çok sığ olabilir. Ayrıca bir sel nedeniyle ölmüş ve ardından bir gölete gömülmüş olabilir. Bununla birlikte, çamur ve kum katmanları su baskını değil, muhtemelen kuru bir ortam ve bozulmuş sedimanlar Fosilin etrafı, hayvanın bölgeye geldiğinde yaşadığını gösterir. Yazarlar bu nedenle izlerin uzun bir süre ve kuruma süresi boyunca yapıldığını ve muhtemelen hiçbirinin ayağa sahip olan kişi tarafından üretilmediğini öne sürdüler. Hayvan, yağmur veya suyla ıslanırken bir göletin tabanından geçerek tortu katmanını izlerle kırmış olabilir. Hayvan bu pozisyonda susuzluktan, açlıktan veya başka bir nedenle ölmüş olabilir ve daha sonra çamurun üzerine çökelmiş, böylece izleri ve karkası kaplayıp koruyabilir. Ayak çürürken kenetlenmiş ve serbest kalmış olabilir, bu da tendonlar esnedi ve daha sonra ağır dinozorlar tarafından basıldı. Alan tek olabilir kemik yatağı (olası haşlanmış örnek sayısına dayalı olarak) bir Gallimimus belki de kuraklık veya kıtlık nedeniyle kitlesel ölümler. Hayvanların aynı anda ölmüş gibi görünmesi (boş kazı çukurları stratigrafik olarak özdeşti) şunu gösterebilir: Gallimimus oldu sokulgan (gruplar halinde yaşadı), diğer ornitomimidler için de önerildi.[12]
Beslenme ve diyet

Osmólska ve meslektaşları, boynun ön kısmının Gallimimus çok hareketli olurdu (arka kısım daha sertti), o bölgenin omurlarındaki sinir kemerleri tavuk ve diğer Galliformlara benziyordu, bu da benzer beslenme alışkanlıklarını gösteriyordu. Gagasını buldular Gallimimus bir ördek ya da kazınkine benzer ve bütün olarak yediği küçük, canlı avla beslenmiş olması. Gözler kafatasının yan taraflarına yerleştirildiğinden, boynun hareketliliği avın yerde bulunmasında faydalı olabilirdi. Tüm ornitomimidlerin benzer beslenme alışkanlıklarına sahip olduğunu varsaydılar ve Russel'in ornitomimidlerin gagalarını ornitomimidlerin gagalarıyla karşılaştırdığını belirttiler. böcek yiyen kuşlar. Osmólska ve meslektaşları şunu önerdi: Gallimimus yapabiliyordu kafatası kinesis (kafatasının arkasındaki bazı kemikler arasındaki görünüşte gevşek bağlantı nedeniyle), kafatasının tek tek kemiklerinin birbirine göre hareket etmesine izin veren bir özellik. Ayrıca, kısa elli ön ayaklarını yiyecekleri ağza götürmek için değil, yiyeceğe ulaşmak için tırmıklamak veya toprağı kazmak için kullanmasını önerdiler.[1] Elleri Gallimimus örneğin aşağıdakilerden daha zayıf olabilir Struthiomimuspaleontologlar Elizabeth L. Nicholls ve Anthony P. Russell tarafından yazılan 1985 tarihli bir makaleye göre, bunun yerine ellerini kancalamak ve kavramak için kullanmış olabilir.[31]
1988'de Paul, ornitomimidlerin omnivorlar daha önce önerildiği gibi küçük hayvanları ve yumurtaları ve bitkileri yiyenler. Devekuşlarının ve emüslerin ağırlıklı olarak otlayanlar ve tarayıcılar ve ornitomimidlerin kafataslarının soyu tükenmiş kafataslarına en çok benzediğini moas Bağırsak içeriğinden de anlaşılacağı gibi, dalları ısıracak kadar güçlü olan. Ayrıca, ornitomimidlerin sert bitkilere göz atmak için iyi adapte olduklarını ve dalları çenelerine yaklaştırmak için ellerini kullanacaklarını öne sürdü.[14] Paleontolog Jørn Hurum 2001 yılında benzer çene yapısı nedeniyle, Gallimimus martılar gibi fırsatçı, omnivor bir diyet yapmış olabilir. O da sıkı olduğunu gözlemledi intramandibular eklem alt çenenin ön ve arka kısımları arasındaki herhangi bir hareketi engelleyecektir.[21]

2001 yılında paleontologlar Mark A. Norell, Makovicky ve Currie bir Gallimimus kafatası (IGM 100/1133) ve bir Ornitomimus gagadaki yumuşak doku yapılarını koruyan kafatası. İç tarafı Gallimimus gaga, yazarların benzer buldukları sütunlu yapılara sahipti. lameller gagalarında Anseriform Bunları yiyecekleri manipüle etmek, tortuları süzmek için kullanan kuşlar, filtre besleme Gıda maddelerini diğer malzemelerden ayırarak ve otlarken bitkileri kesmek için. Buldular Kuzey kürekçi bitkiler, yumuşakçalar ile beslenen, ostrakodlar, ve foraminiferans anatomi bakımından en çok benzer yapılara sahip modern anseriform olmak Gallimimus. Yazarlar, ornitomimidlerin muhtemelen gagalarını büyük hayvanları avlamak için kullanmadıklarını ve mesic ortamlar, daha kurak ortamlarda daha nadir olmakla birlikte, sucul gıda kaynaklarına bağlı olabileceklerini düşündürmektedir.[22] Bu yorum doğruysa, Gallimimus bilinen en büyük karasal filtre besleyicilerden biri olurdu.[17]
2005 yılında paleontolog Paul Barrett, lamel benzeri yapıların Gallimimus filtre besleyen kuşlarınki gibi esnek kıllar gibi görünmüyordu (çünkü bu yapıların üst üste geldiğine veya çöktüğüne dair hiçbir gösterge yok), bunun yerine kaplumbağaların gagalarındaki ince, düzenli aralıklı dikey çıkıntılara daha çok benziyordu ve hadrosaurid dinozorlar. Bu hayvanlarda, bu tür çıkıntıların, sert bitki örtüsünü hasat etmek için kullanılan otçul diyetlerle ilişkili olduğu düşünülmektedir. Barrett, gagasındaki çıkıntıların Gallimimus gaganın iç yüzeyinin doğal bir dökümünü temsil ediyordu, bu da hayvanın lif içeriği yüksek olan bir otobur olduğunu gösteriyordu. Birçok kişinin keşfi mide taşı (taşlık Bazı ornitomimidlerdeki taşlar) bir mide değirmeni varlığını gösterir ve bu nedenle otçul bir diyete işaret eder, çünkü bunlar gerekli çiğneme aparatına sahip olmayan hayvanların yiyeceklerini öğütmek için kullanılır. Barrett ayrıca 440 kilogram (970 lb) Gallimimus olup olmadığına bağlı olarak, günde 0.07 ila 3.34 kilogram (0.15 ila 7.36 lb) yiyecek gerekirdi. endotermik veya bir ektotermik ("sıcak" veya "soğuk" -kanlı) metabolizma, bir filtre besleyici olsaydı gerçekleştirilemez bulduğu bir alım. Ayrıca ornitomimidlerin sadece mezik ortamları temsil eden oluşumlarda değil, aynı zamanda filtreli beslemeye dayalı bir diyet sürdürmek için yetersiz suyun olduğu kurak ortamlarda da bol olduğunu buldu.[20] 2007 yılında, paleontolog Espen M. Knutsen, ornitomimidlerin gaga şeklinin, modern kuşlara kıyasla, omnivori veya yüksek lifli otçul hayvanlarla tutarlı olduğunu yazdı.[32]
Geliştirme

Kafatasının şekli ve oranları büyüme sırasında önemli ölçüde değişti. Kafatasının arkası ve yörüngelerin boyutu küçüldü, oysa burun nispeten uzadı; modernde benzer değişiklikler meydana gelir timsahlar. Kafatası aynı zamanda genç örneklerde orantılı olarak daha büyüktü ve burnun üst profilinin eğimi daha az belirgindi. Boyundaki kaburgalar sadece yetişkinlerde omurlarla kaynaşmıştır. Ön ayaklar, büyüme sırasında orantılı olarak daha uzun görünürken, arka bacaklardaki kemiklerin orantılı uzunluğu çok az değişmiştir.[1][21] 2012 yılında, paleontolog Darla K. Zelenitsky ve meslektaşları, yetişkin ornitomimozorların kollarında kanat benzeri yapılara sahipken gençlerin kanat benzeri yapılara sahip olmadıkları sonucuna varmışlardır. Ornitomimus), bu yapılar orijinal olarak ikincil cinsel özellikler gibi üreme davranışları için kullanılmış olabilir kur, Görüntüle ve düşünceli.[18]
Biyologlar Roman Pawlicki ve P.Bolechała tarafından 1987 yılında yapılan bir araştırma, içeriğinde yaşa bağlı farklılıklar gösterdi. kalsiyum ve fosfor (kemik oluşumunda önemli bileşenler) Gallimimus örnekler. Oranın genç-orta yaşlı hayvanlarda en yüksek olduğunu, yaşla birlikte azaldığını buldular.[33] 1991 yılında, yaşlı bireylerin kemiklerinin en yüksek miktarda kurşun ve demir içerdiğini, genç hayvanlarda ise daha düşük olduğunu bildirdiler.[34] Kemik çalışması histoloji 2000 yılında biyologlar John M. Rensberger ve Mahito Watabe tarafından çeşitli dinozorların kanaliküller (kemik hücrelerini birbirine bağlayan kanallar) ve kolajen lif demetleri Gallimimus ve diğer ornitomimidler kuşlardakilere memelilerden farklı olarak ornitorik memelilere daha çok benzeyen dinozorlar. Bu farklılıklar, kemiğin oluştuğu süreç ve hız ile ilişkili olabilir.[35]
Paleoçevre

Gallimimus dan bilinmektedir Nemegt Oluşumu Güney Moğolistan'ın Gobi Çölü'nde. Bu jeolojik oluşum hiç olmadı radyometrik olarak tarihli, ama fauna fosil kayıtlarında bulunanlar, muhtemelen erken dönemde çökelmiş olduğunu göstermektedir. Maastrihtiyen aşamanın sonunda Geç Kretase yaklaşık 70 milyon yıl önce.[36][37][38] Çökeltileri Gallimimus yerellik yazın Tsaagan Khushuu şunlardan oluşur: alüvyon, silttaşı, çamurtaşları, kumlar ve daha seyrek ince kumtaşı yatakları.[3] Kaya fasiyes Nemegt Formasyonu, nehir kanallarının, çamur tabakalarının, sığ göllerin ve taşkın yatakları benzer bir ortamda Okavango Deltası günümüzün Botsvana.[39] Büyük nehir kanalları ve toprak birikintileri, daha yaşlı olanlardan çok daha nemli bir iklime sahip olduğunun kanıtıdır. Barun Goyot ve Djadochta oluşumlara rağmen kaliş mevduatlar periyodik kuraklıkların meydana geldiğini göstermektedir.[40] Nemegt Havzası'ndaki fosil kemikler Gallimimus, daha fazla radyoaktif çevredeki fosillerden daha büyük olasılıkla uranyum kemiklerde birikmiş, oraya yer altı suyu süzülerek taşınmıştır.[6][41]
Nemegt nehirleri, nerede Gallimimus yaşadı, çok çeşitli organizmalara ev sahipliği yaptı. Ara sıra yumuşakça fosiller ve balık, kaplumbağalar gibi diğer çeşitli su hayvanları,[36] ve krokodilomorflar, dahil olmak üzere Shamosuchus, bu bölgede keşfedilmiştir.[42] Memeli Nemegt Formasyonunda fosiller nadirdir, ancak birçok kuş enantiornitin Gurilynia, hesperornithiform Judinornis, Hem de Teviornis, olası bir anseriform bulunmuştur. Nemegt Formasyonunda keşfedilen otçul dinozorlar şunları içerir: ankilozoridler gibi Tarchia, pachycephalosaurian Prenocephale, large hadrosaurids such as Saurolophus ve Barsboldia, and sauropods such as Nemegtosaurus ve Opisthocoelicaudia.[36][43] Predatory theropods that lived alongside Gallimimus Dahil etmek Tyrannosauroidler gibi Tarbosaurus, Alioramus ve Bagaraatan, and troodontids such as Borogovia, Tochisaurus ve Zanabazar. Herbivorous or omnivorous theropods include therizinosaurs, such as Therizinosaurus, as well as oviraptorosaurians, such as Elmisaurus, Nemegtomaia, ve Rinchenia.[44] Other ornithomimosaurs, including Anserimimus ve Deinocheirus, are also found, but Gallimimus is the most common member of the group in the Nemegt.[8]
Kültürel önem

Gallimimus was featured in the 1993 movie Jurassic Park yönetmen tarafından Steven Spielberg; a similar scene in the original 1990 novel instead featured hadrosaurs. Spielberg had wanted a izdiham sequence with animal herds in the movie, but did not know how to achieve it, and it was initially going to be visualised through hareketli animasyonu durdur. At the time, there was little faith in creating animals through bilgisayar animasyonu, ama görsel efektler şirket Endüstriyel Işık ve Büyü was given a go-ahead by the movie's producers to explore possibilities. ILM created a Gallimimus skeleton in the computer and animated a test showing a herd of running skeletons, and later a Tyrannosaurus chasing a fully rendered Gallimimus sürü. The production team became very enthusiastic as nothing similar had previously been achieved, and Spielberg was convinced to write the scene into the script, and to also use computer graphics for other dinosaur shots in the movie instead of stop motion.[45][46] Gallimimus were animated by tracing frames from footage of ostriches, and footage of herding ceylanlar ayrıca referans alınmıştır.[47] Kielan-Jaworowska, who discovered the holotype specimen, called it a "beautiful scene".[6] The movie's dinosaurs were one of the most widely publicised applications of bilgisayar tarafından oluşturulan görüntüler in film, and were considered more lifelike than what had been previously accomplished with özel efektler.[48]
Emphasising the bird-like flocking behaviour of the Gallimimus herd was a point in Jurassic Park's story, as they were supposed to represent the precursors to birds. The herd was shown moving as a whole, rather than individual animals running around, and the smaller Gallimimus were shown in the middle of the group, as though they were being protected.[45] During the scene, the palaeontologist Alan Grant says that the herd moves with "uniform direction changes, just like a flock of birds evading a predator" and "bet you'll never look at birds the same way again" as he watches the movements of the fast, graceful Gallimimus. This contrasted with how dinosaurs were traditionally depicted in mass media as lumbering, tail-dragging animals, and the movie helped change the common perception of dinosaurs. This and other scenes reflected then-recent theories of kuş evrimi encouraged by the movie's scientific advisor, the palaeontologist John R. Horner, ideas which were still contentious at the time.[49][50][51] Despite such theories, Gallimimus and other dinosaurs of the movie were depicted without feathers, in part because it was unknown at the time how widespread these were among the group.[49][52]
İddia edildi ki Lark Quarry tracks (one of the world's largest concentrations of dinosaur tracks) in Queensland, Australia, served as inspiration and "scientific underpinning" for the Gallimimus stampede scene in Jurassic Park; these tracks were initially interpreted as representing a dinosaur stampede caused by the arrival of a theropod predator. The idea that the tracks represent a stampede has since been contested (the "theropod" may instead have been a herbivore similar to Muttaburrasaurus ), and a consultant to Jurassic Park has denied the tracks served as inspiration for the movie.[53][54][55]
Ayrıca bakınız
Referanslar
- ^ a b c d e f g h ben j k l m n Osmolska, H.; Roniewicz, E.; Barsbold, R. (1972). "A new dinosaur, Gallimimus bullatus n. gen., n. sp. (Ornithomimidae) from the Upper Cretaceous of Mongolia" (PDF). Paleontoloji Polonica. 27: 103–143.
- ^ Kielan-Jaworowska, Z.; Dovchin, N. (1968). "Narrative of the Polish-Mongolian palaeontological expeditions 1963–1965" (PDF). Palaeontologica Polonica: 7–30.
- ^ a b Gradzihski, R.; Kazmierczak, J .; Lefeld, J. (1969). "Geographical and geological data from the Polish-Mongolian palaeontological expeditions" (PDF). Palaeontologica Polonica. 19: 33–82.
- ^ Dodson, P. (1998). Boynuzlu Dinozorlar: Doğal Bir Tarih. Princeton: Princeton Üniversitesi Yayınları. s. 9. ISBN 978-0-691-05900-6.
- ^ a b c Kielan-Jaworowska, Z. (1975). "Late Cretaceous mammals and dinosaurs from the Gobi Desert: Fossils excavated by the Polish-Mongolian paleontological expeditions of 1963–71 cast new light on primitive mammals and dinosaurs and on faunal interchange between Asia and North America". Amerikalı bilim adamı. 63 (2): 150–159. Bibcode:1975AmSci..63..150K. JSTOR 27845359.
- ^ a b c d e Kielan-Jaworowska, Z. (2013). In Pursuit of Early Mammals. Geçmişin Hayatı. Bloomington: Indiana University Press. pp. 47–48, 50–51. ISBN 978-0-253-00824-4.
- ^ a b Glut, D. F. (1997). Dinozorlar: Ansiklopedi. Jefferson: McFarland & Co. pp.429 –432. ISBN 978-0-89950-917-4.
- ^ a b c d e f Kobayashi, Y .; Barsbold, R. (2006). "Ornithomimids from the Nemegt Formation of Mongolia" (PDF). Journal of the Paleontological Society of Korea. 22 (1): 195–207.
- ^ "Cretaceous Mongolian Dinosaurs". DinoCasts.com. Arşivlenen orijinal 2 Şubat 2015.
- ^ Oest, S. G.; Hansen, S.; Oest, T. (1997). "Dinosaurer, æg og unger – en særudstilling om en forsvunden verden" (PDF). GeologiskNyt (Danca). 6: 24–25. Arşivlenen orijinal (PDF) 2018-05-14 tarihinde. Alındı 2018-05-13.
- ^ Fanti, F .; Bell, P. R.; Tighe, M.; Milan, L. A.; Dinelli, E. (2018). "Geochemical fingerprinting as a tool for repatriating poached dinosaur fossils in Mongolia: A case study for the Nemegt Locality, Gobi Desert". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 494: 51–64. Bibcode:2018PPP...494...51F. doi:10.1016/j.palaeo.2017.10.032.
- ^ a b Lee, H.-J .; Lee, Y.-N .; Adams, T. L.; Currie, P. J .; Kobayashi, Y .; Jacobs, L. L .; Koppelhus, E. B. (2018). "Theropod trackways associated with a Gallimimus foot skeleton from the Nemegt Formation, Mongolia". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 494: 160–167. Bibcode:2018PPP...494..160L. doi:10.1016/j.palaeo.2017.10.020.
- ^ Gannon, M. (2014). "Stolen 'Nest of Dinosaurs' Returned to Mongolia". uk.news.yahoo.com. LiveScience.com. Alındı 21 Mayıs 2019.
- ^ a b c d Paul, G. S. (1988). Dünyanın Yırtıcı Dinozorları. New York: Simon ve Schuster. pp.384–386, 393–394. ISBN 978-0-671-61946-6.
- ^ Paul, G. S. (2016). Princeton Dinozorlar Saha Rehberi (2 ed.). Princeton: Princeton Üniversitesi Yayınları. s. 131. ISBN 978-0-691-16766-4.
- ^ Paul, G. S. (2010). Princeton Dinozorlar Saha Rehberi (1 ed.). Princeton: Princeton Üniversitesi Yayınları. pp.112–114. ISBN 978-0-691-13720-9.
- ^ a b c d e f Makovicky, P. J .; Kobayashi, Y .; Currie, P. J. (2004). "Ornithomimosauria". Weishampel, D. B .; Dodson, P .; Osmolska, H. (editörler). Dinosauria (2 ed.). Berkeley: California Üniversitesi Yayınları. pp.137 –150. ISBN 978-0-520-24209-8.
- ^ a b Zelenitsky, D. K.; Therrien, F .; Erickson, G. M .; DeBuhr, C. L.; Kobayashi, Y .; Eberth, D. A.; Hadfield, F. (2012). "Feathered non-avian dinosaurs from North America provide insight into wing origins". Bilim. 338 (6106): 510–514. Bibcode:2012Sci...338..510Z. doi:10.1126/science.1225376. PMID 23112330. S2CID 2057698.
- ^ a b c Kobayashi, Y .; Lü, J.-C. (2003). "Çin'in Geç Kretase'sinden kalma alışkanlıklara sahip yeni bir ornitoimid dinozor". Acta Palaeontologica Polonica. 48 (2): 235–259.
- ^ a b Barrett, P.M. (2005). "The diet of ostrich dinosaurs (Theropoda: Ornihomimosauria)". Paleontoloji. 48 (2): 347–358. doi:10.1111 / j.1475-4983.2005.00448.x.
- ^ a b c d Hurum, J. (2001). "Lower jaw of Gallimimus bullatus". Tanke, D. H .; Carpenter, K .; Skrepnick, M.W. (editörler). Mesozoik Omurgalı Yaşamı. Bloomington: Indiana University Press. pp.34–41. ISBN 978-0253339072.
- ^ a b Norell, M. A .; Makovicky, P. J .; Currie, P.J. (2001). "Devekuşu dinozorlarının gagaları". Doğa. 412 (6850): 873–874. Bibcode:2001Natur.412..873N. doi:10.1038/35091139. PMID 11528466. S2CID 4313779.
- ^ a b c Watanabe, A.; Eugenia Leone Gold, M.; Brusatte, S. L .; Benson, R. B. J .; Choiniere, J.; Davidson, A .; Norell, M. A .; Claessens, L. (2015). "Vertebral pneumaticity in the ornithomimosaur Archaeornithomimus (Dinosauria: Theropoda) revealed by computed tomography imaging and reappraisal of axial pneumaticity in ornithomimosauria". PLOS ONE. 10 (12): e0145168. Bibcode:2015PLoSO..1045168W. doi:10.1371/journal.pone.0145168. PMC 4684312. PMID 26682888.
- ^ Xu, L .; Kobayashi, Y .; Lü, J .; Lee, Y. N.; Liu, Y .; Tanaka, K.; Zhang, X .; Jia, S.; Zhang, J. (2011). "Çin'in Henan Eyaletindeki Geç Kretase Qiupa Formasyonundan Kuzey Amerika yakınlıklarına sahip yeni bir ornitomimid dinozor." Kretase Araştırmaları. 32 (2): 213–222. doi:10.1016 / j.cretres.2010.12.004.
- ^ Hendrickx, C .; Hartman, S. A.; Mateus, O. (2015). "An overview on non-avian theropod discoveries and classification". PalArch'ın Omurgalı Paleontoloji Dergisi. 12 (1): 1–73.
- ^ Holtz, T. R. (1994). "The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics". Paleontoloji Dergisi. 68 (5): 1100–1117. doi:10.1017/S0022336000026706. JSTOR 1306180.
- ^ Holtz, T. R. (1998). "A new phylogeny of the carnivorous dinosaurs". GAIA. 15: 5–61. CiteSeerX 10.1.1.211.3044. ISSN 0871-5424.
- ^ Naish, D .; Martill, D. M .; Frey, E. (2004). "Ecology, Systematics and biogeographical relationships of dinosaurs, including a new theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil". Tarihsel Biyoloji. 16 (2–4): 57–70. CiteSeerX 10.1.1.394.9219. doi:10.1080/08912960410001674200. S2CID 18592288.
- ^ Russell, D. A. (1972). "Ostrich dinosaurs from the Late Cretaceous of Western Canada". Kanada Yer Bilimleri Dergisi. 9 (4): 375–402. Bibcode:1972CaJES...9..375R. doi:10.1139/e72-031.
- ^ Thulborn, R. A. (1982). "Speeds and gaits of dinosaurs". Paleocoğrafya, Paleoklimatoloji, Paleoekoloji. 38 (3–4): 227–256. Bibcode:1982PPP....38..227T. doi:10.1016/0031-0182(82)90005-0.
- ^ Nicholls, E. L .; Russell, A.P. (1985). "Göğüs kemeri ve ön ayağının yapısı ve işlevi Struthiomimus altus (Theropoda: Ornithomimidae)". Paleontoloji. 28 (4): 64–677.
- ^ Madsen, E. K. (2007). "Beak morphology in extant birds with implications on beak morphology in ornithomimids". Det Matematisk-Naturvitenskapelige Fakultet – Thesis: 1–21.
- ^ Pawlicki, R.; Bolechała, P. (1987). "X-ray microanalysis of fossil dinosaur bone: age differences in the calcium and phosphorus content of Gallimimus bullatus bones". Folia Histochemica et Cytobiologica. 25 (3–4): 241–244. PMID 3450541.
- ^ Pawlicki, R.; Bolechała, P. (1991). "X-ray microanalysis of fossil dinosaur bone: age differences in lead, iron, and magnesium content". Folia Histochemica et Cytobiologica. 29 (2): 81–83. PMID 1804726.
- ^ Rensberger, J. M.; Watabe, M. (2000). "Fine structure of bone in dinosaurs, birds and mammals". Doğa. 406 (6796): 619–622. Bibcode:2000Natur.406..619R. doi:10.1038/35020550. PMID 10949300. S2CID 4384549.
- ^ a b c Jerzykiewicz, T .; Russell, D. A. (1991). "Late Mesozoic stratigraphy and vertebrates of the Gobi Basin". Kretase Araştırmaları. 12 (4): 345–377. doi:10.1016/0195-6671(91)90015-5.
- ^ Sullivan, R. M. (2006). "A taxonomic review of the Pachycephalosauridae (Dinosauria: Ornithischia)" (PDF). Lucas, Spencer G .; Sullivan, Robert M. (editörler). Late Cretaceous vertebrates from the Western Interior. 35. New Mexico Doğa Tarihi ve Bilim Müzesi Bülteni. pp. 347–366.
- ^ Gradstein, F. M .; Ogg, J. G.; Smith, A. G. (2005). Jeolojik Zaman Ölçeği 2004. Cambridge: Cambridge University Press. pp.344 –371. doi:10.1017/CBO9780511536045. ISBN 978-0-521-78142-8.
- ^ Holtz, T. R. (2014). "Paleontology: Mystery of the horrible hands solved". Doğa. 515 (7526): 203–205. Bibcode:2014Natur.515..203H. doi:10.1038/nature13930. PMID 25337885. S2CID 205241353.
- ^ Novacek, M. (1996). Dinosaurs of the Flaming Cliffs. New York: Anchor. s.133. ISBN 978-0-385-47775-8.
- ^ Jaworowski, Z.; Pensko, J. (1967). "Unusually radioactive fossil bones from Mongolia". Doğa. 214 (5084): 161–163. Bibcode:1967Natur.214..161J. doi:10.1038/214161a0. S2CID 4148855.
- ^ Efimov, M. B. (1983). "Peresmotr iskopayemykh krokodilov Mongolii" [Revision of the fossil crocodiles of Mongolia]. Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya Trudy (Rusça). 24: 76–96.
- ^ Hurum, J. H .; Sabath, K. (2003). "Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar ve Tyrannosaurus rex compared" (PDF). Acta Palaeontologica Polonica. 48 (2): 188.
- ^ Holtz, T. R. (2004). "Tyrannosauroidea". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (editörler). Dinosauria (2 ed.). Berkeley: California Üniversitesi Yayınları. s.124. ISBN 978-0-520-24209-8.
- ^ a b Shay, D.; Duncan, J. (1993). The Making of Jurassic Park. New York: Boxtree Ltd. pp. 48–52, 135–136. ISBN 978-1-85283-774-7.
- ^ Crichton, M. (1990). Jurassic Park. London: Random Century Group. pp.137–138. ISBN 978-0-394-58816-2.
- ^ Baird, R. (1998). "Animalizing Jurassic Park's dinosaurs: Blockbuster schemata and cross-cultural cognition in the threat scene". Sinema Dergisi. 37 (4): 82–103. doi:10.2307/1225728. JSTOR 1225728.
- ^ Prince, S. (1996). "True lies: Perceptual realism, digital images, and film theory". Üç Aylık Film (Gönderilen makale). 49 (3): 27–37. doi:10.2307/1213468. hdl:10919/25811. JSTOR 1213468.
- ^ a b Montanari, S. (2015). "What we've learned about dinosaurs since Jurassic Park came out". Forbes. Alındı 7 Mayıs 2018.
- ^ Kirby, D. A. (2011). Lab Coats in Hollywood: Science, Scientists, and Cinema. Massachusetts: MIT Basın. pp.127 –129. ISBN 978-0262294867.
- ^ Kirby, D. A. (2003). "Science consultants, fictional films, and scientific practice". Bilim Sosyal Çalışmaları. 33 (2): 231–268. doi:10.1177/03063127030332015. JSTOR 3183078. S2CID 111227921.
- ^ Phillips, I. (2015). "Nasıl Jurassic World dinosaurs looked in real life". Business Insider. Alındı 3 Aralık 2016.
- ^ Martin, A. J. (2014). Dinosaurs Without Bones: Dinosaur Lives Revealed by their Trace Fossils. London: Pegasus Books. sayfa 66–67. ISBN 978-1605984995.
- ^ Romilio, A. (2015). "Dinosaur stampede stopped in its tracks". Avustralasya Bilimi. 36 (2): 24–27. ISSN 1442-679X.
- ^ Shapiro, A. D. (2013). "The great dinosaur stampede that never was?". BBC haberleri. Alındı 7 Mayıs 2018.
Dış bağlantılar
 İle ilgili medya Gallimimus Wikimedia Commons'ta
İle ilgili medya Gallimimus Wikimedia Commons'ta














| Yetki kontrolü |
|---|